Recent Advances in Chitosan-Based Carriers for Gene Delivery



Abstract
1. Introduction
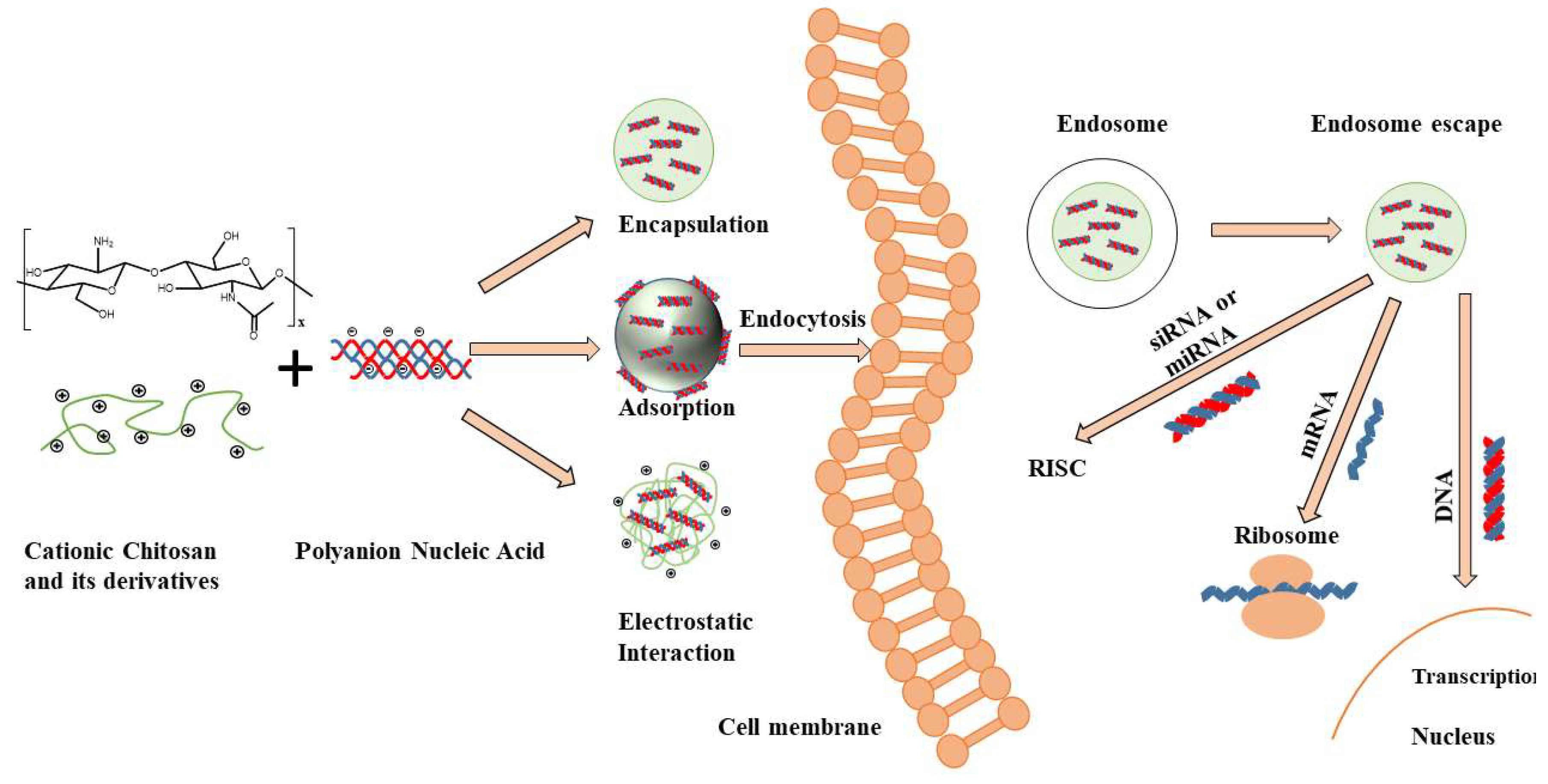
2. Design Criteria and Requirements for Gene Delivery Using Chitosan-Based Carriers
2.1. Design Considerations for Chitosan-Based Carriers
2.2. Basic Properties of Chitosan
2.3. Chitosan Molecular Weight
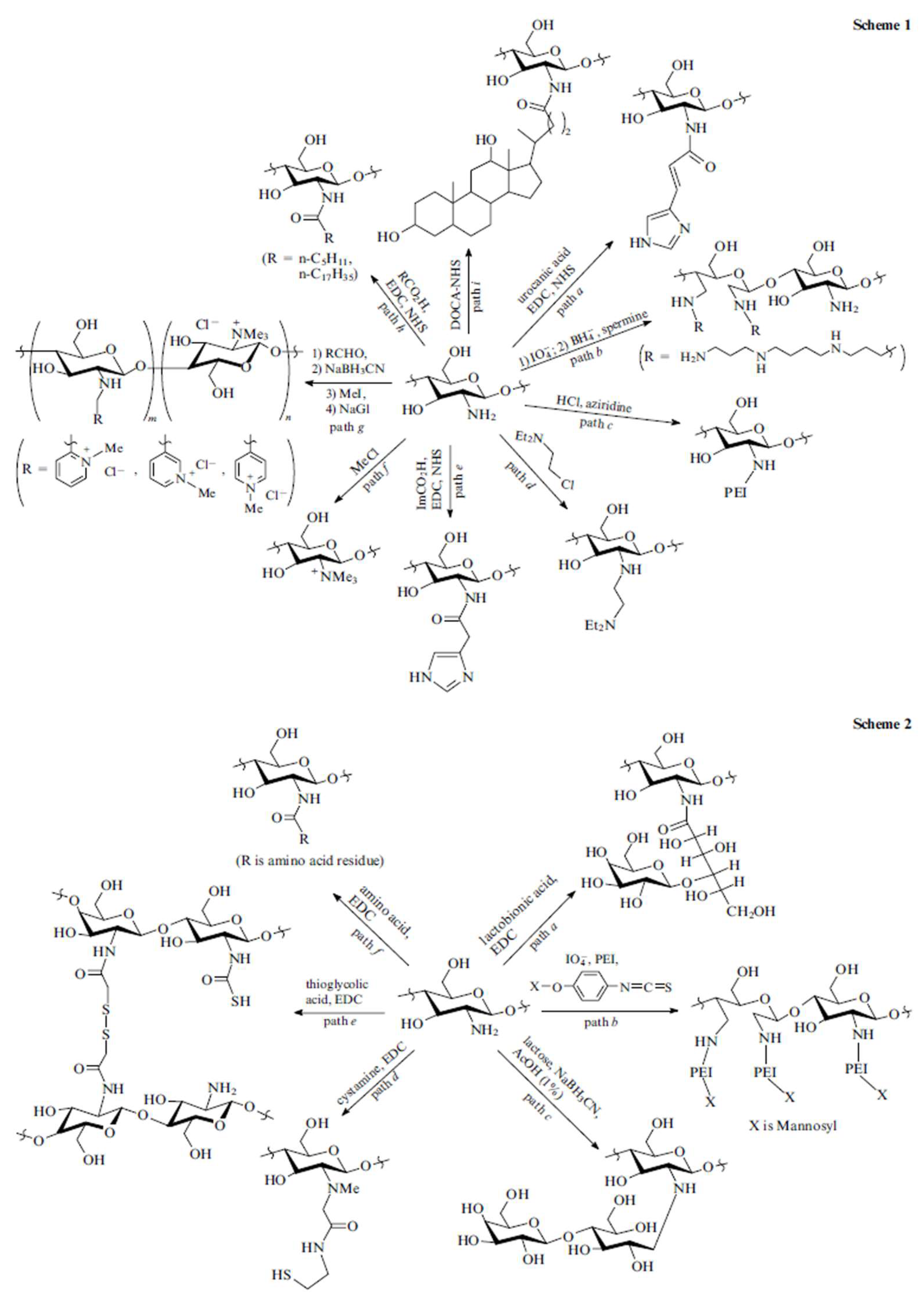
2.4. Chitosan Derivatives
2.5. Chitosan Deacetylation Degree
2.6. N/P Ratio
2.7. Chitosan’s Toxicity
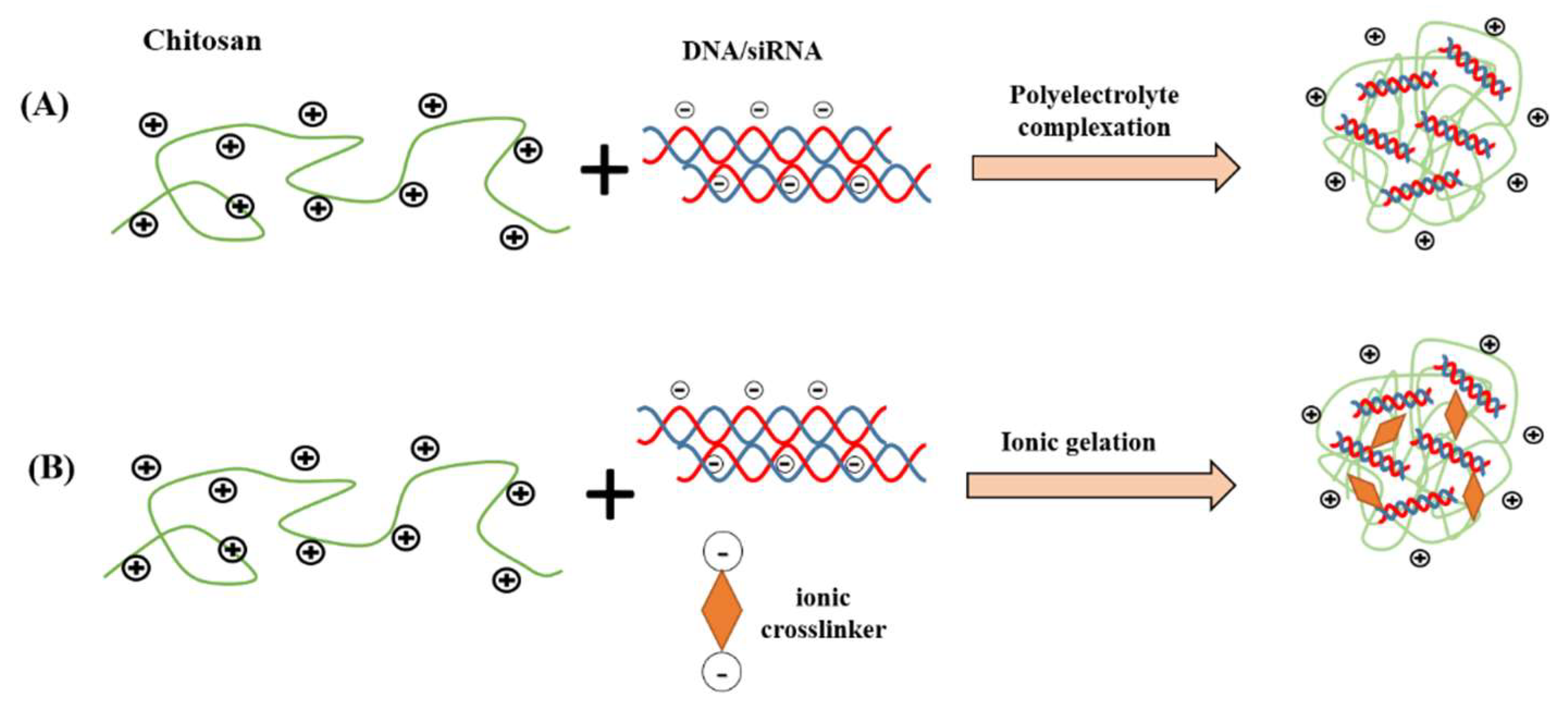
3. Preparation Methods of Chitosan Micro/Nano-Particles
4. Chitosan Based Formulations for siRNA/DNA Delivery
4.1. Formulations with Enhanced Stability
4.2. Formulations with Enhanced Cell Penetration
4.3. Formulations with Enhanced DNA/siRNA Release
4.4. Formulations with Enhanced siRNA Prolonged Gene Silencing
4.5. Recent Enhanced siRNA/DNA Delivery Systems
5. Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Anguela, X.M.; High, K.A. Entering the modern era of gene therapy. Annu. Rev. Med. 2019, 70, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Büning, H.; Ling, C. Rational Design of Gene Therapy Vectors. Mol. Ther. Methods Clin. Dev. 2019, 12, 246–247. [Google Scholar] [CrossRef] [PubMed]
- Manfredsson, F.P.; Benskey, M.J. Viral Vectors for Gene Therapy: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Hidai, C.; Kitano, H. Nonviral gene therapy for cancer: A review. Diseases 2018, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Panizo, M.V.; Domínguez-Bajo, A.; Portolés, M.T.; Serrano, M.C.; Marciello, M. Chapter 11—Nonviral Gene Therapy: Design and Application of Inorganic Nanoplexes. In Nucleic Acid Nanotheranostics; Filice, M., Ruiz-Cabello, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 365–390. [Google Scholar]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, T.; Kamimura, K.; Kanefuji, T.; Suda, T.; Terai, S. Nucleic Acid-Based Therapy: Development of a Nonviral-Based Delivery Approach. In In Vivo and Ex Vivo Gene Therapy for Inherited and Non-Inherited Disorders; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar]
- Guo, X.; Huang, L. Recent Advances in Nonviral Vectors for Gene Delivery. Acc. Chem. Res. 2012, 45, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Ozpolat, B.; Sood, A.K.; Lopez-Berestein, G. Liposomal siRNA nanocarriers for cancer therapy. Adv. Drug Deliv. Rev. 2014, 66, 110–116. [Google Scholar] [CrossRef]
- Crespo-Barreda, A.; Encabo-Berzosa, M.M.; González-Pastor, R.; Ortíz-Teba, P.; Iglesias, M.; Serrano, J.L.; Martin-Duque, P. Chapter 11—Viral and Nonviral Vectors for In Vivo and Ex Vivo Gene Therapies. In Translating Regenerative Medicine to the Clinic; Laurence, J., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 155–177. [Google Scholar]
- Lostalé-Seijo, I.; Montenegro, J. Synthetic materials at the forefront of gene delivery. Nat. Rev. Chem. 2018, 2, 258–277. [Google Scholar] [CrossRef]
- Arya, G.; Mankamna Kumari, R.; Sharma, N.; Gupta, N.; Chandra, R.; Nimesh, S. Chapter 18—Polymeric nanocarriers for site-specific gene therapy. In Drug Targeting and Stimuli Sensitive Drug Delivery Systems; Grumezescu, A.M., Ed.; William Andrew Publishing: Norwich, NY, USA, 2018; pp. 689–714. [Google Scholar]
- Kritchenkov, A.S.; Andranovitš, S.; Skorik, Y.A. Chitosan and its derivatives: Vectors in gene therapy. Russ. Chem. Rev. 2017, 86, 231–239. [Google Scholar] [CrossRef]
- Opanasopit, P.; Tragulpakseerojn, J.; Apirakaramwong, A.; Ngawhirunpat, T.; Rojanarata, T.; Ruktanonchai, U. The development of poly-L-arginine-coated liposomes for gene delivery. Int. J. Nanomed. 2011, 6, 2245–2252. [Google Scholar] [CrossRef]
- Alameh, M.; Lavertu, M.; Tran-Khanh, N.; Chang, C.-Y.; Lesage, F.; Bail, M.; Darras, V.; Chevrier, A.; Buschmann, M.D. siRNA delivery with chitosan: Influence of chitosan molecular weight, degree of deacetylation, and amine to phosphate ratio on in vitro silencing efficiency, hemocompatibility, biodistribution, and in vivo efficacy. Biomacromolecules 2017, 19, 112–131. [Google Scholar] [CrossRef] [PubMed]
