Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss)


 , and
, and
Abstract
1. Introduction
2. Results
2.1. Resveratrol Impairs Body Weight Gain in Rainbow Trout
2.2. Experimental Diets Affect the Fatty Acid Composition in Whole Body Homogenates of Rainbow Trout
2.3. Experimental Diets Affect Lipid Levels and Fatty Acid Composition in Livers of Rainbow Trout
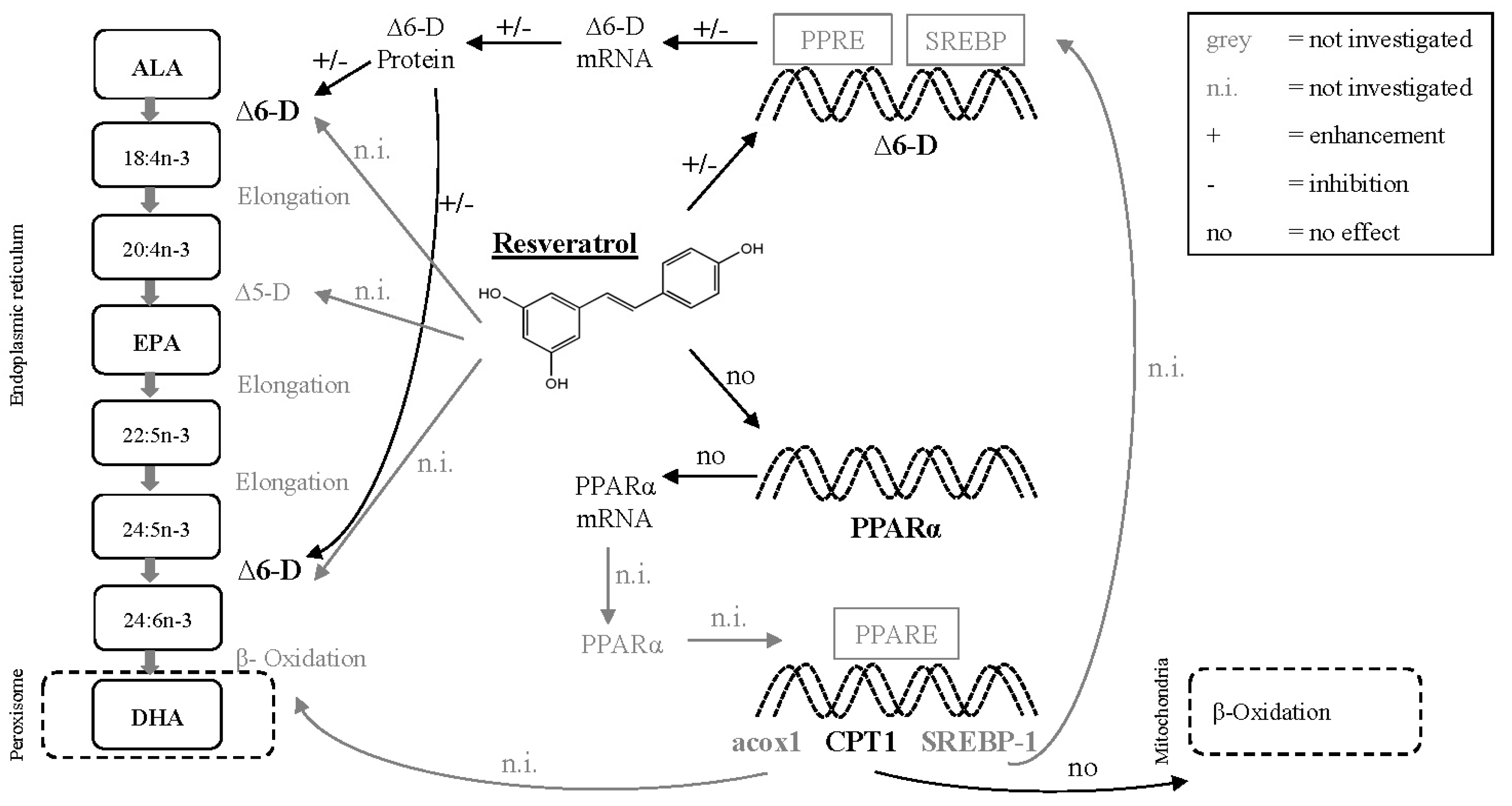
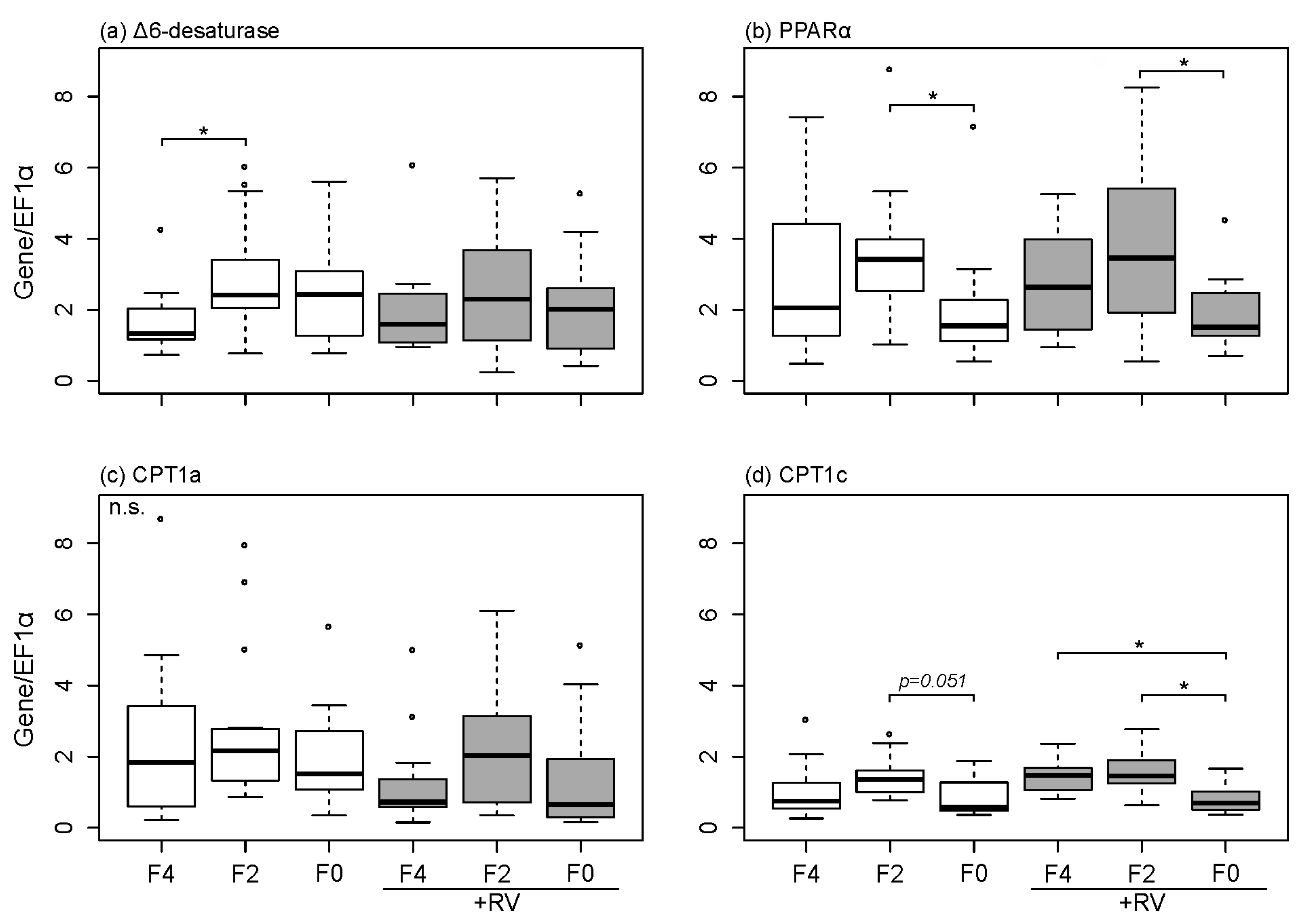
2.4. Dietary Fish Oil Content Affects Hepatic mRNA Expression Levels
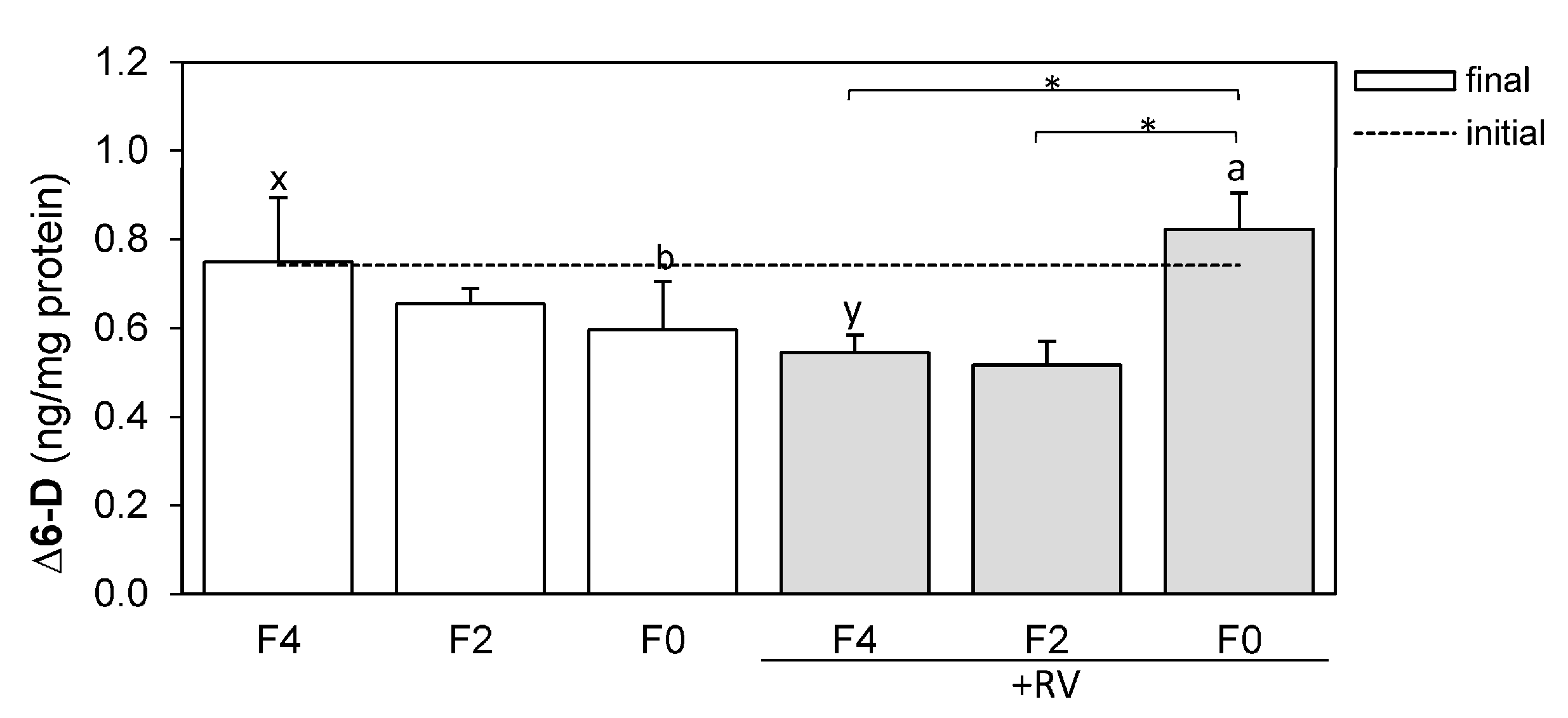
2.5. Resveratrol Affects Hepatic ∆6-Desaturase Protein Levels
3. Discussion
4. Materials and Methods
4.1. Experimental Diets and Housing Conditions of Rainbow Trout
4.2. Sampling
4.3. Lipid Extraction and Measurement of Fatty Acids
4.4. mRNA Extraction and qRT-PCR
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harris, W.S.; Mozaffarian, D.; Lefevre, M.; Toner, C.D.; Colombo, J.; Cunnane, S.C.; Holden, J.M.; Klurfeld, D.M.; Morris, M.C.; Whelan, J. Towards Establishing Dietary Reference Intakes for Eicosapentaenoic and Docosahexaenoic Acids. J. Nutr. 2009, 139, 804–819. [Google Scholar] [CrossRef] [PubMed]
- Fats and Fatty Acids in Human Nutrition-Report of an Expert Consultation; Sanders, T., Ed.; FAO: Rome, Italy, 2010. [Google Scholar]
- Diet, Nutrition and the Prevention of Chronic Diseases; WHO: Geneva, Switzerland, 2003; Volume 916.
