Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio

 and
and 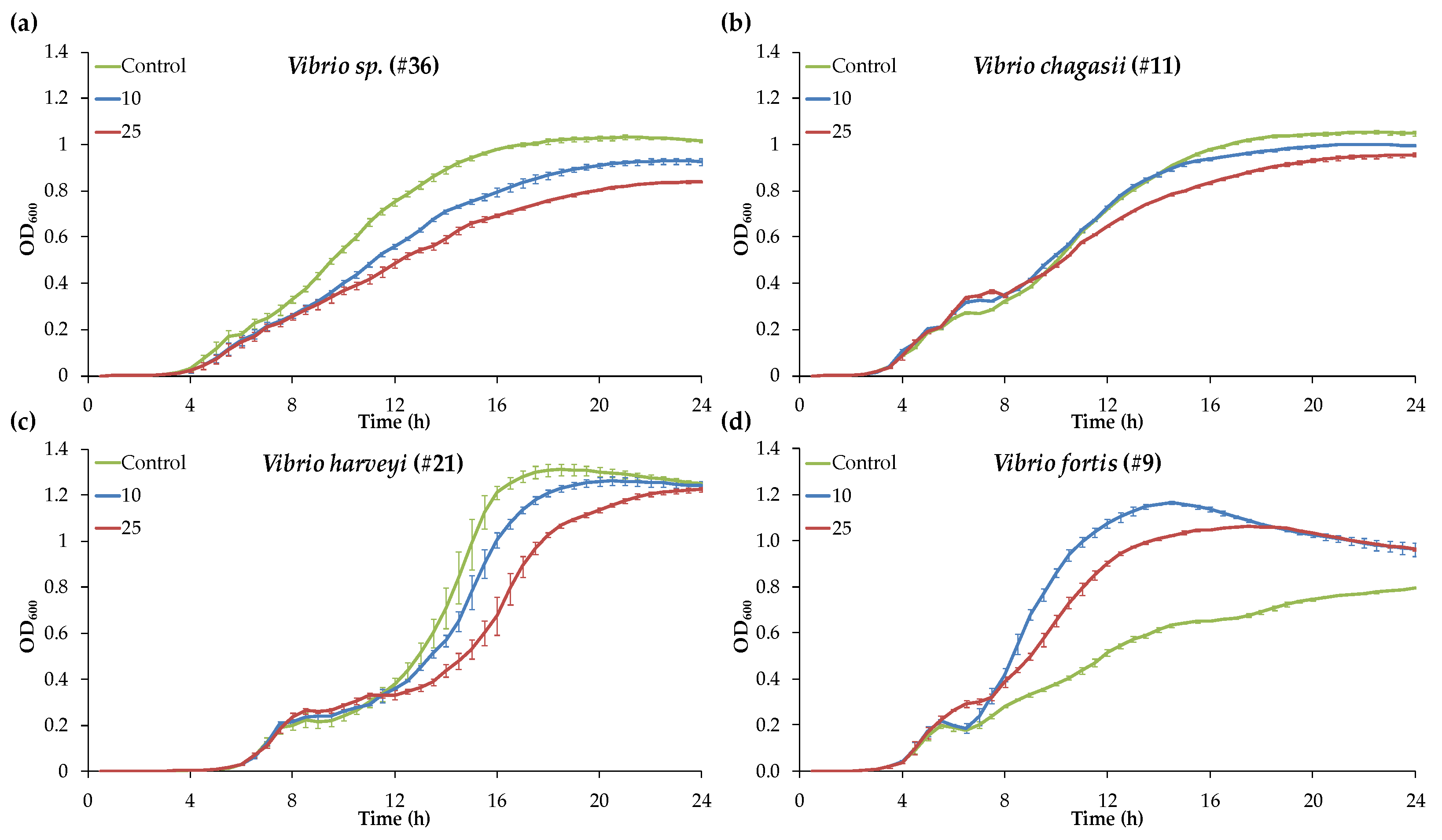{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Vibrio Strains
2.2. Vibrio Exposure to Blue Water Solutions
2.2.1. Preparation of Bacterial Inocula
2.2.2. Blue Water (BW) Production
2.2.3. Antibacterial Essay
2.3. Growth Curve Analyses and Statistics
3. Results
3.1. Different Patterns of Vibrio Growth Curves Evidenced by the Screening Experiment
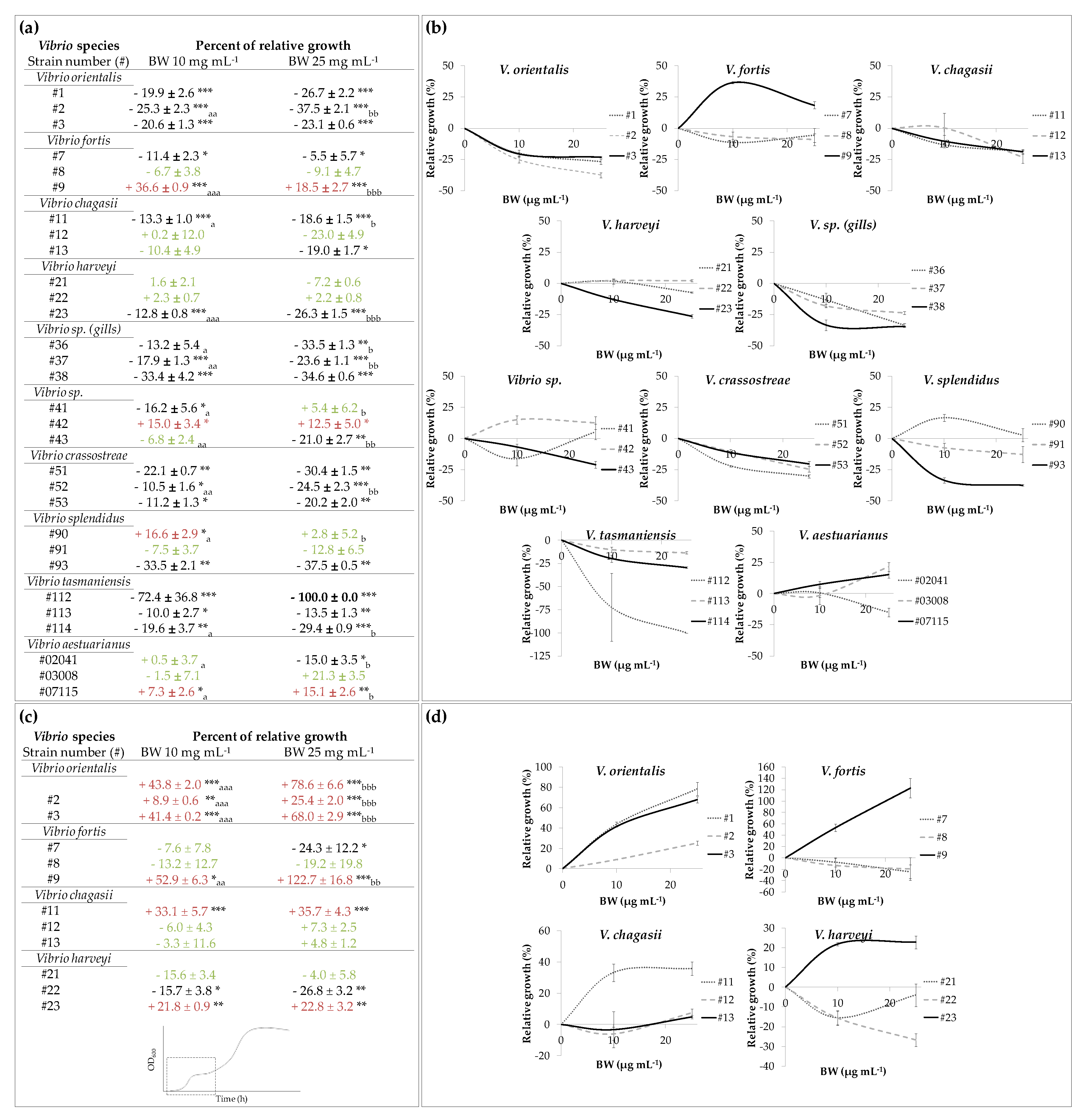
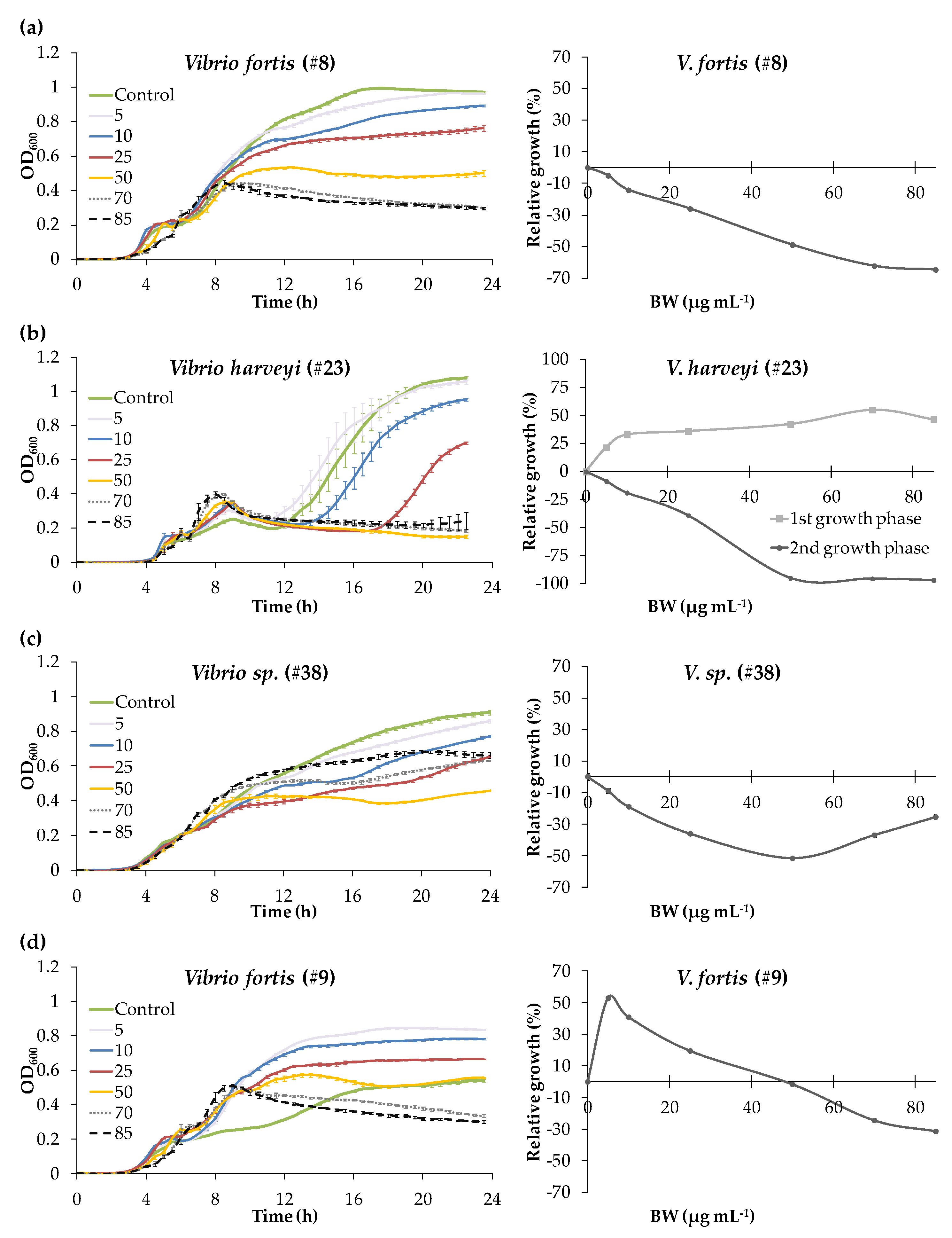
3.2. Experiment with Blue Water Concentration Range
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nassiri, Y.; Robert, J.-M.; Rincé, Y.; Ginsburger-Vogel, T. The cytoplasmic fine structure of the diatom Haslea ostrearia (Bacillariophyceae) in relation to marennine production. Phycologia 1998, 37, 84–91. [Google Scholar] [CrossRef]
- Gastineau, R.; Davidovich, N.A.; Bardeau, J.-F.; Caruso, A.; Leignel, V.; Hardivillier, Y.; Jacquette, B.; Davidovich, O.I.; Rincé, Y.; Gaudin, P.; et al. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the Black Sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar] [CrossRef]
- Gastineau, R.; Hansen, G.; Davidovich, N.A.; Davidovich, O.; Bardeau, J.-F.; Kaczmarska, I.; Ehrman, J.M.; Leignel, V.; Hardivillier, Y.; Jacquette, B.; et al. A new blue-pigmented hasleoid diatom, Haslea provincialis, from the Mediterranean Sea. Eur. J. Phycol. 2016, 51, 156–170. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Gastineau, R.; Poulin, M.; Lemieux, C.; Turmel, M.; Syakti, A.D.; Hardivillier, Y.; Widowati, I.; Risjani, Y.; Iskandar, I.; et al. Haslea nusantara, a new blue diatom from the Java Sea, Indonesia: Morphology, biometry and molecular characterization. Plant. Ecol. Evol. 2019, 151. (in press).