- Ragelle, H.; Vandermeulen, G.; Préat, V. Chitosan-based siRNA delivery systems. J. Control. Release 2013, 172, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Howard, K.A.; Dong, M.; Andersen, M.Ø.; Rahbek, U.L.; Johnsen, M.G.; Hansen, O.C.; Besenbacher, F.; Kjems, J. The influence of polymeric properties on chitosan/siRNA nanoparticle formulation and gene silencing. Biomaterials 2007, 28, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Liu, G.; Sun, B.; Zhou, S.; Zuo, A.; Zhao, R.; Liang, D. Phosphorylatable short peptide conjugated low molecular weight chitosan for efficient siRNA delivery and target gene silencing. Int. J. Pharm. 2012, 422, 445–453. [Google Scholar] [CrossRef] [PubMed]
- MacLaughlin, F.C.; Mumper, R.J.; Wang, J.; Tagliaferri, J.M.; Gill, I.; Hinchcliffe, M.; Rolland, A.P. Chitosan and depolymerized chitosan oligomers as condensing carriers for in vivo plasmid delivery. J. Control. Release 1998, 56, 259–272. [Google Scholar] [CrossRef]
- Huang, M.; Fong, C.-W.; Khor, E.; Lim, L.-Y. Transfection efficiency of chitosan vectors: Effect of polymer molecular weight and degree of deacetylation. J. Control. Release 2005, 106, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Malmo, J.; Sørgård, H.; Vårum, K.M.; Strand, S.P. siRNA delivery with chitosan nanoparticles: Molecular properties favoring efficient gene silencing. J. Control. Release 2012, 158, 261–268. [Google Scholar] [CrossRef]
- Sato, T.; Ishii, T.; Okahata, Y. In vitro gene delivery mediated by chitosan. Effect of pH, serum, and molecular mass of chitosan on the transfection efficiency. Biomaterials 2001, 22, 2075–2080. [Google Scholar] [CrossRef]
- Chuan, D.; Jin, T.; Fan, R.; Zhou, L.; Guo, G. Chitosan for gene delivery: Methods for improvement and applications. Adv. Colloid Interface Sci. 2019, 268, 25–38. [Google Scholar] [CrossRef]
- Jiang, H.-L.; Xing, L.; Luo, C.-Q.; Zhou, T.-J.; Li, H.-S.; Cho, C.-S. Chemical Modification of Chitosan as a Gene Transporter. Curr. Org. Chem. 2018, 22, 668–689. [Google Scholar] [CrossRef]
- Layek, B.; Singh, J. 8—Chitosan for DNA and gene therapy. In Chitosan Based Biomaterials Volume 2; Jennings, J.A., Bumgardner, J.D., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 209–244. [Google Scholar]
- Guţoaia, A.; Schuster, L.; Margutti, S.; Laufer, S.; Schlosshauer, B.; Krastev, R.; Stoll, D.; Hartmann, H. Fine-tuned PEGylation of chitosan to maintain optimal siRNA-nanoplex bioactivity. Carbohydr. Polym. 2016, 143, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Inamdar, N.; Mourya, V.K. Chitosan and Anionic Polymers—Complex Formation and Applications; Nova Science Publishers, Inc: New York, NY, USA, 2011; pp. 333–377. [Google Scholar]
- Yan, J.; Du, Y.-Z.; Chen, F.-Y.; You, J.; Yuan, H.; Hu, F.-Q. Effect of proteins with different isoelectric points on the gene transfection efficiency mediated by stearic acid grafted chitosan oligosaccharide micelles. Mol. Pharm. 2013, 10, 2568–2577. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Wu, J.; Yi, H.; Liu, J.; Lu, B.; Yuan, M.; Huang, X.; Yuan, H.; Hu, F. A spermine conjugated stearic acid-g-chitosan oligosaccharide polymer with different types of amino groups for efficient p53 gene therapy. Colloids Surf. B Biointerfaces 2016, 145, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, M.; Qian, R.; Liu, C.; Zong, X.; Li, Y.-Q.; Li, W. Binding efficacy and kinetics of chitosan with DNA duplex: The effects of deacetylation degree and nucleotide sequences. Carbohydr. Polym. 2017, 169, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Köping-Höggård, M.; Tubulekas, I.; Guan, H.; Edwards, K.; Nilsson, M.; Vårum, K.M.; Artursson, P. Chitosan as a nonviral gene delivery system. Structure–property relationships and characteristics compared with polyethylenimine in vitro and after lung administration in vivo. Gene Ther. 2001, 8, 1108. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Sun, W.; Kissel, T. Chitosan-based formulations for delivery of DNA and siRNA. Adv. Drug Deliv. Rev. 2010, 62, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, E.J.; Nielsen, J.M.; Becker, D.; Karlas, A.; Prakash, H.; Glud, S.Z.; Merrison, J.; Besenbacher, F.; Meyer, T.F.; Kjems, J. Pulmonary gene silencing in transgenic EGFP mice using aerosolised chitosan/siRNA nanoparticles. Pharm. Res. 2010, 27, 2520–2527. [Google Scholar] [CrossRef] [PubMed]
