The Biosynthesis of Rare Homo-Amino Acid Containing Variants of Microcystin by a Benthic Cyanobacterium

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
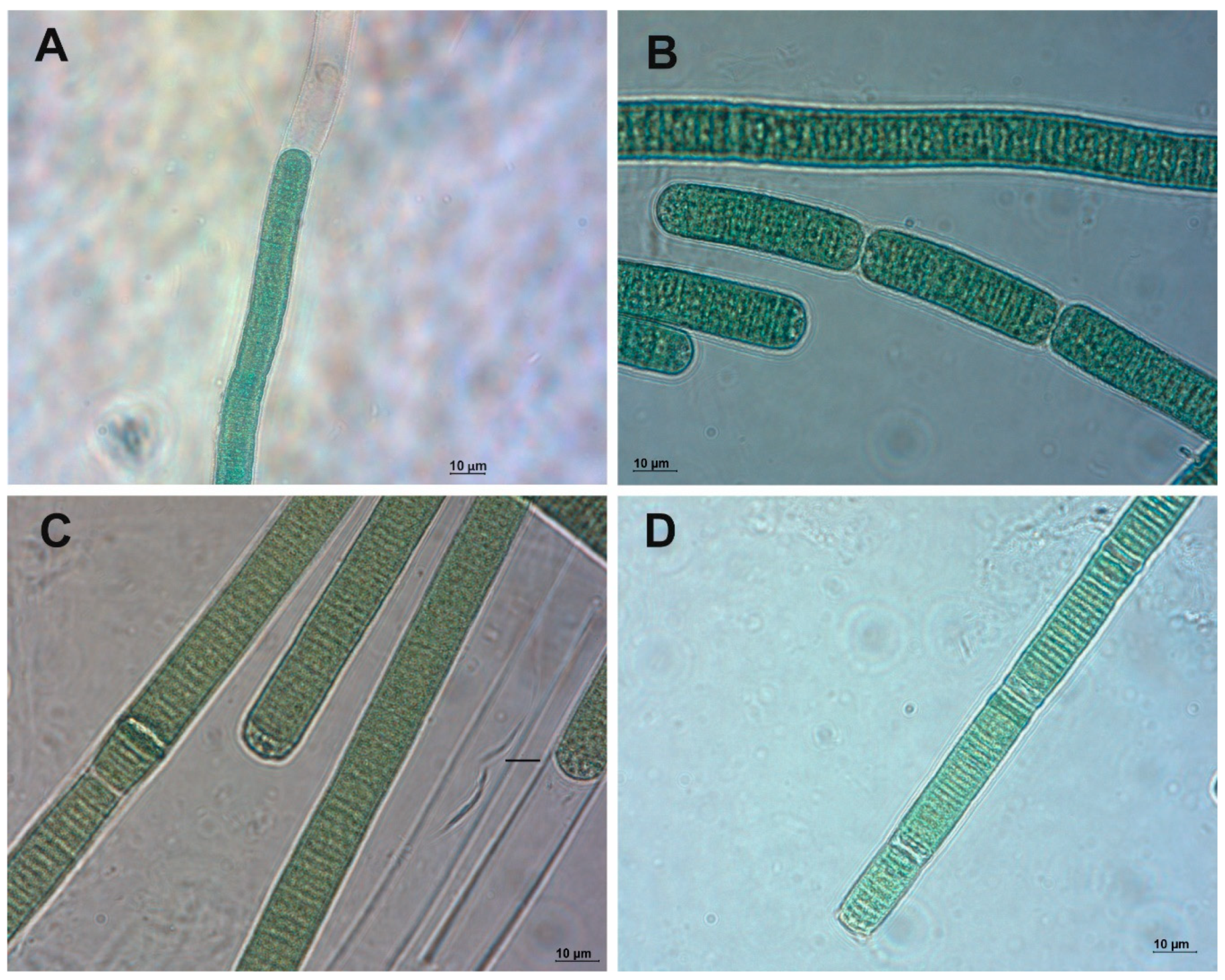
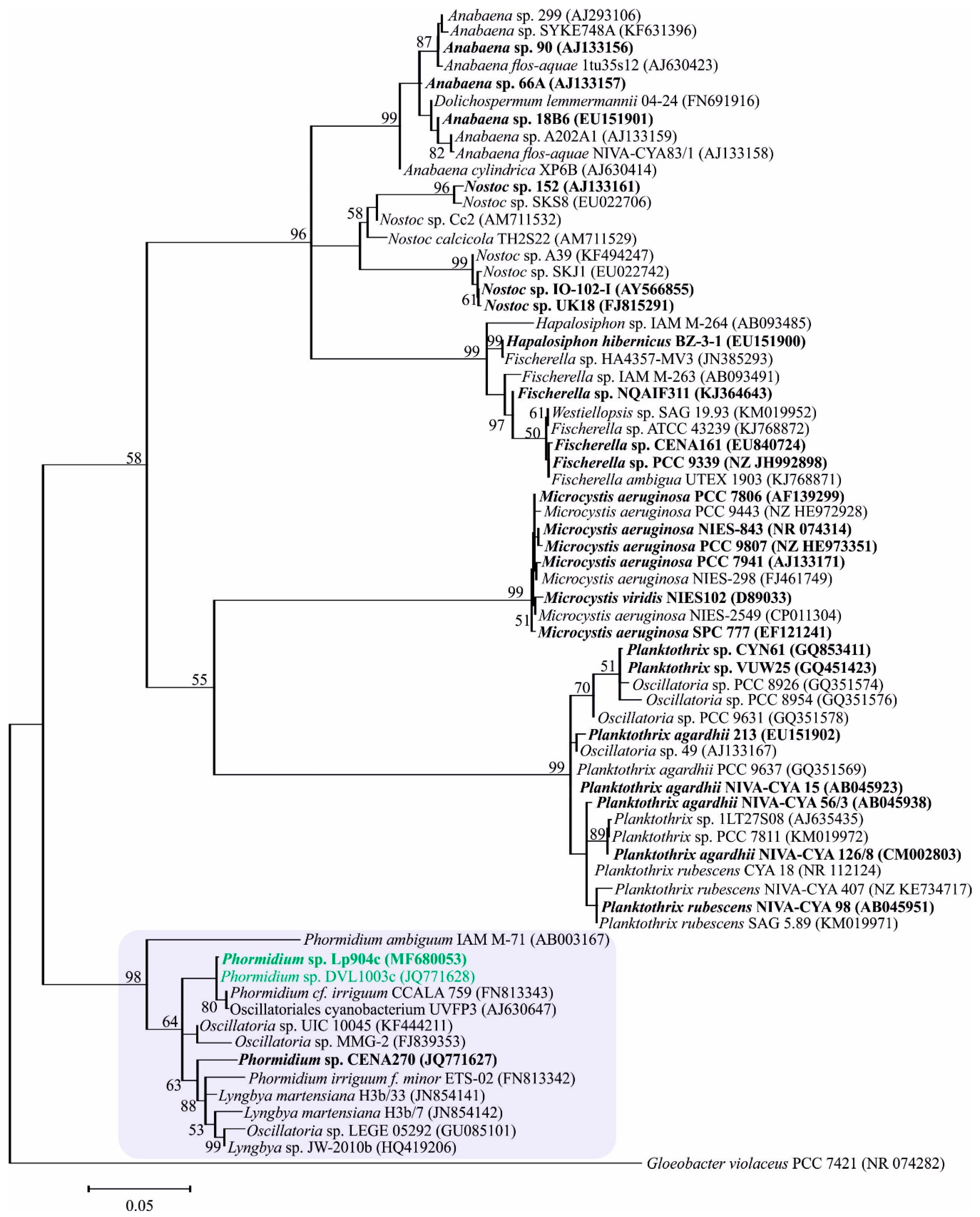
2.1. Phormidium Strains Producing Unusual Microcystins (MCs)
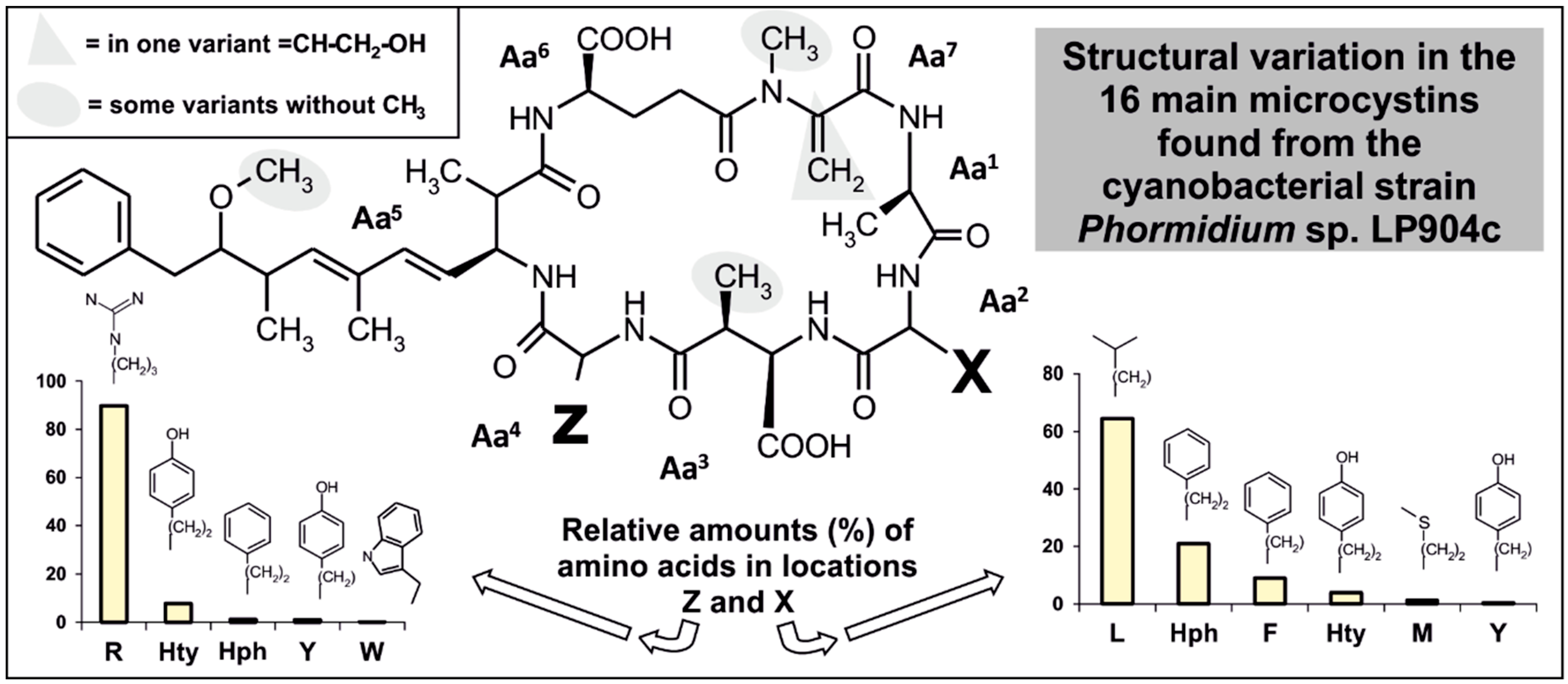
2.2. Microcystin and Homophenylalanine Biosynthetic Genes from Phormidium
2.3. McyB1 and McyC from Phormidium sp. LP904c
3. Discussion
3.1. Benthic Cyanobacteria Producing Unusual Microcystins
3.2. Microcystins Containing Homo-Amino Acids
3.3. Microcystin Gene Cluster from Phormidium
3.4. Lack of Microcystin from Phormidium sp. DVL1003c
3.5. Diversity of Benthic Phormidium Strains
4. Materials and Methods
4.1. Cultivation of Phormidium Strains
4.2. Chemical Analysis
4.3. DNA Extraction, PCR and Genome Sequencing and Assembly
4.4. Bioinformatics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K. Cyanobacterial toxins. In Encyclopedia of Microbiology; Moselio, S., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Meriluoto, J.; Spoof, L.; Codd, G. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; ISBN 978-1-119-06868-6. [Google Scholar]
- Tillett, D.; Dittmann, E.; Erhard, M.; Von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Christiansen, G.; Fastner, J.; Erhard, M.; Börner, T.; Dittmann, E. Microcystin biosynthesis in Planktothrix: Genes, Evolution, and Manipulation. J. Bacteriol. 2003, 185, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes coding for hepatotoxic heptapeptides (microcystins) in the cyanobacterium Anabaena strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef]
- Fewer, D.P.; Wahlsten, M.; Österholm, J.; Jokela, J.; Rouhiainen, L.; Kaasalainen, U.; Rikkinen, J.; Sivonen, K. The genetic basis for O-acetylation of the microcystin toxin in cyanobacteria. Chem. Biol. 2013, 20, 861–869. [Google Scholar] [CrossRef]
- Welker, M.; von Döhren, H. Cyanobacterial peptides-nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Laakso, K.; Wang, H.; Sivonen, K. Recurrent adenylation domain replacement in the microcystin synthetase gene cluster. BMC Evol. Biol. 2007, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Fewer, D.P.; Rohrlack, T.; Jokela, J.; Rouhiainen, L.; Sivonen, K.; Kristensen, T.; Jakobsen, K.S. Evidence for positive selection acting on microcystin synthetase adenylation domains in three cyanobacterial genera. BMC Evol. Biol. 2008, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Alexova, R.; Haynes, P.A.; Ferrari, B.C.; Neilan, B.A. Comparative protein expression in different strains of the bloom-forming cyanobacterium Microcystis aeruginosa. Mol. Cell. Proteom. 2011, 10, M110.003749. [Google Scholar] [CrossRef]
