Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Chemical Composition and Antioxidant Capacity of Padina pavonica Extract
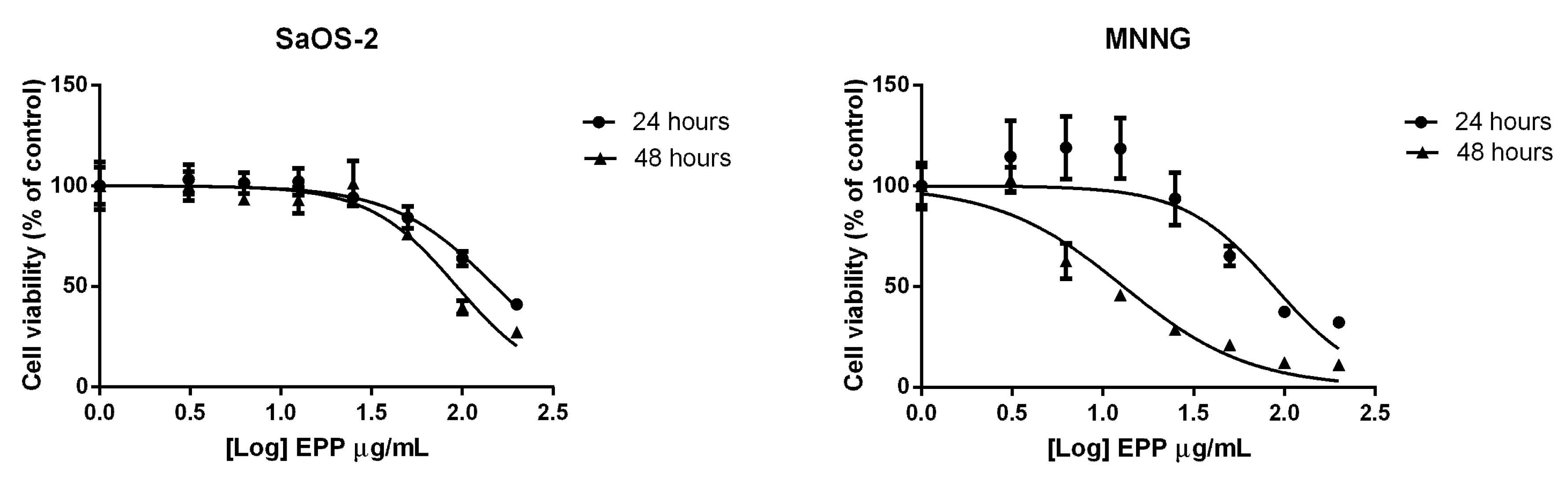
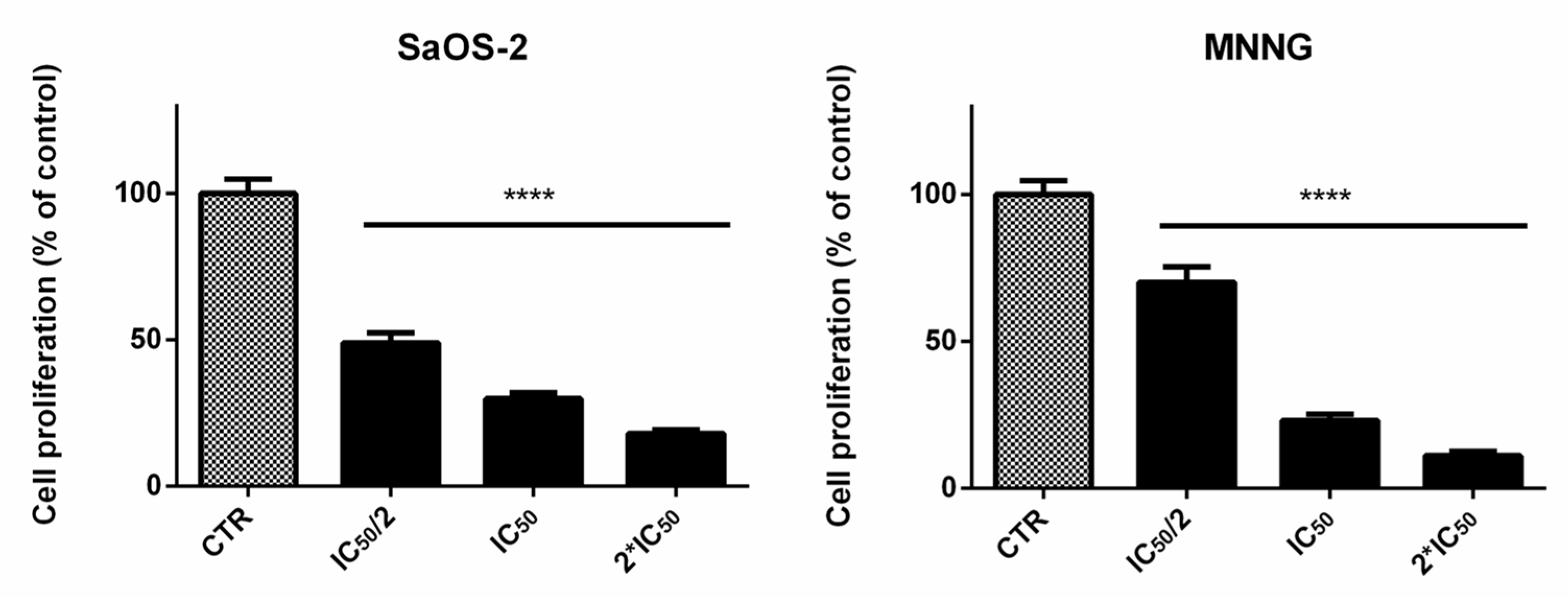
2.2. EPP Effects on OS Cell Viability and Proliferation
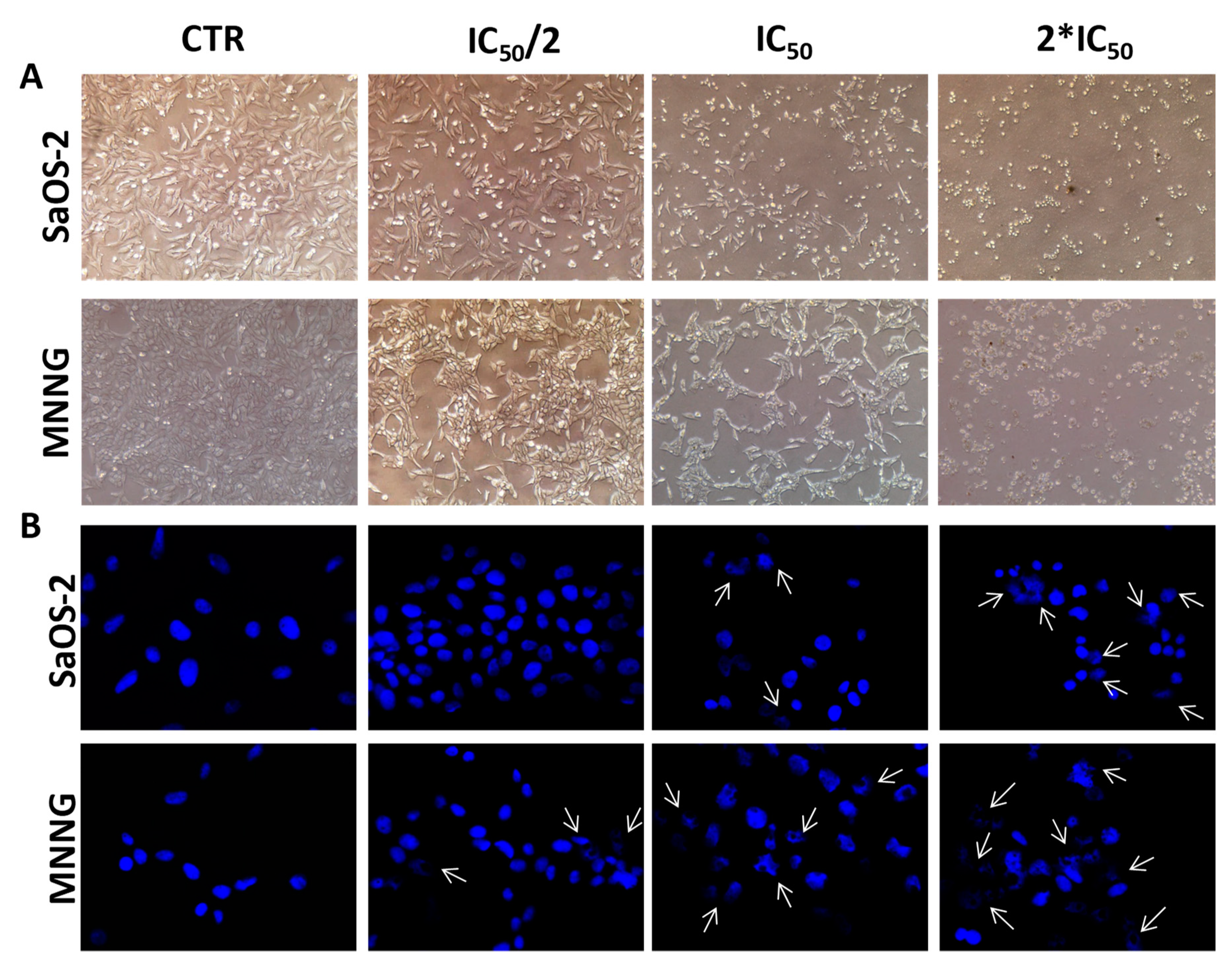
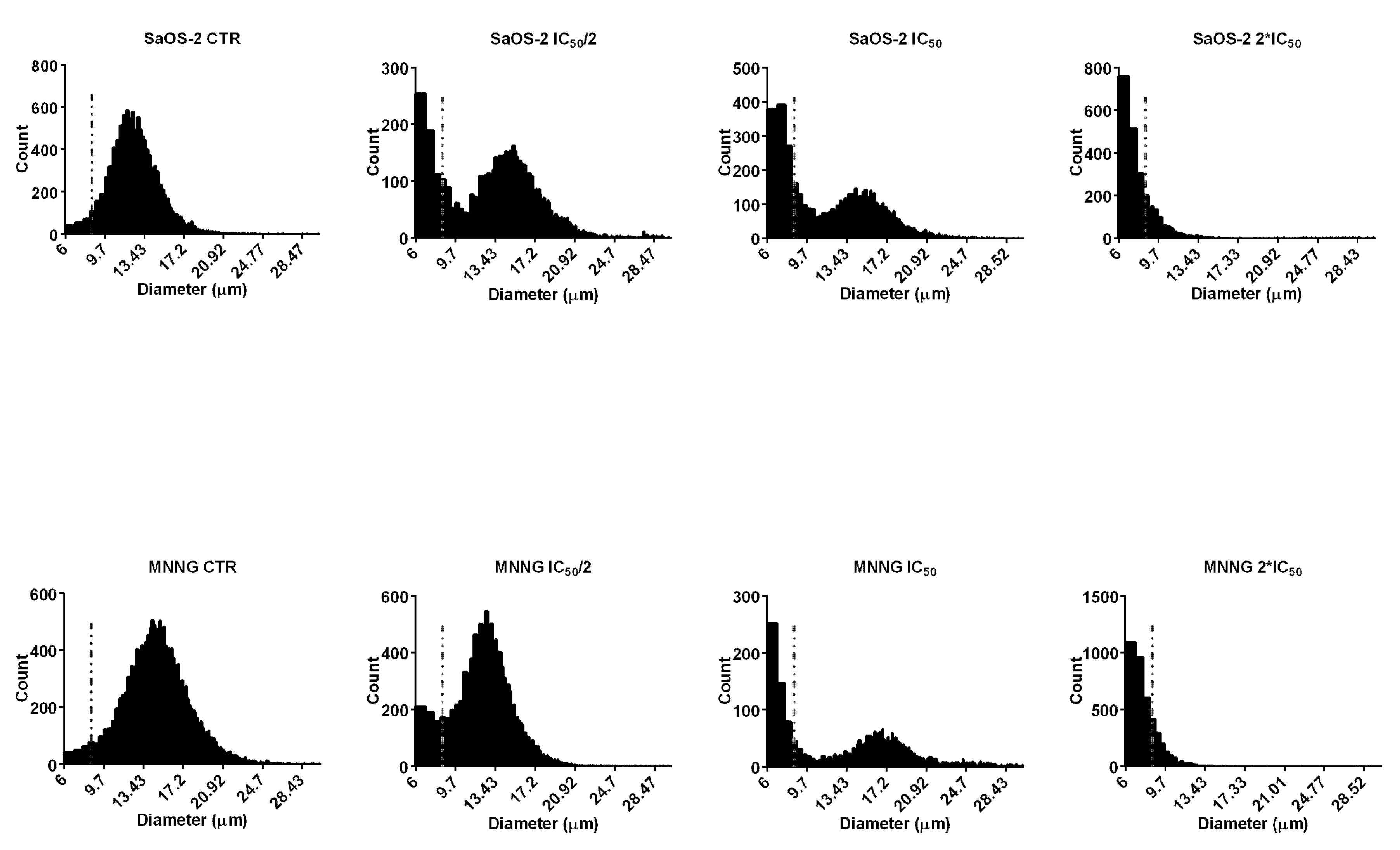
2.3. EPP Effects on OS Cell and Nuclear Morphology
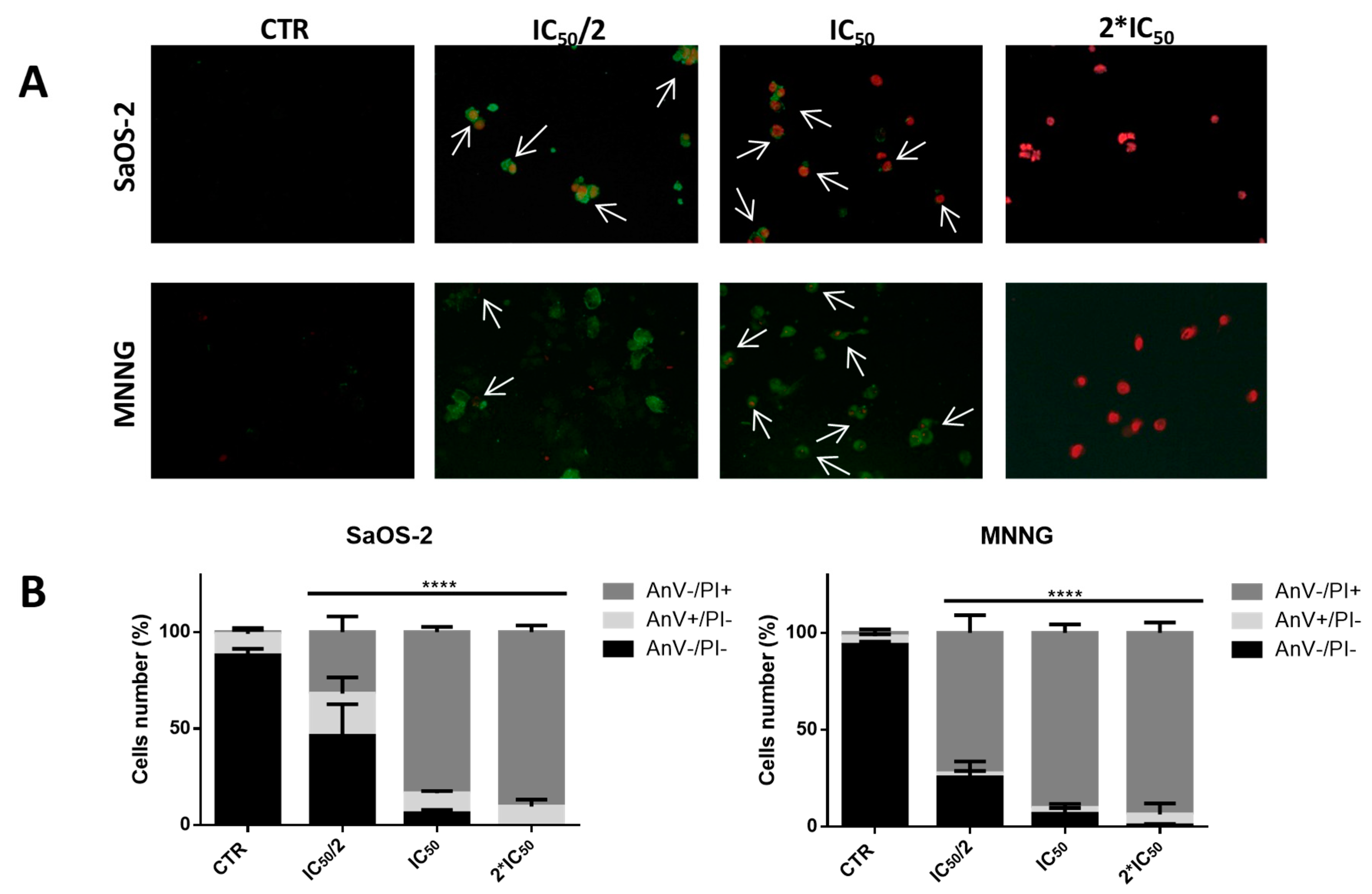
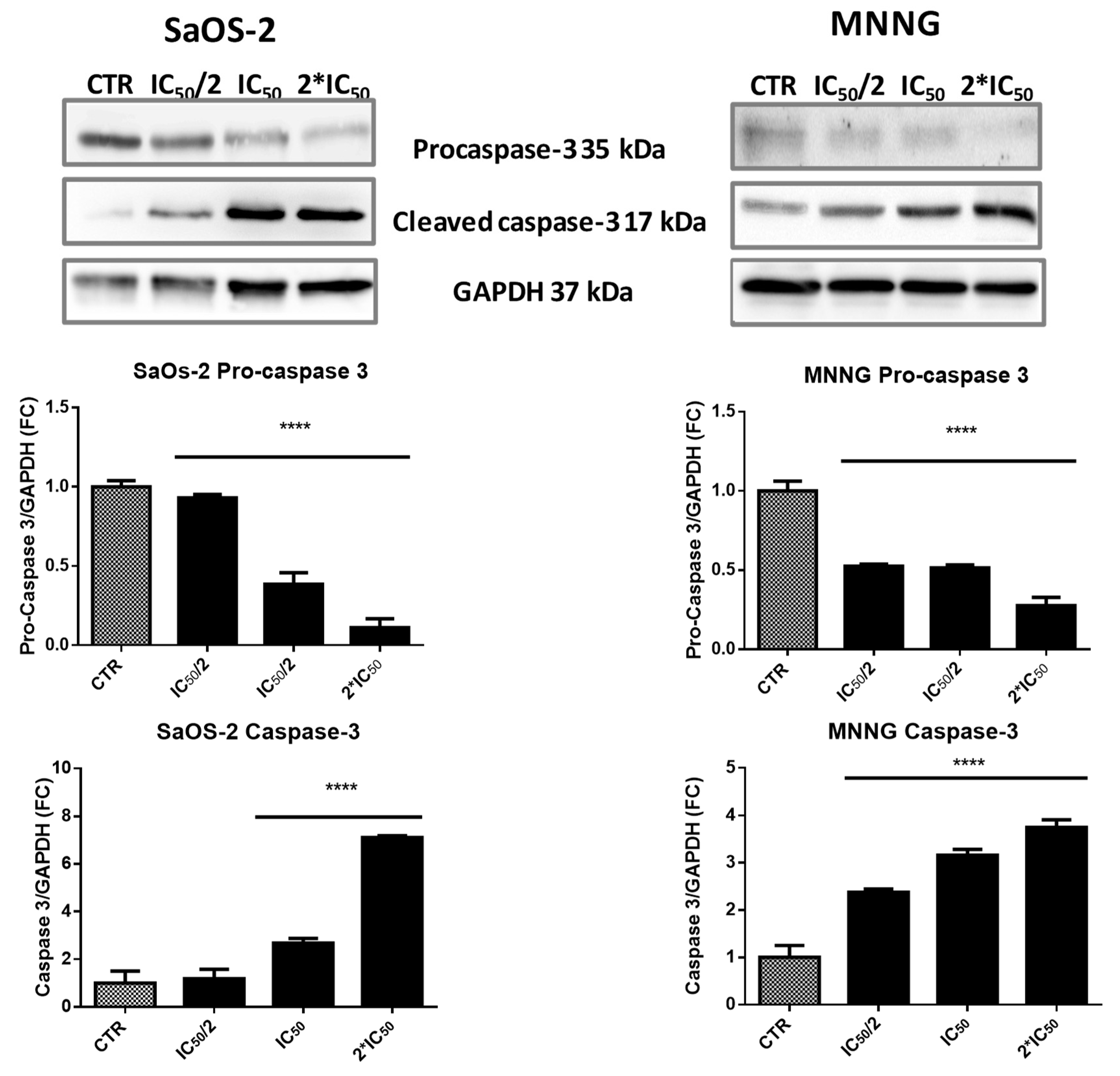
2.4. EPP Induces Apoptosis in OS Cells
3. Discussion
4. Materials and Methods
4.1. Collection of Padina pavonica and Preparation of the Algae Extract (EPP)
4.2. Total Phenolic Content
4.3. Total Flavonoid Content
4.4. Total Condensed Tannins Content
4.5. FRAP Scavenging Ability for the Antioxidant Activity
4.6. GC-MS Analysis
4.7. Cell Cultures
4.8. Cell Viability and Proliferation
4.9. Cell Morphology
4.10. Nuclear Staining with 4′, 6-Diamidine Phenylindole (DAPI)
4.11. Cell Diameter Analysis
4.12. Annexin V/Propidium Iodide Assay
4.13. Western Blot Analysis
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, R.C.; Costa-Lotufo, L.V. Bioprospecting of bioactives from seaweeds: Potential, obstacles and alternatives. Braz. J. Pharmacog. 2012, 22, 894–905. [Google Scholar] [CrossRef]
- Kamenarska, Z.; Gasic, M.J.; Zlatovic, M.; Rasovic, A.; Sladic, D.; Kljajic, Z.; Stefanov, K.; Seizova, K.; Najdenski, H.; Kujumgiev, A.; et al. Chemical composition of the brown alga Padina pavonia (L.) Gaill. from the Adriatic Sea. Bot. Mar. 2002, 45, 339–345. [Google Scholar] [CrossRef]
- Taskin, E.; Caki, Z.; Ozturk, M. Assessment of in vitro antitumoral and antimicrobial activities of marine algae harvested from the eastern Mediterranean sea. Afr. J. Biotechnol. 2010, 9, 4272–4277. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Lipej, L.; Orfanidis, S. Benthic macrophytes as a tool for delineating, monitoring and assessing ecological status: The case of Slovenian coastal waters. Mar. Pollut. Bull. 2008, 56, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T.; Olszewski, N.; Sebastiani, J.; Palka, S.; Sparacino, G.; Sciarrno, E.; Grebenok, R.J. Plant phloem sterol content: Forms, putative functions, and implications for phloem-feeding insects. Front. Plant Sci. 2013, 4, 370. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, G.; Laschi, M.; Geminiani, M.; Santucci, A. Proteomics of osteosarcoma. Expert Rev. Proteom. 2014, 11, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, G.; Braconi, D.; Spreafico, A.; Santucci, A. Post-genomics of bone metabolic dysfunctions and neoplasias. Proteomics 2012, 12, 708–721. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine Bioactive Compounds and Their Health Benefits: A Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Zubia, M.; Fabre, M.S.; Kerjean, V.; Le Lann, K.; Stiger-Pouvreau, V.; Fauchon, M.; Deslandes, E. Antioxidant and antitumor activities of some Phaeophyta from Brittany Coasts. Food Chem. 2009, 116, 693–701. [Google Scholar] [CrossRef]
