1. Introduction
Microcystins (MCs) are typical compounds produced by cyanobacteria, such as
Microcystis,
Anabaena, and
Planktothrix [
1]. They are cyclic heptapeptides showing potent hepatotoxicity and tumor-promoting activity [
1]. In the environment, there are many bacteria which work to degrade such hazardous and harmful compounds. The first MC-degrading bacterium was isolated and identified as a
Sphingomonas strain (ACM-3962) in 1994 [
2]. Similar bacteria capable of degrading MC were reported by Dziga et al. [
3]. As per their review [
3], many MC-degrading microorganisms have been found and identified, and the corresponding genetic aspects with respect to MC degradation have been studied. However, in related published papers, no substrates other than MCs have been applied [
4]. The purpose of the present study is to elucidate the inherent function and role of MC-degrading microorganisms in the aquatic environment.
Strain B-9, isolated from Lake Tsukui, Japan, exhibits degradation activity against MCs [
1]. This strain belongs to the genus
Sphingosinecella sp., and, based on the 16S rDNA sequence (GenBank accession no. AB084247), has 99% similarity to
Sphingosinecella microcystinivorans strain Y2, a type of MC-degrading bacteria [
5]. The
Sphingomonas sp. strain ACM-3962 [
2] was the first strain reported to degrade MCs. The cloning and molecular characterization of four genes from strain ACM-3962 revealed the presence of three hydrolytic enzymes (MlrA, MlrB, and MlrC), together with a putative oligopeptide transporter (MlrD) [
6,
7]. The three hydrolytic enzymes were putatively characterized as metalloproteases (MlrA and MlrC) or serine proteases (MlrB). The microcystinase MlrA catalyzes the initial ring opening of microcystin-LR (MC-LR) at the (
2S,
3S,
8S,
9S)-3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4(
E),6(
E)-dienoic acid (Adda)-Arg peptide bond to give linearized MC-LR (Adda-Glu-Mdha-Ala-Leu-β-MeAsp-Arg). This further degrades to Adda-Glu-Mdha-Ala by MlrB, and the third enzyme, MlrC, hydrolyzes the tetrapeptide into smaller peptides and amino acids [
6,
7,
8]. In terms of advancements, recombinant MlrA and MlrC have been prepared, and the degradation scheme has been almost completely verified [
9,
10]. The use of typical protease inhibitors, such as ethylenediaminetetraacetic acid (EDTA) and 1,10-phenanthroline, results in the accumulation of linear MC-LR and the tetrapeptide, which allows for the classification of the enzymes MlrA and MlrC as metalloproteases, [
6,
7]. Meanwhile, phenylmethylsulfonyl fluoride (PMSF) characterizes MlrB as a possible serine protease [
6,
7].
We extended the area of application of strain B-9 for bioremediation and applied it to the secondary fungal metabolites of mycotoxins that may have mutagenic, carcinogenic, cytotoxic, and endocrine-disrupting effects. These substances frequently contaminate agricultural commodities despite efforts for prevention, so successful detoxification methods are needed. The application of microorganisms to degrade mycotoxins is a possible strategy that shows potential for example in food and feed processing [
11]; in antibiotics used in human and veterinary medicine (which can enter the environment via wastewater treatment plant effluents, hospitals, and processing plant effluents); in the application of agricultural waste and biosolids to fields; and in the case of leakage from waste-storage containers and landfills [
12,
13]. Such antibiotic pollution may facilitate the development and spread of antibiotic resistance [
4].
Strain B-9 degrades MC-LR, the most toxic of the MCs, within 24 h [
14,
15]. After the discovery of strain B-9, we advanced our research on the degradation of the following compounds: non-toxic cyanobacterial cyclic peptides that are structurally different from MCs [
16]; representative cyclic peptides (antibiotics) produced by bacteria [
17]; physiologically-active cardiovascular and neuropeptides [
18]; and the glucagon/vasoactive intestinal polypeptide (VIP) family of peptides [
19]. The aforementioned experiments [
17,
18,
19] confirmed that strain B-9 could degrade the tested peptides completely. During the application of strain B-9 to remove mycotoxins and drugs released in the aquatic environment, we obtained interesting results concerning the function of this strain. The purpose of this study was to demonstrate the additional hydrolytic enzymes (such as protease and/or esterase) of strain B-9.
3. Discussion
Strain B-9, which has 99% similarity to
Sphingosinicella microcystinivorans strain Y2 [
5], is Gram-negative and has, in several studies, shown promise for the degradation of MC-related compounds and nodularin [
14,
15]. Subsequently, we applied strain B-9 to other types of substrates, such as cyanobacterial peptides including depsipeptides [
16], and bacterial cyclic peptides including depsipeptides [
16], which are structurally different from the MCs and nodularin. Based on these results, the hydrolytic behavior using this strain is suggested as follows: (1) the reaction essentially occurs at a peptide bond in a cyclic peptide moiety to give a linearized peptide, which is more quickly hydrolyzed compared to their original ones; (2) strain B-9 primarily hydrolyzes an ester bond in a depsipeptide, in which the resulting peptides are further hydrolyzed; (3) a cyclic peptide is hydrolyzed at the acyclic part, and no further reaction occurs; and (4) the resulting linearized peptide is more quickly hydrolyzed compared to the cyclic one. In some cases, it is hard to detect the degraded peptides or amino acids due to rapid hydrolysis [
16].
To confirm these observations and to further investigate the hydrolytic activities of the strain, we extended our study to physiologically active peptides such as neuropeptides and cardiovascular peptides [
18]. The tested peptides were classified into two groups: (1) linear peptides, and (2) cyclic peptides with a loop formed by disulfide bond formation. The linearized peptides degraded faster than the loop-containing peptides because the loop formed by the disulfide bond was regarded as one of the degradation-resistant factors. Hydrolysis of the peptides occurred through the cleavage of various peptide bonds, and strain B-9 may bear similarities to the mammalian neutral endopeptidase (NEP) 24.11, a 94-kDa zinc metalloendoprotease widely distributed in humans and involved in the processing of peptide hormones due to its broad selectivity [
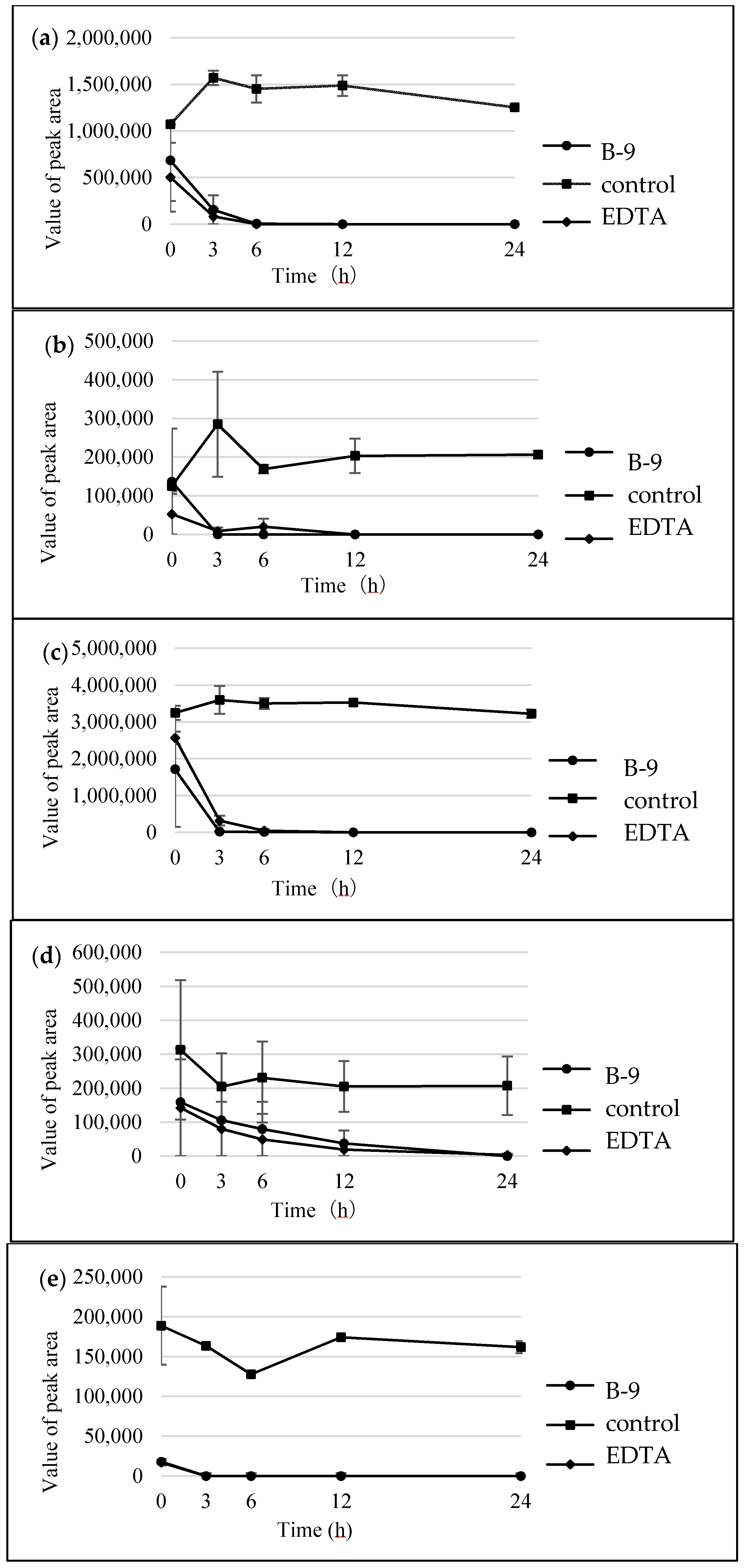
20]. In a separate study, we observed the degradation behavior of the linear peptides—the glucagon/VIP family peptides (3200–5000 Da)—by strain B-9 in the absence or presence of two protease inhibitors, EDTA and PMSF. Consequently, we confirmed that one of the B-9 proteases (presumed to be MlrB), which is not inhibited by EDTA, cleaved bioactive peptides in the manner of an endopeptidase similar to NEP. Another protease, which is not inhibited by PMSF, corresponded to MlrC and cleaved the resulting middle-sized peptides to smaller peptides or amino acids [
19].