- Shishido, T.K.; Kaasalainen, U.; Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Fiore, M.F.; Yunes, J.S.; Rikkinen, J.; Sivonen, K. Convergent evolution of [D-Leucine(1)] microcystin-LR in taxonomically disparate cyanobacteria. BMC Evol. Biol. 2013, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Koketsu, K.; Mitsuhashi, S.; Tabata, K. Identification of homophenylalanine biosynthetic genes from the cyanobacterium Nostoc punctiforme PCC73102 and application to its microbial production by Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 2201–2208. [Google Scholar] [CrossRef]
- Rouhiainen, L.; Jokela, J.; Fewer, D.P.; Urmann, M.; Sivonen, K. Two alternative starter modules for the non-ribosomal biosynthesis of specific anabaenopeptin variants in Anabaena (Cyanobacteria). Chem. Biol. 2010, 17, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Shishido, T.K.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Fiore, M.F.; Sivonen, K. Simultaneous production of anabaenopeptins and namalides by the cyanobacterium Nostoc sp. CENA543. ACS Chem. Biol. 2017, 12, 2746–2755. [Google Scholar] [CrossRef]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier/Spektrum Akademischer Verlag: Heidelberg, Germany, 2005; pp. 390–391. [Google Scholar]
- Silva, C.S.; Genuário, D.B.; Vaz, M.G.; Fiore, M.F. Phylogeny of culturable cyanobacteria from Brazilian mangroves. Syst. Appl. Microbiol. 2014, 37, 100–112. [Google Scholar] [CrossRef]
- Andreote, A.P.; Vaz, M.G.; Genuário, D.B.; Barbiero, L.; Rezende-Filho, A.T.; Fiore, M.F. Nonheterocytous cyanobacteria from Brazilian saline-alkaline lakes. J. Phycol. 2014, 50, 675–684. [Google Scholar] [CrossRef]
- Bravakos, P.; Kotoulas, G.; Skaraki, K.; Pantazidou, A.; Economou-Amilli, A. A polyphasic taxonomic approach in isolated strains of Cyanobacteria from thermal springs of Greece. Mol. Phylogenet. Evol. 2016, 98, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Khattar, J.; Singh, D.P.; Rahi, P.; Gulati, A. Limnology and cyanobacterial diversity of high altitude lakes of Lahaul-Spiti in Himachal Pradesh, India. J. Biosci. 2014, 39, 643–657. [Google Scholar] [CrossRef]
- Hawes, I.; Sumner, D.Y.; Andersen, D.T.; Mackey, T.J. Legacies of recent environmental change in the benthic communities of Lake Joyce, a perennially ice-covered Antarctic lake. Geobiology 2011, 9, 394–410. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; El-Sharouny, H.M.; Ali, W.S.M. Microcystin production in benthic mats of cyanobacteria in the Nile River and irrigation canals, Egypt. Toxicon 2006, 47, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, G.; Jungblut, A.D.; Neilan, B.A. Benthic cyanobacteria (Oscillatoriaceae) that produce microcystin-LR, isolated from four reservoirs in southern California. Water Res. 2007, 41, 492–498. [Google Scholar] [CrossRef]