- Khaled, N.; Hiba, M.; Asma, C. Antioxidant and Antifungal activities of Padina pavonica and Sargassum vulgare from the Lebanese Mediterranean Coast. Adv. Environ. Biol. 2012, 6, 42–48. [Google Scholar]
- Caf, F.; Yilmaz, Ö.; Durucan, F.; Özdemir, N.S. Biochemical components of three marine macroalgae (Padina pavonica, Ulva lactuca and Taonia atomaria) from the levantine sea coast of antalya, Turkey. J. Biodivers. Environ. Sci. 2015, 6, 401–411. [Google Scholar]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Rodrigues, A.I.; Mendes, S.; Pedrosa, R. Cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Hlila, M.B.; Hichri, A.O.; Mahjoub, M.A.; Mighri, Z.; Mastouri, M. Antioxidant and antimicrobial activities of Padina pavonica and Enteromorpha sp. from the Tunisian Mediterranean coast. J. Coast. Life Med. 2017, 5, 336–342. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.; Domínguez, R.; Carballo, J.; Franco, D.; Lorenzo, J.M. Proximate composition, phenolic content and in vitro antioxidant activity of aqueous extracts of the seaweeds Ascophyllum nodosum, Bifurcaria bifurcata and Fucus vesiculosus. Effect of addition of the extracts on the oxidative stability of canola oil under accelerated storage conditions. Food Res. Int. 2017, 99, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Kelman, D.; Kromkowski Posner, E.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant Activity of Hawaiian Marine Algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Kanias, G.D.; Skaltsa, H.; Tsitsa, E.; Loukis, A.; Bitis, J. Study of the correlation between trace elements, sterols and fatty acids in brown algae from the Saronikos Gulf of Greece. Fresenius’ J. Anal. Chem. 1992, 344, 334–339. [Google Scholar] [CrossRef]
- Petkov, G.; Furnadzieva, S.; Popov, S. Petrol-induced changes in the lipid and sterol composition of three microalgae. Phytochemistry 1992, 31, 1165–1166. [Google Scholar] [CrossRef]
- Combaut, G.; Yacoubou, A.; Piovetti, L.; Kornprobst, J.M. Sterols of the Senegalese brown alga Padina vickersiae. Phytochemistry 1985, 24, 618–619. [Google Scholar] [CrossRef]
- Abdul, Q.A.; Choi, R.J.; Jung, H.A.; Choi, J.S. Health benefit of fucosterol from marine algae: A review. J. Sci. Food Agric. 2016, 96, 1856–1866. [Google Scholar] [CrossRef]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Chiu, Y.H.; Duh, C.Y. Cytotoxic sterols from the formosan brown alga Turbinaria ornata. Planta Med. 1997, 63, 571–572. [Google Scholar] [CrossRef] [PubMed]
