Discovery Methodology of Novel Conotoxins from Conus Species


 ,
, 
Abstract
1. Introduction
2. Diversity of Conotoxins
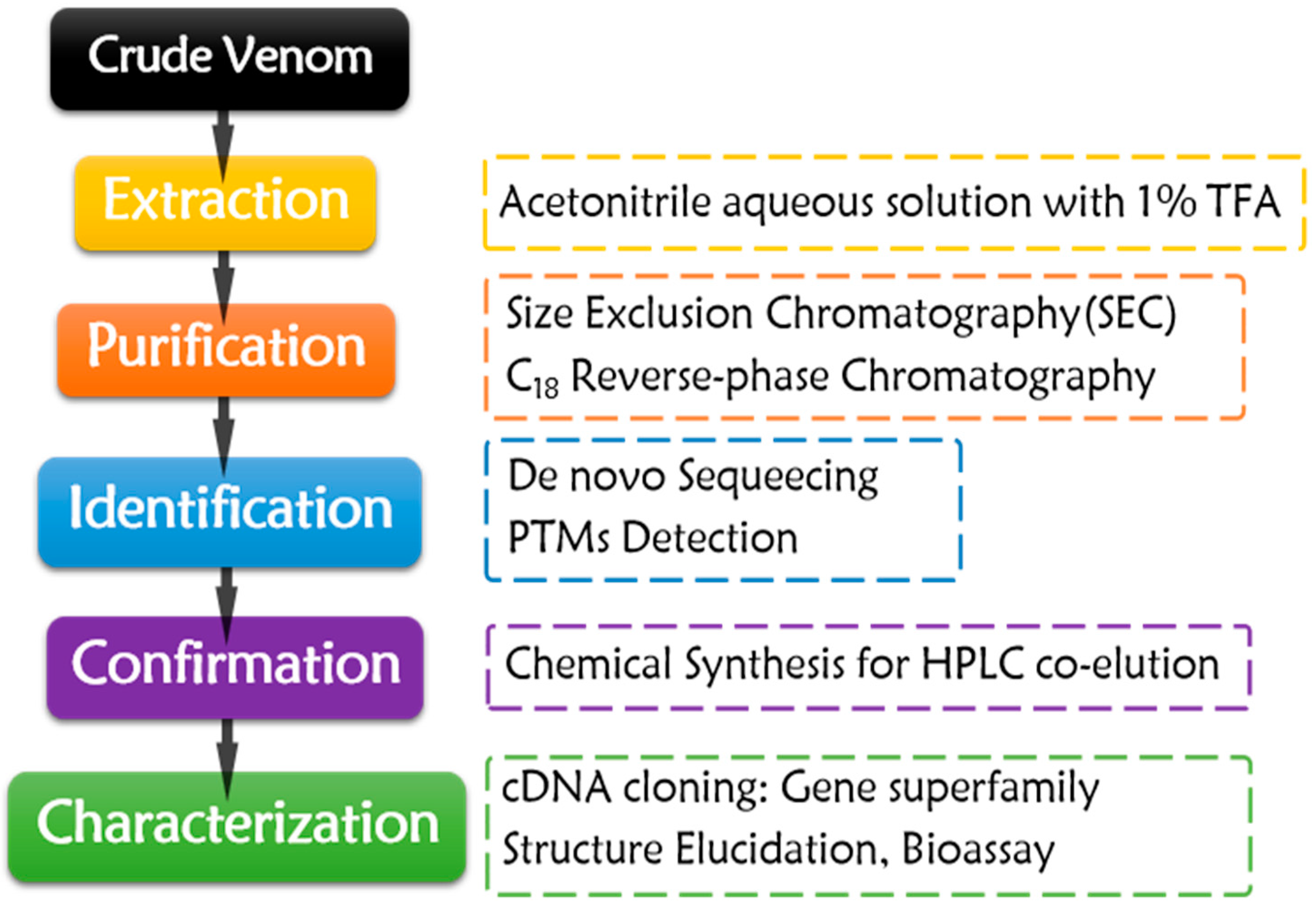
3. Conotoxins Purified from Crude Venom
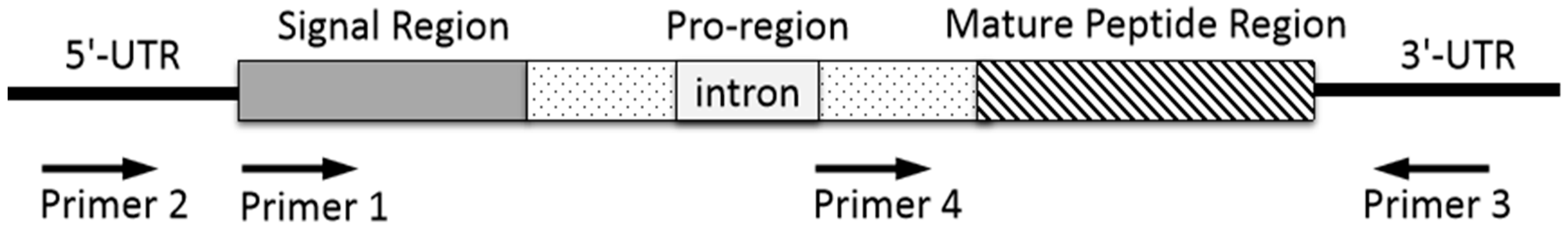
4. Gene Cloning to Discover New Conotoxins
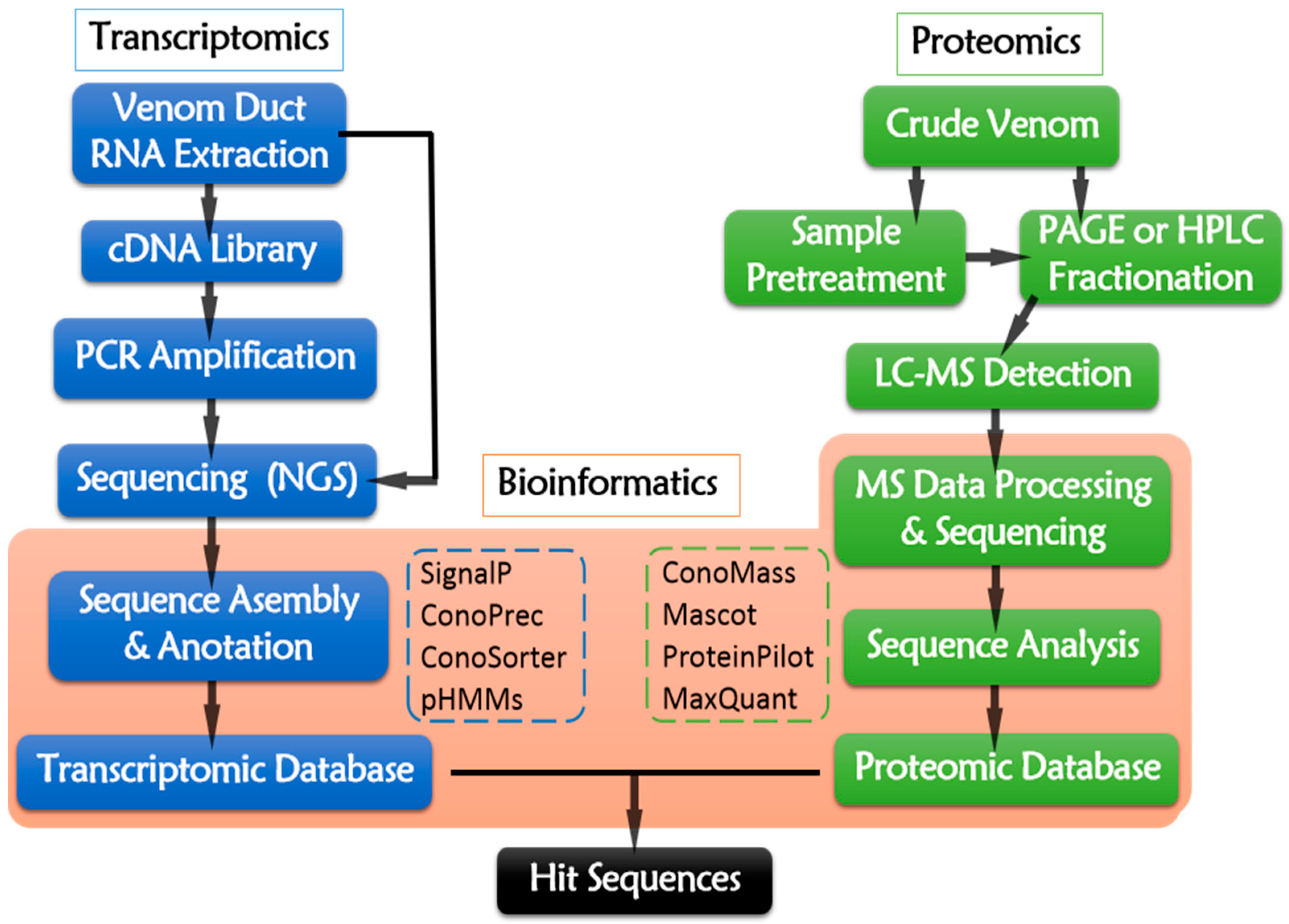
5. Cone Snail Multi-Omics
5.1. Transcriptomics—A Useful Pathway to Identify Putative Conotoxins
5.2. Proteomics—An Effective Approach to Discovery Natural Conotoxins
5.3. Bioinformaics—An Efficient Tool for Massive Data Processing and Integrating
5.4. Multi-Omics Integration
6. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BLAST | Basic local alignment search tool |
| CID | Collision induced dissociation |
| ESI | Electrospray ionization |
| ETD | Electron transfer dissociation |
| EThcD | Electron transfer higher energy collision dissociation |
| GPCRs | G protein-coupled receptors |
| HPLC | High performance liquid chromatography |
| LTQ-Orbitrap | Linear Trap Quatropole-Orbitrap |
| MS | Mass spectrometry |
| MALDI | Matrix-assisted laser desorption ionization |
| nAChRs | Nicotinic acetylcholine receptors |
| NCBI | National center of biotechnology information |
| NMR | Nuclear magnetic resonance |
| NMDA | N-methyl-d-aspartic acid receptor |
| PAGE | Poly acrylamide gel electrophoresis |
| PCR | Polymerase chain reaction |
| RACE | Rapid amplification of cDNA ends |
| SILAC | Stable isotope labeling by amino acids in cell culture |
| SPPS | Solid phase peptide synthesis |
| TOF | Time of flight |
References
- Prashanth, J.R.; Dutertre, S.; Jin, A.H.; Lavergne, V.; Hamilton, B.; Cardoso, F.C.; Griffin, J.; Venter, D.J.; Alewood, P.F.; Lewis, R.J. The role of defensive ecological interactions in the evolution of conotoxins. Mol. Ecol. 2016, 25, 598–615. [Google Scholar] [CrossRef] [PubMed]
- Endean, R.; Duchemin, C. The venom apparatus of Conus magus. Toxicon 1967, 4, 275–284. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.; Kelley, W.P.; Rubakhin, S.S.; Bingham, J.P.; Sweedler, J.V.; Gilly, W.F. Anatomical correlates of venom production in Conus californicus. Biol. Bull. 2002, 203, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, S.M.; Martin, G.G.; Kier, W.M.; Schulz, J.R. Venom kinematics during prey capture in Conus: The biomechanics of a rapid injection system. J. Exp. Biol. 2010, 213, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.J. Cone Shell Stings. Recent Cases of Human Injury due to Venomous Marine Snails of the Genus Conus. Hawaii Med. J. 1958, 17, 528. [Google Scholar] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Tosti, E.; Boni, R.; Gallo, A. µ-Conotoxins Modulating Sodium Currents in Pain Perception and Transmission: A Therapeutic Potential. Mar. Drugs 2017, 15, 295. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.; Mcarthur, J.R.; Adams, D.J. Conotoxins Targeting Neuronal Voltage-Gated Sodium Channel Subtypes: Potential Analgesics? Toxins 2012, 4, 1236–1260. [Google Scholar] [CrossRef]
- Leipold, E.; Ullrich, F.; Thiele, M.; Tietze, A.A.; Terlau, H.; Imhof, D.; Heinemann, S.H. Subtype-specific block of voltage-gated K+ channels by μ-conopeptides. Biochem. Biophys. Res. Commun. 2017, 482, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, D.; Gonzalez, W.; Fissore, R.A.; Carvacho, I. Conotoxins as Tools to Understand the Physiological Function of Voltage-Gated Calcium (CaV) Channels. Mar. Drugs 2017, 15, 313. [Google Scholar] [CrossRef] [PubMed]
- Bourinet, E.; Zamponi, G.W. Block of voltage-gated calcium channels by peptide toxins. Neuropharmacology 2017, 127, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; Mcintosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 771. [Google Scholar] [CrossRef] [PubMed]
- England, L.J.; Imperial, J.; Jacobsen, R.; Craig, A.G.; Gulyas, J.; Akhtar, M.; Rivier, J.; Julius, D.; Olivera, B.M. Inactivation of a serotonin-gated ion channel by a polypeptide toxin from marine snails. Science 1998, 281, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.E.; White, H.S.; Wilcox, K.S. The effect of CGX-1007 and CI-1041, novel NMDA receptor antagonists, on NMDA receptor-mediated EPSCs. Epilepsy Res. 2004, 59, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Harrington, A.M.; Garciacaraballo, S.; Maddern, J.; Grundy, L.; Zhang, J.; Page, G.; Miller, P.E.; Craik, D.J.; Adams, D.J. α-Conotoxin Vc1.1 inhibits human dorsal root ganglion neuroexcitability and mouse colonic nociception via GABAB receptors. Gut 2017, 66, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.T.; Clark, R.J. G-Protein Coupled Receptors Targeted by Analgesic Venom Peptides. Toxins 2017, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Rogge, G.; Hague, C.; Alewood, D.; Colless, B.; Lewis, R.J.; Minneman, K.P. Subtype-selective noncompetitive or competitive inhibition of human alpha1-adrenergic receptors by rho-TIA. J. Biol. Chem. 2004, 279, 35326–35333. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, I.A.; Gehrmann, J.; Loughnan, M.L.; Thomas, L.; Adams, D.A.; Atkins, A.; Palant, E.; Craik, D.J.; Adams, D.J.; Alewood, P.F. Two new classes of conopeptides inhibit the alpha1-adrenoceptor and noradrenaline transporter. Nat. Neurosci. 