- Pouvreau, J.-B.; Morançais, M.; Fleury, F.; Rosa, P.; Thion, L.; Cahingt, B.; Zal, F.; Fleurence, J.; Pondaven, P. Preliminary characterisation of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 757–767. [Google Scholar] [CrossRef]
- Turpin, V.; Robert, J.-M.; Goulletquer, P.; Massé, G.; Rosa, P. Oyster greening by outdoor mass culture of the diatom Haslea ostrearia Simonsen in enriched seawater. Aquac. Res. 2001, 32, 801–809. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Housson, E.; Tallec, L.L.; Morançais, M.; Rincé, Y.; Fleurence, J.; Pondaven, P. Growth inhibition of several marine diatom species induced by the shading effect and allelopathic activity of marennine, a blue-green polyphenolic pigment of the diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Exp. Mar. Biol. Ecol. 2007, 352, 212–225. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Safitri, I.; Widowati, I.; Cognie, B.; Decottignies, P.; Gastineau, R.; Morançais, M.; Windarto, E.; Tremblay, R.; Mouget, J.-L. Does allelopathy affect co-culturing Haslea ostrearia with other microalgae relevant to aquaculture? J. Appl. Phycol. 2016, 28, 2241–2254. [Google Scholar] [CrossRef]
- Falaise, C.; François, C.; Travers, M.-A.; Morga, B.; Haure, J.; Tremblay, R.; Turcotte, F.; Pasetto, P.; Gastineau, R.; Hardivillier, Y.; et al. Antimicrobial compounds from eukaryotic microalgae against human pathogens and diseases in aquaculture. Mar. Drugs 2016, 14, 159. [Google Scholar] [CrossRef]
- Pouvreau, J.-B. Purification et Caractérisation du Pigment Bleu-Vert “Marennine” Synthétisé par la Diatomee Marine Haslea ostrearia (Gaillon/Bory) Simonsen: Propriétés Physico-Chimiques et Activités Biologiques. Ph.D. Thesis, Université de Nantes, Nantes, France, July 2006. [Google Scholar]