- Cantoral Uriza, E.A.; Asencio, A.D.; Aboal, M. Are we underestimating benthic cyanotoxins? Extensive sampling results from Spain. Toxins 2017, 9, 385. [Google Scholar] [CrossRef]
- Wood, S.A.; Heath, M.W.; Holland, P.T.; Munday, R.; McGregor, G.B.; Ryan, K.G. Identification of a benthic microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, T.; Kato, H.; Imanishi, S.; Kawasaki, Y.; Hasegawa, M.; Harada, K. Structural characterization of microcystins by LC/MS/MS under Ion Trap conditions. J. Antibiot. 2006, 59, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Namikoshi, M.; Otsuki, A.; Sivonen, K. Effect of amino acid side-chain on fragmentation of cyclic peptide ions: Differences of electrospray ionization collision-induced decomposition mass spectra of toxic heptapeptide microcystins containing ADMAdda instead of Adda. Eur. Mass. Spectrom 1998, 4, 287–298. [Google Scholar] [CrossRef]
- Diehnelt, C.W.; Dugan, N.R.; Peterman, S.M.; Budde, W.L. Identification of microcystin toxins from a strain of Microcystis aeruginosa by Liquid Chromatography Introduction into a hybrid linear Ion Trap-Fourier Transform Ion Cyclotron Resonance Mass Spectrometer. Anal. Chem. 2006, 78, 501–512. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast calculation of molecular polar surface area as a sum of fragment-based contributions and its application to the prediction of drug transport properties. J. Med. Chem. 2000, 43, 3714–3717. [Google Scholar] [CrossRef]
- Stachelhaus, T.; Mootz, H.D.; Marahiel, M.A. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 1999, 6, 493–505. [Google Scholar] [CrossRef]
- Rausch, C.; Weber, T.; Kohlbacher, O.; Wohlleben, W.; Hudson, D.H. Specificity prediction of adenylation domains in nonribosomal peptide synthetases (NRPS) using transductive support vector machines (TSVMs). Nucleic Acids Res. 2005, 33, 5799–5808. [Google Scholar] [CrossRef]
- Röttig, M.; Medema, M.H.; Bin, K.; Weber, T.; Rausch, C.; Kohbacher, O. NRPSpredictor2—A web server for predicting NRPS adenylation domain specificity. Nucleic Acids Res. 2011, 39, W362–W367. [Google Scholar] [CrossRef]
- Fiore, M.F.; Genuário, D.B.; da Silva, C.S.; Shishido, T.K.; Moraes, L.A.; Cantúsio Neto, R.; Silva-Stenico, M.E. Microcystin production by a freshwater spring cyanobacterium of the genus Fischerella. Toxicon 2009, 53, 754–761. [Google Scholar] [CrossRef]
- Cirés, S.; Alvarez-Roa, C.; Wood, S.A.; Puddick, J.; Loza, V.; Heimann, K. First report of microcystin-producing Fischerella sp. (Stigonematales, Cyanobacteria) in tropical Australia. Toxicon 2014, 88, 62–66. [Google Scholar] [CrossRef]