- Khanavi, M.; Gheidarloo, R.; Sadati, N.; Ardekani, M.R.S.; Nabavi, S.M.B.; Tavajohi, S.; Ostad, S.N. Cytotoxicity of fucosterol containing fraction of marine algae against breast and colon carcinoma cell line. Pharmacogn. Mag. 2012, 8, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Duh, C.Y. New cytotoxic oxygenated fucosterol from the brown alga Turbinaria conoides. J. Nat. Prod. 1999, 62, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Park, S.Y.; Chung, W.S.; Park, J.H.; Shin, H.S.; Hwang, E.; Kim, I.H.; Yi, T.H. The bone regenerative effects of fucosterol in in vitro and in vivo models of postmenopausal osteoporosis. Mol. Nutr. Food Res. 2014, 58, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Huh, G.W.; Lee, D.Y.; In, S.J.; Lee, D.G.; Park, S.Y.; Yi, T.H.; Kang, H.C.; Seo, W.D.; Baek, N.I. Fucosterols from Hizikia fusiformis and their proliferation activities on osteosarcoma-derived cell MG63. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 551–555. [Google Scholar] [CrossRef]
- Bang, M.H.; Kim, H.H.; Lee, D.Y.; Han, M.W.; Baek, Y.S.; Chung, D.K.; Baek, N.I. Anti-osteoporotic activities of fucosterol from sea mustard (Undaria pinnatifida). Food Sci. Biotechnol. 2011, 20, 343–347. [Google Scholar] [CrossRef]
- Smit, A.J. Medicinal and Pharmaceutical Uses of Seaweed Natural Products: A Review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Kelly, G.S. Squalene and its potential clinical uses. Altern. Med. Rev. 1999, 4, 29–36. [Google Scholar] [PubMed]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid Composition, Fatty Acids and Sterols in the Seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An Analysis from Nutritional, Chemotaxonomic, and Antiproliferative Activity Perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.I.; Rhee, S.H.; Park, K.Y. Anticancer activity of phytol and eicosatrienoic acid identified from Perilla leaves. J. Korean Soc. Food Sci. Nutr. 1999, 28, 1107–1112. [Google Scholar]
- Jiang, R.; Sun, L.; Wang, Y.; Liu, J.; Liu, X.; Feng, H.; Zhao, D. Chemical composition, and cytotoxic, antioxidant and antibacterial activities of the essential oil from ginseng leaves. Nat. Prod. Commun. 2014, 9, 865–868. [Google Scholar] [PubMed]
- Pejin, B.; Kojic, V.; Bogdanovic, G. An insight into the cytotoxic activity of phytol at in vitro conditions. Nat. Prod. Res. 2014, 28, 2053–2056. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Lee, H.J.; Jung, J.H.; Kim, Y.H.; Jung, D.B.; Sohn, E.J.; Lee, J.H.; Woo, H.J.; Baek, N.I.; Kim, Y.C.; et al. Activation of Caspase-9/3 and Inhibition of Epithelial Mesenchymal Transition are Critically Involved in Antitumor Effect of Phytol in Hepatocellular Carcinoma Cells. Phytother. Res. 2015, 29, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.W.; Cho, S.K. Phytol induces apoptosis and ROS-mediated protective autophagy in human gastric adenocarcinima AGS cells. Biochem. Anal. Biochem. 2015, 4, 211. [Google Scholar] [CrossRef]