- Gastineau, R.; Pouvreau, J.-B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Bourgougnon, N.; Mouget, J.-L. Biological activities of purified marennine, the blue pigment responsible for the greening of oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.S.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, promising blue pigments from a widespread Haslea diatom species complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [PubMed]
- Travers, M.-A.; Boettcher Miller, K.; Roque, A.; Friedman, C.S. Bacterial diseases in marine bivalves. J. Invertebr. Pathol. 2015, 131, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Hardivillier, Y.; Leignel, V.; Tekaya, N.; Morançais, M.; Fleurence, J.; Davidovich, N.; Jacquette, B.; Gaudin, P.; Hellio, C.; et al. Greening effect on oysters and biological activities of the blue pigments produced by the diatom Haslea karadagensis (Naviculaceae). Aquaculture 2012, 368–369, 61–67. [Google Scholar] [CrossRef]
- Turcotte, F.; Mouget, J.-L.; Genard, B.; Lemarchand, K.; Deschênes, J.-S.; Tremblay, R. Prophylactic effect of Haslea ostrearia culture supernatant containing the pigment marennine to stabilize bivalve hatchery production. Aquat. Living Resour. 2016, 29, 401. [Google Scholar] [CrossRef]
- Bruto, M.; James, A.; Petton, B.; Labreuche, Y.; Chenivesse, S.; Alunno-Bruscia, M.; Polz, M.F.; Le Roux, F. Vibrio crassostreae, a benign oyster colonizer turned into a pathogen after plasmid acquisition. ISME J. 2017, 11, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Document M07-A9. Approved Standard—Ninth Edition; Clinical and Laboratory Institute: Wayne, PA, USA, 2012; Volume 32. [Google Scholar]
- Pouvreau, J.-B.; Morançais, M.; Massé, G.; Rosa, P.; Robert, J.-M.; Fleurence, J.; Pondaven, P. Purification of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 769–781. [Google Scholar] [CrossRef]
- Sprouffske, K.; Wagner, A. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinform. 2016, 17, 1–4. [Google Scholar] [CrossRef]
