Cervical Cancer Correlates with the Differential Expression of Nicotinic Acetylcholine Receptors and Reveals Therapeutic Targets



Abstract
1. Introduction
2. Results
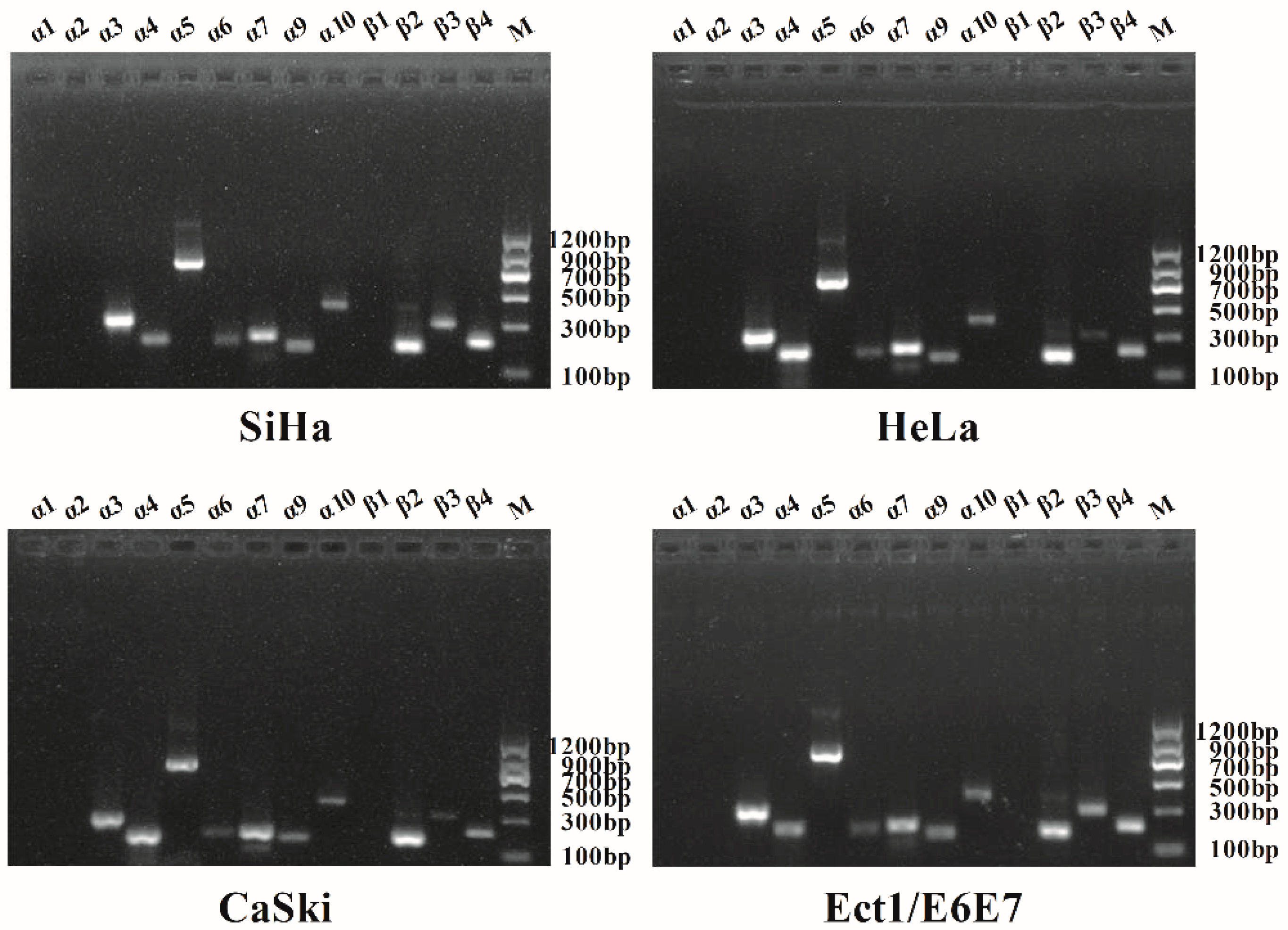
2.1. Qualitative Analysis of nAChR Subunits Expression in Human Cervical Cancer by PCR
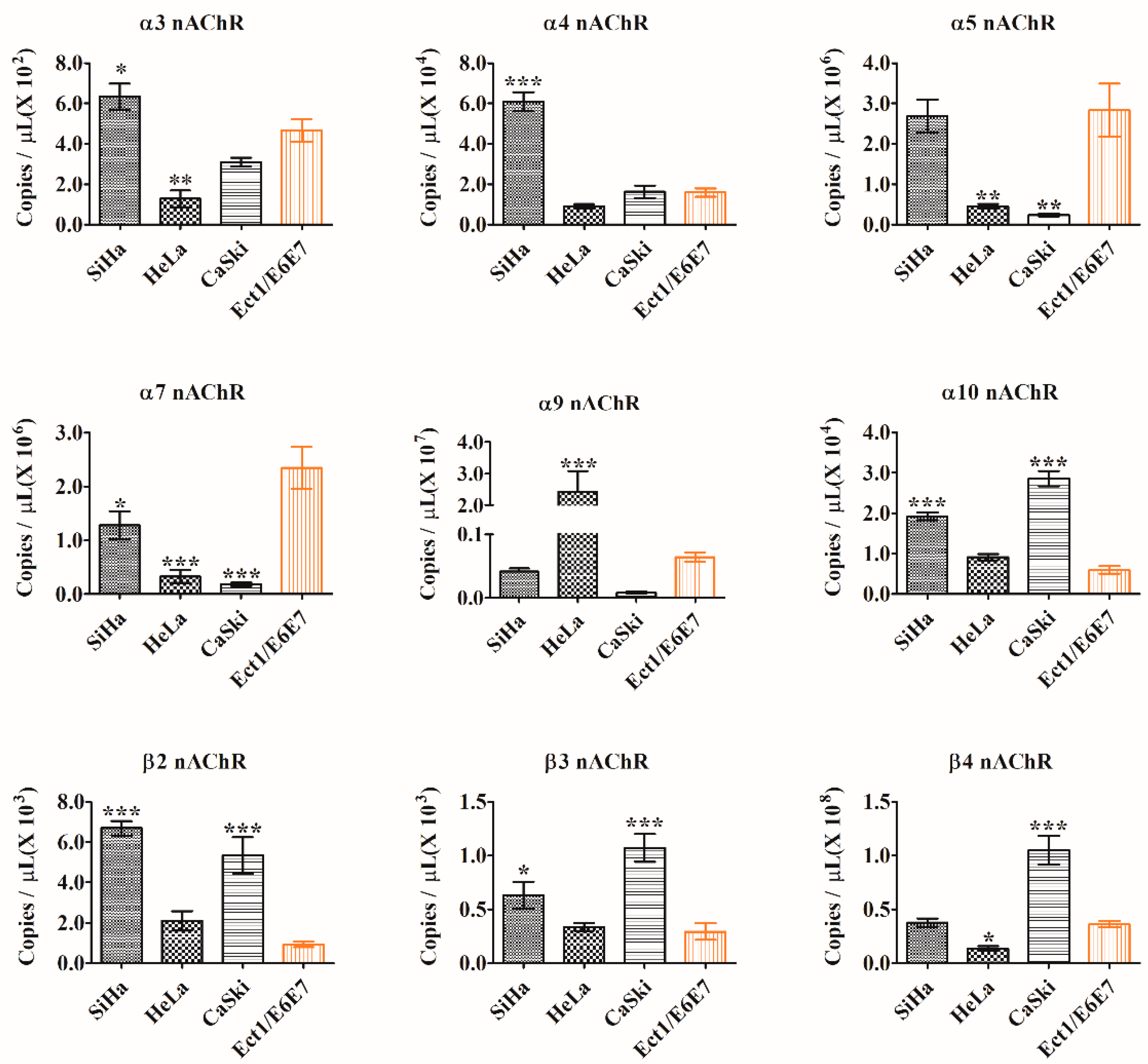
2.2. Quantitative PCR Detection of nAChR Subunits Expression
2.3. Detection of nAChR Subunits by Western Blot
2.4. Effects of α*-Conotoxins on Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. RNA Extraction and Polymerase Chain Reaction (PCR)
4.3. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.4. Protein Extraction and Western Blotting Assay
4.5. MTT Assay
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Vu, M.; Yu, J.; Awolude, O.A.; Chuang, L. Cervical cancer worldwide. Curr. Probl. Cancer 2018, 42, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Crow, J.M. HPV: The global burden. Nature 2012, 488, S2–S3. [Google Scholar]
- Crosbie, E.J.; Einstein, M.H.; Franceschi, S.; Kitchener, H.C. Human papillomavirus and cervical cancer. Lancet 2013, 382, 889–899. [Google Scholar] [CrossRef]
- Chabeda, A.; Yanez, R.J.R.; Lamprecht, R.; Meyers, A.E.; Rybicki, E.P.; Hitzeroth, II. Therapeutic vaccines for high-risk HPV-associated diseases. Papillomavirus Res. 2018, 5, 46–58. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans. Br. J. Pharmacol. 2008, 154, 1558–1571. [Google Scholar] [CrossRef]
- Bierut, L.J. Nicotine dependence and genetic variation in the nicotinic receptors. Drug Alcohol Depend. 2009, 104 (Suppl. 1), S64–S69. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, A.; Nastrucci, C.; Cesario, A.; Russo, P. Nicotine: Specific role in angiogenesis, proliferation and apoptosis. Crit. Rev. Toxicol. 2012, 42, 68–89. [Google Scholar] [CrossRef] [PubMed]
- Schaal, C.; Chellappan, S.P. Nicotine-mediated cell proliferation and tumor progression in smoking-related cancers. Mol. Cancer Res. 2014, 12, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Pillai, S.; Chellappan, S. Nicotinic acetylcholine receptor signaling in tumor growth and metastasis. Int. J. Oncol. 2011, 2011, 1–11. [Google Scholar] [CrossRef]