2001, 4, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Möller, C.; Marí, F. A vasopressin/oxytocin-related conopeptide with gamma-carboxyglutamate at position 8. Biochem. J. 2007, 404, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Zhang, L.; Smith, M.D.; Walewska, A.; Vellore, N.A.; Baron, R.; Mcintosh, J.M.; White, H.S.; Olivera, B.M.; Bulaj, G. A marine analgesic peptide, Contulakin-G, and neurotensin are distinct agonists for neurotensin receptors: Uncovering structural determinants of desensitization properties. Front. Pharmacol. 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Paczkowski, F.A.; Sharpe, I.A.; Dutertre, S.; Lewis, R.J. chi-Conotoxin and tricyclic antidepressant interactions at the norepinephrine transporter define a new transporter model. J. Biol. Chem. 2007, 282, 17837. [Google Scholar] [CrossRef] [PubMed]
- Romero, H.K.; Christensen, S.B.; Di Cesare Mannelli, L.; Gajewiak, J.; Ramachandra, R.; Elmslie, K.S.; Vetter, D.E.; Ghelardini, C.; Iadonato, S.P.; Mercado, J.L.; et al. Inhibition of alpha9alpha10 nicotinic acetylcholine receptors prevents chemotherapy-induced neuropathic pain. Proc. Natl. Acad. Sci. USA 2017, 114, E1825–E1832. [Google Scholar] [CrossRef] [PubMed]
- Hannon, H.E.; Atchison, W.D. Omega-Conotoxins as Experimental Tools and Therapeutics in Pain Management. Mar. Drugs 2013, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Crooks, P.A.; Bardo, M.T.; Dwoskin, L.P. Nicotinic receptor antagonists as treatments for nicotine abuse. Adv. Pharmacol. 2014, 69, 513–551. [Google Scholar] [CrossRef] [PubMed]
- Gandini, M.A.; Sandoval, A.; Felix, R. Toxins targeting voltage-activated Ca2+ channels and their potential biomedical applications. Curr. Top. Med. Chem. 2015, 15, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Irasema, O.P.; Mario, N.; Cervantes-Luevano, K.E.; Carolina, Á.-D.; Guy, S.; Sanchez-Campos, L.N.; Licea-Navarro, A.F. Apoptosis Activation in Human Lung Cancer Cell Lines by a Novel Synthetic Peptide Derived from Conus californicus Venom. Toxins 2016, 8, 38. [Google Scholar] [CrossRef]
- Yang, R.; Liu, Y.; Hou, X.; Fan, Y.; Li, J.; Chen, M.; Wang, Y.; Zhang, X.; Zhang, M. MAPKs-mediated modulation of the myocyte voltage-gated K+ channels is involved in ethanol-induced rat coronary arterial contraction. Eur. J. Pharmacol. 2018, 834, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Dendorfer, A.; Finolurdaneta, R.K.; Terlau, H.; Olivera, B.M. Biochemical Characterization of κM-RIIIJ, a Kv1.2 Channel Blocker. J. Biol. Chem. 2010, 285, 14882–14889. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Lewis, R.J. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top. Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259. [Google Scholar] [CrossRef] [PubMed]
- Layer, R.T.; Mcintosh, J.M. Conotoxins: Therapeutic Potential and Application. Mar. Drugs 2006, 4, 119–142. [Google Scholar] [CrossRef]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure: Activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Peng, C.; Yang, J.; Yi, Y.; Zhang, J.; Shi, Q. Cone Snails: A Big Store of Conotoxins for Novel Drug Discovery. Toxins 2017, 9, 397. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, J.R.; Brust, A.; Jin, A.H.; Alewood, P.F.; Dutertre, S.; Lewis, R.J. Cone snail venomics: From novel biology to novel therapeutics. Future Med. Chem. 2014, 6, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Halai, R.; Craik, D.J. Conotoxins: Natural product drug leads. Nat. Prod. Rep. 2009, 26, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef] [PubMed]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin. Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Obata, H.; Conklin, D.; Eisenach, J.C. Spinal noradrenaline transporter inhibition by reboxetine and Xen2174 reduces tactile hypersensitivity after surgery in rats. Pain 2005, 113, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, N.L.; Campbell, T.J.; Polakowski, J.S.; Bulaj, G.; Layer, R.T.; Moore, J.; Gross, G.J.; Cox, B.F. Postischemic administration of CGX-1051, a peptide from cone snail venom, reduces infarct size in both rat and dog models of myocardial ischemia and reperfusion. J. Cardiovasc. Pharm. 2005, 46, 141–146. [Google Scholar] [CrossRef]
- Clark, R.J.; Fischer, H.; Nevin, S.T.; Adams, D.J.; Craik, D.J. The synthesis, structural characterization, and receptor specificity of the alpha-conotoxin Vc1.1. J. Biol. Chem. 2006, 281, 23254–23263. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Allen, J.; Wagstaff, J.S.; Yaksh, T. The pharmacokinetics of the conopeptide contulakin-G (CGX-1160) after intrathecal administration: An analysis of data from studies in beagles. Anesth. Analg. 2007, 104, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.; Zhang, M.M.; Azam, L.; Olivera, B.M.; Bulaj, G.; Yoshikami, D. Navβ subunits modulate the inhibition of Nav1.8 by the analgesic gating modifier μO-conotoxin MrVIB. J. Pharmacol. Exp. Ther. 2011, 338, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Hieble, J.P.; Robert, R.R., Jr. The use of alpha-adrenoceptor antagonists in the pharmacological management of benign prostatic hypertrophy: An overview. Pharmacol. Res. 1996, 33, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Brust, A.; Palant, E.; Croker, D.E.; Colless, B.; Drinkwater, R.; Patterson, B.; Schroeder, C.I.; Wilson, D.; Nielsen, C.K.; Smith, M.T. chi-Conopeptide pharmacophore development: Toward a novel class of norepinephrine transporter inhibitor (Xen2174) for pain. J. Med. Chem. 2009, 52, 6991–7002. [Google Scholar] [CrossRef] [PubMed]