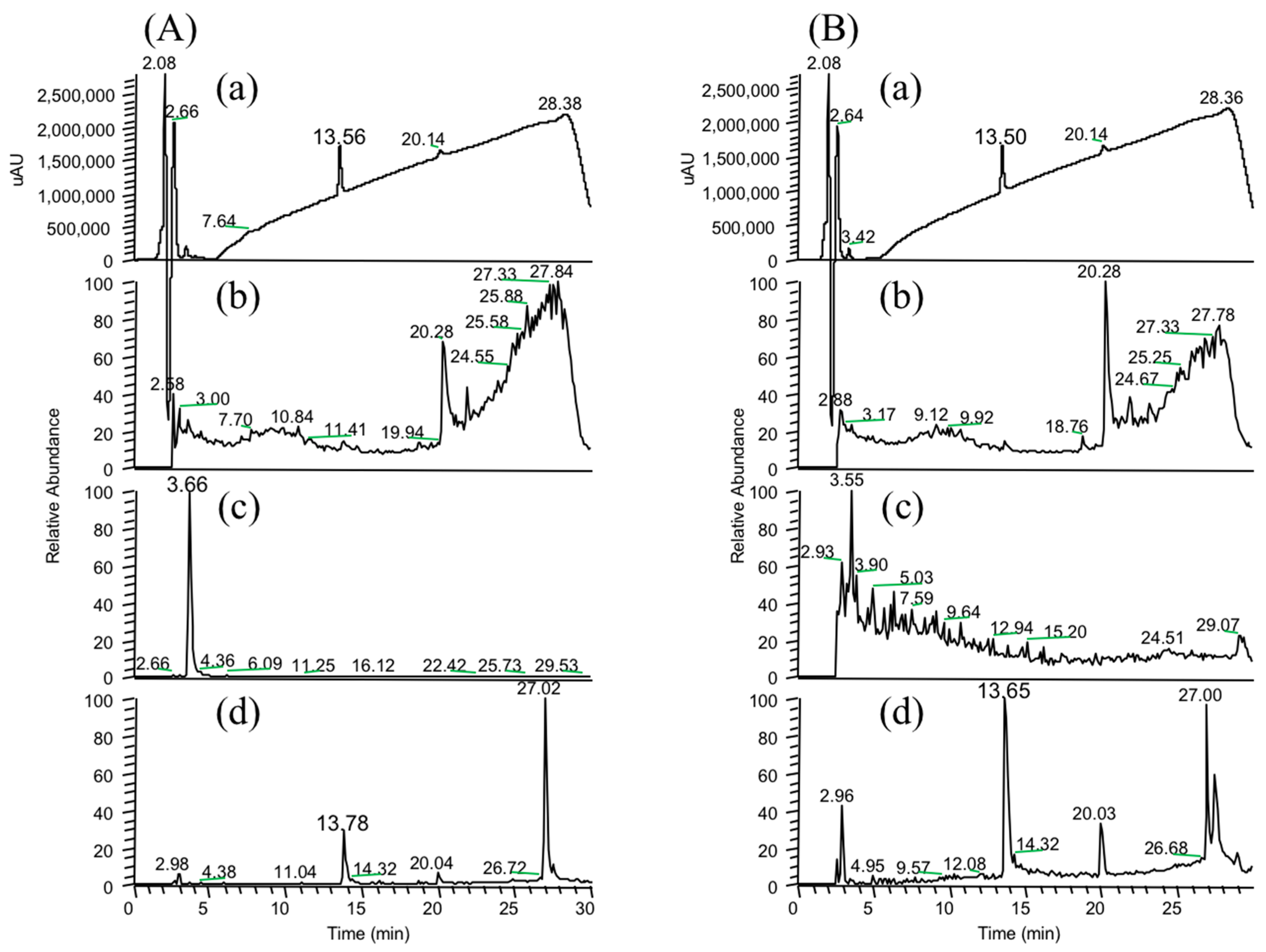
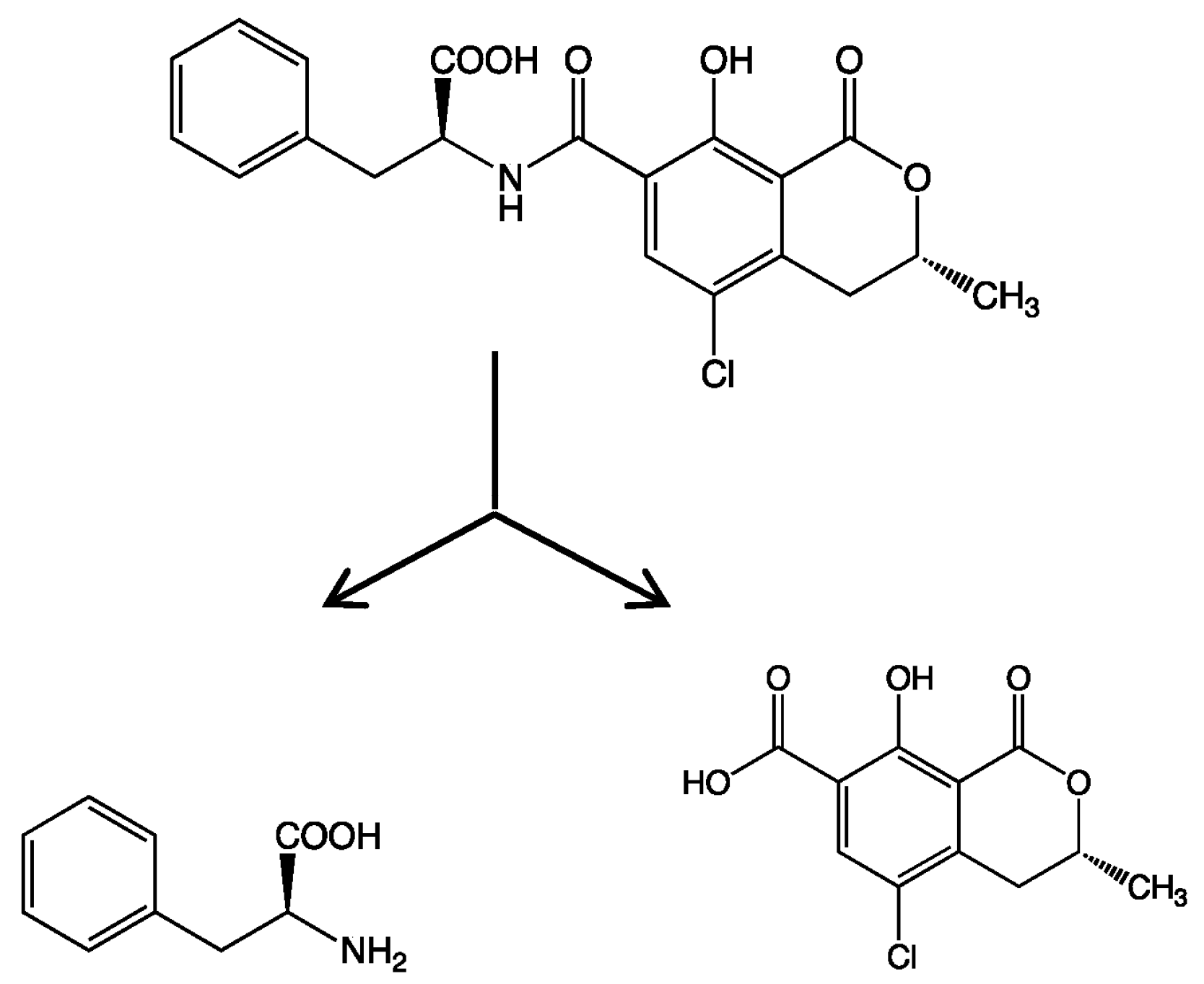
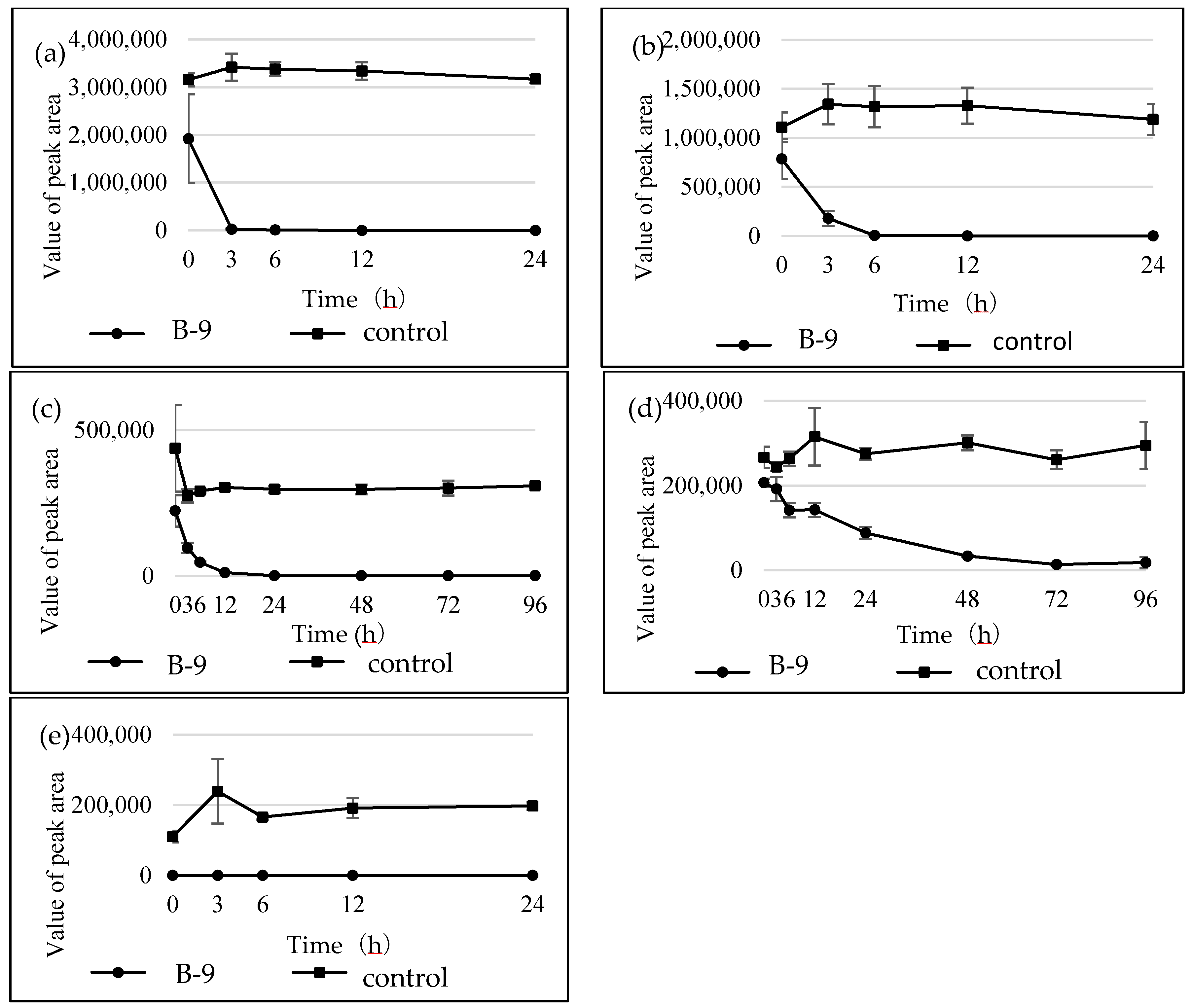
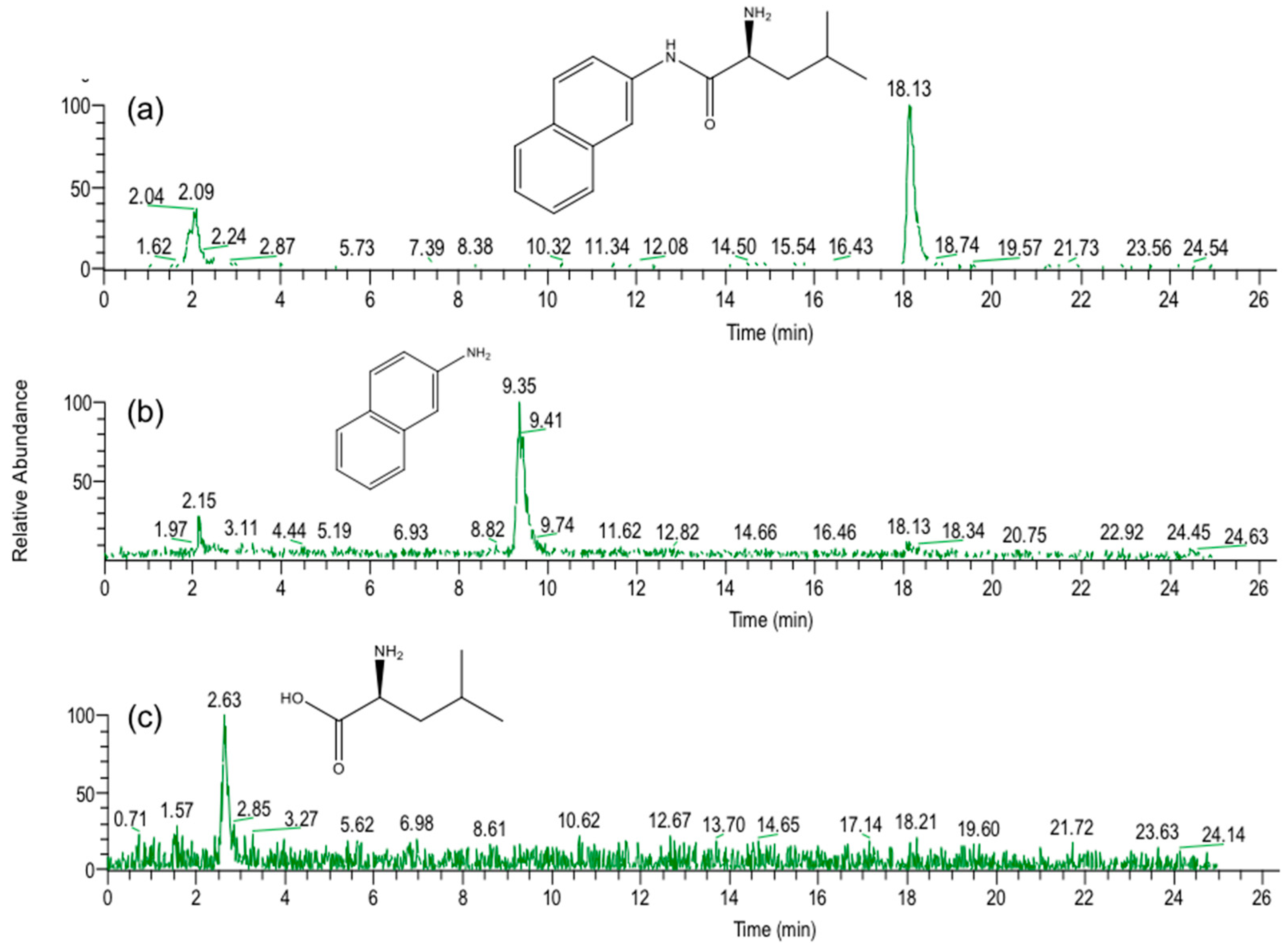
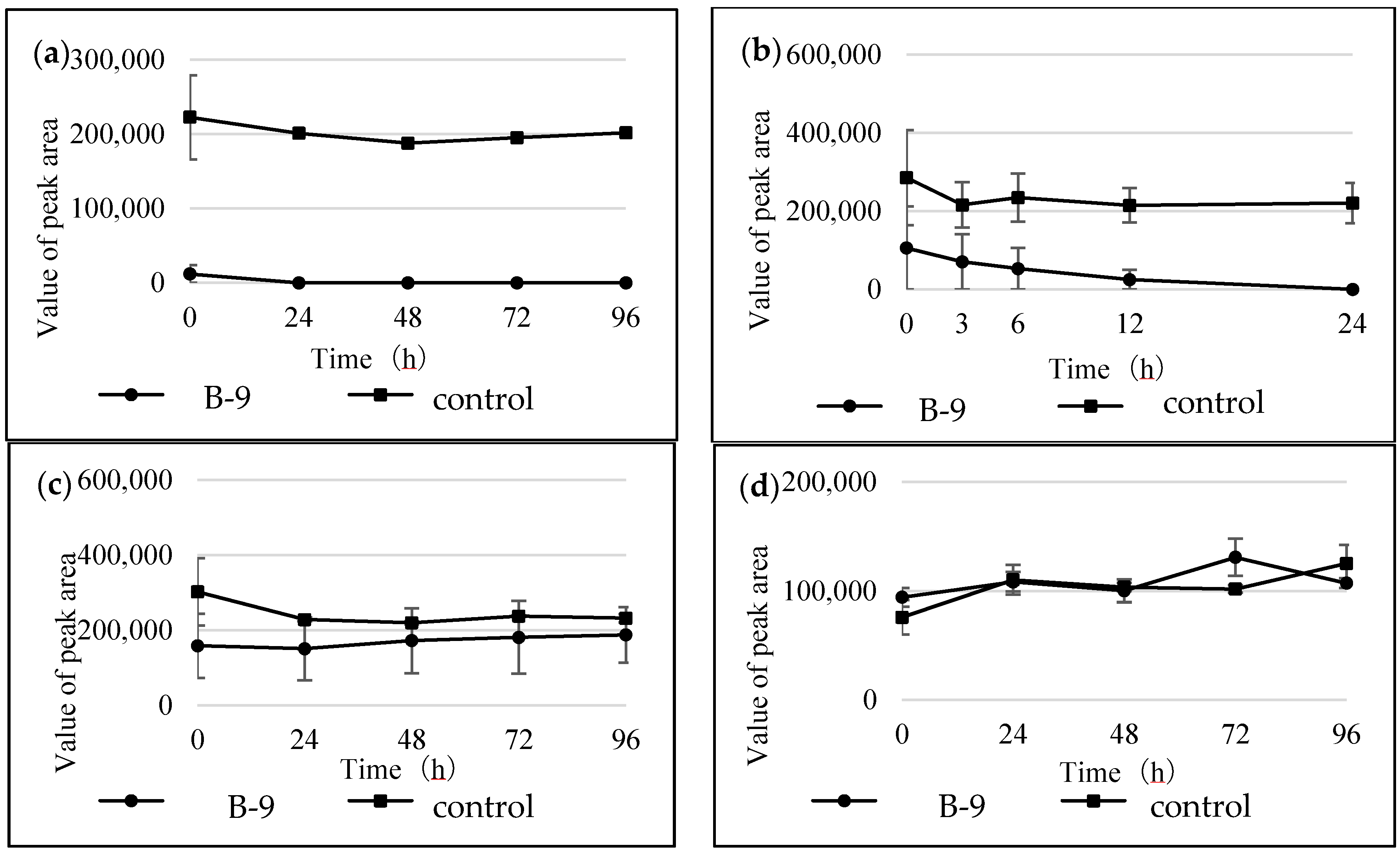
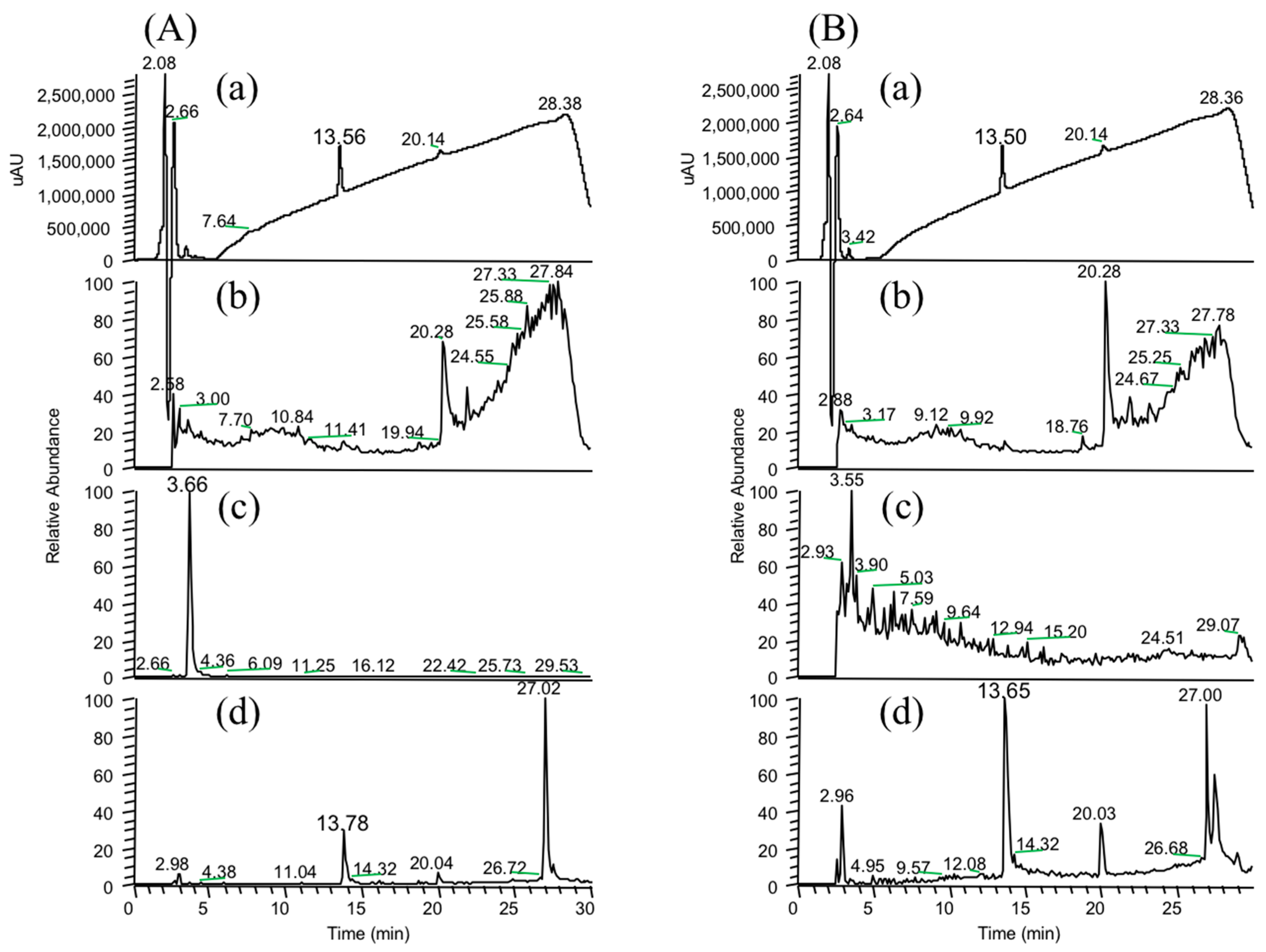
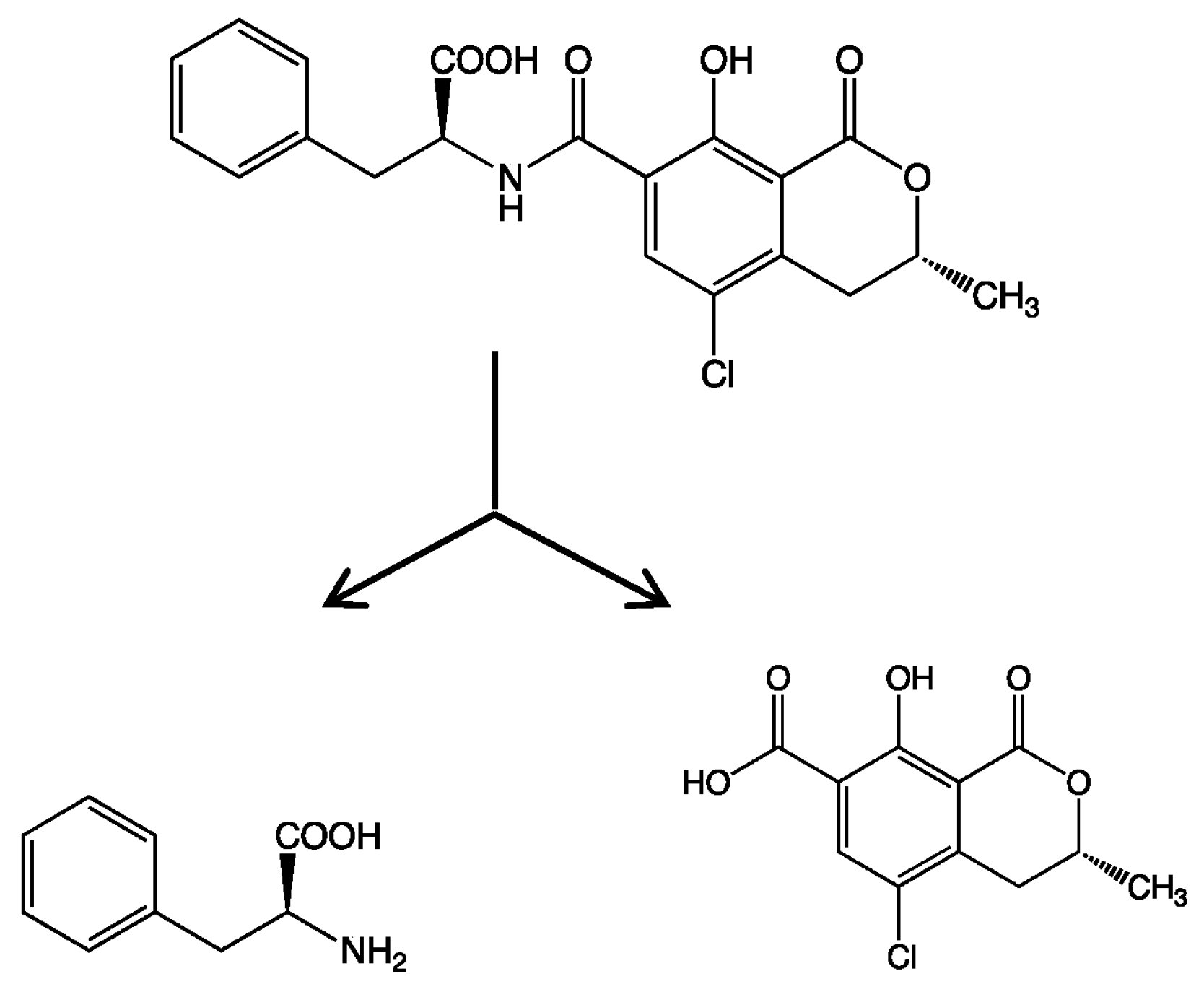