- Gaget, V.; Humpage, A.R.; Huang, Q.; Monis, P.; Brookes, J.D. Benthic cyanobacteria: A source of cylindrospermopsin and microcystin in Australian drinking water reservoirs. Water Res. 2017, 124, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Mez, K.; Beattie, K.; Codd, G.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Aboal, M.; Puig, M.A. Intracellular and dissolved microcystin in reservoirs of the river Segura basin, Murcia, SE Spain. Toxicon 2005, 45, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, I.; Aboal, M.; Zafra, E.; Campillo, D. Significance of microcystin production by benthic communities in water treatment systems of arid zones. Water Res. 2008, 42, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Dasey, M.; Ryan, N.; Wilson, J.; McGregor, G.; Fabbro, L.; Neilan, B.A.; Burns, B.P.; Kankaanpää, H.; Morrison, L.F.; Codd, G.A.; et al. Investigations into the taxonomy, toxicity and ecology of benthic cyanobacterial accumulations in Myall Lake, Australia. Mar. Freshw. Res. 2005, 56, 45–55. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Lampert, C.S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D.R. Toxin production in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Küpper, F.C.; Dietrich, D. Potent toxins in Arctic environments—Presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem. Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef]
- Mehner, C.; Müller, D.; Krick, A.; Kehraus, S.; Löser, R.; Gütschow, M.; Maier, A.; Fiebig, H.-H.; Brun, R.; König, G.M. A novel beta-amino acid in cytotoxic peptides from the cyanobacterium Tychonema sp. Eur. J. Org. Chem. 2008, 2008, 1732–1739. [Google Scholar] [CrossRef]
- An, T.; Kumar, T.K.; Wang, M.; Liu, L.; Lay, J.O., Jr.; Liyanage, R.; Berry, J.; Gantar, M.; Marks, V.; Gawley, R.E.; et al. Structures of pahayokolides A and B, cyclic peptides from a Lyngbya sp. J. Nat. Prod. 2007, 70, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Structures of three new homotyrosine-containing microcystins and a new homophenylalanine variant from Anabaena sp. strain 66. Chem. Res. Toxicol. 1992, 5, 661–666. [Google Scholar] [CrossRef]
- Fewer, D.P.; Jokela, J.; Rouhiainen, L.; Wahlsten, M.; Koskenniemi, K.; Stal, L.J.; Sivonen, K. The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 2009, 73, 924–937. [Google Scholar] [CrossRef] [PubMed]
- Zainuddin, E.N.; Jansen, R.; Nimtz, M.; Wray, V.; Preisitsch, M.; Lalk, M.; Mundt, S. Lyngbyazothrins A–D, antimicrobial cyclic undecapeptides from the cultured cyanobacterium Lyngbya sp. J. Nat. Prod. 2009, 72, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Pereira, A.R.; Liu, W.T.; Ng, J.; Pevzner, P.A.; Dorrestein, P.C.; König, G.M.; Vasconcelos, V.M.; Gerwick, W.H. Synergistic allelochemicals from a freshwater cyanobacterium. Proc. Natl. Acad. Sci. USA 2010, 107, 11183–11188. [Google Scholar] [CrossRef] [PubMed]