- Puillandre1, N.; Duda, T.F.; Meyer, C.; Olivera, B.M.; Bouchet, P. One, four or 100 genera? A new classification of the cone snails. J. Molluscan Stud. 2015, 81, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2015, 112, E3782. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Woodward, S.R.; Cruz, L.J.; Olivera, B.M.; Hillyard, D.R. Constant and hypervariable regions in conotoxin propeptides. EMBO J. 1990, 9, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Yang, L.; Xu, S.; Wang, C. Various Conotoxin Diversifications Revealed by a Venomic Study of Conus flavidus. Mol. Cell. Proteom. 2014, 13, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Dutertre, S.; Kaas, Q.; Lavergne, V.; Kubala, P.; Lewis, R.J.; Alewood, P.F. Transcriptomic messiness in the venom duct of Conus miles contributes to conotoxin diversity. Mol. Cell. Proteom. 2013, 12, 3824–3833. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, J.A.; Kelley, W.P.; Sweedler, J.V. Screening for post-translational modifications in conotoxins using liquid chromatography/mass spectrometry: An important component of conotoxin discovery. Toxicon 2006, 47, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Riveraortiz, J.A.; Cano, H.; Marí, F. Intraspecies variability and conopeptide profiling of the injected venom of Conus ermineus. Peptides 2011, 32, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Gray, W.R.; Olivera, B.M. Purification and properties of a myotoxin from Conus geographus venom. Arch. Biochem. Biophys. 1978, 190, 539–548. [Google Scholar] [CrossRef]
- Edman, P.; Begg, G. A Protein Sequenator. FEBS J. 1967, 1, 80–91. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Pi, C.; Zeng, X.; Zhou, M.; Jiang, X.; Chen, S.; Ren, Z.; Xu, A. Identification of a novel M-superfamily conotoxin with the ability to enhance tetrodotoxin sensitive sodium currents. Arch. Toxicol. 2009, 83, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Van, D.H.A.; Peigneur, S.; Dyubankova, N.; Möller, C.; Marí, F.; Diego-García, E.; Naudé, R.; Lescrinier, E.; Herdewijn, P.; Tytgat, J. Pc16a, the first characterized peptide from Conus pictus venom, shows a novel disulfide connectivity. Peptides 2012, 34, 106–113. [Google Scholar] [CrossRef]
- Bernáldez, J.; Romángonzález, S.A.; Martínez, O.; Jiménez, S.; Vivas, O.; Arenas, I.; Corzo, G.; Arreguín, R.; García, D.E.; Possani, L.D. A Conus regularis Conotoxin with a Novel Eight-Cysteine Framework Inhibits CaV2.2 Channels and Displays an Anti-Nociceptive Activity. Mar. Drugs 2013, 11, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.; Peigneur, S.; Maiti, M.; Mille, B.G.; Devi, P.; Ravichandran, S.; Lescrinier, E.; Waelkens, E.; D’Souza, L.; Herdewijn, P. Discovery of a new subclass of α-conotoxins in the venom of Conus australis. Toxicon 2014, 91, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.M.; Peigneur, S.; Maiti, M.; Devi, P.; Ravichandran, S.; Lescrinier, E.; Ulens, C.; Waelkens, E.; D’Souza, L.; Herdewijn, P. Structure-Function Elucidation of a New α-Conotoxin, Lo1a, from Conus longurionis. J. Biol. Chem. 2014, 91, 170–171. [Google Scholar] [CrossRef]
- Nguyen, B.; Le, C.J.; Aráoz, R.; Thai, R.; Lamthanh, H.; Benoit, E.; Molgó, J. Isolation, purification and functional characterization of alpha-BnIA from Conus bandanus venom. Toxicon 2014, 91, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, T.; Kompella, S.N.; Yan, M.; Lu, A.; Wang, Y.; Shao, X.; Chi, C.; Adams, D.J.; Ding, J.; et al. Conotoxin alphaD-GeXXA utilizes a novel strategy to antagonize nicotinic acetylcholine receptors. Sci. Rep. 2015, 5, 14261. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Liu, J.; Ren, Z.; Yu, C.; Xu, A. Discovery of two P-superfamily conotoxins, lt9a and lt9b, with different modifications on voltage-sensitive sodium channels. Toxicon 2017, 134, 6–13. [Google Scholar] [CrossRef]
- Jiang, S.; Tae, H.S.; Xu, S.; Shao, X.; Adams, D.J.; Wang, C. Identification of a Novel O-Conotoxin Reveals an Unusual and Potent Inhibitor of the Human α9α10 Nicotinic Acetylcholine Receptor. Mar. Drugs 2017, 15, 170. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.D.; Liu, L.; Shao, X.X.; Peng, C.; Chi, C.W.; Guo, Z.Y. New conotoxins define the novel I3-superfamily. Peptides 2009, 30, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Chen, F.; Cai, H.; Xiao, A.; You, Q.; Lu, Y. Isolation and Characterization of Conotoxin bt5a from Conus betulinus. Chin. J. Nat. Med. 2010, 8, 132–136. [Google Scholar] [CrossRef]