- Venkataraman, B.; Samuel, L.A.; Pardha Saradhi, M.; Narashimharao, B.; Naga Vamsi Krishna, A.; Sudhakar, M.; Radhakrishnan, T.M. Antibacterial, antioxidant activity and GC-MS analysis of Eupatorium odoratum. Asian J. Pharm. Clin. Res. 2012, 5, 99–106. [Google Scholar]
- Sri Nurestri, A.M.; Sim, K.S.; Norhanom, A.W. Phytochemical and Cytotoxic Investigations of Pereskia grandifolia Haw. (Cactaceae) Leaves. J. Biol. Sci. 2009, 9, 488–493. [Google Scholar] [CrossRef]
- Rigano, D.; Russo, A.; Formisano, C.; Cardile, V.; Senatore, F. Antiproliferative and cytotoxic effects on malignant melanoma cells of essential oils from the aerial parts of Genista sessilifolia and G. tinctoria. Nat. Prod. Commun. 2010, 5, 1127–1132. [Google Scholar] [PubMed]
- Kusch, P.; Deininger, S.; Specht, S.; Maniako, R.; Haubrich, S.; Pommerening, T.; Kong Thoo Lin, P.; Hoerauf, A.; Kaiser, A. 2,5 In vitro and in vivo antimalarial activity assays of seeds from Balanites aegyptiaca: Compounds of the extract show growth inhibition and activity against plasmodial aminopeptidase. J. Parasitol. Res. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.A.; Jeong, T.S.; Park, D.S.; Xu, M.Z.; Oh, H.W.; Song, K.B.; Lee, W.S.; Park, H.Y. Antioxidant effects of quinoline alkaloids and 2,4-di-tert-butylphenol isolated from Scolopendra subspinipes. Biol. Pharm. Bull. 2006, 29, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Varsha, K.K.; Devendra, L.; Shilpa, G.; Priya, S.; Pandey, A.; Nampoothiri, K.M. 2,4-Di-tert-butyl phenol as the antifungal, antioxidant bioactive purified from a newly isolated Lactococcus sp. Int. J. Food Microbiol. 2015, 211, 44–50. [Google Scholar] [CrossRef]
- Hsouna, A.B.; Trigui, M.; Mansour, R.B.; Jarraya, R.M.; Damak, M.; Jaoua, S. Chemical composition, cytotoxicity effect and antimicrobial activity of Ceratonia siliqua essential oil with preservative effects against Listeria inoculated in minced beef meat. Int. J. Food Microbiol. 2011, 148, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Aknin, M.; Dogbevi, K.; Samb, A.; Kornprobst, J.M.; Gaydou, E.M.; Miralles, J. Fatty acid and sterol composition of eight algae from the Senegalese coast. Comp. Biochem. Physiol. 1992, 102, 841–843. [Google Scholar] [CrossRef]