- Williams, C.M.; Burdge, G. Long-chain n-3 PUFA: Plant v. marine sources. Proc. Nutr. Soc. 2006, 65, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Molendi-Coste, O.; Legry, V.; Leclercq, I.A. Why and How Meet n-3 PUFA Dietary Recommendations? Gastroenterol. Res. Pract. 2011, 2011, 364040. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- De Lorgeril, M.; Salen, P. New insights into the health effects of dietary saturated and omega-6 and omega-3 polyunsaturated fatty acids. BMC Med. 2012, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Ba, G.N.; Couvreur, P.; Tew, K.D. Polyunsaturated fatty acids (PUFA) and eicosanoids in human health and pathologies. Biomed. Pharmacother. 2002, 56, 215–222. [Google Scholar] [CrossRef]
- Tocher, D.R.; Sargent, J.R. Incorporation into phospholipid classes and metabolism via desaturation and elongation of various 14C-labelled (n-3) and (n-6) polyunsaturated fatty acids in trout astrocytes in primary culture. J. Neurochem. 1990, 54, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Kalinowski, T.; Obach, A.; Robaina, L.; Tort, L.; Caballero, M.J.; Izquierdo, M.S. Vegetable lipid sources for gilthead seabream (Sparus aurata): Effects on fish health. Aquaculture 2003, 225, 353–370. [Google Scholar] [CrossRef]
- Caballero, M.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Bell, J.G.; McEvoy, J.; Tocher, D.R.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar) affects tissue lipid compositions and hepatocyte fatty acid metabolism. J. Nutr. 2001, 131, 1535–1543. [Google Scholar] [PubMed]
- Bell, J.G.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Rapeseed oil as an alternative to marine fish oil in diets of post-smolt Atlantic salmon (Salmo salar): Changes in flesh fatty acid composition and effectiveness of subsequent fish oil “wash out”. Aquaculture 2003, 218, 515–528. [Google Scholar] [CrossRef]
- Regost, C.; Arzel, J.; Robin, J.; Rosenlund, G.; Kaushik, S.J. Total replacement of fish oil by soybean or linseed oil with a return to fish oil in turbot (Psetta maxima) 1. Growth performance, flesh fatty acid profile, and lipid metabolism. Aquaculture 2003, 217, 465–482. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Jackson, A.J. Global fishmeal and fish-oil supply: Inputs, outputs and markets. J. Fish Biol. 2013, 83, 1046–1066. [Google Scholar] [CrossRef] [PubMed]
- The State of World Fisheries and Aquaculture-Opportunities and Challenges; FAO: Rome, Italy, 2014.
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef] [PubMed]
- Turchini, G.M.; Francis, D.S.; Keast, R.S.J.; Sinclair, A.J. Transforming salmonid aquaculture from a consumer to a producer of long chain omega-3 fatty acids. Food Chem. 2011, 124, 609–614. [Google Scholar] [CrossRef]
- Haas, S.; Bauer, J.L.; Adakli, A.; Meyer, S.; Lippemeier, S.; Schwarz, K.; Schulz, C. Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.). J. Appl. Phycol. 2015. [Google Scholar] [CrossRef]
- Sprague, M.; Walton, J.; Campbell, P.J.; Strachan, F.; Dick, J.R.; Bell, J.G. Replacement of fish oil with a DHA-rich algal meal derived from Schizochytrium sp. on the fatty acid and persistent organic pollutant levels in diets and flesh of Atlantic salmon (Salmo salar, L.) post-smolts. Food Chem. 2015, 185, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Betiku, O.C.; Barrows, F.T.; Ross, C.; Sealey, W.M. The effect of total replacement of fish oil with DHA-Gold® and plant oils on growth and fillet quality of rainbow trout (Oncorhynchus mykiss) fed a plant based diet. Aquac. Nutr. 2016, 22, 158–169. [Google Scholar] [CrossRef]
- Betancor, M.B.; Sprague, M.; Sayanova, O.; Usher, S.; Campbell, P.J.; Napier, J.A.; Caballero, M.J.; Tocher, D.R. Evaluation of a high-EPA oil from transgenic Camelina sativa in feeds for Atlantic salmon (Salmo salar L.): Effects on tissue fatty acid composition, histology and gene expression. Aquaculture 2015, 444, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Trattner, S.; Kamal-Eldin, A.; Brännäs, E.; Moazzami, A.; Zlabek, V.; Larsson, P.; Ruyter, B.; Gjøen, T.; Pickova, J. Sesamin supplementation increases white muscle docosahexaenoic acid (DHA) levels in rainbow trout (Oncorhynchus mykiss) fed high alpha-linolenic acid (ALA) containing vegetable oil: Metabolic actions. Lipids 2008, 43, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Schiller Vestergren, A.L.; Trattner, S.; Pan, J.; Johnsson, P.; Kamal-Eldin, A.; Brännäs, E.; Moazzami, A.A.; Pickova, J. The effect of combining linseed oil and sesamin on the fatty acid composition in white muscle and on expression of lipid-related genes in white muscle and liver of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2012, 21, 843–859. [Google Scholar] [CrossRef]
- Sargent, J.; Bell, G.; McEvoy, L.; Tocher, D.; Estevez, A. Recent developments in the essential fatty acid nutrition of fish. Aquaculture 1999, 177, 191–199. [Google Scholar] [CrossRef]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The lipids. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 181–257. [Google Scholar]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. 2003, 11, 7–184. [Google Scholar] [CrossRef]
- Burdge, G. α-Linolenic acid metabolism in men and women: Nutritional and biological implications. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Vagner, M.; Santigosa, E. Characterization and modulation of gene expression and enzymatic activity of delta-6 desaturase in teleosts: A review. Aquaculture 2011, 315, 131–143. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R.; Porter, A.E.A. Biosynthesis and tissue deposition of docosahexaenoic acid (22:6n-3) in rainbow trout (Oncorhynchus mykiss). Lipids 2001, 36, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Thiessen, D.L.; Maenz, D.D.; Newkirk, R.W.; Classen, H.L.; Drew, M.D. Replacement of fishmeal by canola protein concentrate in diets fed to rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2004, 10, 379–388. [Google Scholar] [CrossRef]
- Richard, N.; Kaushik, S.; Larroquet, L.; Panserat, S.; Corraze, G. Replacing dietary fish oil by vegetable oils has little effect on lipogenesis, lipid transport and tissue lipid uptake in rainbow trout (Oncorhynchus mykiss). Br. J. Nutr. 2006, 96, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Lazzarotto, V.; Corraze, G.; Leprevost, A.; Quillet, E.; Dupont-Nivet, M.; Médale, F. Three-year breeding cycle of rainbow trout (Oncorhynchus mykiss) fed a plant-based diet, totally free of marine resources: Consequences for reproduction, fatty acid composition and progeny survival. PLoS ONE 2015, 10, e0117609. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Fish and Shrimp; National Research Council, Ed.; The National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-16338-5. [Google Scholar]
- Blanchet, C.; Lucas, M.; Julien, P.; Morin, R.; Gingras, S.; Dewailly, É. Fatty acid composition of wild and farmed Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss). Lipids 2005, 40, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Report of the Sub-Committee on Recommendations for Intake of Polyunsaturated Fatty Acids in Healthy Adults; International Society for the Study of Faty Acids and Lipids: Brighton, UK, 2004.