- Heck, K.; Alvarenga, D.O.; Shishido, T.K.; Varani, A.M.; Dörr, F.A.; Pinto, E.; Rouhiainen, L.; Jokela, J.; Sivonen, K.; Fiore, M.F. Biosynthesis of microcystin hepatotoxins in the cyanobacterial genus Fischerella. Toxicon 2018, 141, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Stachelhaus, T.; Marahiel, M.A.; Brick, P. Structural basis for the activation of phenylalanine in the non-ribosomal biosynthesis of gramicidin S. EMBO J. 1997, 16, 4174–4183. [Google Scholar] [CrossRef]
- Challis, G.L.; Ravel, J.; Townsend, C.A. Predictive, structure-based model of amino acid recognition by nonribosomal peptide synthetase adenylation domains. Chem. Biol. 2000, 7, 211–224. [Google Scholar] [CrossRef]
- Mikalsen, B.; Boison, G.; Skulberg, O.M.; Fastner, J.; Davies, W.; Gabrielsen, T.M.; Rudi, K.; Jacobsen, K.S. Natural variation in the microcystin synthetase operon mcyABC and impact on microcystin production in Microcystis strains. J. Bacteriol. 2003, 185, 2774–2785. [Google Scholar] [CrossRef]
- Tanabe, Y.; Kaya, K.T.; Watanabe, M.M. Evidence for recombination in the microcystin synthetase (mcy) genes of toxic cyanobacteria Microcystis spp. J. Mol. Evol. 2004, 58, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Gumpenberger, M.; Fastner, J. Genetic identification of microcystin ecotypes in toxic cyanobacteria of the genus Planktothrix. Microbiology 2005, 151, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Gumpenberger, M. Diversity of microcystin genotypes among populations of the filamentous cyanobacteria Planktothrix rubescens and Planktothrix agardhii. Mol. Ecol. 2006, 15, 3849–3861. [Google Scholar] [CrossRef]
- Tooming-Klunderud, A.; Mikalsen, B.; Kristensen, T.; Jakobsen, K.S. The mosaic structure of the mcyABC operon in Microcystis. Microbiology 2008b, 154, 1886–1899. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, G.; Yoshida, W.Y.; Blom, J.F.; Portmann, C.; Gademann, K.; Hemscheidt, T.; Kurmayer, R. Isolation and structure determination of two microcystins and sequence comparison of the McyABC adenylation domains in Planktothrix species. J. Nat. Prod. 2008, 71, 1881–1886. [Google Scholar] [CrossRef]
- Hoffmann, D.; Hevel, J.M.; Moore, R.E.; Moore, B.S. Sequence analysis and biochemical characterization of the nostopeptolide A biosynthetic gene cluster from Nostoc sp. GSV224. Gene 2003, 311, 171–180. [Google Scholar] [CrossRef]
- Rounge, T.B.; Rohrlack, T.; Kristensen, T.; Jakobsen, K.S. Recombination and selectional forces in cyanopeptolin NRPS operons from highly similar, but geographically remote Planktothrix strains. BMC Microbiol. 2008, 8, 141. [Google Scholar] [CrossRef]