- Tabarsa, M.; Rezaei, M.; Ramezanpour, Z.; Robert Waaland, J.; Rabiei, R. Fatty acids, amino acids, mineral contents, and proximate composition of some brown seaweeds. J. Phycol. 2012, 48, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Cavia Mdel, M.; Alonso-Torre, S.R. Antitumor effect of oleic acid; mechanisms of action: A review. Nutr. Hosp. 2012, 27, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Chajes, V.; Thiebaut, A.C.; Rotival, M.; Gauthier, E.; Maillard, V.; Boutron-Ruault, M.C.; Joulin, V.; Lenoir, G.M.; Clavel-Chapelon, F. Association between serum trans-monounsaturated fatty acids and breast cancer risk in the E3N-EPIC Study. Am. J. Epidemiol. 2008, 167, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Sianipar, N.F.; Purnamaningsih, R.; Rosaria. Bioactive compounds of fourth generation gamma-irradiated Typhoniumflagelliforme Lodd. mutants based on gas chromatography-mass spectrometry. In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2016; Volume 41, p. 012025. [Google Scholar] [CrossRef]
- Stoneham, M.; Goldacre, M.; Seagroatt, V.; Gill, L. Olive oil, diet and colorectal cancer: An ecological study and a hypothesis. J. Epidemiol. Community Health 2000, 54, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, R.; Fernández-Banares, F.; Navarro, E.; Castella, E.; Mane, J.; Alvarez, M.; Pastor, C.; Cabre, E.; Gassull, M. Effect of olive oil on early and late events of colon carcinogenesis in rats: Modulation of arachidonic acid metabolism and local prostaglandin E2 synthesis. Gut 2000, 46, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, B.; Birk, Y.; Raz, A.; Madar, Z. Nutritional-pharmacological combinations—A novel approach to reducing colon cancer incidence. Eur. J. Nutr. 2004, 43, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, I.A.; Brown, I.; Wahle, K.W.; Heys, S.D. Enhancing cytotoxic therapies for breast and prostate cancers with polyunsaturated fatty acids. Nutr. Cancer 2010, 62, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Menéndez, J.A.; del Mar Barbacid, M.; Montero, S.; Sevilla, E.; Escrich, E.; Solanas, M.; Cortés-Funes, H.; Colomer, R. Effects of gamma-linolenic acid and oleic acid on paclitaxel cytotoxicity in human breast cancer cells. Eur. J. Cancer 2001, 37, 402–413. [Google Scholar] [CrossRef]
- Ito, H.; Kasama, K.; Naruse, S.; Shimura, K. Antitumor effect of palmitoleic acid on Ehrlich ascites tumor. Cancer Lett. 1982, 17, 197–203. [Google Scholar] [CrossRef]
- Wahbeh, M.I. Amino acid and fatty acid profiles of four species of macroalgae from Aqaba and their suitability for use in fish diets. Aquaculture 1997, 159, 101–109. [Google Scholar] [CrossRef]
- Li, X.; Fan, X.; Han, L.; Lou, Q. Fatty acids of some algae from the Bohai Sea. Phytochemistry 2002, 59, 157–161. [Google Scholar] [CrossRef]
