In Vitro Studies Reveal Antiurolithic Effect of Antioxidant Sulfated Polysaccharides from the Green Seaweed Caulerpa cupressoides var flabellata



Abstract
1. Introduction
2. Results and Discussion
2.1. C. cupressoides’s Sulfated Polysaccharides Extraction, Chemical and Physicochemical Analysis
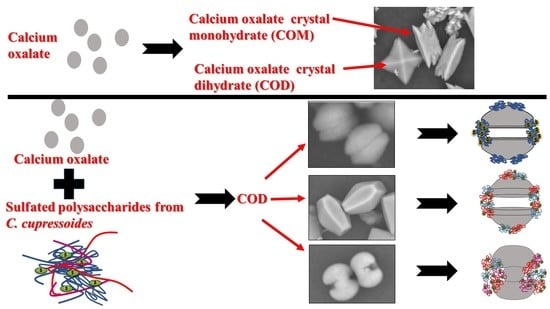
2.2. Effect of Sulfated Polysaccharides C. cupressoides on the Formation of Calcium Oxalate Crystals
2.3. Morphology of the Crystals Formed in the Presence of Caulerpa polysaccharides
2.4. FT-IR Spectrum Analyses
2.5. Zeta Potential
2.6. Fluorescence and Flow Cytometry Analyses of CaOx Crystals and FITC-Labeled SPs
2.7. Stabilization of COD Crystals
3. Materials and Methods
3.1. Sulfated Polysaccharides Extraction from Green Seaweed C. cupressoides
3.2. Chemical and Physicochemical Analysis
3.3. Agarose Gel Electrophoresis in 1,3-Diamino Propane Acetate Buffer (PDA)
3.4. Gel Permeation Chromatography (GPC)
3.5. Fourier Transformed Infrared (FT-IR) Spectroscopy Analysis
3.6. Calcium Oxalate Crystallization Assay
3.7. Scanning Electron Microscopy (SEM) and Dispersive Energy Spectroscopy (EDS)
3.8. Zeta Potential (ζ) Measurements
3.9. Light Microscopy and Fluorescence Microscopy for the Analysis of CaOx Crystal Morphology
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Afsar, B.; Kiremit, M.C.; Sag, A.A.; Tarim, K.; Acar, O.; Esen, T.; Solak, Y.; Covic, A.; Kanbay, M. The role of sodium intake in nephrolithiasis: epidemiology, pathogenesis, and future directions. Eur. J. Intern. Med. 2016, 35, 16–19. [Google Scholar] [CrossRef]
- Sofia, N.H.; Walter, T.M.; Sanatorium, T. Prevalence and risk factors of kidney stone. GJRA 2016, 5, 183–187. [Google Scholar]
- Honeycutt, A.A.; Segel, J.E.; Zhuo, X.; Hoerger, T.J.; Imai, K.; Williams, D. Medical costs of CKD in the medicare population. J. Am. Soc. Nephrol. 2013, 24, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Kerr, M.; Bray, B.; Medcalf, J.; O’Donoghue, D.J.; Matthews, B. Estimating the financial cost of chronic kidney disease to the NHS in England. Nephrol. Dial. Transp. 2012, 27, iii73–iii80. [Google Scholar] [CrossRef] [PubMed]
- Daudon, M.; Bazin, D.; Letavernier, E. Randall’s plaque as the origin of calcium oxalate kidney stones. Urolithiasis 2014, 43, 5–11. [Google Scholar] [CrossRef]
- Fong-Ngern, K.; Peerapen, P.; Sinchaikul, S.; Chen, S.T.; Thongboonkerd, V. Large-scale identification of calcium oxalate monohydrate crystal-binding proteins on apical membrane of distal renal tubular epithelial cells. J. Proteome Res. 2011, 10, 4463–4477. [Google Scholar] [CrossRef]
- Parvaneh, L.S.; Donadio, D.; Sulpizi, M. Molecular mechanism of crystal growth inhibition at the calcium oxalate/water interfaces. J. Phys. Chem. C 2016, 120, 4410–4417. [Google Scholar] [CrossRef]
- Grases, F.; Millan, A.; Conte, A. Production of calcium-oxalate monohydrate, dihydrate or trihydrate—A comparative-study. Urol. Res. 1990, 18, 17–20. [Google Scholar] [CrossRef]