- Jean, M.; Alameh, M.; De Jesus, D.; Thibault, M.; Lavertu, M.; Darras, V.; Nelea, M.; Buschmann, M.D.; Merzouki, A. Chitosan-based therapeutic nanoparticles for combination gene therapy and gene silencing of in vitro cell lines relevant to type 2 diabetes. Eur. J. Pharm. Sci. 2012, 45, 138–149. [Google Scholar] [CrossRef]
- Ramesh Kumar, D.; Saravana Kumar, P.; Gandhi, M.R.; Al-Dhabi, N.A.; Paulraj, M.G.; Ignacimuthu, S. Delivery of chitosan/dsRNA nanoparticles for silencing of wing development vestigial (vg) gene in Aedes aegypti mosquitoes. Int. J. Biol. Macromol. 2016, 86, 89–95. [Google Scholar] [CrossRef]
- Rudzinski, W.E.; Palacios, A.; Ahmed, A.; Lane, M.A.; Aminabhavi, T.M. Targeted delivery of small interfering RNA to colon cancer cells using chitosan and PEGylated chitosan nanoparticles. Carbohydr. Polym. 2016, 147, 323–332. [Google Scholar] [CrossRef]
- Sadreddini, S.; Safaralizadeh, R.; Baradaran, B.; Aghebati-Maleki, L.; Hosseinpour-Feizi, M.A.; Shanehbandi, D.; Jadidi-Niaragh, F.; Sadreddini, S.; Kafil, H.S.; Younesi, V.; et al. Chitosan nanoparticles as a dual drug/siRNA delivery system for treatment of colorectal cancer. Immunol. Lett. 2017, 181, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Dehousse, V.; Garbacki, N.; Jaspart, S.; Castagne, D.; Piel, G.; Colige, A.; Evrard, B. Comparison of chitosan/siRNA and trimethylchitosan/siRNA complexes behaviour in vitro. Int. J. Biol. Macromol. 2010, 46, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Veilleux, D.; Nelea, M.; Biniecki, K.; Lavertu, M.; Buschmann, M.D. Preparation of Concentrated Chitosan/DNA Nanoparticle Formulations by Lyophilization for Gene Delivery at Clinically Relevant Dosages. J. Pharm. Sci. 2016, 105, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Lebre, F.; Borchard, G.; Faneca, H.; Pedroso de Lima, M.; Borges, O. Intranasal administration of novel chitosan nanoparticle/DNA complexes induces antibody response to hepatitis B surface antigen in mice. Mol. Pharm. 2016, 13, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yun, K.-S.; Choi, C.S.; Shin, S.-H.; Ban, H.-S.; Rhim, T.; Lee, S.K.; Lee, K.Y. T cell-specific siRNA delivery using antibody-conjugated chitosan nanoparticles. Bioconjugate Chem. 2012, 23, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Kean, T.; Thanou, M. Biodegradation, biodistribution and toxicity of chitosan. Adv. Drug Deliv. Rev. 2010, 62, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Matica, A.; Menghiu, G.; Ostafe, V. Toxicity of chitosan based products. New Front. Chem. 2017, 26, 65–74. [Google Scholar]
- Csaba, N.; Köping-Höggård, M.; Alonso, M.J. Ionically crosslinked chitosan/tripolyphosphate nanoparticles for oligonucleotide and plasmid DNA delivery. Int. J. Pharm. 2009, 382, 205–214. [Google Scholar] [CrossRef]
- Kean, T.; Roth, S.; Thanou, M. Trimethylated chitosans as non-viral gene delivery vectors: Cytotoxicity and transfection efficiency. J. Control. Release 2005, 103, 643–653. [Google Scholar] [CrossRef]
- Zubareva, A.; Shagdarova, B.; Varlamov, V.; Kashirina, E.; Svirshchevskaya, E. Penetration and toxicity of chitosan and its derivatives. Eur. Polym. J. 2017, 93, 743–749. [Google Scholar] [CrossRef]
- Richardson, S.W.; Kolbe, H.J.; Duncan, R. Potential of low molecular mass chitosan as a DNA delivery system: Biocompatibility, body distribution and ability to complex and protect DNA. Int. J. Pharm. 1999, 178, 231–243. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Bhattacharya, S.; Nandy, A.; Bhattacharyya, A.; Mishra, R.; Kundu, P.P. Assessment of in vivo chronic toxicity of chitosan and its derivates used as oral insulin carriers. Toxicol. Res. 2015, 4, 281–290. [Google Scholar] [CrossRef]
