A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides

 , ,
, , 
 and
and 
Abstract
1. Introduction
2. Results and Discussion
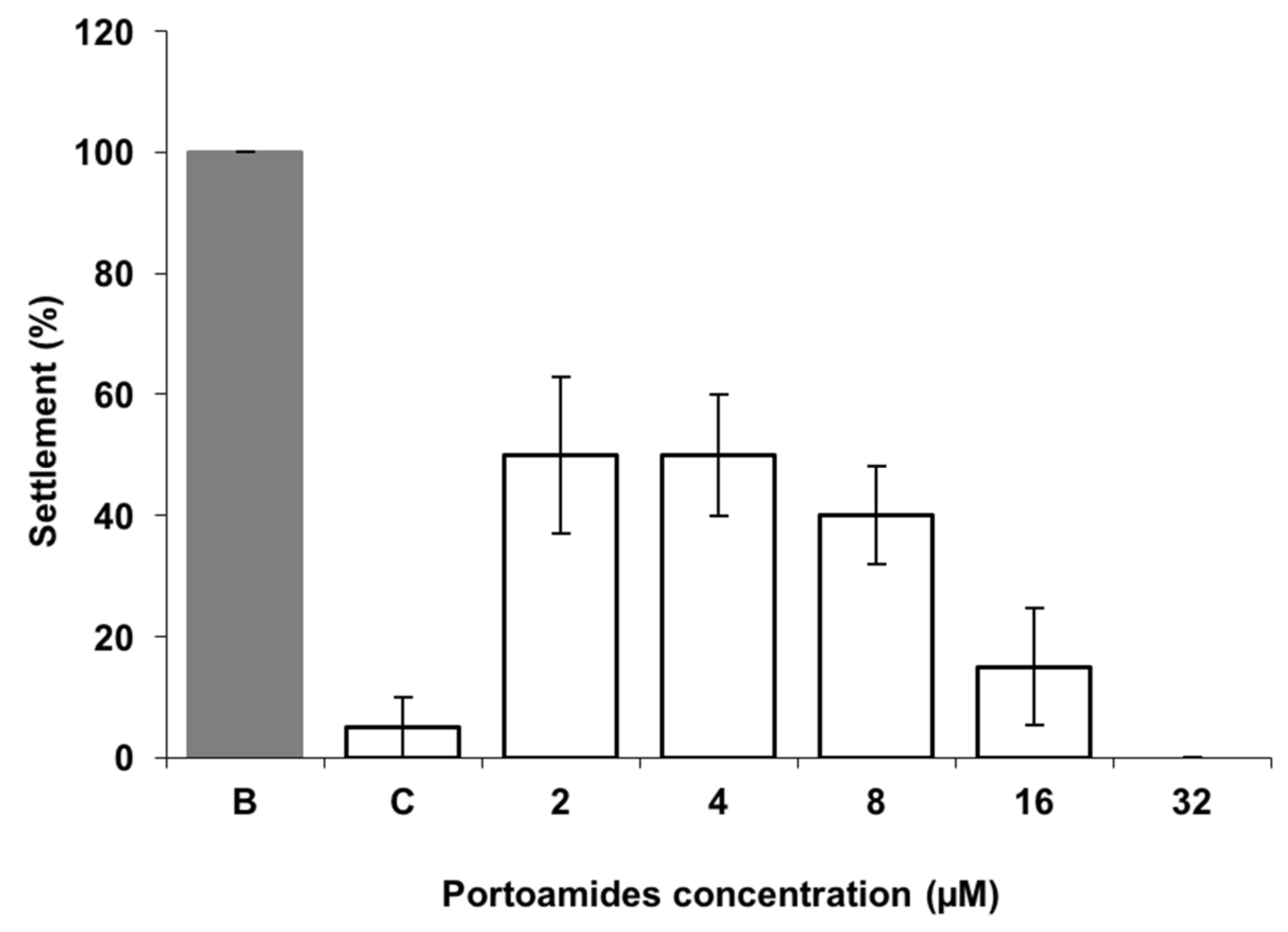
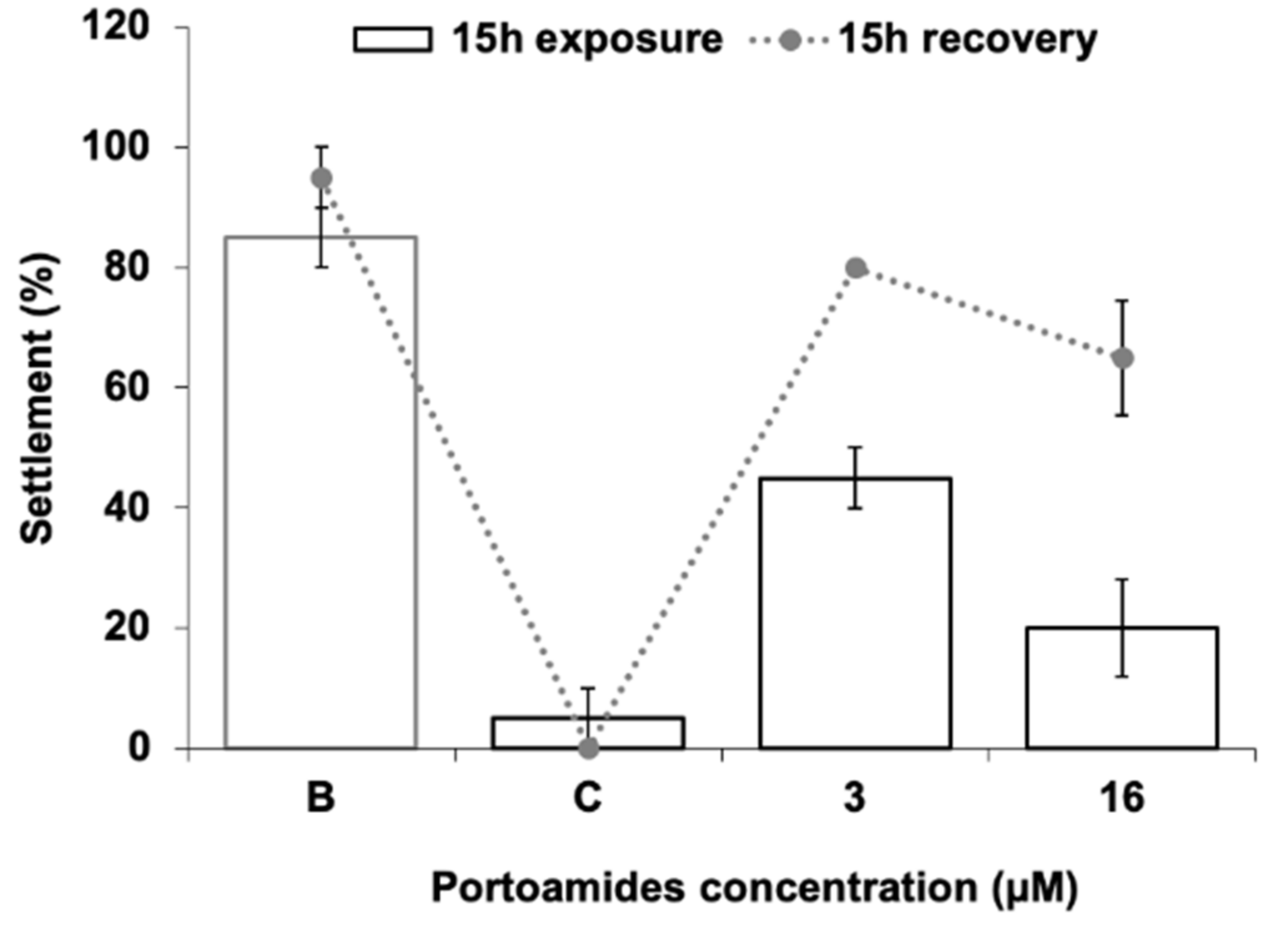
2.1. Mytilus galloprovincialis Larvae Antisettlement Activity and Recovery
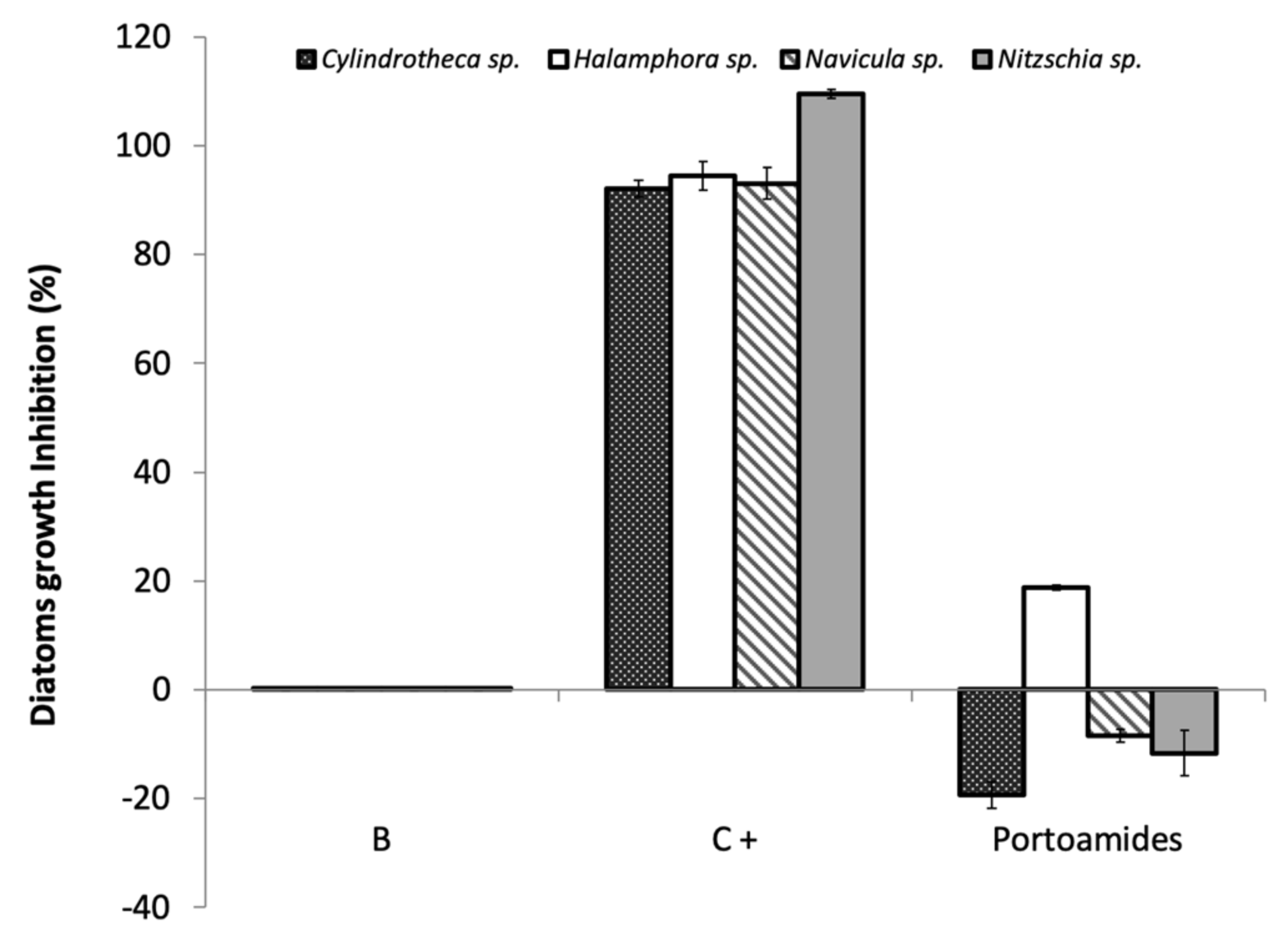
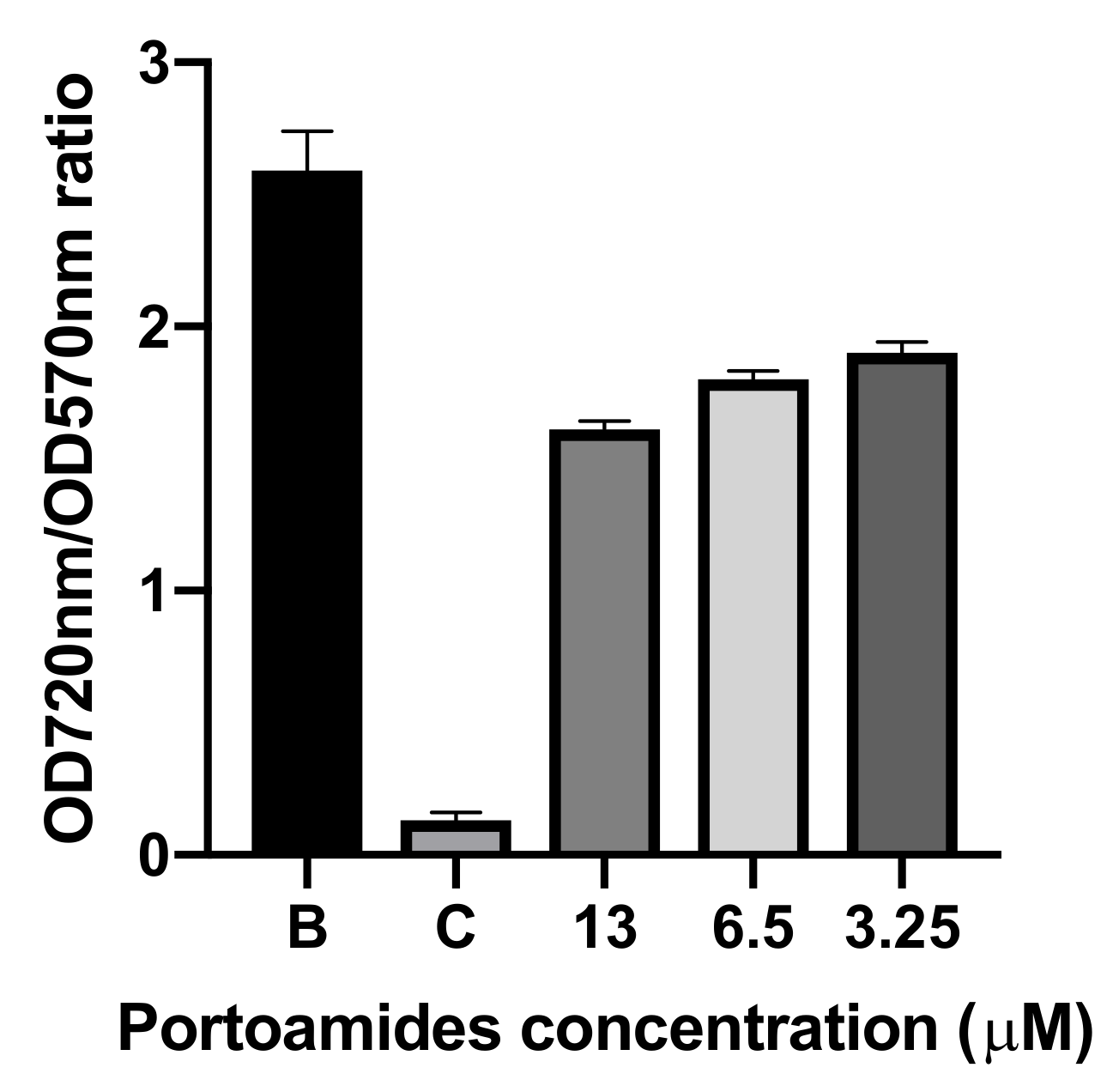
2.2. Antibacterial and Antimicroalgal Activities
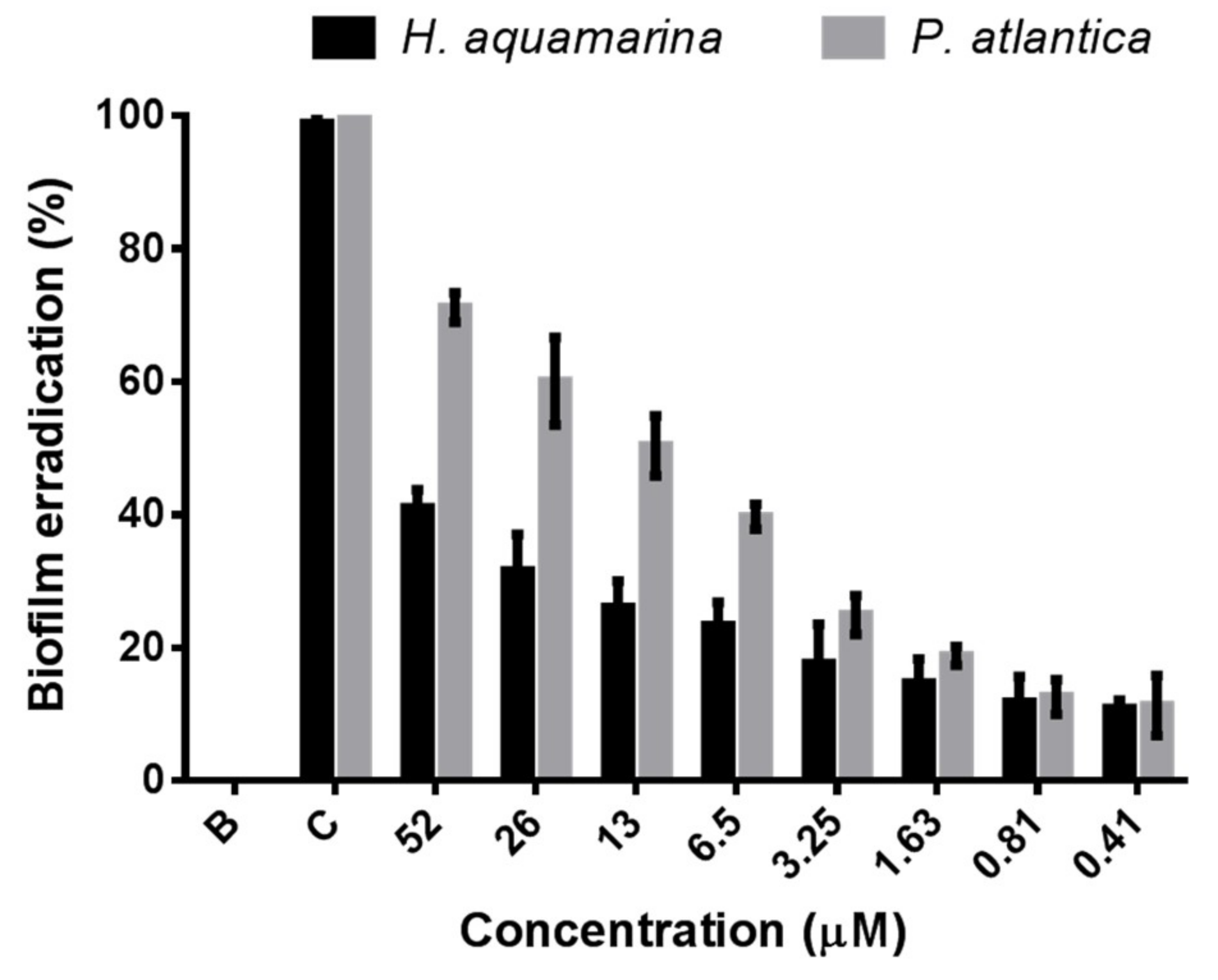
2.3. Antibiofilm Activity
2.4. Quorum-Sensing Inhibition (QSI) Activity
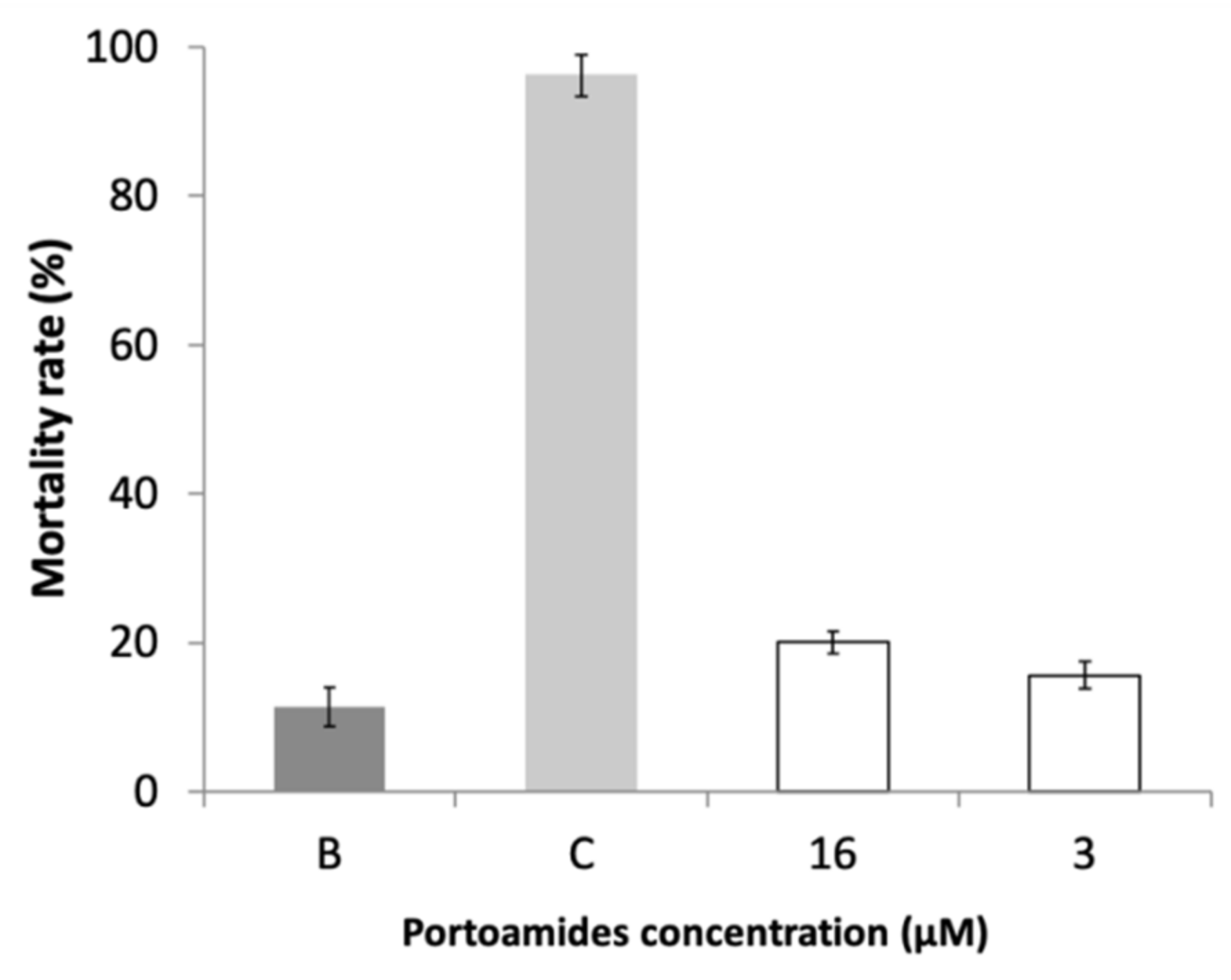
2.5. Ecotoxicity Assessment
2.6. Mode of Action Assessment
3. Materials and Methods
3.1. Portoamides Isolation and Purification
3.2. Antifouling Bioassays
3.2.1. Mussel Larvae Antisettlement Bioassays
3.2.2. Mussel Larvae Settlement Recovery Bioassays
3.2.3. Antibacterial Bioassays
3.2.4. Antimicroalgal Bioassays
3.2.5. Biofilm Inhibition Assay
3.2.6. Quorum-Sensing Inhibition Assay
Disk-Diffusion Assay
Quorum-Sensing Inhibition (QSI) Quantitative Assay (Dose–Response Violacein Inhibition Assay)
3.3. Ecotoxicity Bioassay
3.4. Antifouling Mode of Action Towards Mytilus Larvae
3.4.1. Sample Preparation
3.4.2. LC-MS/MS Analysis
3.4.3. Protein Identification and Quantification
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013, 15, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mo. Bio. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.Y.; Lau, S.C.K.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine Biofilms as Mediators of Colonization by Marine Macroorganisms: Implications for Antifouling and Aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Ann. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Mieszkin, S.; Callow, M.E.; Callow, J. Interactions between microbial biofilms and marine fouling algae: A mini review. Biofouling 2013, 29, 1097–1113. [Google Scholar] [CrossRef]
- Schultz, M.P.; Swain, G.W. The influence of biofilms on skin friction drag. Biofouling 2000, 15, 129–139. [Google Scholar] [CrossRef]
- Schultz, M.P. Effects of coating roughness and biofouling on ship resistance and powering. Biofouling 2007, 23, 331–341. [Google Scholar] [CrossRef]
- Costas, E.; Gonzalez, R.; López-Rodas, V.; Emma Huertas, I. Mutation of Microalgae from Antifouling Sensitivity to Antifouling Resistance Allows Phytoplankton Dispersal through Ships’ Biofouling. Biological. Invasions 2013, 15, 1739–1750. [Google Scholar] [CrossRef]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology―Past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R. Marine biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5, 126. [Google Scholar] [CrossRef]