- Queiroz, M.F.; Melo, K.R.T.; Sabry, D.A.; Sassaki, G.L.; Rocha, H.A.O. Does the use of chitosan contribute to oxalate kidney stone formation? Mar. Drugs 2015, 13, 141–158. [Google Scholar] [CrossRef]
- Ratkalkar, V.N.; Kleinman, J.G. Mechanisms of stone formation. Clin. Rev. Bone Miner. Metab. 2011, 9, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Xue, J.F.; Tan, C.Y.; Gui, B.S.; Sun, X.Y.; Ouyang, J.M. Inhibition of urinary macromolecule heparin on aggregation of nano-COM and nano-COD crystals. Molecules 2015, 20, 1626–1642. [Google Scholar] [CrossRef]
- Li, J.J.; Xue, J.F.; Ouyang, J.M. Stabilization of submicron calcium oxalate suspension by chondroitin sulfate C may be an efficient protection from stone formation. Bioinorg. Chem. Appl. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Bhadja, P.; Tan, C.Y.; Ouyang, J.M.; Yu, K. Repair effect of seaweed polysaccharides with different contents of sulfate group and molecular weights on damaged HK–2 Cells. Polymers 2016, 8, 188. [Google Scholar] [CrossRef]
- Ouyang, J.M.; Wang, M.; Lu, P.; Tan, J. Degradation of sulfated polysaccharide extracted from algal Laminaria japonica and its modulation on calcium oxalate crystallization. Mater. Sci. Eng. C 2010, 30, 1022–1029. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Wu, W.H.; Wang, J.; Lan, M.B. Antioxidant properties of polysaccharide from the brown seaweed Sargassum graminifolium (Turn.), and its effects on calcium oxalate crystallization. Mar. Drugs 2012, 10, 119–130. [Google Scholar] [CrossRef]
- Melo, K.R.T.; Camara, R.B.G.; Queiroz, M.F.; Vidal, A.A.J.; Lima, C.R.M.; Melo-Silveira, R.F.; Almeida-Lima, J.; Rocha, H.A.O. Evaluation of sulfated polysaccharides from the brown seaweed Dictyopteris justii as antioxidant agents and as inhibitors of the formation of calcium oxalate crystals. Molecules 2013, 18, 14543–14563. [Google Scholar] [CrossRef]
- Ouyang, J.M.; Wu, X.M. Morphological and phase changes in calcium oxalate crystals induced by sulfated polysaccharide extracted from algae Eucheuma striatum. Chem. Lett. 2005, 34, 1296–1297. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Rajesh, N.G.; Varalakshmi, P. Mitochondrial dysfunction in an animal model of hyperoxaluria: A prophylactic approach with fucoidan. Eur. J. Pharmacol. 2008, 579, 330–336. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P.; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: The protective role of fucoidan. Life Sci. 2006, 79, 1789–1795. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P. Beneficial role of sulfated polysaccharides from edible seaweed Fucus vesiculosus in experimental hyperoxaluria. Food Chem. 2007, 100, 1552–1559. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Kong, T.; Wu, W.H.; Lan, M.B. The protection of polysaccharide from the brown seaweed Sargassum graminifolium against ethylene glycol–induced mitochondrial damage. Mar. Drugs 2013, 11, 870–880. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida–Lima, J.; Farias, E.H.C.; Leite, E.L.; Rocha, H.A.O. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Barbosa, J.S.; Costa, M.S.S.P.; Melo, L.F.M.; Medeiros, M.J.C.; Pontes, D.L.; Scortecci, K.C.; Rocha, H.A.O. In Vitro immunostimulating activity of sulfated polysaccharides from Caulerpa cupressoides var Flabellata. Mar. Drugs 2019, 17, 105. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.S.S.P.; Costa, L.S.; Cordeiro, S.L.; Almeida-Lima, J.; Dantas-Santos, N.; Magalhães, K.D.; Sabry, D.A.; Albuquerque, I.R.L.; Pereira, M.R.; Leite, E.L.; Rocha, H.A.O. Evaluating the possible anticoagulant and antioxidant effects of sulfated polysaccharides from the tropical green alga Caulerpa cupressoides var flabellate. J. Appl. Phycol. 2012, 24, 1159–1167. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Effect of season on the composition of bioactive polysaccharides from the brown seaweed Saccharina longicruris. Phytochemistry 2009, 70, 1069–1075. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Bourret, E. Effects of season on the yield and quality of agar from Gracilaria species (Gracilariaceae, Rhodophyta). Bioresour. Technol. 2003, 90, 329–333. [Google Scholar] [CrossRef]
- Siddhanta, A.K.; Goswami, A.M.; Ramavat, B.K.; Mody, K.H.; Mairh, O.P. Water soluble polysaccharides of marine algal species of Ulva (Ulvales, Chlorophyta) of Indian waters. Indian J. Mar. Sci. 2001, 30, 166–172. [Google Scholar]
- Grünewald, N.; Growth, I.; Alban, S. Evaluation of seasonal variations of the structure and anti–inflammatory activity of sulfated polysaccharides extracted from the red alga Delesseria sanguinea (Hudson) Lamouroux (Ceramiales, Delesseriaceae). Biomacromolecules. 2009, 10, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Wesson, J.A.; Worcester, E.M.; Kleinman, J.G. Role of anionic proteins in kidney stone formation: Interaction between model anionic polypeptides and calcium oxalate crystals. J. Urol. 2000, 163, 1343–1348. [Google Scholar] [CrossRef]
- He, J.Y.; Deng, S.P.; Ouyang, J.M. Morphology, particle size distribution, aggregation, and crystal phase of nanocrystallites in the urine of healthy persons and lithogenic patients. IEEE Trans. Nanobiosci. 2010, 9, 156–163. [Google Scholar] [CrossRef]
- Sheng, X.; Jung, T.; Wesson, J.A.; Ward, M.D. Adhesion at calcium oxalate crystal surfaces and the effect of urinary constituents. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 267–272. [Google Scholar] [CrossRef]
- Thomas, A.; Rosseeva, E.; Hochrein, O.; Carrillo-Cabrera, W.; Simon, P.; Duchstein, P.; Zahn, D.; Kniep, R. Mimicking the growth of a pathologic biomineral: Shape development and structures of calcium oxalate dihydrate in the presence of polyacrylic acid. Chem. Eur. J. 2012, 18, 4000–4009. [Google Scholar] [CrossRef]
- Duan, C.Y.; Xia, Z.Y.; Zhang, G.N.; Gui, B.S.; Xue, J.F.; Ouyang, J.M. Changes in urinary nanocrystallites in calcium oxalate stone formers before and after potassium citrate intake. Int. J. Nanomed. 2013, 8, 909–918. [Google Scholar] [CrossRef]