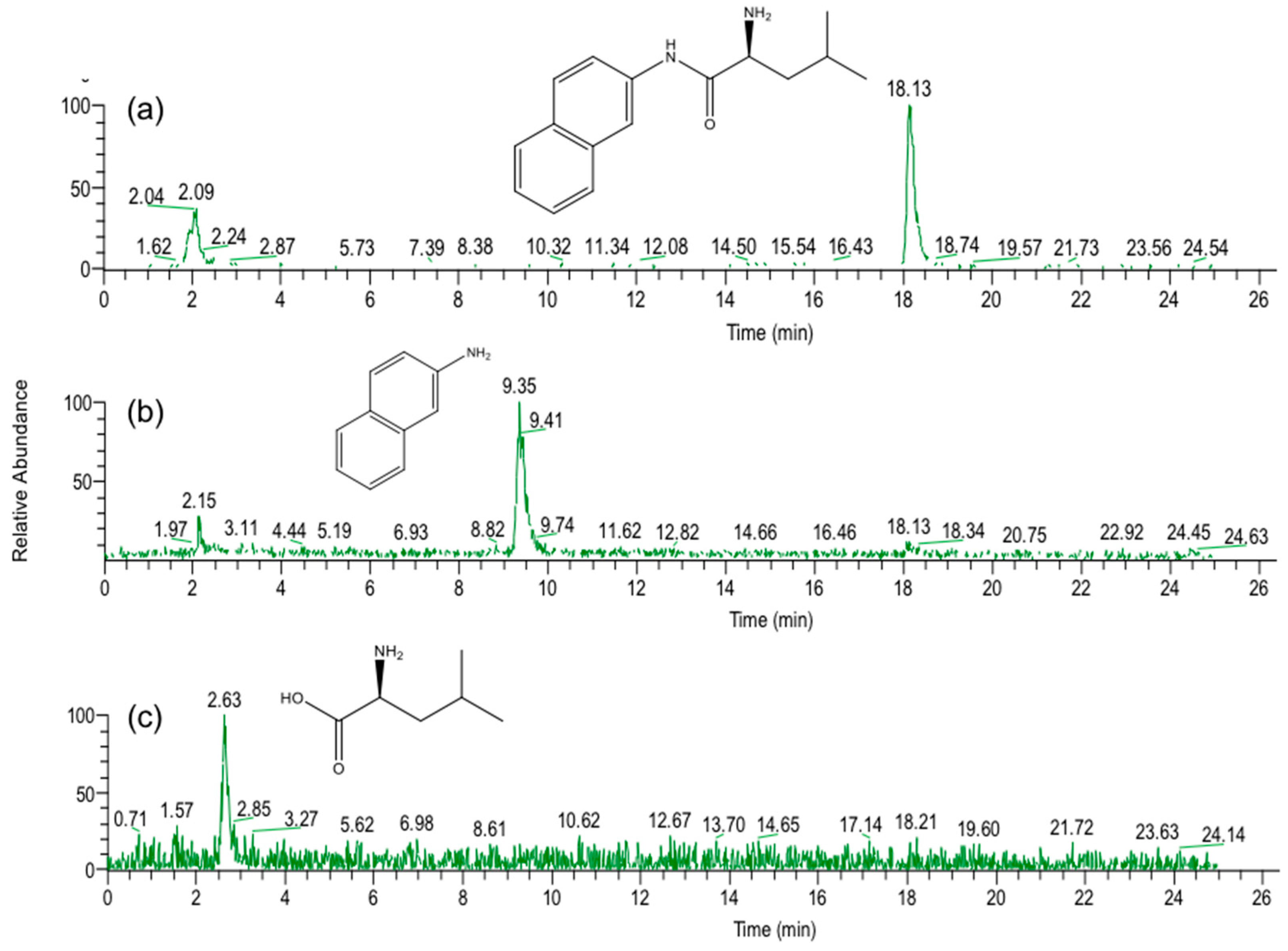
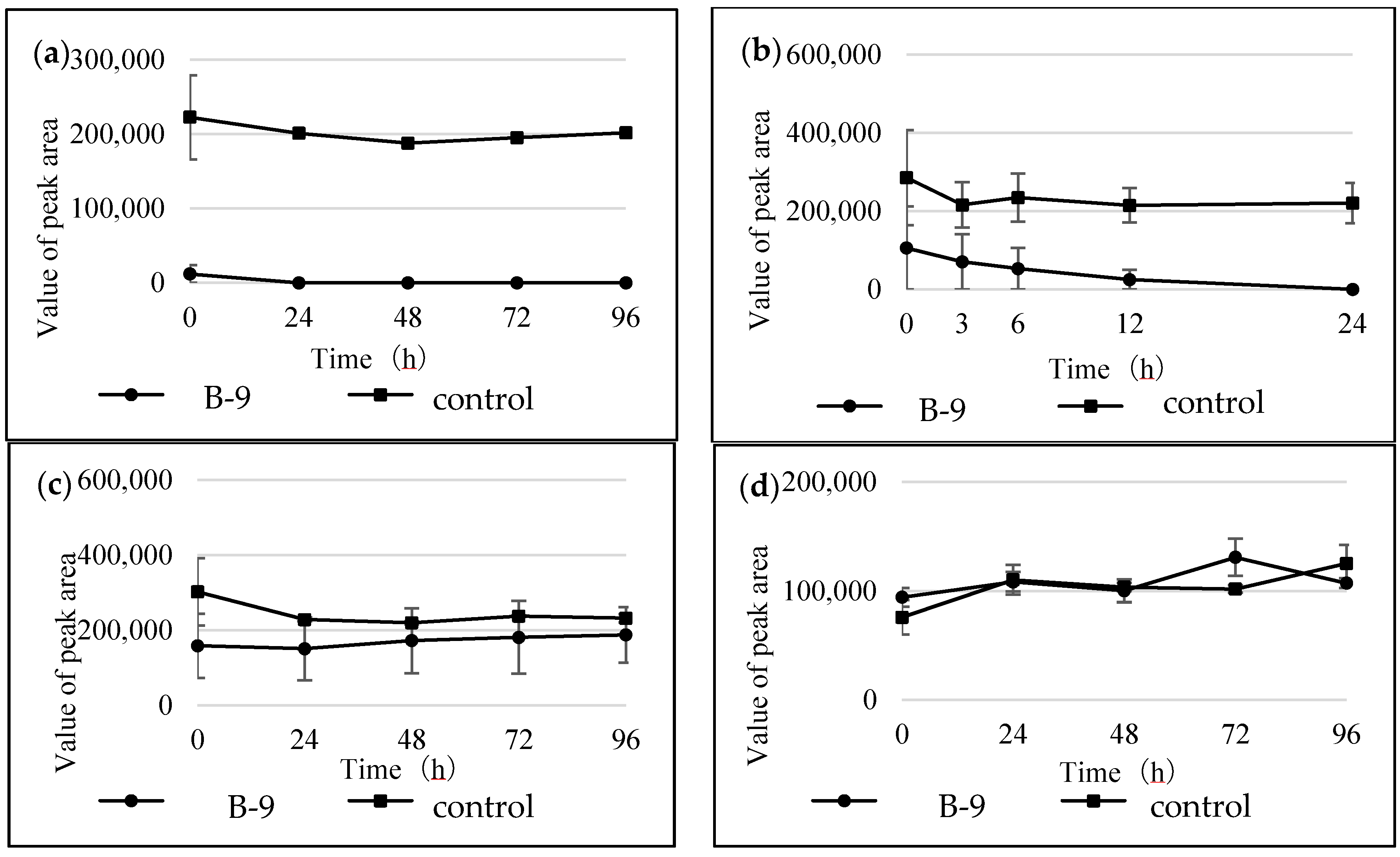
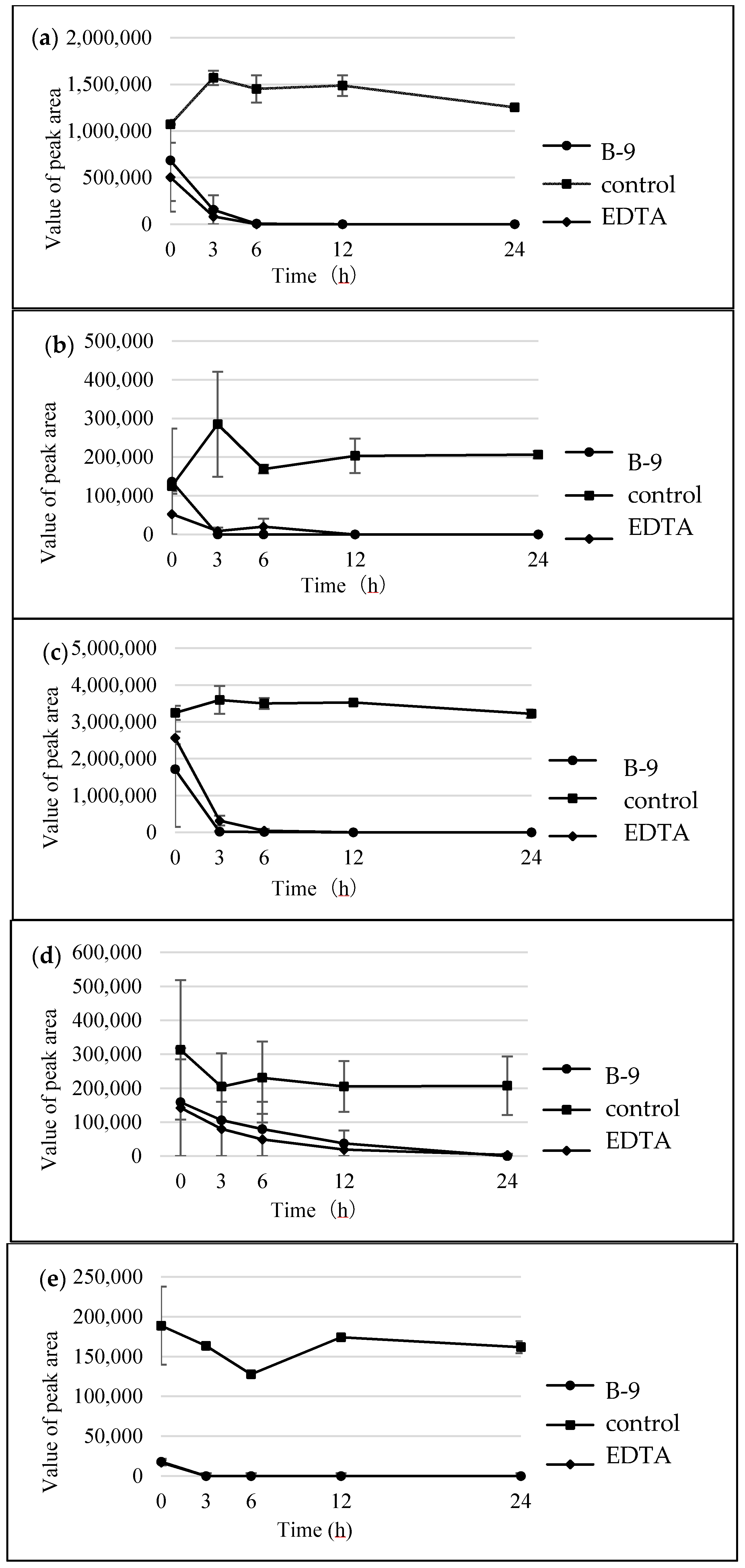
In the present study, we attempted to extend the applications of strain B-9, applying it to mycotoxins produced by fungi. Among the tested mycotoxins, only