- Möller, C.; Marí, F. 9.3 KDa components of the injected venom of Conus purpurascens define a new five-disulfide conotoxin framework. Biopolymers 2011, 96, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, M.B.; Zugasti-Cruz, A.; Falcón, A.; Batista, C.V.F.; Olivera, B.M.; Cotera, E.P.H.D.L. A novel arrangement of Cys residues in a paralytic peptide of Conus cancellatus (jr. syn.: Conus austini), a worm-hunting snail from the Gulf of Mexico. Peptides 2013, 41, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Heghinian, M.D.; Mejia, M.; Adams, D.J.; Godenschwege, T.A.; Marí, F. Inhibition of cholinergic pathways in Drosophila melanogaster by α-conotoxins. FASEB J. 2015, 29, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ye, M.; Wang, Y.; Shao, X.; Yuan, D.; Liu, J.; Hawrot, E.; Wang, C.; Chi, C. A new subfamily of conotoxins belonging to the A-superfamily. Peptides 2010, 31, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.B.; Bandyopadhyay, P.K.; Olivera, B.M.; McIntosh, J.M. αS-conotoxin GVIIIB potently and selectively blocks α9α10 nicotinic acetylcholine receptors. Biochem. Pharmacol. 2015, 96, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Espino, S.S.; Dilanyan, T.; Imperial, J.S.; Aguilar, M.B.; Teichert, R.W.; Bandyopadhyay, P.; Olivera, B.M. Glycine-rich Conotoxins from the Virgiconus clade. Toxicon 2016, 113, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.; Ghequire, M.G.; Peigneur, S.; Mille, B.G.; Devi, P.; Ravichandran, S.; Waelkens, E.; D’Souza, L.; De, M.R.; Tytgat, J. Novel Conopeptides of Largely Unexplored Indo Pacific Conus sp. Mar. Drugs 2016, 14, 199. [Google Scholar] [CrossRef] [PubMed]
- Echterbille, J.; Gilles, N.; Araóz, R.; Mourier, G.; Amar, M.; Servent, D.; Pauw, E.D.; Quinton, L. Discovery and characterization of EII B, a new α-conotoxin from Conus ermineus venom by nAChRs affinity capture monitored by MALDI-TOF/TOF mass spectrometry. Toxicon 2017, 130, 1. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, M.F.; Rodriguez, A.M.; Cano, H.; Clark, E.; Tae, H.S.; Adams, D.J.; Godenschwege, T.A.; Marí, F. In vivo and in vitro testing of native α-conotoxins from the injected venom of Conus purpurascens. Neuropharmacology 2017, 127, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Kauferstein, S.; Kendel, Y.; Nicke, A.; Coronas, F.I.V.; Possani, L.D.; Favreau, P.; Krizaj, I.; Wunder, C.; Kauert, G.; Mebs, D. New conopeptides of the D-superfamily selectively inhibiting neuronal nicotinic acetylcholine receptors. Toxicon 2009, 54, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.C.; Olivera, B.M. Divergent M- and O-superfamily peptides from venom of fish-hunting Conus parius. Peptides 2010, 31, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Hong, J.; Zhou, M.; Huang, L.; Shao, X.; Yang, Y.; Sigworth, F.J.; Chi, C.; Lin, D.; Wang, C. A novel conotoxin, qc16a, with a unique cysteine framework and folding. Peptides 2011, 32, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Khoo, K.K.; Xu, S.; Zhou, M.; Boonyalai, N.; Perugini, M.A.; Shao, X.; Chi, C.; Galea, C.A.; Wang, C. A helical conotoxin from Conus imperialis has a novel cysteine framework and defines a new superfamily. J. Biol. Chem. 2012, 287, 14973–14983. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shao, X.; Yan, M.; Chi, C.; Lu, A.; Wang, C. Identification of Two Novel O2-Conotoxins from Conus generalis. Int. J. Pept. Res. Ther. 2015, 21, 81–89. [Google Scholar] [CrossRef]
- Vetter, I.; Dekan, Z.; Knapp, O.; Adams, D.J.; Alewood, P.F.; Lewis, R.J. Isolation, characterization and total regioselective synthesis of the novel μO-conotoxin MfVIA from Conus magnificus that targets voltage-gated sodium channels. Biochem. Pharmacol. 2012, 84, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Inserra, M.C.; Kompella, S.N.; Vetter, I.; Brust, A.; Daly, N.L.; Cuny, H.; Craik, D.J.; Alewood, P.F.; Adams, D.J.; Lewis, R.J. Isolation and characterization of α-conotoxin LsIA with potent activity at nicotinic acetylcholine receptors. Biochem. Pharmacol. 2013, 86, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Kompella, S.N.; Akondi, K.B.; Melaun, C.; Daly, N.L.; Luetje, C.W.; Alewood, P.F.; Craik, D.J.; Adams, D.J.; Mari, F. RegIIA: An alpha 4/7-conotoxin from the venom of Conus regius that potently blocks alpha 3 beta 4 nAChRs. Biochem. Pharmacol. 2012, 83, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.C.; Nery, A.A.; Ulrich, H.; Konno, K.; Sciani, J.M.; Pimenta, D.C. α-RgIB: A Novel Antagonist Peptide of Neuronal Acetylcholine Receptor Isolated from Conus regius Venom. Int. J. Pept. 2013, 2013, 543028. [Google Scholar] [CrossRef] [PubMed]