- Amaro, H.M.; Fernandes, F.; Valentão, P.; Andrade, P.B.; Sousa-Pinto, I.; Malcata, F.X.; Guedes, A.C. Effect of solvent system on extractability of lipidic components of Scenedesmus obliquus (M2-1) and Gloeothece sp. on antioxidant scavenging capacity thereof. Mar. Drugs 2015, 13, 6453–6471. [Google Scholar] [CrossRef]
- Lauvrak, S.U.; Munthe, E.; Kresse, S.H.; Stratford, E.W.; Namløs, H.M.; Meza-Zepeda, L.A.; Myklebost, O. Functional characterisation of osteosarcoma cell lines and identification of mRNAs and miRNAs associated with aggressive cancer phenotypes. Br. J. Cancer 2013, 109, 2228–2236. [Google Scholar] [CrossRef] [PubMed]
- Roepke, M.; Diestel, A.; Bajbouj, K.; Walluscheck, D.; Schonfeld, P.; Roessner, A.; Schneider-Stock, R.; Gali-Muhtasib, H. Lack of p53 augments thymoquinone-induced apoptosis and caspase activation in human osteosarcoma cells. Cancer Biol. Ther. 2007, 6, 160–169. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Abdella, E.M.; El-Derby, A.M.; Abdella, E.M. Protective Effects of Turbinaria ornata and Padina pavonia against Azoxymethane-Induced Colon Carcinogenesis through Modulation of PPAR Gamma, NF-κB and Oxidative Stress. Phytother. Res. 2015, 29, 737–748. [Google Scholar] [CrossRef]
- Ktari, L.; Guyot, M. A cytotoxic oxysterol from the marine alga Padina pavonica (L.) Thivy. J. Appl. Phycol. 1999, 11, 511–513. [Google Scholar] [CrossRef]
- Stanojković, T.P.; Šavikin, K.; Zdunić, G.; Kljajić, Z.; Grozdanić, N.; Antić, J. In vitro antitumoral Activities of Padina pavonia on human cervix and breast cancer cell Lines. J. Med. Plant Res. 2013, 7, 419–424. [Google Scholar] [CrossRef]
- Jaganathan, S.K.; Mandal, M. Antiproliferative effects of honey and of its polyphenols: A review. J. Biomed. Biotechnol. 2009, 2009, 1–13. [Google Scholar] [CrossRef]
- Zhang, Z.; Teruya, K.; Yoshida, T.; Eto, H.; Shirahata, S. Fucoidan extract enhances the anti-cancer activity of chemotherapeutic agents in MDA-MB-231 and MCF-7 breast cancer cells. Mar. Drugs 2013, 11, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Park, S.Y.; Lee, J.Y.; Park, J.H. Fucoidan present in brown algae induces apoptosis of human colon cancer cells. BMC Gastroenterol. 2010, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Konishi, I.; Hosokawa, M.; Sashima, T.; Kobayashi, H.; Miyashita, K. Halocynthiaxanthin and fucoxanthinol isolated from Halocynthia roretzi induce apoptosis in human leukemia, breast and colon cancer cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.X.; Hu, X.M.; Xu, S.Q.; Jiang, Z.J.; Yang, W. Effects of fucoxanthin on proliferation and apoptosis in human gastric adenocarcinoma MGC-803 cells via JAK/STATsignal pathway. Eur. J. Pharmacol. 