ochratoxin A was completely hydrolyzed to provide the constituents ochratoxin α and
l-phenylalanine (
Figure 2), and fumonisin B1 levels gradually decreased to a certain extent after 96 h due to the formation of a new peak at 12.7 min (
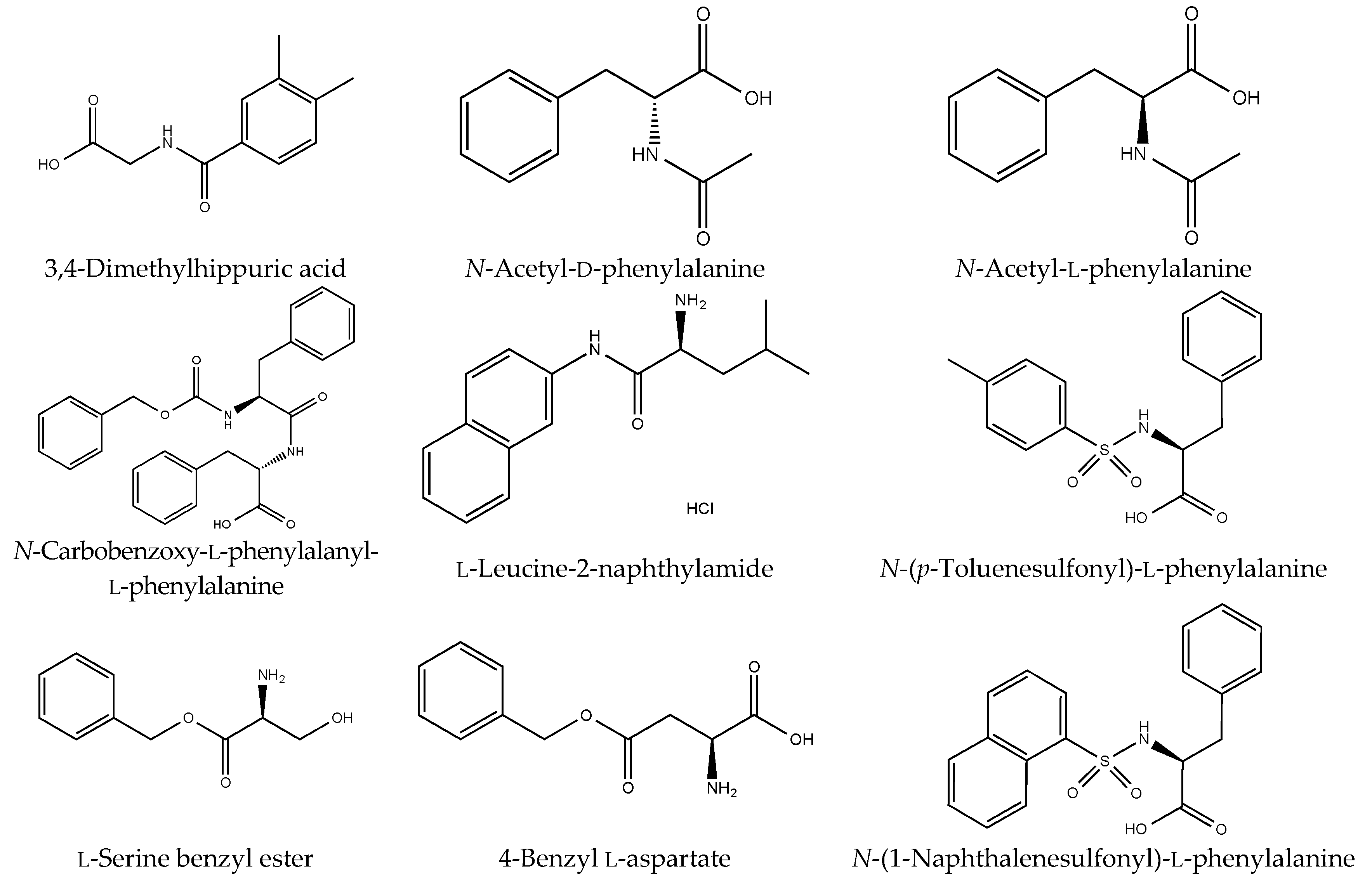