- Favreau, P.; Benoit, E.; Hocking, H.G.; Carlier, L.; Hoedt, D.D.; Leipold, E.; Markgraf, R.; Schlumberger, S.; Córdova, M.A.; Gaertner, H. A novel µ-conopeptide, CnIIIC, exerts potent and preferential inhibition of NaV1.2/1.4 channels and blocks neuronal nicotinic acetylcholine receptors. Br. J. Pharmacol. 2012, 166, 1654–1668. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.; Caer, J.P.; Mourier, G.; Thai, R.; Lamthanh, H.; Servent, D.; Benoit, E.; Molgó, J. Characterization of a Novel Conus bandanus Conopeptide Belonging to the M-Superfamily Containing Bromotryptophan. Mar. Drugs 2014, 12, 3449–3465. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, M.; Miyashita, M.; Kitanaka, A.; Juichi, H.; Sarhan, M.; Fouda, M.; Abdel-Rahman, M.; Saber, S.; Nakagawa, Y. Characterization of the venom of the vermivorous cone snail Conus fulgetrum. Biosci. Biotechnol. Biochem. 2016, 80, 1879–1882. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Du, T.; Liu, Z.; Wu, Q.; Feng, G.; Dong, M.; Zhou, X.; Jiang, L.; Dai, Q. Im10A, a short conopeptide isolated from Conus imperialis and possesses two highly concentrated disulfide bridges and analgesic activity. Peptides 2016, 81, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Johanna, B.; Samanta, J.; Javier, G.L.; Noda, F.J.; Enrique, S.; Emilio, S.; Daniela, C.; Aguilar, M.B.; Alexei, L.N. A New Member of Gamma-Conotoxin Family Isolated from Conus princeps Displays a Novel Molecular Target. Toxins 2016, 8, 39. [Google Scholar] [CrossRef]
- Han, T.S.; Teichert, R.W.; Olivera, B.M.; Bulaj, G. Conus venoms—A rich source of peptide-based therapeutics. Curr. Pharm. Des. 2008, 14, 2462–2479. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, characterization, synthesis, and cloning of the lockjaw peptide from Conus purpurascens venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.D.; Mcintosh, J.M.; Hillyard, D.R.; Cruz, L.J.; Olivera, B.M. The A-superfamily of conotoxins: Structural and functional divergence. J. Biol. Chem. 2004, 279, 17596–17606. [Google Scholar] [CrossRef] [PubMed]
- Mcintosh, J.M.; Plazas, P.V.; Watkins, M.; Gomezcasati, M.E.; Olivera, B.M.; Elgoyhen, A.B. A Novel α-Conotoxin, PeIA, Cloned from Conus pergrandis, Discriminates between Rat α9α10 and α7 Nicotinic Cholinergic Receptors. J. Biol. Chem. 2005, 280, 30107–30112. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.D.; Han, Y.H.; Wang, C.G.; Chi, C.W. From the identification of gene organization of conotoxins to the cloning of novel toxins. Toxicon 2007, 49, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jiang, H.; Han, Y.H.; Yuan, D.D.; Chi, C.W. Two different groups of signal sequence in M-superfamily conotoxins. Toxicon 2008, 51, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Liu, L.; Shao, X.; Chi, C.; Wang, C. Identification of a novel class of conotoxins defined as V-conotoxins with a unique cysteine pattern and signal peptide sequence. Peptides 2008, 29, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.D.; Liu, L.; Shao, X.X.; Peng, C.; Chi, C.W.; Guo, Z.Y. Isolation and cloning of a conotoxin with a novel cysteine pattern from Conus caracteristicus. Peptides 2008, 29, 1521–1525. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Nielsen, H.; Von, H.G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kass, Q.; Wu, Y.; Zhu, X.; Hu, Y.; Li, X.; Tsetlin, V.I.; Christensen, S. Cloning, synthesis, and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, E4026. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Wu, Y.; Zhu, X.; Hu, Y.; Mcintyre, M.; Christensen, S.; Akcan, M.; Craik, D.J.; Mcintosh, J.M. Characterization of a Novel α-Conotoxin from Conus textile that Selectively Targets α6/α3β2β3 Nicotinic Acetylcholine Receptors. J. Biol. Chem. 2013, 288, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; Mcintosh, J.M. Characterization of a Novel Alpha-Conotoxin TxID from Conus textile that Potently Blocks rat Alpha3beta4 Nicotinic Acetylcholine Receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Schroeder, C.I.; Zhu, X.; Hu, Y.; Wu, Y.; Weltzin, M.M.; Eberhard, S.; Kaas, Q.; Craik, D.J.; et al. A novel α4/7-conotoxin LvIA from Conus lividus that selectively blocks α3β2 vs. α6/α3β2β3 nicotinic acetylcholine receptors. FASEB J. 2014, 28, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, L.; Ning, H.; Cai, F.; Liu, Z.; Zhang, L.; Zhou, L.; Dai, Q. Cloning, Synthesis and Functional Characterization of a Novel α-Conotoxin Lt1.3. Mar. Drugs 2018, 16, 112. [Google Scholar] [CrossRef] [PubMed]
- Zhangsun, D.; Luo, S.; Wu, Y.; Zhu, X.; Hu, Y.; Xie, L. Novel O-superfamily conotoxins identified by cDNA cloning from three vermivorous Conus species. Chem. Biol. Drug Des. 2010, 68, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Swa, H.; Lewis, R.J. Venomics-Accelerated Cone Snail Venom Peptide Discovery. Int. J. Mol. Sci. 2018, 19, 788. [Google Scholar] [CrossRef]
- Utkin, Y.N. Modern trends in animal venom research—Omics and nanomaterials. World J. Biol. Chem. 2017, 8, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Animal venom studies: Current benefits and future developments. World J. Biol. Chem. 2015, 6, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Yao, G.; Gao, B.M.; Fan, C.X.; Bian, C.; Wang, J.; Cao, Y.; Wen, B.; Zhu, Y.; Ruan, Z. High-throughput identification of novel conotoxins from the Chinese tubular cone snail (Conus betulinus) by multi-transcriptome sequencing. Gigascience 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. Next-generation sequencing platforms. Annu. Rev. Anal. Chem. 2013, 6, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Quail, M.A.; Miriam, S.; Paul, C.; Otto, T.D.; Harris, S.R.; Connor, T.R.; Anna, B.; Swerdlow, H.P.; Yong, G. A tale of three next generation sequencing platforms: Comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers. BMC Genom. 