- Tapola, N.S.; Lyyra, M.L.; Kolehmainen, R.M.; Sarkkinen, E.S.; Schauss, A.G. Safety aspects and cholesterol-lowering efficacy of chitosan tablets. J. Am. Coll. Nutr. 2008, 27, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, M.; Tomaro-Duchesneau, C.; Saha, S.; Kahouli, I.; Prakash, S. Development and characterization of chitosan-PEG-TAT nanoparticles for the intracellular delivery of siRNA. Int. J. Nanomed. 2013, 8, 2041–2052. [Google Scholar]
- Bozkir, A.; Saka, O. Chitosan Nanoparticles for Plasmid DNA Delivery: Effect of Chitosan Molecular Structure on Formulation and Release Characteristics. Drug Deliv. 2004, 11, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.; Ramesh, R. Multifaceted Applications of Chitosan in Cancer Drug Delivery and Therapy. Mar. Drugs 2017, 15, 96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mysore, K.; Flannery, E.; Michel, K.; Severson, D.W.; Zhu, K.Y.; Duman-Scheel, M. Chitosan/interfering RNA nanoparticle mediated gene silencing in disease vector mosquito larvae. J. Vis. Exp. Jove 2015, 97, e52523. [Google Scholar] [CrossRef] [PubMed]
- Ragelle, H.; Riva, R.; Vandermeulen, G.; Naeye, B.; Pourcelle, V.; Le Duff, C.S.; D’Haese, C.; Nysten, B.; Braeckmans, K.; De Smedt, S.C.; et al. Chitosan nanoparticles for siRNA delivery: Optimizing formulation to increase stability and efficiency. J. Control. Release 2014, 176, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Somavarapu, S.; Colombani, A.; Govind, N.; Taylor, K.M.G. Nebulised siRNA encapsulated crosslinked chitosan nanoparticles for pulmonary delivery. Int. J. Pharm. 2013, 455, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Song, X.; Yang, C.; Gao, S.; Klausen, L.H.; Zhang, Y.; Dong, M.; Kjems, J. Chitosan/siRNA functionalized titanium surface via a layer-by-layer approach for in vitro sustained gene silencing and osteogenic promotion. Int. J. Nanomed. 2015, 10, 2335–2346. [Google Scholar]
- Labala, S.; Jose, A.; Venuganti, V.V.K. Transcutaneous iontophoretic delivery of STAT3 siRNA using layer-by-layer chitosan coated gold nanoparticles to treat melanoma. Colloids Surf. B Biointerfaces 2016, 146, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.; Hill, A.; Uezguen, S.; Weigandt, M.; Goepferich, A. Analysis of immediate stress mechanisms upon injection of polymeric micelles and related colloidal drug carriers: Implications on drug targeting. Biomacromolecules 2012, 13, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Buyens, K.; De Smedt, S.C.; Braeckmans, K.; Demeester, J.; Peeters, L.; van Grunsven, L.A.; du Jeu, X.M.; Sawant, R.; Torchilin, V.; Farkasova, K. Liposome based systems for systemic siRNA delivery: Stability in blood sets the requirements for optimal carrier design. J. Control. Release 2012, 158, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, S.; Strand, S.; de Lange Davies, C.; Stokke, B.T. Glycosaminoglycan destabilization of DNA–chitosan polyplexes for gene delivery depends on chitosan chain length and GAG properties. Biochim. Biophys. Acta Gen. Subj. 2005, 1721, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Ruponen, M.; Honkakoski, P.; Tammi, M.; Urtti, A. Cell-surface glycosaminoglycans inhibit cation-mediated gene transfer. J. Gene Med. Cross Discip. J. Res. Sci. Gene Transf. Clin. Appl. 2004, 6, 405–414. [Google Scholar] [CrossRef]
- Mao, H.-Q.; Roy, K.; Troung-Le, V.L.; Janes, K.A.; Lin, K.Y.; Wang, Y.; August, J.T.; Leong, K.W. Chitosan-DNA nanoparticles as gene carriers: Synthesis, characterization and transfection efficiency. J. Control. Release 2001, 70, 399–421. [Google Scholar] [CrossRef]
- Strand, S.P.; Issa, M.M.; Christensen, B.E.; Vårum, K.M.; Artursson, P. Tailoring of chitosans for gene delivery: Novel self-branched glycosylated chitosan oligomers with improved functional properties. Biomacromolecules 2008, 9, 3268–3276. [Google Scholar] [CrossRef]
- Hashimoto, M.; Morimoto, M.; Saimoto, H.; Shigemasa, Y.; Sato, T. Lactosylated chitosan for DNA delivery into hepatocytes: The effect of lactosylation on the physicochemical properties and intracellular trafficking of pDNA/chitosan complexes. Bioconjugate Chem. 2006, 17, 309–316. [Google Scholar] [CrossRef]