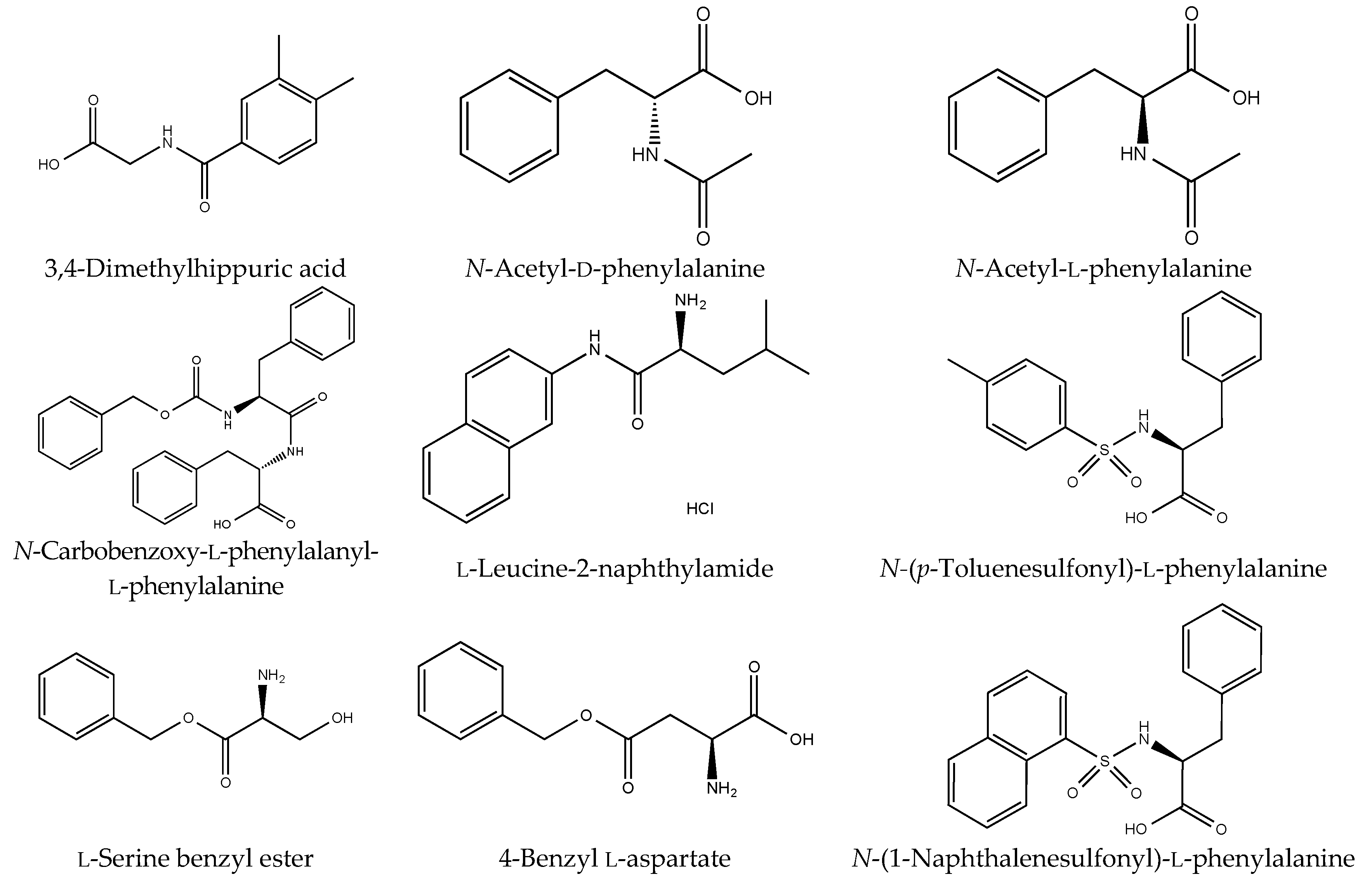
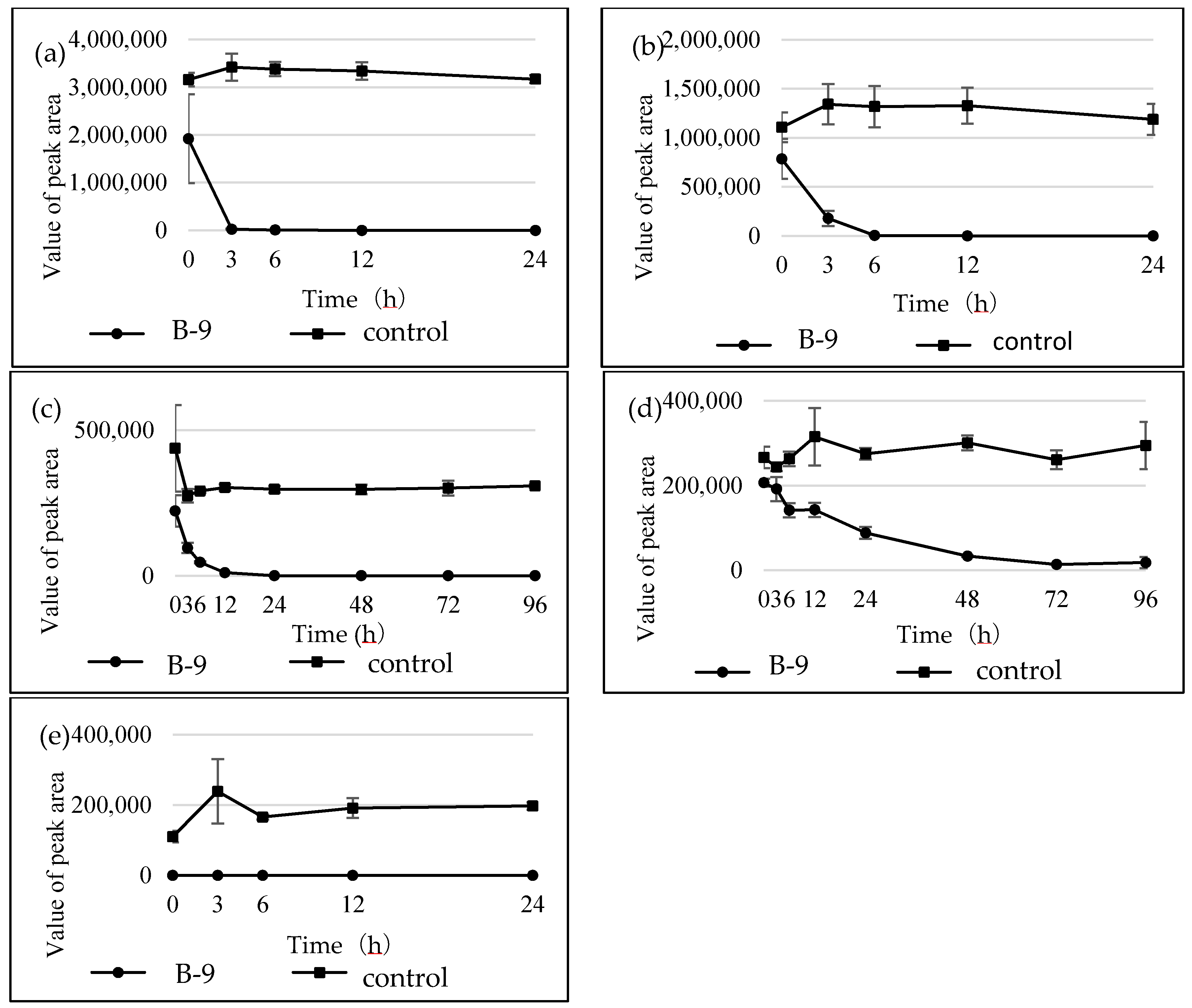
Figure S1). However, although drugs including antibiotics released into the aquatic environment were applied for microbial degradation using strain B-9, no degradation occurred. These results suggest that strain B-9 can only degrade amino acid-containing compounds. As expected, the tested compounds with amide and ester bonds were readily hydrolyzed by strain B-9, although the sulfonamides were not degraded (
Figure 4 and
Figure 6). In particular, the ester compounds were characteristically and rapidly hydrolyzed as soon as they came into contact with strain B-9 (
Figure 6). Furthermore, the degradations of the amide and ester compounds containing amino acids were not inhibited by the addition of EDTA, suggesting that the responsible enzyme is not MlrC.