2012, 13, 341. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Young, N.D.; Williamson, N.A.; Purcell, A.W. Proteomic interrogation of venom delivery in marine cone snails: Novel insights into the role of the venom bulb. J. Proteome Res. 2010, 9, 5610–5619. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.; Jin, A.H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. Comparison of the Venom Peptides and Their Expression in Closely Related Conus Species: Insights into Adaptive Post-speciation Evolution of Conus Exogenomes. Genome Biol. Evol. 2015, 7, 1797–1814. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Von, G.H.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Dutertre, S.; Jin, A.H.; Lewis, R.J.; Taft, R.J.; Alewood, P.F. Systematic interrogation of the Conus marmoreus venom duct transcriptome with ConoSorter reveals 158 novel conotoxins and 13 new gene superfamilies. BMC Genom. 2013, 14, 708. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavihemami, H.; Mcintosh, L.D.; Purcell, A.W.; Norton, R.S.; Papenfuss, A.T. Diversity of Conotoxin Gene Superfamilies in the Venomous Snail, Conus victoriae. PLoS ONE 2014, 9, e87648. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.C.; Halai, R.; Wang, C.K.L.; Craik, D.J. ConoServer, a database for conopeptide sequences and structures. Bioinformatics 2008, 24, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Yu, R.; Jin, A.H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef] [PubMed]
- Magrane, M.; Martin, M.J.; O’Donovan, C.; Apweiler, R. Protein Sequence Databases. Curr. Opin. Chem. Biol. 2004, 8, 76–80. [Google Scholar] [CrossRef]
- Consortium, U.P. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Holford, M.; Zhang, M.M.; Gowd, K.H.; Azam, L.; Green, B.R.; Watkins, M.; Ownby, J.P.; Yoshikami, D.; Bulaj, G.; Olivera, B.M. Pruning nature: Biodiversity-derived discovery of novel sodium channel blocking conotoxins from Conus bullatus. Toxicon 2009, 53, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Gilly, W.F.; Richmond, T.A.; Duda, T.F., Jr.; Elliger, C.; Lebaric, Z.; Schulz, J.; Bingham, J.P.; Sweedler, J.V. A diverse family of novel peptide toxins from an unusual cone snail, Conus californicus. J. Exp. Biol. 2011, 214, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Helena, S.H.; Lu, A.; Li, Q.; Fedosov, A.E.; Jason, B.; Patrice, S.C.; Jon, S.; Mark, Y.; Olivera, B.M. Venom Insulins of Cone Snails Diversify Rapidly and Track Prey Taxa. Mol. Biol. Evol. 2016, 33, 2924–2934. [Google Scholar] [CrossRef]
- Gao, B.; Peng, C.; Lin, B.; Chen, Q.; Zhang, J.; Shi, Q. Screening and Validation of Highly-Efficient Insecticidal Conotoxins from a Transcriptome-Based Dataset of Chinese Tubular Cone Snail. Toxins 2017, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Tayo, L.L.; Lu, B.; Cruz, L.J.; Rd, Y.J. Proteomic analysis provides insights on venom processing in Conus textile. J. Proteome Res. 2010, 9, 2292. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Characterization of the Conus bullatus genome and its venom-duct transcriptome. BMC Genom. 2011, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Terrat, Y.; Biass, D.; Dutertre, S.; Favreau, P.; Remm, M.; Stöcklin, R.; Piquemal, D.; Ducancel, F. High-resolution picture of a venom gland transcriptome: Case study with the marine snail Conus consors. Toxicon 2012, 59, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Lluisma, A.O.; Milash, B.A.; Moore, B.; Olivera, B.M.; Bandyopadhyay, P.K. Novel venom peptides from the cone snail Conus pulicarius discovered through next-generation sequencing of its venom duct transcriptome. Mar. Genom. 2012, 5, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. High Conopeptide Diversity in Conus tribblei Revealed Through Analysis of Venom Duct Transcriptome Using Two High-Throughput Sequencing Platforms. Mar. Biotechnol. 2015, 17, 81. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Vetter, I.; Himaya, S.W.A.; Alewood, P.F.; Lewis, R.J.; Dutertre, S. Transcriptome and proteome of Conus planorbis identify the nicotinic receptors as primary target for the defensive venom. Proteomics 2015, 15, 4030–4040. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Li, Q.; Lu, A.; Bandyopadhyay, P.K.; Yandell, M.; Olivera, B.M.; Safavihemami, H. The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea. Mar. Drugs 2017, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Domon, B.; Aebersold, R. Mass spectrometry and protein analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Ueberheide, B.M.; Fenyö, D.; Alewood, P.F.; Chait, B.T. Rapid sensitive analysis of cysteine rich peptide venom components. Proc. Natl. Acad. Sci. USA 2009, 106, 6910–6915. [Google Scholar] [CrossRef] [PubMed]
- Phuong, M.A.; Mahardika, G.N.; Alfaro, M.E. Dietary breadth is positively correlated with venom complexity in cone snails. BMC Genom. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nature 2016, 537, 347. [Google Scholar] [CrossRef] [PubMed]