- Monod, J. The Growth of Bacterial Cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef]
- Falaise, C.; Cormier, P.; Tremblay, R.; Audet, C.; Deschênes, J.-S.; Turcotte, F.; François, C.; Seger, A.; Hallegraeff, G.; Lindquist, N.; et al. Harmful or harmless: Biological effects of marennine on marine organisms. Aquatic Toxicol. 2019, 209, 13–25. [Google Scholar] [CrossRef]
- Bag, J. Glucose inhibition of the transport and phosphoenolpyruvate-dependent phosphorylation of galactose and fructose in Vibrio cholerae. J. Bacteriol. 1974, 118, 764–767. [Google Scholar]
- Bhattacharya, D.; Ghosh, D.; Bhattacharya, S.; Sarkar, S.; Karmakar, P.; Koley, H.; Gachhui, R. Antibacterial activity of polyphenolic fraction of Kombucha against Vibrio cholerae: Targeting cell membrane. Lett. Appl. Microbiol. 2018, 66, 145–152. [Google Scholar] [CrossRef]
- Chatterjee, T.; Chatterjee, B.K.; Chakrabarti, P. Modelling of growth kinetics of Vibrio cholerae in presence of gold nanoparticles: Effect of size and morphology. Sci. Rep. 2017, 7, 9671. [Google Scholar] [CrossRef] [PubMed]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. In vitro growth characteristics of five candidate aquaculture probiotics and two fish pathogens grown in fish intestinal mucus. FEMS Microbiol. Lett. 2004, 231, 145–152. [Google Scholar] [CrossRef]
- Banerjee, S.; Kim, L.M.; Shariff, M.; Khatoon, H.; Yusoff, F.M. Antibacterial activity of neem (Azadirachta indica) leaves on Vibrio spp. isolated from cultured shrimp. Asian J. Anim. Vet. Adv. 2013, 8, 355–361. [Google Scholar]
- Cavallo, R.; Acquaviva, M.; Stabili, L.; Cecere, E.; Petrocelli, A.; Narracci, M. Antibacterial activity of marine macroalgae against fish pathogenic Vibrio species. Open Life Sci. 2013, 8, 646–653. [Google Scholar]