- Meyer, S.; Kehr, J.C.; Mainz, A.; Dehm, D.; Petras, D.; Süssmuth, R.D.; Dittmann, E. Biochemical dissection of the natural diversification of microcystin provides lessons for synthetic biology of NRPS. Cell. Chem. Biol. 2016, 23, 462–471. [Google Scholar] [CrossRef]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasärkkä, K.; Li, Z.; et al. Genome-derived insights into the biology of the hepatotoxic bloom-forming cyanobacterium Anabaena sp. strain 90. BMC Genom. 2012, 13, 613. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bachmann, B.O.; Ravel, J. In silico prediction of microbial secondary metabolic pathways from DNA sequence data. Methods Enzymol. 2009, 458, 181–217. [Google Scholar] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Microcystin | Aa in Position | Rt | [M + H]+ (m/z) | Error | RA (%) | |||
|---|---|---|---|---|---|---|---|---|---|
| X | Z | (min) | Calculated | Measured | (ppm) | LP904c | DVL1003c | ||
| 1 | MC-YR | Y | R | 15.4 | 1045.5353 | 1045.5329 | –2.35 | <1 | <1 |
| 2 | MC-HtyR | Hty | R | 15.8 | 1059.5510 | 1059.5510 | –0.01 | 4 | 4 |
| 3 | MC-MR | M | R | 15.8 | 1013.5125 | 1013.5148 | 2.26 | 1 | 1 |
| 4 | [DMAdda5]MC-LR | L | R | 15.8 | 981.5404 | 981.5404 | –0.04 | <1 | <1 |
| 5 | [L-MeSer7]MC-LR | L | R | 16.6 | 1013.5666 | 1013.5649 | –1.73 | <1 | <1 |
| 6 | [D-Asp3]MC-LR | L | R | 17 | 981.5404 | 981.5404 | –0.04 | <1 | <1 |
| 7 | MC-LR | L | R | 17 | 995.5560 | 995.5566 | 0.51 | 55 | 49 |
| 8 | MC-FR | F | R | 17.3 | 1029.5404 | 1029.5401 | –0.33 | 9 | 7 |
| 9 | [Dha7]MC-LR | L | R | 17.5 | 981.5404 | 981.5404 | –0.04 | <1 | <1 |
| 10 | MC-HphR | Hph | R | 18.7 | 1043.5560 | 1043.5561 | 0.01 | 17 | 14 |
| 11 | MC-LY | L | Y | 22.8 | 1002.5183 | 1002.5178 | –0.51 | 1 | 2 |
| 12 | MC-LHty | L | Hty | 23.2 | 1016.5339 | 1016.5347 | 0.73 | 5 | 12 |
| 13 | MC-HphHty | Hph | Hty | 24.3 | 1064.5339 | 1064.5337 | –0.24 | 3 | 8 |
| 14 | MC-LW | L | W | 24.6 | 1025.5342 | 1025.5343 | 0.00 | <1 | <1 |
| 15 | MC-LHph | L | Hph | 26.7 | 1000.5390 | 1000.5369 | –2.15 | <1 | 1 |
| 16 | MC-HphHph | Hph | Hph | 27.7 | 1048.5390 | 1048.5403 | 1.20 | <1 | <1 |
| Organism | Strain | McyB1 | P * | % | Aa # | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 235 | 236 | 239 | 278 | 299 | 301 | 322 | 330 | 331 | 517 | |||||
| Phormidium sp. | LP904c | D | I | C | V | F | G | L | V | H | K | Ser-Thr | 60 | Leu, Hph, Phe, Hty, Tyr, Arg |
| Phormidium sp. | DVL1003c | D | I | C | V | F | G | L | V | H | K | Ser-Thr | 60 | Leu, Hph, Phe, Hty, Tyr, Arg |
| Planktothrix agardhii | CYA 126/8 | D | A | L | F | F | G | L | V | D | K | Leu | 100 | Arg, Leu |
| Planktothrix agardhii | 213 | D | A | L | F | F | G | L | V | D | K | Leu | 100 | Arg, Leu |
| Planktothrix agardhii | NIVA-CYA56/3 | D | A | L | F | F | G | L | V | D | K | Leu | 100 | Leu, Arg, Tyr |
| Planktothrix prolifica | NIVA-CYA 98 | D | A | L | F | F | G | L | V | D | K | Leu | 100 | Leu, Arg |
| Planktothrix rubescens | NIVA-CYA 407 | D | A | L | L | F | G | L | V | D | K | Leu | 90 | Leu, Arg, Hty |
| Dolichospermum sp. ∞ | 90 | D | V | W | F | F | G | L | V | D | K | Ser | 80 | Leu, Arg, Hil |