- Petras, D.; Heiss, P.; Süssmuth, R.D.; Calvete, J.J. Venom proteomics of Indonesian king cobra, Ophiophagus hannah: Integrating top-down and bottom-up approaches. J. Proteome Res. 2015, 14, 2539–2556. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Craik, D.J. Bioinformatics-Aided Venomics. Toxins 2015, 7, 2159–2187. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target/Mode of Action | Conotoxin | Clinical Potential | Ref. | |
|---|---|---|---|---|
| Voltage-gated Ion Channels | Cav 2.2 inhibitor | MVIIA | Analgesia (On Market) | [40] |
| Nav 1.8 inhibitor | MrVIB | Analgesia | [45] | |
| Kv inhibitor | PVIIA | Cardiac reperfusion | [42] | |
| Ligand-gated Ion Channels | α9α10 nAChRs inhibitor | Vc1.1 | Analgesia (Phase II) | [43] |
| NMDA-R inhibitor | Conantokin G | Analgesia/anti-epileptic | [17] | |
| 5-HT3 inhibitor | GVIIIA | — | [16] | |
| GPCRs | α1-adrenoceptor inhibitor | TIA | Cardiovascular/Benign Prostate Hyperplasia | [20,46] |
| vasopressin receptor agonist | Conopressin-G | Cardiovascular/mood | [22] | |
| neurotensin receptor agonist | Contulakin-G | Analgesia (Phase Ia) | [23] | |
| Neurotransmitter Transporters | noradrenaline transporter | MrIA | Analgesia (Phase I) | [47] |
| Name | Species | Super-Family | Cystine Pattern | Sequence | Target/IC50 | Year | Ref. |
|---|---|---|---|---|---|---|---|
| RegIIA | C. regius | A | I | GCCSHPACNVNNPHIC # | nAChR: α7/103 nM, α3β2/33 nM, α3β4/97 nM | 2011 | [60] |
| α-LsIA | C. limpusi | - | I | SGCCSNPACRVNNPNIC | nAChRs: α3β2/10 nM, α3α5β2/31 nM, α7/10 nM | 2013 | [87] |
| α-RgIB | C. regius | - | I | TWEECCKNPGCRNNHVDRCRGQV | α3β4 and/or α3β4α5 nAChRs | 2013 | [61] |
| α-BruIB | C. brunneus | - | I | DYCCRROTCIPIC # | Dα7 nAChR | 2014 | [62] |
| α-AusIA | C. australis | - | I | SCCARNPACRHNHPCV | α7 nAChR: 11.68 mM for AusIA (g), 9.67 mM for AusIA (r) | 2014 | [63] |
| Lo1a | C. longurionis | A | I | EGCCSNPACRTNHPEVCD | α7 nAChR/3.24 μM | 2014 | [64] |
| BnIA | C. bandanus | A | I | GCCSHPACSVNNPDIC # | α7 nAChR | 2014 | [65] |
| Im10A | C. imperialis | T | I | NTICCEGCMCY # | unknown | 2016 | [91] |
| α-EIIB | C. ermineus | - | I | ZTOGCCWHPACGKNRC # | nAChRs | 2017 | [66] |
| PIC | C. purpurascens | A | I | SGCCKHPACGKNRC | rα1β1δε nAChR | 2017 | [67] |
| PIC[O7] | SGCCKHOACGKNRC | ||||||
| lt3a | C. litteratus | M | III | DγCCγOQWCDGACDCCS | unknown | 2009 | [68] |
| κ-RIIIJ | C. radiates | M | III | LOSCCSLNLRLCOVOACKRNOCCT # | hKv1.2 channels/33 nM | 2010 | [69] |
| pr3a | C. parius | M | III | CCNWPCSFGCIPCCY | unknown | 2010 | [70] |
| pr3b | ERVCCGYOMSCKSRACKOSYCC # | ||||||
| CnIIIC | C. consors | M | III | ZGCCNGPKGCSSKWCRDHARCC # | Nav1.4/1.3 nM α3β2 nAChR/450 nM | 2012 | [71] |
| BnIIID | C. bandanus | M | III | CCDBγNCDHLCSCCD # | unknown | 2014 | [72] |
| Asi3a | C. asiaticus | M | III | CCQWPCSHGCIPCCY # | unknown | 2016 | [91] |
| bt5a | C. betulinus | T | V | SγCCIRNFLCC | unknown | 2010 | [73] |
| pr6a | C. parius | O | VI/VII | TCLARDELCGASFLSNFLCCDGLCLLICV | unknown | 2010 | [70] |
| pr6b | FGSFIOCAHKGEOCTICCROLRCHEEKTOTCV | ||||||
| pr6c | DQCTYCGIYCCPPKFCTSSGCRSP | ||||||
| pr6d | YGNFOTCSETGEDCSAMHCCRSMTCRNNICAD | ||||||
| MfVIA | C. imperialis | O | VI/VII | RDCQEKWEYCIVPILGFVYCCPGLICGPFVCV | Nav1.8/95.9 nM, Nav1.4/81 nM | 2012 | [88] |
| ge6b | C. geneis | O2O2 | VI/VII | ACGGGGAPCGSSLDCCYPFECSYNSCG | unknown | 2015 | [74] |
| ge6c | VI/VII | ACGGGGAPCGSSLDCCYPFγCSYNSCG | |||||
| PiVIIA | C. princeps | O2 | VI/VII | CDAOTHYCTNYWγCCSGYCγHSHCW | unknown | 2016 | [75] |
| vi6a | C. virgo | O1 | VI/VII | DCGGQGEGCYTQOCCOGLRCRGGGTGGGVCQL | unknown | 2016 | [76] |
| Lo6/7a | C. longurionis | - | VI/VII | DQCSYCGIYCCPPKFCTSAGCRSP # | unknown | 2016 | [91] |
| Lo6/7b | SCLSSGALCGIDSNCCNGCNVPRNQCY # | ||||||
| fu6a | C. fulgetrum | O | VI/VII | TCREKGEOCSVYVγCCSRICGYYACA | unknown | 2016 | [77] |
| α-GVIIIB | C. geographus | S | VIII | SGSTCTCFTSTNCQGSCECLSPPGCYCSNNGIRQPGCSCTCPGT #G | α9α10 nAChR/9.8 nM | 2015 | [92] |
| lt9a | C. litteratus | P | IX | IWFCASRTCSAPADCNPCTCESGVCVDWL | tetrodotoxin-sensi-tive sodium channels/300 nM | 2017 | [78] |
| lt9b | IWFCASRTCSAOADCNOCTCγSGVCVDWL | tetrodotoxin-sensi-tive sodium channels/504 nM | |||||
| Ca11a | C. caracteristicus | I | XI | AWPCGGVRASCSRHDDCCGSLCCFGTSTGCRVAVRPCW | unknown | 2009 | [79] |
| Ca11b | ALLCGGTHARCNRDNDCCGSLCCFGTCISAFVPC | ||||||
| ts14a | C. tessulatus | A | XIV | DGCPPHPVPGMHPCMCTNTC | unknown | 2010 | [80] |
| Asi14a | C. asiaticus | - | XIV | SCGYPCSHCGIPGCYPG # | unknown | 2016 | [92] |
| pc16a | C. pictus | M | XVI | SCSCKRNFLCC # | unknown | 2011 | [81] |
| qc16a | C. quercinus | - | XVI | DCQPCGHNVCC | unknown | 2011 | [82] |
| αD-Ms | C. mustelinus | D | XX | DVRECQVNTPGSKWGKCCMTRMCGTMCCARSGCTCVYHWRRGHGCSCPG | nAChR: α7/0.12 nM, α3β2/1.08 nM, α4β2/4.5 nM | 2009 | [31] |
| αD-Cp | C. capitaneus | D | XX | EVQECQVDTPGSSWGKCCMTRMCGTMCCSRSVCTCVYHWRRGHGCSCPG | showed the same selectivity profile as αD-Ms, but has a lower potency | ||
| α-GeXXA | C. generalis | D | XX | DVHRPCQSVRPGRVWGKCCLTRLCSTMCCARADCTCVYHTWRGHGCSCVM (dimer) | α9α10 nAChR | 2015 | [83] |
| im23a | C. imperialis | K | XXIII | IPYCGQTGAECYSWCIKQDLSKDWCCDFVKDIRMNPPADKCP | unknown | 2012 | [84] |
| im23b | IPYCGQTGAECYSWCIKQDLSKDWCCDFVKTIARLPPAHICSQ | ||||||