- Chambers, L.D; Wharton, J.A.; Wood, R.J.K.; Walsh, F.C.; Stokes, K.R. Techniques for the measurement of natural product incorporation into an antifouling coating. Prog. Org. Coat. 2014, 77, 473–484. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Lewis, J.A.; Johnston, E.L. Antifouling strategies: History and regulation, ecological impacts and mitigation. Mar. Pollut. Bull. 2011, 62, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Callow, J.A.; Callow, M.E. Trends in the development of environmentally friendly fouling resistant marine coatings. Nat. Commun. 2011, 2, 244. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.-Y.; Li, Z.; Xu, Y.; Li, Y.; Fusetani, N. Mini-review: Marine natural products and their synthetic analogs as antifouling compounds: 2009–2014. Biofouling 2015, 31, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.; Wu, Z.H.; Wang, Y.; Wang, C.Y.; Xu, Y. Mini-Review: Antifouling natural products from marine microorganisms and their synthetic analogs. Mar. Drugs 2017, 15, 266. [Google Scholar] [CrossRef]
- Dahms, H.U.; Ying, X.; Pfeiffer, C. Antifouling potential of cyanobacteria: A mini-review. Biofouling 2006, 22, 317–327. [Google Scholar] [CrossRef]
- Almeida, J.R.; Vasconcelos, V. Natural antifouling compounds: Effectiveness in preventing invertebrate settlement and adhesion. Biotechnol. Adv. 2015, 33, 343–357. [Google Scholar] [CrossRef]
- Fusetani, N. Biofouling and antifouling. Nat. Prod. Rep. 2004, 21, 94–104. [Google Scholar] [CrossRef]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–441. [Google Scholar] [CrossRef]
- Chen, L.; Qian, P.-Y. Review on molecular mechanisms of antifouling compounds: An update since 2012. Mar. Drugs 2017, 15, 264. [Google Scholar] [CrossRef] [PubMed]
- Dahms, H.U.; Dobretsov, S. Antifouling Compounds from Marine Macroalgae. Mar. Drugs 2017, 15, 265. [Google Scholar] [CrossRef] [PubMed]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria―A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Qian, P.Y.; Chen, L.; Xu, Y. Mini-Review: Molecular Mechanisms of Antifouling Compounds. Biofouling 2013, 29, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Leao, P.N.; Pereira, A.R.; Liu, W.T.; Ng, J.; Pevzner, P.A.; Dorrestein, P.C.; Konig, G.M.; Vasconcelos, V.M.; Gerwick, W.H. Synergistic allelochemicals from a freshwater cyanobacterium. Proc. Natl. Acad. Sci. USA 2010, 107, 11183–11188. [Google Scholar] [CrossRef] [PubMed]