- Chen, R.J.; Ho, Y.S.; Guo, H.R.; Wang, Y.J. Rapid activation of Stat3 and ERK1/2 by nicotine modulates cell proliferation in human bladder cancer cells. Toxicol. Sci. 2008, 104, 283–293. [Google Scholar] [CrossRef]
- Wei, P.L.; Chang, Y.J.; Ho, Y.S.; Lee, C.H.; Yang, Y.Y.; An, J.; Lin, S.Y. Tobacco-specific carcinogen enhances colon cancer cell migration through alpha7-nicotinic acetylcholine receptor. Ann. Surg. 2009, 249, 978–985. [Google Scholar] [CrossRef]
- Wang, W.; Chin-Sheng, H.; Kuo, L.J.; Wei, P.L.; Lien, Y.C.; Lin, F.Y.; Liu, H.H.; Ho, Y.S.; Wu, C.H.; Chang, Y.J. NNK enhances cell migration through α7-nicotinic acetylcholine receptor accompanied by increased of fibronectin expression in gastric cancer. Ann. Surg. Oncol. 2012, 19 (Suppl. 3), 580–588. [Google Scholar] [CrossRef]
- Nieh, S.; Jao, S.W.; Yang, C.Y.; Lin, Y.-S.; Tseng, Y.H.; Liu, C.L.; Lee, T.Y.; Liu, T.Y.; Chu, Y.H.; Chen, S.F. Regulation of tumor progression via the Snail-RKIP signaling pathway by nicotine exposure in head and neck squamous cell carcinoma. Head Neck 2015, 37, 1712–1721. [Google Scholar]
- Jin, T.; Hao, J.; Fan, D. Nicotine induces aberrant hypermethylation of tumor suppressor genes in pancreatic epithelial ductal cells. Biochem. Biophys. Res. Commun. 2018, 499, 934–940. [Google Scholar] [CrossRef]
- Medjber, K.; Freidja, M.L.; Grelet, S.; Lorenzato, M.; Maouche, K.; Nawrocki-Raby, B.; Birembaut, P.; Polette, M.; Tournier, J.M. Role of nicotinic acetylcholine receptors in cell proliferation and tumour invasion in broncho-pulmonary carcinomas. Lung Cancer 2015, 87, 258–264. [Google Scholar] [CrossRef]
- Brown, K.C.; Lau, J.K.; Dom, A.M.; Witte, T.R.; Luo, H.; Crabtree, C.M.; Shah, Y.H.; Shiflett, B.S.; Marcelo, A.J.; Proper, N.A. MG624, an α7-nAChR antagonist, inhibits angiogenesis via the Egr-1/FGF2 pathway. Angiogenesis 2012, 15, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Rizwani, W.; Pillai, S.; Kinkade, R.; Kovacs, M.; Rastogi, S.; Banerjee, S.; Carless, M.; Kim, E.; Coppola, D. Nicotine induces cell proliferation, invasion and epithelial-mesenchymal transition in a variety of human cancer cell lines. Int. J. Cancer 2009, 124, 36–45. [Google Scholar] [CrossRef]