| as25a | C. cancellatus | - | XXV | CKCPSCNFNDVTENCKCCIFRQP # | unknown | 2013 | [85] |
| as25b | CKCOSCNFNDVTENCKCCIFRQO? | ||||||
| RsXXIVA | C. regularis | - | XXVI | CKGQSCSSCSTKEFCLSKGSRLMYDCCTGSCCGVKTAGVT | Cav2.2 | 2013 | [89] |
| GeXXVIIA | C. generalis | O | - | ALMSTGTNYRLLKTCRGSGRYCRSPYDCRRRYCRRISDACV | α9α10 nAChR/16.2 nM | 2017 | [93] |
| p21a | C. purpurascens | - | - | FELLPSQDRSCCIQKTLECLENYOGQASQRAHYCQQDATTNCODTYYFGCCPGYATCMSINAGNNVRSAFDKCINRLCFDPGH # | unknown | 2011 | [86] |
| Conotoxin | Super-Family | Primer | Sequence | Target (nAChRs)/IC50 | Ref. |
|---|---|---|---|---|---|
| Pu14.1 | A | signal sequence & 3′-UTR | MGMRMMFAVFLLVVLATTVVSFNSDRASDGRNAAANVKASDLMARVLEKDCPPHPVPGMHKCVCLKTC | rα1β1δε > rα6α3β2 > rα3β2 | [73] |
| GeXIVA | O1 | signal sequence | MKLTCVLIITVLFLTACQLTTAVTYSRGEHKHRALMSTGTNYRLPKTCRSSGRYCRSPYDRRRRYCRRITDACV | rα9α10/4.6 nM | [102] |
| TxIB | - | intron & 3′-UTR | FDGRNTSANNKATDLMALPVRGCCSDPPCRNKHPDLC # | rα6/α3β2β4/28 nM | [103] |
| TxID | - | intron & 3′-UTR | FDGRNAAGNDKMSALMALTTRGCCSHPVCSAMSPIC | rα3β4/12.5 nM, rα6/α3β4/94 nM | [104] |
| LvIA | - | intron & 3′-UTR | FRGRDAAAKASGLVGLTDRRGCCSHPACNVDHPEIC # | rα3β2 (8.7 nM) > rα6/α3β2β3 ≈ rα6/α3β4 ≈ rα3β4 > α7 | [105] |
| Lt1.3 | - | intron & 3′-UTR | FDGRNAAPSDKASDLISLAVRGCCSHPACSGNNPYFC # | α3β2/44.8 nM | [106] |
| VxXXIVA | B | cDNA sequencing | METLTLLWRASSSCLLVVLSHSLLRLLGVRCLEKSGAQPNKLFRPPCCQKGPSFARHSRCVYYTQSRE | rα9α10/1.2 μM, Mouse α1β1γδ/6.6 μM | [107] |
| Species | Number of Precursors | Number of Gene Superfamily | Sequencing Platforms | Number of Confirmed Conotoxins by Proteomics | MS Instruments | Year | Ref. |
|---|---|---|---|---|---|---|---|
| C. textile | - | - | - | 31 | ESI-LTQ-Orbitrap | 2010 | [128] |
| C. bullatus | 30 | 6 | Illumina, Roche 454 | - | - | 2011 | [129] |
| C. consors | 53 | 11 | Roche 454 | - | - | 2012 | [130] |
| C. pulicarius | 82 (79 new) | 14 | Roche 454 | - | - | 2012 | [131] |
| C. marmoreus | 105 | 13 | Roche 454 | 2710–6254 | MALDI-TOF, ESI-Q-TOF | 2013 | [49] |
| C. marmoreus | 158 | 13 new | Roche454 | 106 | ESI-MS/MS | 2013 | [118] |
| C. miles | 662 | 16 (8 new) | Roche 454 | 48 | ESI-Q-TOF | 2013 | [54] |
| C. flavidus | - | - | - | 31 | ESI-LTQ-Orbitrap | 2013 | [53] |
| C. victoriae | 113 | 20 | Roche454 | - | - | 2014 | [119] |
| C. geographus | 127 | 16 (4 new) | Roche454 | 43 | ESI-TripleTOF | 2014 | [3] |
| C. catus | 104 | 11 | Roche 454 | 51 | ESI-Q-TOF | 2015 | [115] |
| C. episcopatus | 3305 | 25 (16 new) | Illumina | 1,448 | ESI-MS/MS ESI-Q-TOF | 2015 | [50] |
| C. tribblei | 136 | 30 (6 new) | Illumina, Roche 454 | - | - | 2015 | [132] |
| C. tribblei C. lenavati | 100 (45 new) 132 | 39 40 | ABI 3730XL | - | - | 2015 | [116] |
| C. planorbis | 182 | 25 | Roche 454 | 23 | ESI-TripleTOF | 2015 | [133] |
| C. betulinus | 215 (183 new) | 9 new | Illumina | - | - | 2016 | [111] |
| C. vexillum C. capitaneus | 220 | 19 (4 new) | Roche 454 | 24 | ESI-Q-TOF, MALDI-TOF | 2016 | [1] |
| C. gloriamaris | 108 (98 new) | 31 | Illumina | - | - | 2017 | [134] |
| Tool | Developer | Function |
|---|---|---|
| Tools for transcriptomics | ||
| SignalP | Technical University of Denmark, Denmark | Predict and locate the signal peptides and their cleavage sites |
| ConoPrec | The university of Queensland, Australia | Identify ORF and analyze contigs coding for conopeptide precursors, predict signal peptides and their cleavage site; Superfamily categorization |
| ConoSorter | — | Identify and classify precursor conotoxins into gene superfamilies; Provide relevant information (frequency of protein sequences, length, number of cysteine residues, hydrophobicity rate of N-terminal region etc.) |
| pHMMs | Technical University of Denmark, Denmark | Identify precursor peptides and classify the sequences into gene superfamily |
| Tools for proteomics | ||
| ConoMass | The university of Queensland, Australia | Match experimental proteomic mass list against the mass predicted from transcripts, mass spectrometry comparison; PTMs identification |
| Mascot | Mascot science, UK | Peptide mass fingerprint; MS/MS database searches |
| ProteinPilot | AB SCIEX, USA | Searching and identification of mass sequences; Identification of PTMs |
| MaxQuant | Max Planck Institute of Biochemistry, Germany | Quantitative analysis of label-free and SILAC-based analysis; PTMs identification |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Li, C.; Dong, S.; Wu, Y.; Zhangsun, D.; Luo, S. Discovery Methodology of Novel Conotoxins from Conus Species. Mar. Drugs 2018, 16, 417. https://doi.org/10.3390/md16110417
Fu Y, Li C, Dong S, Wu Y, Zhangsun D, Luo S. Discovery Methodology of Novel Conotoxins from Conus Species. Marine Drugs. 2018; 16(11):417. https://doi.org/10.3390/md16110417
Chicago/Turabian StyleFu, Ying, Cheng Li, Shuai Dong, Yong Wu, Dongting Zhangsun, and Sulan Luo. 2018. "Discovery Methodology of Novel Conotoxins from Conus Species" Marine Drugs 16, no. 11: 417. https://doi.org/10.3390/md16110417
APA StyleFu, Y., Li, C., Dong, S., Wu, Y., Zhangsun, D., & Luo, S. (2018). Discovery Methodology of Novel Conotoxins from Conus Species. Marine Drugs, 16(11), 417. https://doi.org/10.3390/md16110417