- Dias, F.; Antunes, J.T.; Ribeiro, T.; Azevedo, J.; Vasconcelos, V.; Leao, P.N. Cyanobacterial Allelochemicals But Not Cyanobacterial Cells Markedly Reduce Microbial Community Diversity. Front. Microbiol. 2017, 8, 1495. [Google Scholar] [CrossRef] [PubMed]
- Diers, J.A.; Pennaka, H.K.; Peng, J.; Bowling, J.J.; Duke, S.O.; Hamann, M.T. Structural Activity Relationship Studies of Zebra Mussel Antifouling and Antimicrobial Agents from Verongid Sponges. J. Nat. Prod. 2004, 67, 2117–2120. [Google Scholar] [CrossRef] [PubMed]
- Diers, J.A.; Bowling, J.J.; Duke, S.O.; Wahyuono, S.; Kelly, M.; Hamann, M.T. Zebra Mussel Antifouling Activity of the Marine Natural Product Aaptamine and Analogs. Mar. Biotechnol. 2006, 8, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Ortlepp, S.; Pedpradap, S.; Dobretsov, S.; Proksch, P. Antifouling activity of sponge-derived polybrominated diphenyl ethers and synthetic analogues. Biofouling 2008, 24, 201–208. [Google Scholar] [CrossRef]
- Almeida, J.R.; Correia-da-Silva, M.; Sousa, E.; Antunes, J.; Pinto, M.; Vasconcelos, V.; Cunha, I. Antifouling potential of Nature-inspired sulfated compounds. Sci. Rep. 2017, 7, 42424. [Google Scholar] [CrossRef]
- Almeida, J.R.; Moreira, J.; Pereira, D.; Pereira, S.; Antunes, J.; Palmeira, A.; Vasconcelos, V.; Pinto, M.; Correira-da-Siva, M.; Cidade, H. Potential of synthetic chalcone derivatives to prevent marine biofouling. Sci. Total Environ. 2018, 643, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Goh, B.P.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.; Teo, S.L. Natural antifoulants from the marine cyanobacterium Lyngbya majuscula. Biofouling 2010, 2, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Wong, H.F.; Hutchinson, N.; Lee, S.C.; Chan, B.K.K.; Williams, G.A. Chemistry and biology of macula- lactone A from the marine cyanobacterium Kyrtuthrix maculans. Phytochem. Rev. 2004, 3, 381–400. [Google Scholar] [CrossRef]
- Dobretsov, S.; Abed, R.M.M.; Teplitski, M. Mini-review: Inhibition of biofouling by marine microorganisms. Biofouling 2013, 29, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Carl, C.; Poole, A.J.; Vucko, M.J.; Williams, M.R.; Whalan, S.; de Nys, R. Optimising settlement assays of pediveligers and plantigrades of Mytilus galloprovincialis. Biofouling 2011, 27, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.; Leikoski, N.; Wahlsten, M.; Azevedo, J.; Antunes, J.; Jokela, J.; Sivonen, K.; Vasconcelos, V.; Fewer, D.P.; Leão, P. Sphaerocyclamide a prenylated Cyanobactin from the cyanobacterium Sphaerospermopsis sp. LEGE 00249. Sci. Rep. 2018, 8, 14537. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, K.Ø.; Cervin, G.; Trepos, R.; Petitbois, J.; Haug, T.; Hansen, E.; Andersen, J.H.; Pavia, H.; Hellio, C.; Svenson, J. The Bromotyrosine Derivative Ianthelline Isolated from the Arctic Marine Sponge Stryphnus fortis Inhibits Marine Micro- and Macrobiofouling. Mar. Biotechnol. 2014, 16, 684–694. [Google Scholar] [CrossRef]
- Moodie, L.W.K.; Trepos, R.; Cervin, G.; Larsen, L.; Larsen, D.S.; Pavia, H.; Hellio, C.; Cahill, P.; Svenson, J. Probing the structure-activity relationship of the natural antifouling agent polygodial against both micro and macrofoulers by semisynthetic modification. J. Nat. Prod. 2017, 80, 515–525. [Google Scholar] [CrossRef]
- Maréchal, J.P.; Culioli, G.; Hellio, C.; Thomas-Guyon, H.; Callow, M.E.; Clare, A.S.; Ortalo-Magne, A. Seasonal variation in antifouling activity of crude extracts of the brown alga Bifurcaria bifurcata (Cystoseiraceae) against cyprids of Balanus amphitrite and the marine bacteria Cobetia marina and Pseudoalteromonas haloplanktis. J. Exp. Mar. Biol. Ecol. 2004, 313, 47–62. [Google Scholar] [CrossRef]
- Tsoukatou, M.; Marechal, J.P.; Hellio, C.; Novakovic, I.; Tufegdzie, S.; Sladic, D.; Gasic, M.J.; Clare, A.S.; Vagias, C.; Roussis, V. Evaluation of the activity of the sponge metabolites avarol and avarone and their synthetic derivatives against fouling micro- and macroorganisms. Molecules 2012, 12, 1022–1034. [Google Scholar] [CrossRef]
- Viano, Y.; Bonhomme, D.; Camps, M.; Briand, J.; Ortalo-Magne, A.; Blache, Y.; Piovetti, L.; Culioli, G. Diterpenoids from the Mediterranean Brown Alga Dictyota sp. Evaluated as Antifouling Substances against a Marine Bacterial Biofilm. J. Nat. Prod. 2009, 72, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Molino, P.J.; Wetherbee, R. Mini-review: The biology of biofouling diatoms and their role in the development of microbial slimes. Biofouling 2008, 24, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Inbakandan, D.P.; Sriyutha Murthy, R.; Khan, S.J. 16S rDNA Sequence Analysis of Culturable Marine Biofilm Forming Bacteria from a Ship’s Hull. Biofouling 2010, 26, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, I.; Bachmann, R.T.; Edyvean, R.G.J. Type 2 quorum sensing monitoring, inhibition and biofilmformation in marine microrganisms. Curr. Microbiol. 