2011, 657, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Song, F.L.; Gan, R.Y.; Zhang, Y.; Xiao, Q.; Kuang, L.; Li, H.B. Total Phenolic Contents and Antioxidant Capacities of Selected Chinese Medicinal Plants. Int. J. Mol. Sci. 2010, 11, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Yang, M.H.; Wen, H.-M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Broadhurst, R.B.; Jones, W.T. Analysis of Condensed Tannins Using Acidified Vanillin. J. Sci. Food Agric. 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Rezaee, M.; Assadia, Y.; Milani Hosseini, M.R.; Aghaee, E.; Ahmadi, F.; Berijani, S. Determination of organic compounds in water using dispersive liquid–liquid microextraction. J. Chromatogr. A 2006, 1116, 1–9. [Google Scholar] [CrossRef]
- Bernardini, G.; Geminiani, M.; Gambassi, S.; Orlandini, M.; Petricci, E.; Marzocchi, B.; Laschi, M.; Taddei, M.; Manetti, F.; Santucci, A. Novel smoothened antagonists as anti-neoplastic agents for the treatment of osteosarcoma. J. Cell. Physiol. 2018, 233, 4961–4971. [Google Scholar] [CrossRef]
- Laschi, M.; Bernardini, G.; Geminiani, M.; Ghezzi, L.; Amato, L.; Braconi, D.; Millucci, L.; Frediani, B.; Spreafico, A.; Franchi, A.; et al. Establishment of Four New Human Primary Cell Cultures from Chemo-Naïve Italian Osteosarcoma Patients. J. Cell. Physiol. 2015, 230, 2718–2727. [Google Scholar] [CrossRef] [PubMed]
- Laschi, M.; Bernardini, G.; Geminiani, M.; Manetti, F.; Mori, M.; Spreafico, A.; Campanacci, D.; Capanna, R.; Schenone, S.; Botta, M.; et al. Differentially activated Src kinase in chemo-naïve human primary osteosarcoma cells and effects of a Src kinase inhibitor. Biofactors 2017, 43, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Tahara, M.; Inoue, T.; Miyakura, Y.; Horie, H.; Yasuda, Y.; Fujii, H.; Kotake, K.; Sugano, K. Cell diameter measurements obtained with a handheld cell counter could be used as a surrogate marker of G2/M arrest and apoptosis in colon cancer cell lines exposed to SN-38. Biochem. Biophys. Res. Commun. 2013, 434, 753–759. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPP (%) | |
|---|---|
| Hydrocarbons | |
| Fatty acids | 68.83 |
| Squalene | 0.19 |
| Other hydrocarbons | 10.86 |
| Total hydrocarbons | 79.88 |
| Sterols | |
| Cholesterol | 0.97 |
| Fucosterol | 7.40 |
| Total sterols | 8.37 |
| Other compounds | |
| α-Tocopherol | 0.17 |
| δ-Tocopherol | 0.19 |
| Phytol | 5.27 |
| Neophytadiene | 5.56 |
| 2,4-di-tert-butylphenol (DTBP) | 0.18 |
| 2(4H)-Benzofuranone, 5,6,7,7a-tetrahydro-4,4,7a-trimethyl-, (R)-(Dihydroactinidiolide) | 0.37 |
| EPP (%) | |
|---|---|
| Hydrocarbons | |
| Fatty acids | 17.11 |
| Squalene | 0.19 |
| Other hydrocarbons | 1.65 |
| Total hydrocarbons | 18.95 |
| Sterols | |
| Cholesterol | 4.57 |
| Fucosterol | 17.57 |
| Total sterols | 22.14 |
| Other compounds | |