- Improgo, M.R.; Schlichting, N.A.; Cortes, R.Y.; Zhao-Shea, R.; Tapper, A.R.; Gardner, P.D. ASCL1 regulates the expression of the CHRNA5/A3/B4 lung cancer susceptibility locus. Mol. Cancer Res. 2010, 8, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Improgo, M.R.; Soll, L.G.; Tapper, A.R.; Gardner, P.D. Nicotinic acetylcholine receptors mediate lung cancer growth. Front Physiol. 2013, 4, 251. [Google Scholar] [CrossRef]
- Lee, C.H.; Huang, C.S.; Chen, C.S.; Tu, S.H.; Wang, Y.J.; Chang, Y.J.; Tam, K.W.; Wei, P.L.; Cheng, T.C.; Chu, J.-S. Overexpression and activation of the α9-nicotinic receptor during tumorigenesis in human breast epithelial cells. J. Natl. Cancer Inst. 2010, 102, 1322–1335. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.H.; Ku, C.Y.; Ho, C.T.; Chen, C.S.; Huang, C.S.; Lee, C.H.; Chang, Y.J. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits nicotine- and estrogen-induced alpha9-nicotinic acetylcholine receptor upregulation in human breast cancer cells. Mol. Nutr. Food Res. 2011, 55, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kaas, Q.; Wu, Y.; Zhu, X.; Romero, H.K. Cloning, synthesis, and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, 4026–4035. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a Novel α-Conotoxin TxID from Conus textile That Potently Blocks Rat α3β4 Nicotinic Acetylcholine Receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [CrossRef]
- Zhangsun, D.; Zhu, X.; Kaas, Q.; Wu, Y.; Craik, D.J.; McIntosh, J.M.; Luo, S. αO-Conotoxin GeXIVA disulfide bond isomers exhibit differential sensitivity for various nicotinic acetylcholine receptors but retain potency and selectivity for the human α9α10 subtype. Neuropharmacology 2017, 127, 243–252. [Google Scholar] [CrossRef]
- Grando, S.A. Connections of nicotine to cancer. Nat. Rev. Cancer 2014, 14, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. The Oncogenic Functions of Nicotinic Acetylcholine Receptors. J. Oncol. 2016, 2016, 9650481. [Google Scholar] [CrossRef] [PubMed]
- Sciamanna, M.A.; Griesmann, G.E.; Williams, C.L.; Lennon, V.A. Nicotinic acetylcholine receptors of muscle and neuronal (α7) types coexpressed in a small cell lung carcinoma. J. Neurochem. 1997, 69, 2302–2311. [Google Scholar] [CrossRef]