2014, 68, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Melander, C.; Moeller, P.D.R.; Ballard, T.E.; Richards, J.J.; Huigens III, R.W.; Cavanagh, J. Evaluation of dihydrooroidin as an antifouling additive in marine paint. Int. Biodeter. Biodegr. 2009, 63, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Bernbom, N.; Ng, Y.Y.; Kjelleberg, S.; Harder, T.; Gram, L. Marine bacteria from danish coastal waters show antifouling activity against the marine fouling bacterium Pseudoalteromonas sp. strain S91 and zoospores of the green alga Ulva australis independent of bacteriocidal activity. Appl. Environ. Microbiol. 2011, 77, 8557–8567. [Google Scholar] [CrossRef]
- Bernbom, N.; Ng, Y.Y.; Olsen, S.M.; Gram, L. Pseudoalteromonas spp. serve as initial bacterial attractants in mesocosms of coastal waters but have subsequent antifouling capacity in mesocosms and when embedded in paint. Appl. Environ. Microb. 2013, 79, 6885–6893. [Google Scholar] [CrossRef]
- Brian-Jaisson, F.; Ortalo-Magné, A.; Guentas-Dombrowsky, L.; Armougom, F.; Blache, Y.; Molmeret, M. Identification of Bacterial Strains Isolated from the Mediterranean Sea Exhibiting Different Abilities of Biofilm Formation. Microb. Ecol. 2004, 68, 94–110. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar]
- Dusane, D.H.; Damare, S.R.; Nancharaiah, Y.V.; Ramaiah, N.; Venugopalan, V.P.; Kumar, A.R.; Zinjarde, S.S. Disruption of microbial biofilms by an extracellular protein isolated from epibiotic tropical marine strain of Bacillus licheniformis. PLoS ONE 2013, 8, e64501. [Google Scholar] [CrossRef]
- Richards, J.J.; Melander, C. Controlling Bacterial Biofilms. ChemBioChem 2009, 10, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Salta, M.; Wharton, J.A.; Dennington, S.P.; Stoodley, P.; Stokes, KR. Anti-biofilm performance of three natural products against initial bacterial attachment. Int. J. Mol. Sci. 2013, 14, 21757–21780. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, E.; Boyd, K.G.; Burgess, J.G. Prevention of marine biofouling using natural compounds from marine organisms. Biotech. Ann. Rev. 2000, 6, 221–241. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Paul, C. Mini-Review: Quorum Sensing in the Marine Environment and Its Relationship to Biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E.; Jacobsen, L.B.; Nichols, D.E.; McLaughlin, J.L. Brine shrimp: A convenient general bioassay for active plant constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.; Lemos, F.; Preto, M.; Azevedo, J.; Sousa, M.L.; Leao, P.N.; Vitorino, R.; Vasconcelos, V.; Urbatzka, R. Cytotoxicity of portoamides in human cancer cells and analysis of the molecular mechanisms of action. PLoS ONE 2017, 12, e188817. [Google Scholar] [CrossRef] [PubMed]