| Dolichospermum flos-aquae∞ | 18B6 | D | V | W | S | F | G | L | V | D | K | Ser | 80 | Arg, X |
| Dolichospermum lemmermannii∞ | 66 A | D | V | W | S | F | G | L | V | Y | K | Ser | 70 | Hty, X, Hph, Leu, Tyr, Phe |
| Nostoc sp. | 152 | D | A | L | F | F | G | L | I | Y | K | Leu | 80 | Leu, Hil, X, Val |
| Nostoc sp. | IO-102-I | D | I | K | N | F | G | A | I | V | K | Gln | 50 | Leu, X, Phe, Hil, Tyr |
| Fischerella sp. | PCC9339 | D | V | L | I | F | G | L | I | Y | K | Pro | 70 | Leu |
| Hapalosiphon hibernicus | BZ-3-1 | D | V | W | F | F | G | L | V | D | K | Ser | 80 | Leu, Arg |
| Microcystis aeruginosa | PCC 7806 | D | A | W | F | L | G | N | V | V | K | Leu | 100 | Val |
| Microcystis aeruginosa | FCY-28 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Leu |
| Microcystis aeruginosa | FCY-26 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Ni |
| Microcystis aeruginosa | UV027 | D | V | W | T | I | G | A | V | E | K | Arg | 100 | Arg |
| Microcystis aeruginosa | K139 | D | A | W | F | L | G | N | V | V | K | Leu | 100 | Leu |
| Microcystis aeruginosa | DIANCHI905 | D | A | W | F | L | G | N | V | V | K | Leu | 100 | Ni |
| Microcystis aeruginosa | PCC 9807 | D | A | W | F | L | G | N | V | V | K | Leu | 100 | Ni |
| Microcystis aeruginosa | PCC 7941 | D | A | W | F | L | G | N | V | V | K | Leu | 100 | Ni |
| Microcystis aeruginosa | PCC 9443 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Ni |
| Microcystis aeruginosa | NIES-843 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Arg, Leu, Tyr |
| Microcystis aeruginosa | SPC777 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Arg |
| Microcystis viridis | NIES 102 | D | G | W | T | I | G | A | V | E | K | Arg | 90 | Hil, Leu, Arg, Tyr, Trp, Phe, Hty, X |
| Organism | Strain | McyC | P * | % | Aa # | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 235 | 236 | 239 | 278 | 299 | 301 | 322 | 330 | 331 | 517 | |||||
| Phormidium sp. | LP904c | D | A | L | F | F | G | L | V | D | K | Leu | 100 | Arg, Hty, Hph, Trp |
| Phormidium sp. | DVL1003c | D | A | L | C | F | G | L | V | D | K | Leu | 100 | Arg, Hty, Hph, Trp |
| Planktothrix agardhii | CYA 126/8 | D | P | W | G | F | G | L | V | D | K | Gln | 70 | Arg |
| Planktothrix agardhii | 213 | D | P | W | C | F | G | L | V | D | K | Gln | 70 | Arg |
| Planktothrix agardhii | NIVA-CYA 56/3 | D | P | W | G | F | G | L | V | D | K | Gln | 70 | Arg |
| Planktothrix prolifica | NIVA-CYA 98 | D | P | W | G | F | G | L | V | D | K | Gln | 70 | Arg |
| Planktothrix rubescens | NIVA-CYA 407 | D | P | W | G | F | G | L | V | D | K | Gln | 70 | Arg |
| Dolichospermum sp. ∞ | 90 | D | V | W | C | F | G | L | V | D | K | Ser | 80 | Arg |
| Dolichospermum flos-aquae∞ | 18B6 | D | V | W | S | F | G | L | V | D | K | Ser | 80 | Arg |