- Calleja-Macias, I.E.; Kalantari, M.; Bernard, H.U. Cholinergic signaling through nicotinic acetylcholine receptors stimulates the proliferation of cervical cancer cells: An explanation for the molecular role of tobacco smoking in cervical carcinogenesis? Int. J. Cancer 2009, 124, 1090–1096. [Google Scholar] [CrossRef]
- De Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef]
- Day, D.A.; Tuite, M.F. Post-transcriptional gene regulatory mechanisms in eukaryotes: An overview. J. Endocrinol. 1998, 157, 361–371. [Google Scholar] [CrossRef]
- Kong, J.; Lasko, P. Translational control in cellular and developmental processes. Nat. Rev. Genet. 2012, 13, 383–394. [Google Scholar] [CrossRef]
- Yang, J.R. Does mRNA structure contain genetic information for regulating co-translational protein folding? Zool. Res. 2017, 38, 36–43. [Google Scholar]
- Wu, J.; Lukas, R.J. Naturally-expressed nicotinic acetylcholine receptor subtypes. Biochem. Pharmacol. 2011, 82, 800–807. [Google Scholar] [CrossRef]
- Chen, C.S.; Lee, C.H.; Hsieh, C.D.; Ho, C.T.; Pan, M.H.; Huang, C.S.; Wei, P.L. Nicotine-induced human breast cancer cell proliferation attenuated by garcinol through down-regulation of the nicotinic receptor and cyclin D3 proteins. Breast Cancer Res. Treat. 2011, 125, 73–87. [Google Scholar] [CrossRef]
- Shih, Y.L.; Liu, H.C.; Chen, C.S.; Hsu, C.H.; Pan, M.H.; Chang, H.W.; Ho, Y.S. Combination treatment with luteolin and quercetin enhances antiproliferative effects in nicotine-treated MDA-MB-231 cells by down-regulating nicotinic acetylcholine receptors. J. Agric. Food Chem. 2010, 58, 235–241. [Google Scholar] [CrossRef]
- Ma, X.; Jia, Y.; Zu, S.; Li, R.; Jia, Y.; Zhao, Y.; Xiao, D.; Dang, N.; Wang, Y. α5 Nicotinic acetylcholine receptor mediates nicotine-induced HIF-1alpha and VEGF expression in non-small cell lung cancer. Toxicol. Appl. Pharmacol. 2014, 278, 172–179. [Google Scholar] [CrossRef]
- Sun, H.; Ma, X. α5-nAChR modulates nicotine-induced cell migration and invasion in A549 lung cancer cells. Exp. Toxicol. Pathol. 2015, 67, 477–482. [Google Scholar] [CrossRef]
- Sun, H.J.; Jia, Y.F.; Ma, X.L. Alpha5 Nicotinic Acetylcholine Receptor Contributes to Nicotine-Induced Lung Cancer Development and Progression. Front Pharmacol. 2017, 8, 573. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, X.P.; Zhao, Q.N.; Yang, X.J.; An, S.M.; Wang, H.; Xu, L.; Zhu, L.; Chen, H.Z. Role of α7-nicotinic acetylcholine receptor in nicotine-induced invasion and epithelial-to-mesenchymal transition in human non-small cell lung cancer cells. Oncotarget 2016, 7, 59199–59208. [Google Scholar]
