Biotechnological Potential of Cold Adapted Pseudoalteromonas spp. Isolated from ‘Deep Sea’ Sponges

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
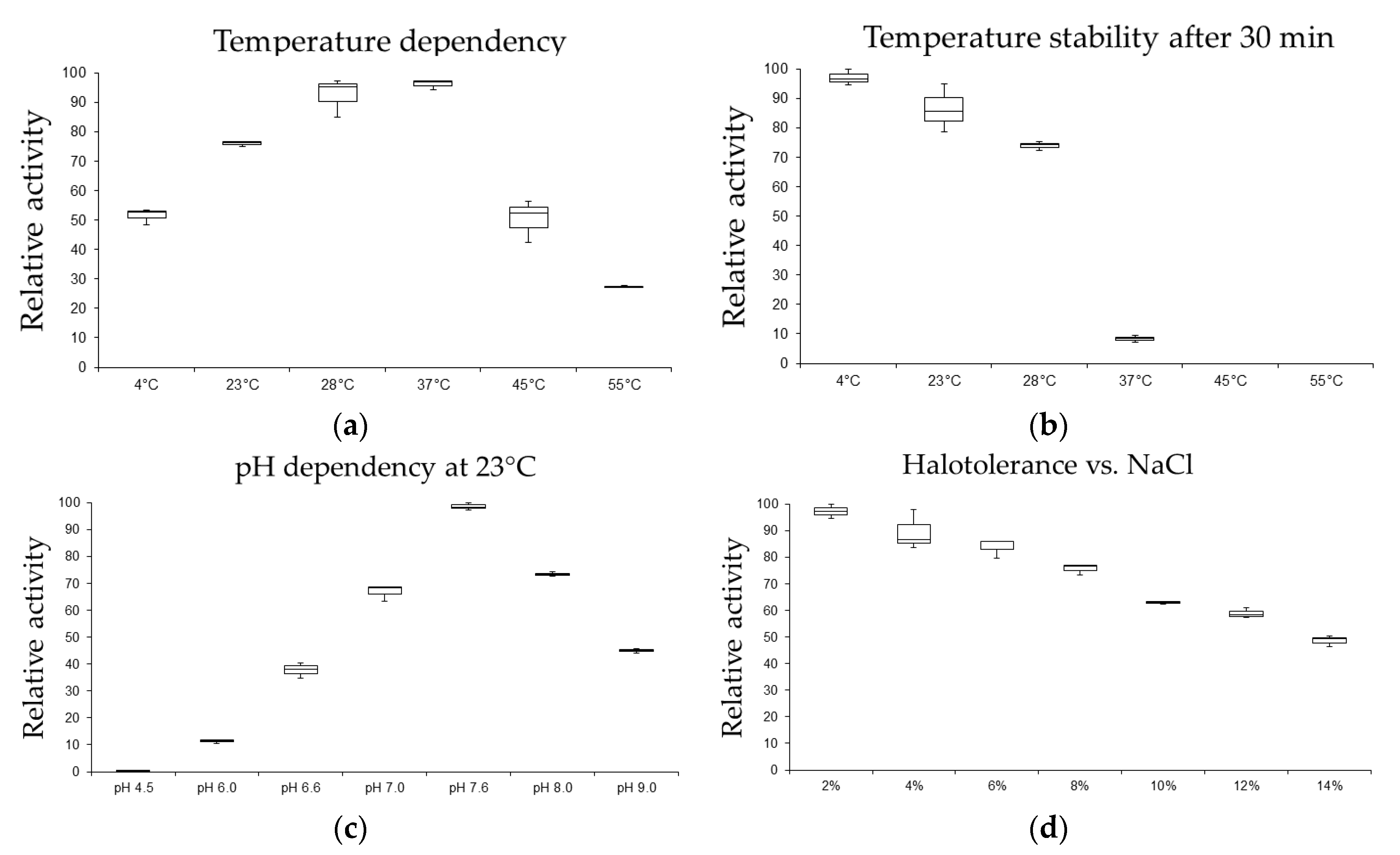
2.1. Enzymatic Activity Profile