- Das, B.K.; Pradhan, J.; Pattnaik, P.; Samantaray, B.R.; Samal, S.K. Production of antibacterials from the freshwater alga Euglena viridis (Ehren). World J. Microbiol. Biotechnol. 2005, 21, 45–50. [Google Scholar] [CrossRef]
- Solopova, A.; van Gestel, J.; Weissing, F.J.; Bachmann, H.; Teusink, B.; Kok, J.; Kuipers, O.P. Bet-hedging during bacterial diauxic shift. PNAS 2014, 111, 7427–7432. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M. A functional perspective on phenotypic heterogeneity in microorganisms. Nat. Rev. Microbiol. 2015, 13, 497–508. [Google Scholar] [CrossRef]
- Grimbergen, A.J.; Siebring, J.; Solopova, A.; Kuipers, O.P. Microbial bet-hedging: The power of being different. Curr. Opin.Microbiol. 2015, 25, 67–72. [Google Scholar] [CrossRef]
- Martins, B.M.; Locke, J.C. Microbial individuality: How single-cell heterogeneity enables population level strategies. Curr. Opin. Microbiol. 2015, 24, 104–112. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormetic mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Defining hormesis. Hum. Exp. Toxicol 2002, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G.; Duke, S.O. Herbicides and plant hormesis. Pest. Manag. Sci. 2014, 70, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: A Generalizable and Unifying Hypothesis. Crit. Rev. Toxicol. 2001, 31, 353–424. [Google Scholar] [CrossRef] [PubMed]
- Cedergreen, N.; Streibig, J.C.; Kudsk, P.; Mathiassen, S.K.; Duke, S.O. The occurrence of hormesis in plants and algae. Dose Response 2006, 5, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Kendig, E.L.; Le, H.H.; Belcher, S.M. Defining Hormesis: Evaluation of a Complex Concentration Response Phenomenon. Int. J. Toxicol. 2010, 29, 235–246. [Google Scholar] [CrossRef]
- Lushchak, V.I. Dissection of the Hormetic Curve: Analysis of Components and Mechanisms. Dose Response 2014, 12, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. A quantitatively-based methodology for the evaluation of chemical hormesis. Hum. Ecolog. Risk Assess. An. Int. J. 1997, 3, 545–554. [Google Scholar] [CrossRef]