- DeMartini, D.G.; Errico, J.M.; Sjoestroem, S.; Fenster, A.; Waite, J.H. A cohort of new adhesive proteins identified from transcriptomic analysis of mussel foot glands. J. R. Soc. Interface 2017, 14, 131. [Google Scholar] [CrossRef]
- Dral, A.D.G. Movements and Co-ordination of the Latero-frontal Cilia of the Gill Filaments of Mytilus. Nature 1966, 210, 1170–1171. [Google Scholar] [CrossRef]
- Fields, P.A.; Zuzow, M.J.; Tomanek, L. Proteomic responses of blue mussel (Mytilus) congeners to temperature acclimation. J. Exp. Biol. 2012, 215, 1106–1116. [Google Scholar] [CrossRef]
- Sepčić, K.; Turk, T. 3-Alkylpyridinium Compounds as Potential Non-Toxic Antifouling Agents. In Antifouling Compounds. Marine Molecular Biotechnology; Fusetani, N., Clare, A.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Piazza, V.; Dragić, I.; Sepčić, K.; Faimali, M.; Garaventa, F.; Turk, T.; Berne, S. Antifouling activity of synthetic alkylpyridinium polymers using the barnacle model. Mar. Drugs 2014, 12, 1959–1976. [Google Scholar] [CrossRef]
- McLean, R.J.; Pierson, L.S., III; Fuqua, C. A Simple screening protocol for the identification of quorum signal antagonists. J. Microbiol. Methods 2004, 58, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, D.; Grossmann, G.; Sequin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4. [Google Scholar] [CrossRef] [PubMed]
- Gemiarto, A.; Ninyo, N.N.; Lee, W.L.; Logis, J.; Fatima, A.; Chan, E.W.C.; Lim, C.S.Y. Isoprenyl caffeate, a major compound in manuka propolis, is a quorum-sensing inhibitor in Chromobacterium violaceum. Antonie Van Leeuwenhoek. 2015, 108, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | EC30 (μM) | ||
|---|---|---|---|
| Portoamides | Cobetia marina | Halomonas aquamarina | Pseudoalteromonas atlantica |
| 14.56 (95% CI: 10.21–23.75) | 14.83 (95% CI: 10.41–24.66) | 13.59 (95% CI: 9.67–21.45) | |
| Mean of log10 (x + 1)-Transformed Values | p-Value | Accession | Protein Name | Metabolic Process | ||
|---|---|---|---|---|---|---|
| Control | Portoamides | |||||
| Energy Metabolism | 2.465 | 0.304 | 0.004 | XP_009028970.1 | ATP synthase subunit d, mitochondrial | ATP synthesis |