| δ-Tocopherol | 0.27 |
| Phytol | 53.85 |
| 2,4-di-tert-butylphenol (DTBP) | 2.99 |
| 6-Hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)-one | 0.99 |
| 2(4H)-Benzofuranone, 5,6,7,7a-tetrahydro-4,4,7a-trimethyl-, (R)-(Dihydroactinidiolide) | 0.62 |
| n-Nonadecanol-1 | 0.20 |
| Fatty Acids (FAs) | EPP (%) |
|---|---|
| Saturated fatty acids (SFAs) | |
| Lauric acid 12:0 | 0.47 |
| Myristic acid 14:0 | 0.43 |
| Pentadecanoic acid 15:0 | 1.95 |
| Palmitic acid 16:0 | 34.15 |
| Stearic acid 18:0 | 3.25 |
| Arachidic acid 20:0 | 0.74 |
| Behenic acid 22:0 | 0.04 |
| Other SFAs | 2.42 |
| Total SFAs | 43.45 |
| Monounsaturated fatty acids (MUFAs) | |
| Oleic acid (18:1 n-9) | 7.79 |
| Palmitelaidic acid (16:1 n-7 (E)) | 7.82 |
| Palmitoleic acid (16:1 n-7 (Z)) | 6.29 |
| Other MUFAs | 1.76 |
| Total MUFAs | 23.67 |
| Polyunsaturated fatty acids (PUFAs) | |
| Linoleic acid (18:2 n-6) | 0.53 |
| Eicosapentanoic acid (20:5 n-3) | 0.24 |
| Arachidonic acid (20:4 n-6) | 0.64 |
| Other PUFAs | 0.29 |
| Total PUFAs | 1.70 |
| Total n-6 PUFAs | 1.36 |
| Total n-3 PUFAs | 0.24 |
| Ratio n-6/n-3 | 5.61 |
| SaOS-2 OS Cells | Non-Apoptotic AnV−/PI− (%) ± SD | Early Apoptotis AnV+/PI− (%) ± SD | Late Apoptotis AnV+/PI+ (%) ± SD | p-Value |
| CTR | 88.04 ± 3.33 | 11.17 ± 2.94 | 0.79 ± 0.47 | |
| IC50/2 | 46.36 ± 16.31 | 21.80 ± 8.31 | 31.83 ± 8.20 | **** |
| IC50 | 6.11 ± 1.72 | 10.46 ± 1.07 | 83.43 ± 2.76 | **** |
| 2*IC50 | 0.00 | 9.60 ± 3.53 | 90.40 ± 3.53 | **** |
| MMNG OS Cells | Non-apoptotic AnV−/PI− (%) ± SD | Early Apoptotis AnV+/PI− (%) ± SD | Late Apoptotis AnV+/PI+ (%) ± SD | p-Value |
| CTR | 93.90 ± 1.54 | 4.30 ± 1.06 | 1.78 ± 1.78 | |
| IC50/2 | 25.48 ± 8.11 | 2.05 ± 1.26 | 72.46 ± 9.12 | **** |
| IC50 | 6.67 ± 2.89 | 2.99 ± 2.12 | 90.34 ± 4.27 | **** |
| 2*IC50 | 0.69 ± 0.69 | 5.63 ± 5.63 | 93.67 ± 5.32 | **** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardini, G.; Minetti, M.; Polizzotto, G.; Biazzo, M.; Santucci, A. Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells. Mar. Drugs 2018, 16, 504. https://doi.org/10.3390/md16120504
Bernardini G, Minetti M, Polizzotto G, Biazzo M, Santucci A. Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells. Marine Drugs. 2018; 16(12):504. https://doi.org/10.3390/md16120504
Chicago/Turabian StyleBernardini, Giulia, Mariagiulia Minetti, Giuseppe Polizzotto, Manuele Biazzo, and Annalisa Santucci. 2018. "Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells" Marine Drugs 16, no. 12: 504. https://doi.org/10.3390/md16120504
APA StyleBernardini, G., Minetti, M., Polizzotto, G., Biazzo, M., & Santucci, A. (2018). Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells. Marine Drugs, 16(12), 504. https://doi.org/10.3390/md16120504