- Schiller Vestergren, A.L.; Trattner, S.; Mráz, J.; Ruyter, B.; Pickova, J. Fatty acids and gene expression responses to bioactive compounds in Atlantic salmon (Salmo salar L.) hepatocytes. Neuroendocrinol. Lett. 2011, 32 (Suppl. 2), 41–50. [Google Scholar] [PubMed]
- Trattner, S.; Ruyter, B.; Østbye, T.K.; Gjøen, T.; Zlabek, V.; Kamal-Eldin, A.; Pickova, J. Sesamin increases alpha-linolenic acid conversion to docosahexaenoic acid in atlantic salmon (Salmo salar L.) hepatocytes: Role of altered gene expression. Lipids 2008, 43, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Soleas, G.J.; Diamandis, E.P.; Goldberg, D.M. Resveratrol: A molecule whose time has come? And gone? Clin. Biochem. 1997, 30, 91–113. [Google Scholar] [CrossRef]
- Brisdelli, F.; D’Andrea, G.; Bozzi, A. Resveratrol: A natural polyphenol with multiple chemopreventive properties. Curr. Drug Metab. 2009, 10, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Namkoong, K.; Shin, M.; Park, J.; Yang, E.; Ihm, J.; Thu, V.T.; Kim, H.K.; Han, J. Cardiovascular Protective Effects and Clinical Applications of Resveratrol. J. Med. Food 2017, 20, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Arichi, H.; Kimura, Y.; Okuda, H.; Baba, K.; Kozawa, M.; Arichi, S. Effects of Stilbene Components of the Roots of Polygonum cuspidatum Sieb. et Zucc. on Lipid Metabolism. Chem. Pharm. Bull. 1982, 30, 1766–1770. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Ohminami, H.; Okuda, H.; Kimiye, B.; Mitsugi, K.; Arichi, S. Effects of Stilbene Components of Roots of Polygonum ssp. on Liver Injury in Peroxidized Oil-fed Rats. Planta Med. 1983, 49, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhang, X.; Gao, D.; Jiang, X.; Dong, W. Resveratrol inhibits MMP-9 expression by up-regulating PPAR alpha expression in an oxygen glucose deprivation-exposed neuron model. Neurosci. Lett. 2009, 451, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Nakata, R.; Takahashi, S.; Inoue, H. Recent advances in the study on resveratrol. Biol. Pharm. Bull. 2012, 35, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Jiang, X.F.; Katayama, T.; Osada, S.; Umesono, K.; Namura, S. Brain protection by resveratrol and fenofibrate against stroke requires peroxisome proliferator-activated receptor α in mice. Neurosci. Lett. 2003, 352, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Nakata, R.; Tamura, E.; Kosuge, Y.; Kariya, A.; Katsukawa, M.; Mishima, S.; Ito, T.; Iinuma, M.; Akao, Y.; et al. Vaticanol C, a resveratrol tetramer, activates PPARα and PPARβ/δ in vitro and in vivo. Nutr. Metab. (Lond.) 2010, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Momchilova, A.; Petkova, D.; Staneva, G.; Markovska, T.; Pankov, R.; Skrobanska, R.; Nikolova-Karakashian, M.; Koumanov, K. Resveratrol alters the lipid composition, metabolism and peroxide level in senescent rat hepatocytes. Chem. Biol. Interact. 2014, 207, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Caro, M.; Sansone, A.; Amézaga, J.; Navarro, V.; Ferreri, C.; Tueros, I. Wine lees modulate lipid metabolism and induce fatty acid remodelling in Zebrafish. Food Funct. 2017. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Madrigal, J.; Karalazos, V.; Campbell, P.J.; Bell, J.G.; Tocher, D.R. Influence of dietary palm oil on growth, tissue fatty acid compositions, and fatty acid metabolism in liver and intestine in rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2005, 11, 241–250. [Google Scholar] [CrossRef]
- Drew, M.D.; Ogunkoya, A.E.; Janz, D.M.; Van Kessel, A.G. Dietary influence of replacing fish meal and oil with canola protein concentrate and vegetable oils on growth performance, fatty acid composition and organochlorine residues in rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 267, 260–268. [Google Scholar] [CrossRef]
- Emery, J.A.; Hermon, K.; Hamid, N.K.A.; Donald, J.A.; Turchini, G.M. Δ-6 Desaturase substrate competition: Dietary linoleic acid (18:2n-6) has only trivial effects on α-linolenic acid (18:3n-3) bioconversion in the teleost rainbow trout. PLoS ONE 2013, 8, e57463. [Google Scholar] [CrossRef] [PubMed]
- Sissener, N.H.; Sanden, M.; Torstensen, B.E.; Waagbø, R.; Stubhaug, I.; Rosenlund, G. High dietary 18:2n-6/18:3n-3 ratio does not inhibit elongation and desaturation of 18:3n-3 to EPA and DHA in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2016, 1–11. [Google Scholar] [CrossRef]