- Wang, B.; He, C.; Tang, C.; Yin, C. Effects of hydrophobic and hydrophilic modifications on gene delivery of amphiphilic chitosan based nanocarriers. Biomaterials 2011, 32, 4630–4638. [Google Scholar] [CrossRef]
- Katas, H.; Alpar, H.O. Development and characterisation of chitosan nanoparticles for siRNA delivery. J. Control. Release 2006, 115, 216–225. [Google Scholar] [CrossRef]
- Thanou, M.; Florea, B.; Geldof, M.; Junginger, H.; Borchard, G. Quaternized chitosan oligomers as novel gene delivery vectors in epithelial cell lines. Biomaterials 2002, 23, 153–159. [Google Scholar] [CrossRef]
- Lee, S.J.; Huh, M.S.; Lee, S.Y.; Min, S.; Lee, S.; Koo, H.; Chu, J.U.; Lee, K.E.; Jeon, H.; Choi, Y. Tumor-homing poly-siRNA/glycol chitosan self-cross-linked nanoparticles for systemic siRNA delivery in cancer treatment. Angew. Chem. Int. Ed. 2012, 51, 7203–7207. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Dai, H.; Leong, K.W.; Goh, S.H.; Mao, H.Q.; Yang, Y.Y. Chitosan-g-PEG/DNA complexes deliver gene to the rat liver via intrabiliary and intraportal infusions. J. Gene Med. Cross Discip. J. Res. Sci. Gene Transf. Clin. Appl. 2006, 8, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Ping, Y.; Liu, C.; Zhang, Z.; Liu, K.L.; Chen, J.; Li, J. Chitosan-graft-(PEI-β-cyclodextrin) copolymers and their supramolecular PEGylation for DNA and siRNA delivery. Biomaterials 2011, 32, 8328–8341. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Osburg, B.; Petersen, H.; Kissel, T.; Bickel, U. Effect of poly (ethylene imine) molecular weight and pegylation on organ distribution and pharmacokinetics of polyplexes with oligodeoxynucleotides in mice. Drug Metab. Dispos. 2004, 32, 983–992. [Google Scholar] [PubMed]
- Lee, H.; Jeong, J.H.; Park, T.G. PEG grafted polylysine with fusogenic peptide for gene delivery: High transfection efficiency with low cytotoxicity. J. Control. Release 2002, 79, 283–291. [Google Scholar] [CrossRef]
- Braeckmans, K.; Buyens, K.; Bouquet, W.; Vervaet, C.; Joye, P.; Vos, F.D.; Plawinski, L.; Doeuvre, L.; Angles-Cano, E.; Sanders, N.N. Sizing nanomatter in biological fluids by fluorescence single particle tracking. Nano Lett. 2010, 10, 4435–4442. [Google Scholar] [CrossRef] [PubMed]
- Buyens, K.; Meyer, M.; Wagner, E.; Demeester, J.; De Smedt, S.C.; Sanders, N.N. Monitoring the disassembly of siRNA polyplexes in serum is crucial for predicting their biological efficacy. J. Control. Release 2010, 141, 38–41. [Google Scholar] [CrossRef]
- Morrissey, D.V.; Lockridge, J.A.; Shaw, L.; Blanchard, K.; Jensen, K.; Breen, W.; Hartsough, K.; Machemer, L.; Radka, S.; Jadhav, V.; et al. Potent and persistent in vivo anti-HBV activity of chemically modified siRNAs. Nat. Biotechnol. 2005, 23, 1002–1007. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Z.; Chen, L.; Gu, W.; Li, Y. Chitosan N-betainates/DNA self-assembly nanoparticles for gene delivery: In vitro uptake and transfection efficiency. Int. J. Pharm. 2009, 371, 156–162. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Zhang, Y.; Pan, Y.; Zhao, J.; Ren, L.; Liao, M.; Hu, Z.; Kong, L.; Wang, J. A novel PEGylation of chitosan nanoparticles for gene delivery. Biotechnol. Appl. Biochem. 2007, 46, 197–204. [Google Scholar] [PubMed]
- Nimesh, S.; Thibault, M.M.; Lavertu, M.; Buschmann, M.D. Enhanced gene delivery mediated by low molecular weight chitosan/DNA complexes: Effect of pH and serum. Mol. Biotechnol. 2010, 46, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Thibault, M.; Nimesh, S.; Lavertu, M.; Buschmann, M.D. Intracellular trafficking and decondensation kinetics of chitosan–pDNA polyplexes. Mol. Ther. 2010, 18, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.-G.; Wei, W.; Lv, P.-P.; Yue, H.; Wang, L.-Y.; Su, Z.-G.; Ma, G.-H. Surface charge affects cellular uptake and intracellular trafficking of chitosan-based nanoparticles. Biomacromolecules 2011, 12, 2440–2446. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, M.D.; Merzouki, A.; Lavertu, M.; Thibault, M.; Jean, M.; Darras, V. Chitosans for delivery of nucleic acids. Adv. Drug Deliv. Rev. 2013, 65, 1234–1270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Mardyani, S.; Chan, W.C.; Kumacheva, E. Design of biocompatible chitosan microgels for targeted pH-mediated intracellular release of cancer therapeutics. Biomacromolecules 2006, 7, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.; Kurisawa, M.; Chung, J.E.; Yang, Y.-Y. Synthesis and characterization of chitosan-g-poly (ethylene glycol)-folate as a non-viral carrier for tumor-targeted gene delivery. Biomaterials 2007, 28, 540–549. [Google Scholar] [CrossRef]
- Kim, T.H.; Nah, J.W.; Cho, M.-H.; Park, T.G.; Cho, C.S. Receptor-mediated gene delivery into antigen presenting cells using mannosylated chitosan/DNA nanoparticles. J. Nanosci. Nanotechnol. 2006, 6, 2796–2803. [Google Scholar] [CrossRef]
- Gao, S.; Chen, J.; Xu, X.; Ding, Z.; Yang, Y.-H.; Hua, Z.; Zhang, J. Galactosylated low molecular weight chitosan as DNA carrier for hepatocyte-targeting. Int. J. Pharm. 2003, 255, 57–68. [Google Scholar] [CrossRef]
- Park, I.; Kim, T.; Park, Y.; Shin, B.; Choi, E.; Chowdhury, E.; Akaike, T.; Cho, C. Galactosylated chitosan-graft-poly (ethylene glycol) as hepatocyte-targeting DNA carrier. J. Control. Release 2001, 76, 349–362. [Google Scholar] [CrossRef]
- Kim, T.H.; Park, I.K.; Nah, J.W.; Choi, Y.J.; Cho, C.S. Galactosylated chitosan/DNA nanoparticles prepared using water-soluble chitosan as a gene carrier. Biomaterials 2004, 25, 3783–3792. [Google Scholar] [CrossRef] [PubMed]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and development of polymers for gene delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, M.; Tomaro-Duchesneau, C.; Prakash, S. Synthesis of TAT peptide-tagged PEGylated chitosan nanoparticles for siRNA delivery targeting neurodegenerative diseases. Biomaterials 2013, 34, 1270–1280. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, J.; Szoka, F.C. Nucleic acid delivery: The missing pieces of the puzzle? Acc. Chem. Res. 2012, 45, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E. Application of membrane-active peptides for nonviral gene delivery. Adv. Drug Deliv. Rev. 1999, 38, 279–289. [Google Scholar] [CrossRef]
- Li, W.; Nicol, F.; Szoka, F.C., Jr. GALA: A designed synthetic pH-responsive amphipathic peptide with applications in drug and gene delivery. Adv. Drug Deliv. Rev. 2004, 56, 967–985. [Google Scholar] [CrossRef] [PubMed]
- El Ouahabi, A.; Thiry, M.; Pector, V.; Fuks, R.; Ruysschaert, J.M.; Vandenbranden, M. The role of endosome destabilizing activity in the gene transfer process mediated by cationic lipids. FEBS Lett. 1997, 414, 187–192. [Google Scholar]
- Ma, Z.; Yang, C.; Song, W.; Wang, Q.; Kjems, J.; Gao, S. Chitosan hydrogel as siRNA vector for prolonged gene silencing. J. Nanobiotechnol. 2014, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Revuri, V.; Lee, S.-J.; Cho, S.; Park, I.-K.; Cho, K.J.; Bae, W.K.; Lee, Y.-K. Oral siRNA delivery to treat colorectal liver metastases. ACS Nano 2017, 11, 10417–10429. [Google Scholar] [CrossRef] [PubMed]
- Mittnacht, U.; Hartmann, H.; Hein, S.; Oliveira, H.; Dong, M.; Pêgo, A.P.; Kjems, J.; Howard, K.A.; Schlosshauer, B. Chitosan/siRNA nanoparticles biofunctionalize nerve implants and enable neurite outgrowth. Nano Lett. 2010, 10, 3933–3939. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, Z.-Y.; Zhang, J.; Zhang, Y.; Huo, H.; Wang, T.; Jiang, T.; Wang, S. RVG-peptide-linked trimethylated chitosan for delivery of siRNA to the brain. Biomacromolecules 2014, 15, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Yhee, J.Y.; Song, S.; Lee, S.J.; Park, S.-G.; Kim, K.-S.; Kim, M.G.; Son, S.; Koo, H.; Kwon, I.C.; Jeong, J.H. Cancer-targeted MDR-1 siRNA delivery using self-cross-linked glycol chitosan nanoparticles to overcome drug resistance. J. Control. Release 2015, 198, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, S.H.; Oh, M.H.; Kim, J.S.; Park, T.G.; Nam, Y.S. Prolonged gene silencing by siRNA/chitosan-g-deoxycholic acid polyplexes loaded within biodegradable polymer nanoparticles. J. Control. Release 2012, 162, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Baghdan, E.; Pinnapireddy, S.R.; Strehlow, B.; Engelhardt, K.H.; Schäfer, J.; Bakowsky, U. Lipid coated chitosan-DNA nanoparticles for enhanced gene delivery. Int. J. Pharm. 