| 3.742 | 3.335 | 0.011 | XP_021356377.1 | ATP synthase subunit beta mitochondrial | ||
| 3.783 | 3.494 | 0.012 | ABJ51956.1 | H+ ATPase a subunit mitochondrial | ||
| 3.424 | 3.059 | 0.018 | XP_009064140.1 | ATP synthase subunit gamma | ||
| 3.455 | 2.890 | 0.018 | AFI56365.1 | Isocitrate dehydrogenase | Tricarboxylic Acid cycle | |
| 3.401 | 3.023 | 0.027 | AAF27650.1 | Malate dehydrogenase | ||
| 2.825 | 2.405 | 0.031 | XP_022314664.1 | Glutamate dehydrogenase mitochondrial | ||
| Structural | 4.116 | 3.771 | 0.050 | XP_022317649.1 | Myosin heavy chain striated muscle | Muscle structural protein; contractionmovement |
| 3.855 | 3.469 | 0.046 | CAB64663.1 | Pedal retractor muscle myosin heavy chain | ||
| 4.386 | 3.882 | 0.050 | XP_014664190.1 | Tubulin beta chain | Cytoskeleton constituent | |
| 3.857 | 3.379 | 0.038 | XP_021370666.1 | Tubulin alpha-1A chain | ||
| 3.166 | 2.623 | 0.035 | XP_022332848.1 | Cilia- and flagella-associated protein | ||
| 3.756 | 3.064 | 0.021 | KFB49247.1 | Myosin heavy chain | ||
| 3.149 | 2.888 | 0.018 | NP_001292292.1 | Tubulin beta chain | ||
| 3.497 | 2.929 | 0.015 | XP_009029528.1 | Tubulin beta chain | ||
| 3.269 | 2.819 | 0.004 | XP_021350592.1 | Myosin heavy chain | ||
| 2.660 | 1.971 | 0.024 | XP_011416098.1 | Tektin-4 | Structural, cell motility | |
| 2.440 | 1.054 | 0.027 | XP_021370563.1 | Radial spoke head protein 4 | ||
| 2.343 | 1.106 | 0.031 | XP_021363337.1 | Radial spoke head protein 9 | ||
| 3.31471 | 2.727 | 0.031 | XP_011450983.1 | Tektin-3 | ||
| 1.974 | 0 | 0.037 | XP_021375365.1 | Collagen, type VI, alpha 3 | Extracellular matrix organization | |
| Gene Transcription/Translation | 2.990 | 2.673 | 0.049 | XP_021341263.1 | Dolichyl-diphosphooligosaccharide protein glycosyltransferase | Protein modification, PTMs |
| 4.011 | 3.579 | 0.040 | AKS48144.1 | Arginine kinase | ||
| 3.020 | 2.360 | 0.016 | XP_021367901.1 | Dolichyl-diphosphooligosaccharide protein glycosyltransferase 48 kDa subunit | ||
| 3.854 | 3.596 | 0.048 | XP_018019106.1 | 14-3-3 protein | Chaperone, protein activity stabilization/regulation | |
| 3.474 | 2.958 | 0.044 | XP_011444269.1 | Peptidyl-prolyl cis-trans isomerase | ||
| 3.578 | 3.200 | 0.041 | CAJ85741.1 | Heat shock protein 90 | ||
| 2.515 | 2.191 | 0.038 | XP_011446200.1 | Protein disulfide-isomerase A6 | ||
| 2.431 | 2.025 | 0.019 | Q6WV66.3 | Histone H2A | Chromatin function, gene transcription | |
| 2.493 | 2.080 | 0.027 | XP_021377317.1 | Histone H2A | ||