- Ajiboye, O.O.; Yakubu, A.F.; Adams, T.E. A Perspective on the Ingestion and Nutritional Effects of Feed Additives in Farmed Fish Species. World J. Fish Mar. Sci. 2012, 4, 87–101. [Google Scholar] [CrossRef]
- Bureau, D.P.; Harris, A.M.; Cho, C.Y. The effects of purified alcohol extracts from soy products on feed intake and growth of chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss). Aquaculture 1998, 161, 27–43. [Google Scholar] [CrossRef]
- Peragón, J.; Barroso, J.B.; García-Salguero, L.; de la Higuera, M.; Lupiáñez, J.A. Dietary alterations in protein, carbohydrates and fat increase liver protein-turnover rate and decrease overall growth rate in the rainbow trout (Oncorhynchus mykiss). Mol. Cell. Biochem. 2000, 209, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Menoyo, D.; López-Bote, C.J.; Obach, A.; Bautista, J.M. Effect of dietary fish oil substitution with linseed oil on the performance, tissue fatty acid profile, metabolism, and oxidative stability of Atlantic salmon. J. Anim. Sci. 2005, 83, 2853–2862. [Google Scholar] [CrossRef] [PubMed]
- Kjær, M.A.; Vegusdal, A.; Gjøen, T.; Rustan, A.C.; Todorčević, M.; Ruyter, B. Effect of rapeseed oil and dietary n-3 fatty acids on triacylglycerol synthesis and secretion in Atlantic salmon hepatocytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2008, 1781, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Michel, F.; Fouret, G.; Casas, F.; Jullien, M.; Wrutniak-Cabello, C.; Ramos, J.; Cristol, J.-P.; Coudray, C.; Carbonneau, M.-A.; et al. A polyphenol extract modifies quantity but not quality of liver fatty acid content in high-fat-high-sucrose diet-fed rats: Possible implication of the sirtuin pathway. Br. J. Nutr. 2010, 104, 1760–1770. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.D.; Abraham, S. Gene expression: Nutrient control of pre- and posttranscriptional events. J. Fed. Am. Soc. Exp. Biol. 1992, 6, 3146–3152. [Google Scholar] [PubMed]
- Walle, T.; Hsieh, F.; Delegge, M.H.; Oatis, J.E.; Walle, U.K. High absortion but very low bioavaibility of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Delmas, D.; Aires, V.; Limagne, E.; Dutartre, P.; Mazué, F.; Ghiringhelli, F.; Latruffe, N. Transport, stability, and biological activity of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Frémont, L.; Belguendouz, L.; Delpal, S. Antioxidant activity of resveratrol and alcohol-free wine polyphenols related to LDL oxidation and polyunsaturated fatty acids. Life Sci. 1999, 64, 2511–2521. [Google Scholar] [CrossRef]
- Urquiaga, I.; Guasch, V.; Marshall, G.; San Martín, A.; Castillo, Ó.; Rozowski, J.; Leighton, F. Effect of Mediterranean and Occidental diets, and red wine, on plasma fatty acids in humans. An intervention study. Biol. Res. 2004, 37, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Natella, F.; Ghiselli, A.; Alessia, G.; Ursini, F.; Scaccini, C. Red wine mitigates the postprandial increase of LDL susceptibility to oxidation. Free Radic. Biol. Med. 2001, 30, 1036–1044. [Google Scholar] [CrossRef]
- Wu, J.M.; Hsieh, T. Resveratrol: A cardioprotective substance. Ann. N. Y. Acad. Sci. 2011, 1215, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-J.; Wang, Q.; Lv, Z.-M.; Wang, C.-L.; Li, C.-P.; Rong, Y.-L. Resveratrol appears to protect against oxidative stress and steroidogenesis collapse in mice fed high-calorie and high-cholesterol diet. Andrologia 2015, 47, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome Proliferator-Activated Receptors: Nuclear Control of Metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Tejera, N.; O’Neill, C.; Booz, V.; Jude, B.; Wolf, I.M.A.; Rigby, N.; Silvan, J.M.; Curtis, P.J.; Cassidy, A.; et al. Anthocyanins do not influence long-chain n-3 fatty acid status: Studies in cells, rodents and humans. J. Nutr. Biochem. 2015, 26, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Noh, K.; Han, C.Y.; Kim, S.G. Transactivation of genes encoding for phase II enzymes and phase III transporters by phytochemical antioxidants. Molecules 2010, 15, 6332–6348. [Google Scholar] [CrossRef] [PubMed]