2018, 535, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.N.; Han, Z.; Merwin, M.; Al Taai, M.; Conley, S.M.; Naash, M.I. Synthesis and characterization of glycol chitosan DNA nanoparticles for retinal gene delivery. ChemMedChem 2014, 9, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Shiraki, K.; Yasuda, R.; Danjo, K.; Watanabe, Y. Chitosan–interferon-β gene complex powder for inhalation treatment of lung metastasis in mice. J. Control. Release 2011, 150, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Raftery, R.M.; Tierney, E.G.; Curtin, C.M.; Cryan, S.-A.; O’Brien, F.J. Development of a gene-activated scaffold platform for tissue engineering applications using chitosan-pDNA nanoparticles on collagen-based scaffolds. J. Control. Release 2015, 210, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Raftery, R.M.; Castaño, I.M.; Chen, G.; Cavanagh, B.; Quinn, B.; Curtin, C.M.; Cryan, S.A.; O’Brien, F.J. Translating the role of osteogenic-angiogenic coupling in bone formation: Highly efficient chitosan-pDNA activated scaffolds can accelerate bone regeneration in critical-sized bone defects. Biomaterials 2017, 149, 116–127. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Chitosan MW (kDa) | Genes | Chitosan Type | DD (%) | N/P Ratios | Polyplex Size [32] | Cell Type | Transfection Efficiency (%) | References |
|---|---|---|---|---|---|---|---|---|
| 10 | siRNA | Chitosan | 80, 92 | 5, 10 | 78–235 | HepG2 | 56, 72 | [35] |
| Medium MW(Sigma) | dsRNA | Chitosan | 75 | - | 100–200 | Sf21 cell line | 85 | [36] |
| Low MW(Sigma) | siRNA | PEGylated chitosan/ Chitosan | - | - | 100–150 | HCT-116 | 72/69 | [37] |
| 400 | siRNA | Chitosan (with dextran) | 87.3 | - | 111–172 | HCT-116 | - | [38] |
| 5~15 | siRNA | Chitosan | 84 | 30 | 108 | HEK293 | 3.9 | [39] |
| 150~400 | Chitosan | 86 | 187 | 12.3 | ||||
| 42~50 | Trimethyl chitosan | 84 | 138–162 | 24.3–37.1 | ||||
| 7.5-11.3 | pDNA | Chitosan | 92–94 | - | 80–100 | HEK293 | >65 | [40] |
| Low MW | pDNA | Chitosan | 90 | 5,7.5,10 | 300–1000 | A549 | - | [41] |
| 470 | siRNA | Chitosan glutamate | 86 | - | 311–350 | Jurkat, A3.01 | 90 | [42] |
| Disease Model/Goal | siRNA Formulation | Target Gene | Protein/mRNA Silencing (%) | Cell Model | References |
| Enhance bone binding ability in bone implant materials. | Multilayers of sodium hyaluronate and chitosan/siRNA nanoparticles on smooth titanium surfaces via Layer-by-Layer. | GFP, Ckip-1 | Reduction of GFP at day 3, 5 and 7 by 70–80%. Osteogenic differentiation of MG63 increased with Ckip-1 silenced. Both in vitro. | MG63, H1299 | [57] |
| Protect alveolar bone loss in periodontal diseases. | Thermosensitive chitosan hydrogel as siRNA reservoir and vector. | R-RANK | Reduction of mRNA at day 3 (30%), day 5 (50%) and day 7 (60%) in vitro. | RAW264.7 | [95] |
| Promote nerve regeneration and local nanotherapeutics delivery. | Polymer filaments nerve implants loaded with chitosan/siRNA nanoparticles. | RhoA | Reduction of mRNA at day 2 (65–75%). Undisclosed reduction of protein at day 3. | PC12 | [98] |
| Delivery of therapeutic molecules to the brain for the treatment of Neurodegenerative diseases. | Chemoselective conjugation of monomethoxy PEG, at the C2 hydroxyl group of chitosan polymer, with conjugation of PEG to a cell-penetrating peptide, Trans-Activator of Transcription. | Ataxin-1 | Reduction of protein (100%) at day 2 in vitro. | Neuro2a | [91] |
| Efficient delivery of siRNA to the brain to combat Alzheimer's disease. | A peptide derived from rabies virus glycoprotein linked to siRNA/trimethylated chitosan through PEG. | BACE1 | Reduction of protein (50–57%) after 2 days in vitro. | Neuro2a | [99] |