- Cesario, A.; Russo, P.; Nastrucci, C.; Granone, P. Is α7-nAChR a possible target for lung cancer and malignant pleural mesothelioma treatment? Curr. Drug Targets 2012, 13, 688–694. [Google Scholar]
- Schuller, H.M. Regulatory Role of the α7 nAChR in Cancer. Curr. Drug Targets 2012, 13, 680–687. [Google Scholar] [CrossRef]
- Cesario, A.; Russo, P.; Viaggi, S.; Trombino, S.; Imperatori, A.; Margaritora, S.; Dominioni, L.; Festi, L.; Porziella, V.; Granone, P. Malignant pleural mesothelioma: Time for translational research. Lancet Oncol. 2004, 5, 591. [Google Scholar] [CrossRef]
- Luo, S.Q.; Kulak, J.M.; Cartier, G.E.; Jacobsen, R.B.; Yoshikami, D.; Olivera, B.M.; Mcintosh, J.M. α-conotoxin AuIB selectively blocks α3β4 nicotinic acetylcholine receptors and nicotine-evoked norepinephrine release. J. Neurosci. 1998, 18, 8571–8579. [Google Scholar] [CrossRef]
- Louw-du Toit, R.; Hapgood, J.P.; Africander, D. Medroxyprogesterone acetate differentially regulates interleukin (IL)-12 and IL-10 in a human ectocervical epithelial cell line in a glucocorticoid receptor (GR)-dependent manner. J. Biol. Chem. 2014, 289, 31136–31149. [Google Scholar] [CrossRef]
- Fichorova, R.N.; Rheinwald, J.G.; Anderson, D.J. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol. Reprod. 1997, 57, 847–855. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| nAChR Subunits | Regression Equation of Standard Curve | R2 | CT Range |
|---|---|---|---|
| α3 | y = −3.087x + 33.55 | 0.997 | 8.18–30.05 |
| α4 | y = −3.207x + 39.08 | 0.995 | 10.63–29.56 |
| α5 | y = −3.042x + 40.70 | 0.995 | 8.69–26.07 |
| α7 | y = −3.3594x + 42.339 | 0.991 | 13.09–29.26 |
| α9 | y = −3.058x + 45.17 | 0.991 | 12.24–31.01 |
| α10 | y = −3.2786x + 33.164 | 0.999 | 8.15–28.20 |
| β2 | y = −3.304x + 35.73 | 0.992 | 10.91–29.61 |
| β3 | y = −3.036x + 33.70 | 0.998 | 8.74–27.24 |
| β4 | y = −3.358x + 48.09 | 0.996 | 10.15–26.78 |
| nAChR Subunits | SiHa | HeLa | CaSki | Ect1/E6E7 (Control) | |||
|---|---|---|---|---|---|---|---|
| Copies/μL | Significance | Copies/μL | Significance | Copies/μL | Significance | Copies/μL | |