- Baile, C.A.; Yang, J.Y.; Rayalam, S.; Hartzell, D.L.; Lai, C.Y.; Andersen, C.; Della-Fera, M.A. Effect of resveratrol on fat mobilization. Ann. N. Y. Acad. Sci. 2011, 1215, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, H.; Pereira, T.C.B.; Rico, E.P.; Rosemberg, D.B.; Bonan, C.D.; Bogo, M.R.; Souto, A.A. Modulatory effect of resveratrol on SIRT1, SIRT3, SIRT4, PGC1α and NAMPT gene expression profiles in wild-type adult zebrafish liver. Mol. Biol. Rep. 2012, 39, 3281–3289. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Chang, E.J.; Bae, S.J.; Shim, S.M.; Park, H.D.; Rhee, C.H.; Park, J.H.; Choi, S.W. Cytotoxic and antimutagenic stilbenes from seeds of Paeonia lactiflora. Arch. Pharm. Res. 2002, 25, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.; Hanson, S.; Malison, J.A.; Wentworth, B.; Barry, T.P. Genistein and other isoflavones found in soybeans inhibit estrogen metabolism in salmonid fish. Aquaculture 2006, 254, 658–665. [Google Scholar] [CrossRef]
- Gao, J.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Edward, R.; Mamauag, P.; Han, Y. Effects of dietary oxidized fish oil with vitamin E supplementation on growth performance and reduction of lipid peroxidation in tissues and blood of red sea bream Pagrus major. Aquaculture 2012, 356–357, 73–79. [Google Scholar] [CrossRef]
- Faizan, M.; Stubhaug, I.; Menoyo, D.; Esatbeyoglu, T.; Wagner, A.E.; Struksnæs, G.; Koppe, W.; Rimbach, G. Dietary Alpha-Tocopherol Affects Tissue Vitamin E and Malondialdehyde Levels but Does not Change Antioxidant Enzymes and Fatty Acid Composition in Farmed Atlantic Salmon (Salmo salar L.). Int. J. Vitam. Nutr. Res. 2013, 83, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Aluru, N.; Vijayan, M.M. Aryl hydrocarbon receptor activation impairs cortisol response to stress in rainbow trout by disrupting the rate-limiting steps in steroidogenesis. Endocrinology 2006, 147, 1895–1903. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, G. Effects of resveratrol on longevity, cognitive ability and aging-related histological markers in the annual fish Nothobranchius guentheri. Exp. Gerontol. 2012, 47, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.N.; Baumgarner, B.L.; Watanabe, W.O.; Alam, M.S.; Kinsey, S.T. Effects of resveratrol on growth and skeletal muscle physiology of juvenile southern flounder. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 183, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Fontana, S.; Laforgia, F.; Dragone, T.; Jirillo, E.; Passantino, L. Administration of a Polyphenol-Enriched Feed to Farmed Sea Bass (Dicentrarchus labrax L.) Modulates Intestinal and Spleen Immune Responses. Oxid. Med. Cell. Longev. 2016, 2016, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Rodehutscord, M.; Becker, A.; Pack, M.; Pfeffer, E. Response of rainbow trout (Oncorhynchus mykiss) to supplements of individual essential amino acids in a semipurified diet, including an estimate of the maintenance requirement for essential amino acids. J. Nutr. 1997, 127, 1166–1175. [Google Scholar] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Bretz, F.; Hothorn, T.; Westfall, P. Multiple Comparisons Using R; CRC Press: Boca Raton, FL, USA, 2011; Volume 6, ISBN 9781584885740. [Google Scholar]
- Schaarschmidt, F.; Vaas, L. Analysis of trials with complex treatment structure using multiple contrast tests. HortScience 2009, 44, 188–195. [Google Scholar]
- Laird, N.M.; Ware, J.H. Random-Effects Models for Longitudinal Data. Biometrics 1982, 38, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, G.; Molenberghs, G. Linear Mixed Models for Longitudinal Data; Springer: New York, NY, USA, 2000; ISBN 9781441913050. [Google Scholar]
- Konietschke, F.; Hothorn, L.A.; Brunner, E. Rank-based multiple test procedures and simultaneous confidence intervals. Electron. J. Stat. 2012, 6, 738–759. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| F4 | F2 | F0 | F4 + RV | F2 + RV | F0 + RV | |
|---|---|---|---|---|---|---|
| IBW 1 | 35.54 ± 0.20 | 35.34 ± 0.15 | 35.69 ± 0.42 | 35.66 ± 0.15 | 35.81 ± 0.18 | 35.63 ± 0.50 |
| FBW 2 | 46.15 ± 1.26 m | 46.37 ± 0.18 x | 47.27 ± 2.19 a | 41.96 ± 0.90 n | 41.60 ± 1.15 y | 40.57 ± 1.62 b |
| DFI 3 | 1.94 ± 0.07 m | 1.95 ± 0.06 x | 1.90 ± 0.11 a | 1.49 ± 0.04 n | 1.49 ± 0.07 y | 1.36 ± 0.08 b |