- Peng, H.; Ouyang, J.M.; Yao, X.Q.; Yang, R.E. Interaction between submicron COD crystals and renal epithelial cells. Int. J. Nanomed. 2012, 7, 4727–4737. [Google Scholar] [CrossRef][Green Version]
- Sun, X.Y.; Ouyang, J.M.; Liu, A.J.; Ding, Y.M.; Gan, Q.Z. Preparation, characterization, and in vitro cytotoxicity of COM and COD crystals with various sizes. Mater. Sci. Eng. C 2015, 57, 147–156. [Google Scholar] [CrossRef]
- Silva, T.M.A.; Alves, L.G.; Queiroz, K.C.S.; Santos, M.G.L.; Marques, C.T.; Chavante, S.F.; Rocha, H.A.O.; Leite, E.L. Partial characterization and anticoagulant activity of a heterofucan from the brown seaweed Padina gymnospora. Brazilian J. Med. Biol. Res. 2005, 38, 523–533. [Google Scholar] [CrossRef]
- Cao, Y.; Ikeda, I. Antioxidant activity and antitumor activity (in vitro) of xyloglucan selenious ester and surfated xyloglucan. Int. J. Biol. Macromol. 2009, 45, 231–235. [Google Scholar] [CrossRef]
- Grohe, B.; O’Young, J.; Ionescu, D.A.; Lajoie, G.; Rogers, K.A.; Karttunen, M.; Goldberg, H.A.; Hunter, G.K. Control of calcium oxalate crystal growth by face–specific adsorption of an osteopontin phosphopeptide. J. Am. Chem. Soc. 2007, 129, 14946–14951. [Google Scholar] [CrossRef]
- Chien, Y.C.; Masica, D.L.; Gray, J.J.; Nguyen, S.; Vali, H.; McKee, M.D. Modulation of calcium oxalate dihydrate growth by selective crystal–face binding of phosphorylated osteopontin and polyaspartate peptide showing occlusion by sectoral (compositional) zoning. J. Biol. Chem. 2009, 35, 23491–23501. [Google Scholar] [CrossRef]
- Hernandez, S.E.R.; de Leeuw, N.H. Effect of Chondroitin 4-Sulfate on the growth and morphology of calcium oxalate monohydrate: a molecular dynamics study. Cryst. Growth Des. 2015, 15, 4438–4447. [Google Scholar] [CrossRef]
- Qiu, S.R.; Wierzbicki, A.; Orme, C.A.; Cody, A.M.; Hoyer, J.R.; Nancollas, G.H.; Zepeda, S.; De Yoreo, J.J. Molecular modulation of calcium oxalate crystallization by osteopontin and citrate. Proc. Natl. Acad. Sci. USA 2004, 101, 1811–1815. [Google Scholar] [CrossRef]
- Escobar, C.; Neira-Carrillo, A.; Fernández, M.S.; Arias, J.L. Biomineralization: From Paleontology to Materials Science; Role of Sulfated Macromolecules in Urinary Stone Formation; Editorial Universitaria: Santiago, Chile, 2007; pp. 343–358. [Google Scholar]
- Wynne, M.J. A checklist of benthic marine algae of the tropical and subtropical western Atlantic. Can. J. Bot. 1986, 64, 2239–2281. [Google Scholar] [CrossRef]
- Dodgson, K.; Price, R. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Spector, T. Refinement of the Coomassie blue method of protein quantitation. A simple and linear spectrophotometric assay for ≤0.5 to 50 μg of protein. Anal. Biochem. 1978, 86, 142–146. [Google Scholar] [CrossRef]