| Dolichospermum lemmermannii∞ | 66 A | D | V | W | S | F | G | L | V | D | K | Ser | 80 | Arg |
| Nostoc sp. | 152 | D | V | W | N | F | G | F | I | D | K | Gln | 70 | Arg, Har |
| Nostoc sp. | IO-102-I | D | V | W | N | F | G | F | V | D | K | Glu | 70 | Arg |
| Fischerella sp. | PCC9339 | D | V | W | F | F | G | L | V | D | - | Ser | 70 | Arg |
| Hapalosiphon hibernicus | BZ-3-1 | D | V | W | F | F | G | L | V | D | K | Ser | 80 | Ala, Leu, Val |
| Microcystis aeruginosa | PCC 7806 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Arg |
| Microcystis aeruginosa | FCY-28 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Ni |
| Microcystis aeruginosa | FCY-26 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Ni |
| Microcystis aeruginosa | UV027 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Arg |
| Microcystis aeruginosa | K139 | D | V | W | T | I | G | A | V | E | K | Arg | 100 | Arg |
| Microcystis aeruginosa | DIANCHI905 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Ni |
| Microcystis aeruginosa | PCC 9807 | D | V | W | T | I | G | I | V | D | K | Arg | 90 | Ni |
| Microcystis aeruginosa | PCC 7941 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Ni |
| Microcystis aeruginosa | PCC 9443 | D | V | W | T | I | G | I | V | D | K | Arg | 90 | Ni |
| Microcystis aeruginosa | NIES-843 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Arg |
| Microcystis aeruginosa | SPC777 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Arg |
| Microcystis viridis | NIES 102 | D | V | W | T | I | G | A | V | D | K | Arg | 100 | Arg |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shishido, T.K.; Jokela, J.; Humisto, A.; Suurnäkki, S.; Wahlsten, M.; Alvarenga, D.O.; Sivonen, K.; Fewer, D.P. The Biosynthesis of Rare Homo-Amino Acid Containing Variants of Microcystin by a Benthic Cyanobacterium. Mar. Drugs 2019, 17, 271. https://doi.org/10.3390/md17050271
Shishido TK, Jokela J, Humisto A, Suurnäkki S, Wahlsten M, Alvarenga DO, Sivonen K, Fewer DP. The Biosynthesis of Rare Homo-Amino Acid Containing Variants of Microcystin by a Benthic Cyanobacterium. Marine Drugs. 2019; 17(5):271. https://doi.org/10.3390/md17050271
Chicago/Turabian StyleShishido, Tânia Keiko, Jouni Jokela, Anu Humisto, Suvi Suurnäkki, Matti Wahlsten, Danillo O. Alvarenga, Kaarina Sivonen, and David P. Fewer. 2019. "The Biosynthesis of Rare Homo-Amino Acid Containing Variants of Microcystin by a Benthic Cyanobacterium" Marine Drugs 17, no. 5: 271. https://doi.org/10.3390/md17050271
APA StyleShishido, T. K., Jokela, J., Humisto, A., Suurnäkki, S., Wahlsten, M., Alvarenga, D. O., Sivonen, K., & Fewer, D. P. (2019). The Biosynthesis of Rare Homo-Amino Acid Containing Variants of Microcystin by a Benthic Cyanobacterium. Marine Drugs, 17(5), 271. https://doi.org/10.3390/md17050271