| HSI 4 | 1.52 ± 0.40 | 1.49 ± 0.23 | 1.40 ± 0.16 | 1.32 ± 0.24 | 1.28 ± 0.30 | 1.51 ± 0.34 |
| [% of FAMEs] | F4 | F2 | F0 | F4 + RV | F2 + RV | F0 + RV |
|---|---|---|---|---|---|---|
| 14:0 | 1.79 | 1.04 | 0.70 | 1.77 | 1.04 | 0.35 |
| 16:0 | 11.24 | 9.75 | 9.76 | 11.22 | 9.74 | 8.89 |
| 18:0 | 3.58 | 3.18 | 3.17 | 3.55 | 3.11 | 2.95 |
| Σ SFA a | 16.60 | 13.97 | 13.64 | 16.54 | 13.90 | 12.20 |
| 16:1 | 1.72 | 1.09 | 0.83 | 1.73 | 1.08 | 0.52 |
| 18:1n-9c | 33.46 | 40.03 | 42.00 | 33.38 | 39.67 | 45.18 |
| 18:1n-7c | 2.65 | 2.92 | 3.14 | 2.68 | 2.99 | 3.21 |
| Σ MUFA b | 37.83 | 44.04 | 45.95 | 37.79 | 43.74 | 48.91 |
| 18:2n-6c | 28.83 | 28.26 | 27.90 | 28.87 | 28.61 | 27.79 |
| 18:3n-3 | 10.95 | 10.11 | 9.29 | 11.00 | 10.07 | 8.93 |
| 20:5n-3 | 2.28 | 1.44 | 1.20 | 2.35 | 1.40 | 0.68 |
| 22:5n-3 | 0.49 | 0.18 | 0.27 | 0.32 | 0.33 | 0.21 |
| 22:6n-3 | 3.02 | 2.00 | 1.80 | 3.12 | 1.96 | 1.27 |
| Σ PUFA c | 45.57 | 41.99 | 40.42 | 45.67 | 42.37 | 38.89 |
| Σ EPA + DHA d | 5.30 | 3.43 | 2.35 | 5.47 | 3.36 | 1.96 |
| EPA/DHA | 0.75 | 0.72 | 0.61 | 0.75 | 0.72 | 0.54 |
| 18:3n-3/18:2n-6 | 0.38 | 0.36 | 0.33 | 0.38 | 0.35 | 0.32 |
| Σ EPA + DHA % DM e | 0.74 | 0.56 | 0.34 | 0.91 | 0.56 | 0.26 |
| [% of FAMEs] | Initial | F4 | F2 | F0 | F4 + RV | F2 + RV | F0 + RV |
|---|---|---|---|---|---|---|---|
| 14:0 | 4.97 | 3.67 ± 0.29 | 3.29 ± 0.49 | 3.47 ± 0.26 a | 3.19 ± 0.32 A | 2.47 ± 0.19 A,B | 1.73 ± 0.69 b,B |
| 16:0 | 17.73 | 19.44 ± 0.84 | 20.15 ± 1.50 | 19.26 ± 0.52 | 18.54 ± 1.62 | 19.57 ± 0.47 | 20.16 ± 0.63 |
| 18:0 | 4.31 | 5.78 ± 0.48 | 6.26 ± 0.73 | 5.95 ± 0.47 b | 5.73 ± 0.96 B | 6.59 ± 0.34 A,B | 7.96 ± 0.73 a,A |
| Σ SFA a | 27.01 | 28.89 ± 1.05 | 29.70 ± 2.09 | 28.68 ± 0.73 | 27.46 ± 2.71 | 28.63 ± 0.65 | 29.85 ± 0.71 |
| 16:1 | 5.05 | 4.18 ± 0.52 | 3.70 ± 0.32 x | 3.75 ± 0.29 a | 3.50 ± 0.41 A | 2.71 ± 0.13 y,A,B | 2.15 ± 0.69 b,B |
| 18:1n-9c | 27.25 | 25.03 ± 0.91 | 23.94 ± 3.65 | 25.88 ± 1.77 a | 27.46 ± 6.18 | 20.48 ± 0.53 | 17.91 ± 1.30 b |
| 18:1n-7c | 3.36 | 3.12 ± 0.02 | 3.10 ± 0.03 + | 3.17 ± 0.11 a | 3.17 ± 0.21 A | 2.8 ± 0.08 A,+ | 2.48 ± 0.25 b,B |
| Σ MUFA b | 35.66 | 32.33 ± 1.34 | 30.74 ± 3.72 | 32.80 ± 2.11 a | 34.13 ± 6.61 A | 26.00 ± 0.61 B | 22.55 ± 2.24 b,B |
| 18:2n-6c | 14.11 | 11.81 ± 1.63 | 10.31 ± 2.67 | 11.41 ± 1.13 a | 12.50 ± 3.44 A | 8.72 ± 0.12 A,B | 5.46 ± 1.63 b,B |
| 18:3n-3 | 3.44 | 2.28 ± 0.35 | 1.92 ± 0.47 | 2.02 ± 0.21 a | 2.66 ± 0.73 A | 1.46 ± 0.10 B | 0.97 ± 0.26 b,B |
| 20:5n-3 | 4.29 | 3.91 ± 0.10 | 4.03 ± 0.63 | 3.74 ± 0.28 b | 3.65 ± 1.13 B | 4.68 ± 0.20 A,B | 5.54 ± 0.41 a,A |
| 22:5n-3 | 1.97 | 2.24 ± 0.05 | 2.28 ± 0.40 | 2.11 ± 0.12 + | 1.95 ± 0.71 B | 2.57 ± 0.04 A,B | 2.83 ± 0.21 A,+ |
| 22:6n-3 | 13.53 | 18.55 ± 2.09 | 21.02 ± 3.88 + | 19.24 ± 2.33 b | 17.65 ± 6.16 B | 27.95 ± 0.84 A,+ | 32.81 ± 3.11 a,A |
| Σ PUFA c | 37.34 | 38.78 ± 0.65 | 39.56 ± 1.74 y | 38.52 ± 1.39 b | 38.41 ± 3.91 B | 45.38 ± 0.93 x,A | 47.60 ± 1.55 a,A |
| Σ EPA + DHA d | 17.82 | 22.46 ± 2.19 | 25.05 ± 4.48 | 22.98 ± 2.60 b | 21.29 ± 7.29 B | 32.63 ± 0.89 A | 38.35 ± 3.38 a,A |
| EPA/DHA | 0.32 | 0.21 ± 0.02 | 0.19 ± 0.01 + | 0.20 ± 0.01 ++ | 0.21 ± 0.01 A | 0.17 ± 0.01 B,+ | 0.17 ± 0.01 B,++ |
| Σ n-3/Ʃ n-6 | 1.65 | 2.33 ± 0.45 | 3.03 ± 1.14 | 2.42 ± 0.57 b | 2.34 ± 1.45 B | 4.20 ± 0.07 B | 8.24 ± 2.61 a,A |
| Initial | F4 | F2 | F0 | F4 + RV | F2 + RV | F0 + RV | |
|---|---|---|---|---|---|---|---|
| Total fat [mg/g] | 13.60 | 16.70 ± 1.56 N,n | 21.47 ± 3.10 x | 26.80 ± 2.55 M | 24.80 ± 1.06 m | 30.08 ± 3.11 y | 26.70 ± 0.99 |
| FA [% of FAMEs] | |||||||
| 14:0 | 1.63 | 1.45 ± 0.19 | 1.24 ± 0.13 | 1.33 ± 0.15 | 1.32 ± 0.08 | 1.4 ± 0.15 | 1.33 ± 0.11 |
| 16:0 | 20.75 | 22.80 ± 1.01 | 24.07 ± 1.35 | 24.30 ± 1.01 a | 23.01 ± 0.6 | 22.89 ± 2.61 | 21.28 ± 0.71 b |
| 18:0 | 6.90 | 8.14 ± 0.59 | 8.66 ± 0.58 | 8.15 ± 0.38 | 7.74 ± 0.38 | 7.52 ± 1.48 | 7.12 ± 0.18 |
| Σ SFA a | 29.28 | 32.39 ± 1.22 | 33.96 ± 1.79 | 33.78 ± 1.11 a | 32.06 ± 0.89 | 31.81 ± 3.95 | 29.74 ± 0.79 b |
| 16:1 | 2.20 | 1.64 ± 0.21 | 1.46 ± 0.25 | 1.92 ± 0.33 | 1.56 ± 0.25 | 1.91 ± 0.53 | 1.70 ± 0.32 |
| 18:1n9c | 14.93 | 18.60 ± 4.28 | 15.14 ± 1.39 | 14.68 ± 1.00 | 14.58 ± 0.78 | 15.61 ± 1.24 | 16.40 ± 1.64 |
| 18:1n7c | 2.37 | 2.55 ± 0.29 | 2.36 ± 0.23 | 2.66 ± 0.46 | 2.46 ± 0.33 | 2.72 ± 0.37 | 2.73 ± 0.29 |