It is understood that MlrC is found in the final stage in the microbial degradation of MC, catalyzing the degradation from linearized MC and tetrapeptide to smaller peptides and amino acids. Indeed, Dziga et al. reported the role of MlrC in MC degradation, in which linearized MC and tetrapeptides could be degraded to provide Adda by the cleavage of a peptide bond between Adda and Glu by a recombinant MlrC [
14]. These results suggest that strain B-9 possesses an additional hydrolytic enzyme that should be designated as MlrE. Furthermore, the results of the present study suggest that strain B-9 possesses an esterase. As mentioned above, strain B-9 degraded depsipeptides such as aeruginosins and mikamycin A, in which the cleavage at the ester bond was predominant over that of other peptide bonds [
16]. However, there may be a possibility that known proteases are responsible for the ester bond cleavage.
Since their discovery in 1994, it has been believed that MC-degrading microorganisms are only responsible for MC degradation [
2]. Although many reports on MC-degrading microorganisms have appeared since then [
3], few papers have described their substantial function and role in the aquatic environment. As reported by our group, one of the MC-degrading microorganisms, strain B-9, is applicable to structurally diverse peptide compounds, suggesting a different function. We should investigate the detailed function of each hydrolytic and transporter enzyme, as well as a system composed of these enzymes to understand their inherent roles under aquatic conditions.
4. Materials and Methods
4.1. Chemicals
As protease inhibitors, EDTA-2Na (purity: >99.5%) and PMSF (purity: >98.5%), were purchased from Dojindo Laboratories (Kumamoto, Japan) and Sigma-Aldrich Japan (Tokyo, Japan), respectively. Acetonitrile (ACN, LC/MS grade, purity: 99.8%), methanol (MeOH, LC/MS grade, purity: 99.7%), ethanol (EtOH, special grade, purity: 99.5%), formic acid (FA, LC/MS grade, purity: 99.5%), acetic acid (AcOH, LC/MS grade, purity: 99.5%), trifluoroacetic acid (TFA, special grade, purity: 98.0%), ammonium carbonate (special grade), and 28% ammonia solution (NH4OH, special grade) were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Water used for the preparation of all the solutions was purified using a Milli-Q apparatus (Millipore, Billerica, MA, USA); LC/MS analysis used ultrapure water from Wako. The mycotoxins (ochratoxin A, fumonisin B1, zearalenone, deoxynivalenol, patulin) were purchased from Sigma (St. Louis, MO, USA). Drugs with amide, ester, and sulfonamide bonds and amino acid-containing compounds were obtained from the following companies: Aldrich and Sigma Japan (Tokyo, Japan), Nacalai Tesque (Kyoto, Japan), Tokyo Chemical Industry (Tokyo, Japan), and Wako Pure Chemical Industries, Ltd. (Osaka, Japan).
4.2. MC-Degrading Bacterium
Bacterial strain B-9, isolated from the surface water of Lake Tsukui, Kanagawa, Japan, was previously reported to degrade various MCs and nodularin [
15]. This bacterium was inoculated into a flask containing 100 mL of Sakurai medium composed of peptone, yeast extract, and glucose, and incubated at 27 °C at 200 revolutions per minute (rpm) for 3 days.
4.3. Degradation of Tested Compounds
Two milligrams of the tested compounds was dissolved in 1 mL of EtOH and 50 μL of the solution was evaporated to dryness. One milliliter of the preincubated cell broth of strain B-9 (containing approximately 3 × 106 colony forming units (CFU) mL−1) was added to the residue. The resulting solution was mixed, and then incubated at 27 °C for 5, 15, 30, 60, and 120 min. After incubation, 50 μL of each of these mixtures was added to 50 μL of MeOH containing 0.2% FA and filtered using an Ultrafree-MC membrane centrifuge-filtration unit (hydrophilic polytetrafluoroethylene (PTFA), 0.20 μm, Millipore, Bedford, MA, USA) to stop the degradation and to eliminate proteins. Each supernatant was then analyzed by HPLC and LC/MS.
4.4. Enzyme Inhibition
Enzyme inhibitors were prepared as follows: EDTA was prepared as a 200-mM stock solution in water and was used at an assay concentration of 10 mM. PMSF was prepared as a 200-mM stock solution in EtOH and was used at an assay concentration of 10 mM. The cell broth and required inhibitor were preincubated at 27 °C for 30 min.
4.5. High-Performance Liquid Chromatography
The degradation process was monitored by HPLC-photo diode array (PDA) at 220 or 254 nm. The system consisted of a pair of LC 10AD VP pumps, a DGU 12A degasser, a CTO 6A column oven, an SPD 10A VP photodiode array detector, and an SCL 10A VP system controller (Shimadzu, Kyoto, Japan). Five microliters of the filtered sample were loaded onto a TSK-gel Super ODS column (2.0 μm, 2.0 × 100 mm, TOSOH, Tokyo, Japan) at 40 °C. The mobile phase was 0.1% formic acid in water (A) and 0.1% formic acid in methanol (B). The gradient conditions were initially 40–90% B for 20 min, and the flow rate was 200 μL/min.
4.6. Liquid Chromatography/Ion Trap Mass Spectrometry
The sample, column, mobile phase, and gradient conditions were the same as those used for the HPLC analysis (12). The LC separation was performed using the Agilent 1100 HPLC system (Agilent Technologies, Palo Alto, CA, USA). Five microliters of the sample was filtrated using an Ultrafree-MC membrane centrifuge filtration unit (hydrophilic PTFE, 0.20 μm, Millipore, Bedford, MA, USA) and loaded onto a TSK-gel Super ODS column (2.0 μm, 2.0 × 100 mm, TOSOH, Tokyo, Japan) at 40 °C. The mobile phase was 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B). The flow rate was 200 μL/min with UV detection at 254 nm. The gradient conditions were initially 10–90% B for 40 min. The entire eluate was directed into the mass spectrometer, where it was diverted to waste 2.5 min after injection to avoid any introduction of salts into the ion source. The MS analysis was accomplished using a Finnigan LCQ Deca XP plus ITMS (Thermo Fischer Scientific, San Jose, CA, USA) equipped with an electrospray ionization (ESI) interface. The ESI conditions in the positive ion mode were as follows: capillary temperature 300 °C, sheath gas flow rate 35 (arbitrary unit), ESI source voltage 5000 V, capillary voltage 43 V, and tube lens offset 15 V. Various scan ranges were used according to the molecular weights of the tested compounds.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}