- Wang, D.; Lin, Z.; Wang, T.; Ding, X.; Liu, Y. An analogous wood barrel theory to explain the occurrence of hormesis: A case study of sulfonamides and erythromycin on Escherichia coli growth. PLoS ONE 2017, 12, e0181321. [Google Scholar] [CrossRef]
- Tardy-Laporte, C.; Arnold, A.A.; Genard, B.; Gastineau, R.; Morançais, M.; Mouget, J.-L.; Tremblay, R.; Marcotte, I. A 2H solid-state NMR study of the effect of antimicrobial agents on intact Escherichia coli without mutating. Biochim. Biophys. Acta (BBA)—Biomembr. 2013, 1828, 614–622. [Google Scholar] [CrossRef]
- Sarwar, S.; Chakraborti, S.; Bera, S.; Sheikh, I.A.; Hoque, K.M.; Chakrabarti, P. The antimicrobial activity of ZnO nanoparticles against Vibrio cholerae: Variation in response depends on biotype. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1499–1509. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falaise, C.; James, A.; Travers, M.-A.; Zanella, M.; Badawi, M.; Mouget, J.-L. Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio. Mar. Drugs 2019, 17, 160. https://doi.org/10.3390/md17030160
Falaise C, James A, Travers M-A, Zanella M, Badawi M, Mouget J-L. Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio. Marine Drugs. 2019; 17(3):160. https://doi.org/10.3390/md17030160
Chicago/Turabian StyleFalaise, Charlotte, Adèle James, Marie-Agnès Travers, Marie Zanella, Myriam Badawi, and Jean-Luc Mouget. 2019. "Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio" Marine Drugs 17, no. 3: 160. https://doi.org/10.3390/md17030160
APA StyleFalaise, C., James, A., Travers, M.-A., Zanella, M., Badawi, M., & Mouget, J.-L. (2019). Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio. Marine Drugs, 17(3), 160. https://doi.org/10.3390/md17030160