| 3.618 | 3.309 | 0.035 | XP_021363857.1 | Leucine-rich repeat flightless-interacting protein 2 | ||
| 4.961 | 4.649 | 0.042 | XP_014590042.1 | Histone H4 | ||
| 1.968 | 0 | 0.024 | XP_012945445.1 | Heterogeneous nuclear ribonucleoprotein 87F | Ribossome constituent/function, protein translation | |
| 2.738 | 2.356 | 0.029 | XP_021369622.1 | 60S ribosomal protein L7a | ||
| 3.071 | 2.707 | 0.036 | XP_022290789.1 | 40S ribosomal protein S4 | ||
| 3.332 | 2.965 | 0.041 | XP_011438308.1 | 40S ribosomal protein S3 | ||
| 3.050 | 2.675 | 0.042 | XP_011441368.1 | 40S ribosomal protein S13 | ||
| 3.250 | 2.727 | 0.043 | XP_021370373.1 | 40S ribosomal protein S25 | ||
| 3.292 | 3.038 | 0.048 | XP_013068480.1 | 60S ribosomal protein L11 | ||
| Transport | 1.482 | 0 | 0.043 | XP_021354511.1 | Clathrin heavy chain 1 | Vesicle mediated transport |
| 2.565 | 2.174 | 0.011 | XP_022317867.1 | Annexin B9 | ||
| 3.565 | 3.113 | 0.038 | SCN46548.1 | ADP, ATP carrier protein | Transporters, carrier proteins | |
| 3.667 | 3.260 | 0.046 | ADI56517.1 | Voltage-dependent anion channel | ||
| Other Processes | 2.595 | 2.247 | 0.0468 | XP_022343934.1 | 6-phosphogluconate dehydrogenase decarboxylating | Synthesis of NADPH and 5-carbon sugar precursor |
| 3.284 | 2.942 | 0.042 | AFQ35984.1 | Glutathione S-transferase sigma 2 | Detoxification, response to oxidative stress | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antunes, J.; Pereira, S.; Ribeiro, T.; Plowman, J.E.; Thomas, A.; Clerens, S.; Campos, A.; Vasconcelos, V.; Almeida, J.R. A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides. Mar. Drugs 2019, 17, 111. https://doi.org/10.3390/md17020111
Antunes J, Pereira S, Ribeiro T, Plowman JE, Thomas A, Clerens S, Campos A, Vasconcelos V, Almeida JR. A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides. Marine Drugs. 2019; 17(2):111. https://doi.org/10.3390/md17020111
Chicago/Turabian StyleAntunes, Jorge, Sandra Pereira, Tiago Ribeiro, Jeffrey E. Plowman, Ancy Thomas, Stefan Clerens, Alexandre Campos, Vitor Vasconcelos, and Joana R. Almeida. 2019. "A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides" Marine Drugs 17, no. 2: 111. https://doi.org/10.3390/md17020111
APA StyleAntunes, J., Pereira, S., Ribeiro, T., Plowman, J. E., Thomas, A., Clerens, S., Campos, A., Vasconcelos, V., & Almeida, J. R. (2019). A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides. Marine Drugs, 17(2), 111. https://doi.org/10.3390/md17020111