| Σ MUFA b | 19.50 | 22.79 ± 4.73 | 18.96 ± 1.86 | 19.25 ± 1.62 | 18.60 ± 1.23 | 20.25 ± 2.13 | 20.83 ± 2.19 |
| 18:2n6c | 8.17 | 6.09 ± 0.99 | 6.05 ± 0.99 | 5.74 ± 0.27 | 6.27 ± 0.62 | 5.73 ± 0.52 | 6.33 ± 0.35 |
| 18:3n3 | 1.45 | 0.69 ± 0.24 | 0.56 ± 0.16 | 0.47 ± 0.11 | 0.58 ± 0.08 | 0.57 ± 0.13 | 0.57 ± 0.20 |
| 20:5n3 | 4.59 | 3.90 ± 0.25 | 3.55 ± 0.37 | 3.65 ± 0.76 | 4.43 ± 0.41 | 4.37 ± 0.81 | 4.61 ± 0.64 |
| 22:5n3 | 1.87 | 1.87 ± 0.21 + | 1.88 ± 0.18 | 1.89 ± 0.22 b | 2.30 ± 0.23 + | 2.22 ± 0.22 | 2.47 ± 0.19 a |
| 22:6n3 | 35.13 | 32.26 ± 4.40 | 35.03 ± 1.48 | 35.21 ± 1.18 | 35.77 ± 0.93 | 35.06 ± 1.71 | 35.45 ± 1.89 |
| Σ PUFA c | 51.21 | 44.81 ± 3.51 | 47.08 ± 0.21 | 46.97 ± 1.96 | 49.34 ± 0.72 | 47.95 ± 2.49 | 49.43 ± 1.79 |
| Σ EPA + DHA d | 39.72 | 36.16 ± 4.50 | 38.58 ± 1.43 | 38.86 ± 1.68 | 40.20 ± 0.63 | 39.44 ± 2.46 | 40.06 ± 1.77 |
| EPA/DHA | 0.13 | 0.12 ± 0.02 | 0.10 ± 0.01 | 0.10 ± 0.02 | 0.12 ± 0.01 | 0.12 ± 0.02 | 0.13 ± 0.02 |
| Σ n-3/Σ n-6 | 5.27 | 6.54 ± 1.67 | 6.92 ± 1.31 | 7.19 ± 0.45 | 6.92 ± 0.77 | 7.37 ± 1.03 | 6.83 ± 0.60 |
| EPA mg/g | 0.58 | 0.72 ± 0.24 + | 0.70 ± 0.03 y | 0.83 ± 0.28 b,+ | 1.02 ± 0.08 | 1.04 ± 0.39 x | 1.29 ± 0.23 a |
| DHA mg/g | 4.44 | 4.79 ± 1.19 N | 7.00 ± 1.18 y | 7.86 ± 1.52 b,M | 8.25 ± 0.42 | 10.20 ± 0.68 x | 11.08 ± 2.75 a |
| Σ EPA + DHA mg/g e | 5.02 | 5.51 ± 1.23 N | 7.71 ± 1.20 y | 8.69 ± 1.78 b,M | 9.27 ± 0.37 | 11.24 ± 0.78 x | 12.38 ± 2.93 a |
| Ingredients [% DM] | F4 | F2 | F0 | F4 + RV | F2 + RV | F0 + RV |
|---|---|---|---|---|---|---|
| Fish meal (Clupea sp.) a | 10 | 10 | 10 | 10 | 10 | 10 |
| Casein b | 5.9 | 5.9 | 5.9 | 5.9 | 5.9 | 5.9 |
| Rapeseed concentrate c | 13 | 13 | 13 | 13 | 13 | 13 |
| Pea protein isolate d | 13.58 | 13.58 | 13.58 | 13.58 | 13.58 | 13.58 |
| Wheat gluten e | 17.95 | 17.95 | 17.95 | 17.95 | 17.95 | 17.95 |
| Wheat starch e | 20 | 20 | 20 | 20 | 20 | 20 |
| Fish oil a | 4 | 2 | 4 | 2 | ||
| Linseed oil f | 1.50 | 0.94 | 0.38 | 1.50 | 0.94 | 0.38 |
| Rapeseed oil g | 3.19 | 6.11 | 9.04 | 3.19 | 6.11 | 9.04 |
| Sunflower oil g | 1.91 | 1.55 | 1.18 | 1.91 | 1.55 | 1.18 |
| Vitamin mineral premix h | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Lysine i | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 |
| Dicalcium-phosphate j | 1 | 1 | 1 | 1 | 1 | 1 |
| Inert filler k | 6.77 | 6.77 | 6.77 | 6.77 | 6.77 | 6.77 |
| Resveratrol (RV) l | 0.3 | 0.3 | 0.3 | |||
| Nutrient composition [% DM] | ||||||
| Dry matter (in % of diet) | 93.2 | 93.1 | 93.2 | 93.3 | 92.8 | 91.7 |
| Crude protein | 50.1 | 50.3 | 50.2 | 50.0 | 50.3 | 50.8 |
| Crude lipid | 14.9 | 15.1 | 15.1 | 15.0 | 15.0 | 15.0 |
| Crude ash | 6.4 | 6.4 | 6.4 | 6.4 | 6.4 | 6.2 |
| Total KOH m | 28.6 | 28.2 | 28.4 | 28.6 | 28.3 | 28.0 |
| Gross energy [MJ kg−1 DM] | 22.72 | 22.72 | 22.70 | 22.75 | 22.75 | 22.93 |
| Primer | Sequence 5′ → 3′ | Annealing Temperature (°C) |
|---|---|---|
| ∆6-D a,* forward | GCTGGAGARGATGCCACGGA | 61 |
| ∆6-D a,* reverse | TGCCAGCTCTCCAATCAGCA | 61 |
| EF1α b,* forward | ACAAGCCCCTYCGTCTGCC | 61 |
| EF1α b,* reverse | GCATCTCCACAGACTTSACCTCAG | 61 |
| PPARα c,§ forward | CTGGAGCTGGATGACAGTGA | 55 |
| PPARα c,§ reverse | GGCAAGTTTTTGCAGCAGAT | 55 |
| CPT1a d,§ forward | TCGATTTTCAAGGGTCTTCG | 55 |
| CPT1a d,§ reverse | CACAACGATCAGCAAACTGG | 55 |
| CPT1c d,§ forward | CGCTTCAAGAATGGGGTGAT | 59 |
| CPT1c d,§ reverse | CAACCACCTGCTGTTTCTCA | 59 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torno, C.; Staats, S.; Pascual-Teresa, S.d.; Rimbach, G.; Schulz, C. Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss). Mar. Drugs 2017, 15, 252. https://doi.org/10.3390/md15080252
Torno C, Staats S, Pascual-Teresa Sd, Rimbach G, Schulz C. Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss). Marine Drugs. 2017; 15(8):252. https://doi.org/10.3390/md15080252
Chicago/Turabian StyleTorno, Claudia, Stefanie Staats, Sonia de Pascual-Teresa, Gerald Rimbach, and Carsten Schulz. 2017. "Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss)" Marine Drugs 15, no. 8: 252. https://doi.org/10.3390/md15080252
APA StyleTorno, C., Staats, S., Pascual-Teresa, S. d., Rimbach, G., & Schulz, C. (2017). Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss). Marine Drugs, 15(8), 252. https://doi.org/10.3390/md15080252