| α3 | 6.35 × 102 | * | 1.28 × 102 | ## | 3.10 × 102 | ns | 4.66 × 102 |
| α4 | 6.09 × 104 | *** | 9.08 × 103 | ns | 1.63 × 104 | ns | 1.60 × 104 |
| α5 | 2.69 × 106 | ns | 4.55 × 105 | ## | 2.37 × 105 | ## | 2.84 × 106 |
| α7 | 1.28 × 106 | # | 3.26 × 105 | ### | 1.82 × 105 | ### | 2.35 × 106 |
| α9 | 4.15 × 105 | ns | 2.42 × 107 | *** | 8.19 × 104 | ns | 6.43 × 105 |
| α10 | 1.92 × 104 | *** | 9.14 × 103 | ns | 2.86 × 104 | *** | 5.96 × 103 |
| β2 | 6.68 × 103 | *** | 2.10 × 103 | ns | 5.34 × 103 | *** | 9.29 × 102 |
| β3 | 6.33 × 102 | * | 3.37 × 102 | ns | 1.07 × 103 | *** | 2.95 × 102 |
| β4 | 3.78 × 107 | ns | 1.35 × 107 | # | 1.05 × 108 | *** | 3.64 × 107 |
| Cell Lines | Organism | Years | Derivation and Disease | |
|---|---|---|---|---|
| No. | Name | |||
| 1 | SiHa | Homo sapiens, human | 1975 | Female, 55 years, Asian, grade II, squamous cell carcinoma |
| 2 | HeLa | Homo sapiens, human | 1951 | Female, 31 years, Black, adenocarcinoma |
| 3 | CaSki | Homo sapiens, human | 1974 | Female, 40 years, Caucasian, epidermoid carcinoma |
| 4 | Ect1/E6E7 | Homo sapiens, human | 1996 | Female, 43 years, unknown, normal |
| Primer | Accession No. | Sequence | Amplified Region | Length | Application | |
|---|---|---|---|---|---|---|
| No | Name | |||||
| 1 | α1 nAChR-F | NM_001039523.2 | CGTCTGGTGGCAAAGCT | (7–587) a | 581bp | PCR |
| α1 nAChR-R | CCGCTCTCCATGAAGTT | |||||
| 2 | α2 nAChR-F | NM_001282455.1 | GAGGACCGGCTCTTCAAACA | (1–289) a | 289bp | PCR |
| α2 nAChR-R | ACTCCCCATCTGCATTGTTGT | |||||
| 3 | α3 nAChR-F | NM_000743.4 | CTGGTGAAGGTGGATGAAGTAAA | (109–410) a | 302bp | PCR |
| α3 nAChR-R | TGGTAATCAAACGGGAAGTAGGT | |||||
| 4 | α3 nAChR-F | AACGTGTCTGACCCAGTCATCAT | (61–199) a | 139bp | qPCR | |
| α3 nAChR-R | AGGGGTTCCATTTCAGCTTGTAG | |||||
| 5 | α4 nAChR-F | L35901.1 | ATGAAATTCGGCTCCTGGACCTA | (1–201) a | 201bp | PCR, qPCR |
| α4 nAChR-R | CGGCAGCCGCCGGATGAC | |||||
| 6 | α5 nAChR-F | BC033639.1 | GAGAGGATTATCTGAACCTTCTTCT | (102–869) b | 768bp | PCR |
| α5 nAChR-R | ACAAGTACTGAAGTGCAGAGACA | |||||
| 7 | α5 nAChR-F | ACTCCACCGGCAAACTACAA | (343–609) a | 267bp | qPCR | |
| α5 nAChR-R | CAGGCGCTTGATTACAAATGA | |||||
| 8 | α6 nAChR-F | NM_004198.3 | GGCTGCGTCACATCTGGAA | (155–356) a | 202bp | PCR |