| Self-crosslinking nanoparticle to deliver polymerized siRNAs for tumour targeting cancer treatment. | Forming stable nanoparticles of thiolated glycol chitosan with poly-siRNA through charge–charge interactions and self-cross-linking simultaneously. | VEGF | Reduction of mRNA (95%) after 2 days in vitro. Reduction in mRNA (64%) after 24 days in vivo (particles injected every 3 days). | PC-3/tumour-bearing mice | [69] |
| Treatment of multi-drug resistance for tumour treatment. | Self-polymerized 5′-end thiol-modified siRNA incorporated to chitosan nanoparticles. | Pgp | Reduction of protein (62%) after 2 days in vitro. Reduction of protein (92%) after 22 days in vivo (particles injected every 4 days.) | MCF-7/ADR/MCF-7/ADR mice | [100] |
| Improving structural stability of siRNA for prolonged therapeutic efficacy. | Nanoparticles of siRNA/chitosan grafted with deoxycholates poly (D, L-lactic-co glycolic acid) (PLGA) | GFP | Reduction of GFP at 5 hour (60%) and 2 days (36%). Reduction of GFP at day 7 (undisclosed%) in vitro. | MDA-MB-435-GFP | [42] |
| Disease Model/Goal | DNA Carrier Formulation | Target Gene | Protein/mRNA Expression | Cell Model | References |
| Enhanced DNA delivery vehicle to efficiently transfect cells under physiological conditions. | Liposome (DPPC, cholesterol) encapsulated chitosan/DNA nanoparticles. | Luciferase/ GFP | Luciferase expression observed in vitro. GFP expression observed in vivo. | HEK 293/ chorioallantic membrane model | [101] |
| DNA delivery nanoparticle to efficiently transfect intraocular retinal cells under physiological conditions. | Non-viral nanoparticles composed of glycol chitosan and plasmid DNA. | CBA-eGFP | GFP expression observed in vivo at day 14 post-injection in the retinal pigment epithelium. | Adult wild-type albino mice (eyes). | [102] |
| Examine effectiveness of dry gene powders for treating lung metastasis. | Dry chitosan-DNA powders prepared by dispersing a chitosan–DNA solution into supercritical carbon dioxide | Luciferase/ GFP/Muβ | Luciferase and GFP expressions observed. Lung metastasis suppressed at day 21. | CT26/mice with CT26 | [103] |
| Enhanced stability and efficacy of DNA delivery vehicle. | Chitosan/DNA complex formulations with chitosan of varying molecular weights. | GFP | GFP expression observed (13 days). Reverse transfection showed 150% increased efficiency compared to standard protocol. | HEK 293 | [103] |
| Development of gene-activated collagen scaffold with chitosan/DNA nanoparticles for tissue engineering. | Oligomeric chitosan with DNA complexation followed by further crosslinking with TPP. Particles are soak-loaded onto hydrated collagen scaffold. | Luciferase/GFP | Luciferase expression showed overall transgene expression. GFP expression sustained over 14 days. | mesenchymal stem cells (MSC) | [104] |
| Using efficient chitosan/DNA activated scaffolds to accelerate bone regeneration in critical-sized bone defects. | Chitosan/DNA complex soak loaded into fabricated collagen scaffold. | BMP-2/VEGF/GFP | Both BMP-2 and VEGF expression observed in vitro over 14 days. GFP expression in in filtrating host cells at day 7 post in vivo implantation of scaffold. | mesenchymal stem cells (MSC)/calvaria of Wistar rats | [105] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Tan, Y.F.; Wong, Y.S.; Liew, M.W.J.; Venkatraman, S. Recent Advances in Chitosan-Based Carriers for Gene Delivery. Mar. Drugs 2019, 17, 381. https://doi.org/10.3390/md17060381
Cao Y, Tan YF, Wong YS, Liew MWJ, Venkatraman S. Recent Advances in Chitosan-Based Carriers for Gene Delivery. Marine Drugs. 2019; 17(6):381. https://doi.org/10.3390/md17060381
Chicago/Turabian StyleCao, Ye, Yang Fei Tan, Yee Shan Wong, Melvin Wen Jie Liew, and Subbu Venkatraman. 2019. "Recent Advances in Chitosan-Based Carriers for Gene Delivery" Marine Drugs 17, no. 6: 381. https://doi.org/10.3390/md17060381
APA StyleCao, Y., Tan, Y. F., Wong, Y. S., Liew, M. W. J., & Venkatraman, S. (2019). Recent Advances in Chitosan-Based Carriers for Gene Delivery. Marine Drugs, 17(6), 381. https://doi.org/10.3390/md17060381