- Mendes-Marques, M.L.; Presa, F.B.; Viana, R.L.S.; Costa, M.S.S.P.; Amorim, M.O.R.; Bellan, D.L.; Alves, M.G.C.F.; Costa, L.S.; Trindade, E.S.; Rocha, H.A.O. Anti-thrombin, anti-adhesive, anti-migratory, and anti-proliferative activities of sulfated galactans from the tropical green seaweed, Udotea flabellum. Mar. Drugs 2019, 17, 5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharide | (SO4)/Total Sugar (%/%) | Protein (%) |
|---|---|---|
| CCB-F0.3 | 1.10 ± 0.02 a | 0.06 ± 0.01 a |
| CCB-F0.5 | 0.86 ± 0.03 b | 0.05 ± 0.01 b |
| CCB-F1.0 | 0.85 ± 0.02 b | 0.11 ± 0.02 c |
| CCB-F2.0 | 0.72 ± 0.01 c | 0.11 ± 0.01 c |
| Total Amount of Crystals (units) | COM (units) | COM Size (µm) | COD (units) | COD Size (µm) | |
|---|---|---|---|---|---|
| Control CaOx | 21 ± 4.4 a | 13 ± 4.1 a | 11.9 ± 0.16 a | 7 ± 2.0 a | 12.8 ± 0.89 a |
| CaOx + CCB-F0.3 | 257 ± 4.2 b | 48 ± 10.9 b | 1.7 ± 0.08 b | 209 ± 12.3 b | 2.55 ± 0.19 b |
| CaOx + CCB-F0.5 | 77 ± 8.3 c | 20 ± 6.9 a | 4.2 ± 0.64 c | 58 ± 9.8 c | 4.9 ± 0.12 c |
| CaOx + CCB-F1.0 | 184 ± 10.8 d | 66 ± 9.1 b | 4.4 ± 0.29 c | 118 ± 9.8 d | 4.7 ± 0.79 c,d |
| CaOx + CCB-F2.0 | 32 ± 9.4 a | 23 ± 7.3 a | 4.8 ± 0.96 c | 9 ± 3.3 a | 6.2 ± 0.83 d |
| ζ (mV) | |
|---|---|
| Control CaOx | +8.85 ± 3.30 a |
| CaOx + CCB-F0.3 | −25.82 ± 6.36 b |
| CaOx+ CCB-F0.5 | −43.87 ± 8.63 c |
| CaOx + CCB-F1.0 | −51.50 ± 2.14 c |
| CaOx + CCB-F2.0 | −68.70 ± 12.01d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, D.L.; Melo, K.R.T.; Queiroz, M.F.; Batista, L.A.N.C.; Santos, P.C.; Costa, M.S.S.P.; Almeida-Lima, J.; Camara, R.B.G.; Costa, L.S.; Rocha, H.A.O. In Vitro Studies Reveal Antiurolithic Effect of Antioxidant Sulfated Polysaccharides from the Green Seaweed Caulerpa cupressoides var flabellata. Mar. Drugs 2019, 17, 326. https://doi.org/10.3390/md17060326
Gomes DL, Melo KRT, Queiroz MF, Batista LANC, Santos PC, Costa MSSP, Almeida-Lima J, Camara RBG, Costa LS, Rocha HAO. In Vitro Studies Reveal Antiurolithic Effect of Antioxidant Sulfated Polysaccharides from the Green Seaweed Caulerpa cupressoides var flabellata. Marine Drugs. 2019; 17(6):326. https://doi.org/10.3390/md17060326
Chicago/Turabian StyleGomes, Dayanne Lopes, Karoline Rachel Teodosio Melo, Moacir Fernandes Queiroz, Lucas Alighieri Neves Costa Batista, Pablo Castro Santos, Mariana Santana Santos Pereira Costa, Jailma Almeida-Lima, Rafael Barros Gomes Camara, Leandro Silva Costa, and Hugo Alexandre Oliveira Rocha. 2019. "In Vitro Studies Reveal Antiurolithic Effect of Antioxidant Sulfated Polysaccharides from the Green Seaweed Caulerpa cupressoides var flabellata" Marine Drugs 17, no. 6: 326. https://doi.org/10.3390/md17060326
APA StyleGomes, D. L., Melo, K. R. T., Queiroz, M. F., Batista, L. A. N. C., Santos, P. C., Costa, M. S. S. P., Almeida-Lima, J., Camara, R. B. G., Costa, L. S., & Rocha, H. A. O. (2019). In Vitro Studies Reveal Antiurolithic Effect of Antioxidant Sulfated Polysaccharides from the Green Seaweed Caulerpa cupressoides var flabellata. Marine Drugs, 17(6), 326. https://doi.org/10.3390/md17060326