| α6 nAChR-R | GCTGGTGGAGTCCAGGTTAT | |||||
| 9 | α7 nAChR-F | NM_000746.5 | CCACCAACATTTGGCTGCAA | (143–360) a | 218bp | PCR, qPCR |
| α7 nAChR-R | TATGCCTGGAGGCAGGTACT | |||||
| 10 | α9 nAChR-F | NM_017581.3 | TGGCACGATGCCTATCTCAC | (169–340) a | 172bp | PCR, qPCR |
| α9 nAChR-R | TGATCAGCCCATCATACCGC | |||||
| 11 | α10 nAChR-F | AF199235.2 | CTGTTCCGTGACCTCTTT | (4–391) a | 388bp | PCR |
| α10 nAChR-R | GGAAGGCTGCTACATCCA | |||||
| 12 | α10 nAChR-F | TGACCTCTTTGCCAACTACAC | (12–149) a | 138bp | qPCR | |
| α10 nAChR-R | CACAGATACAGGGTCAGCAC | |||||
| 13 | β1 nAChR-F | NM_000747.2 | AGACCTGGAGTGGACTGACT | (159–317) a | 159bp | PCR |
| β1 nAChR-R | ACGACGCTAATGTCCAGAGC | |||||
| 14 | β2 nAChR-F | NM_000748.2 | GGCATGTACGAGGTGTCCTT | (289–473) a | 185bp | PCR, qPCR |
| β2 nAChR-R | ACCAAGTCGATCTCTGTGCG | |||||
| 15 | β3 nAChR-F | NM_000749.4 | GGGTCCGCCCTGTATTACATT | (44–348) a | 305bp | PCR |
| β3 nAChR-R | GGTCCAGACAACAGTTCCGT | |||||
| 16 | β3 nAChR-F | GGTCCGCCCTGTATTACATTC | (45–190) a | 146bp | qPCR | |
| β3 nAChR-R | AGCGTAACTTGTGGTCTGTC | |||||
| 17 | β4 nAChR-F | U48861.1 | CGCTACGAGGGTGTGAACAT | (208–420) a | 213bp | PCR, qPCR |
| β4 nAChR-R | GTTCTGCTGGTCGAAGGGAA | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Qian, J.; Sun, Z.; Zhangsun, D.; Luo, S. Cervical Cancer Correlates with the Differential Expression of Nicotinic Acetylcholine Receptors and Reveals Therapeutic Targets. Mar. Drugs 2019, 17, 256. https://doi.org/10.3390/md17050256
Liu Y, Qian J, Sun Z, Zhangsun D, Luo S. Cervical Cancer Correlates with the Differential Expression of Nicotinic Acetylcholine Receptors and Reveals Therapeutic Targets. Marine Drugs. 2019; 17(5):256. https://doi.org/10.3390/md17050256
Chicago/Turabian StyleLiu, Yiqiao, Jiang Qian, Zhihua Sun, Dongting Zhangsun, and Sulan Luo. 2019. "Cervical Cancer Correlates with the Differential Expression of Nicotinic Acetylcholine Receptors and Reveals Therapeutic Targets" Marine Drugs 17, no. 5: 256. https://doi.org/10.3390/md17050256
APA StyleLiu, Y., Qian, J., Sun, Z., Zhangsun, D., & Luo, S. (2019). Cervical Cancer Correlates with the Differential Expression of Nicotinic Acetylcholine Receptors and Reveals Therapeutic Targets. Marine Drugs, 17(5), 256. https://doi.org/10.3390/md17050256