2.2. Genome Sequencing and Assembly
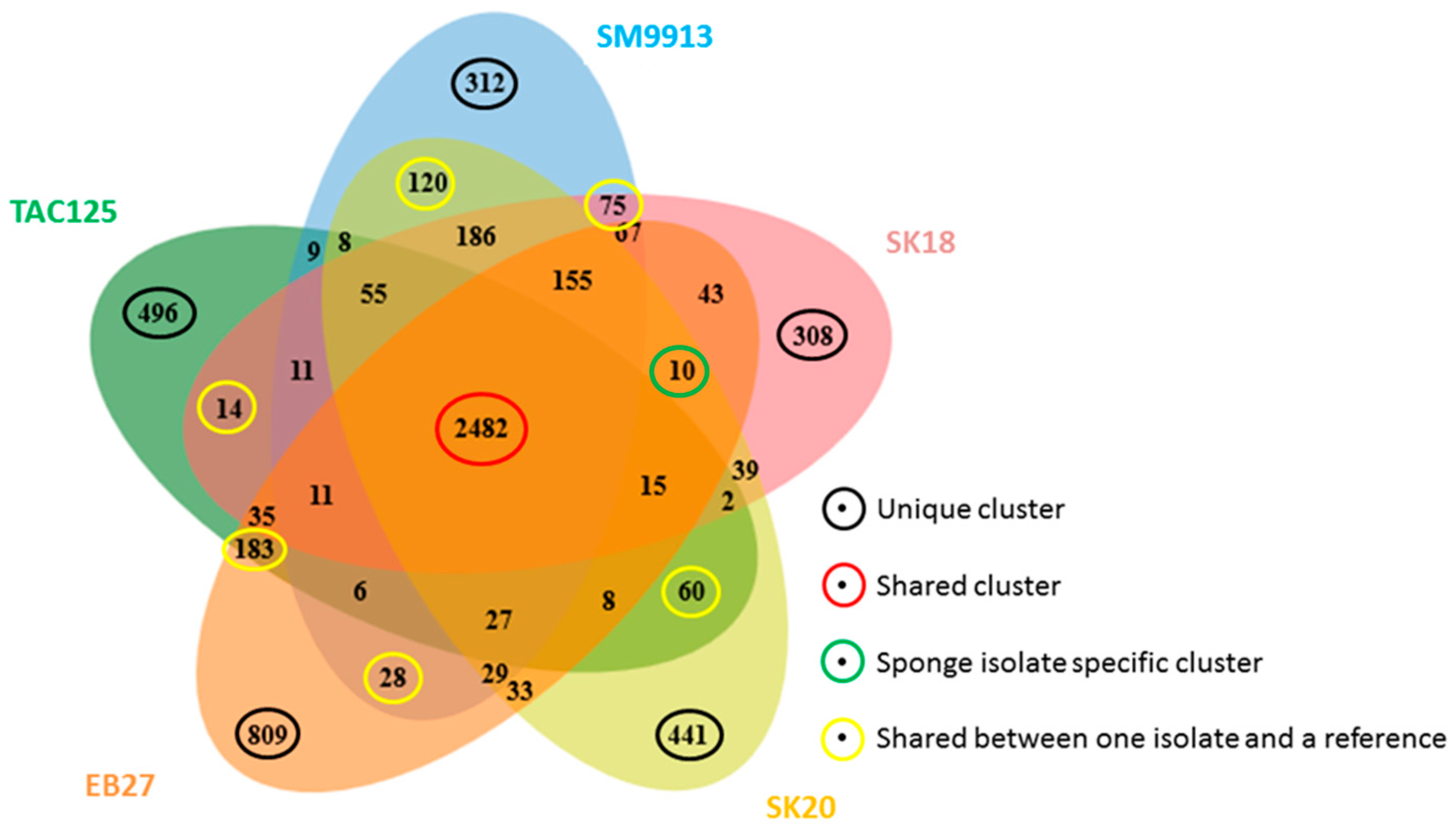
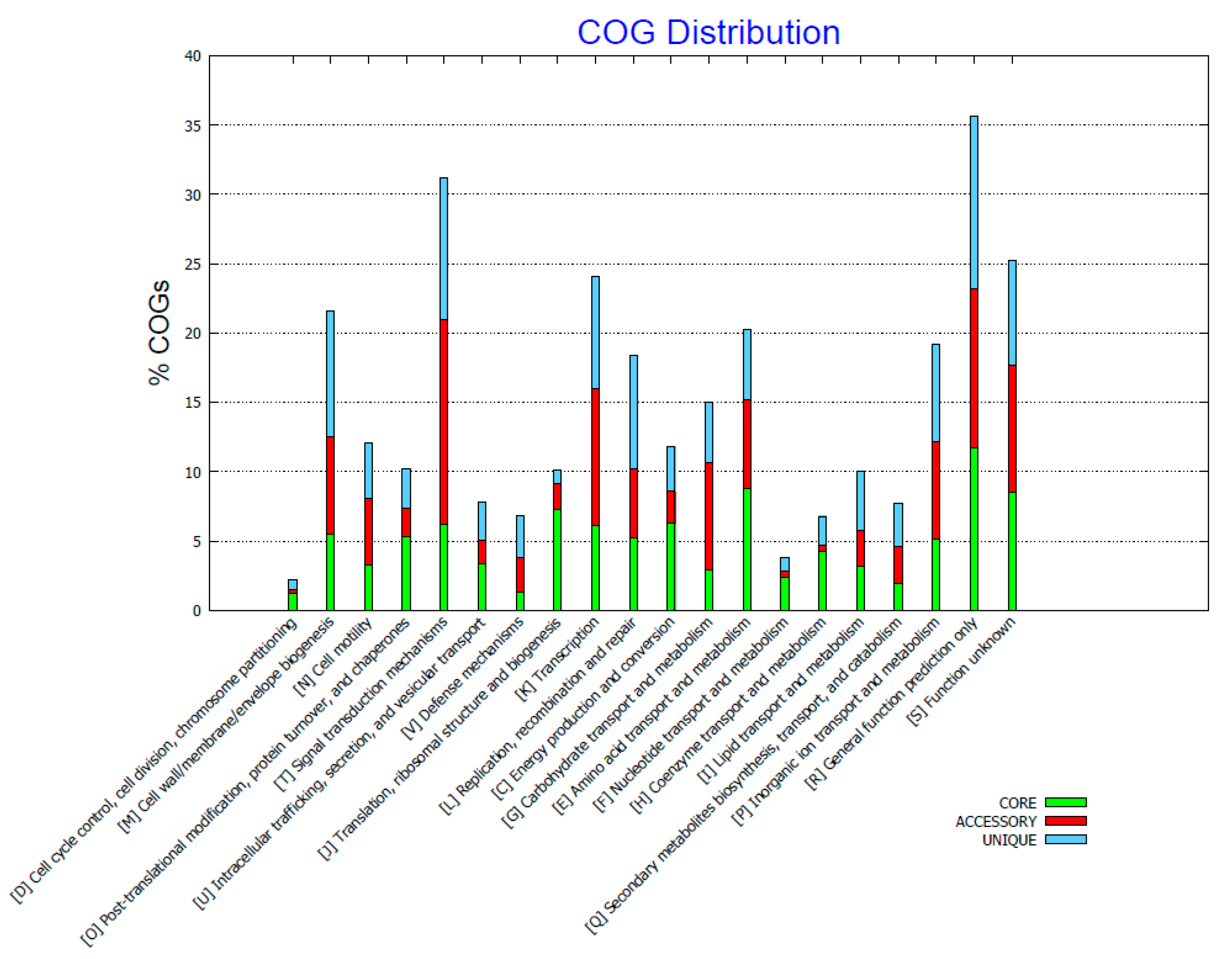
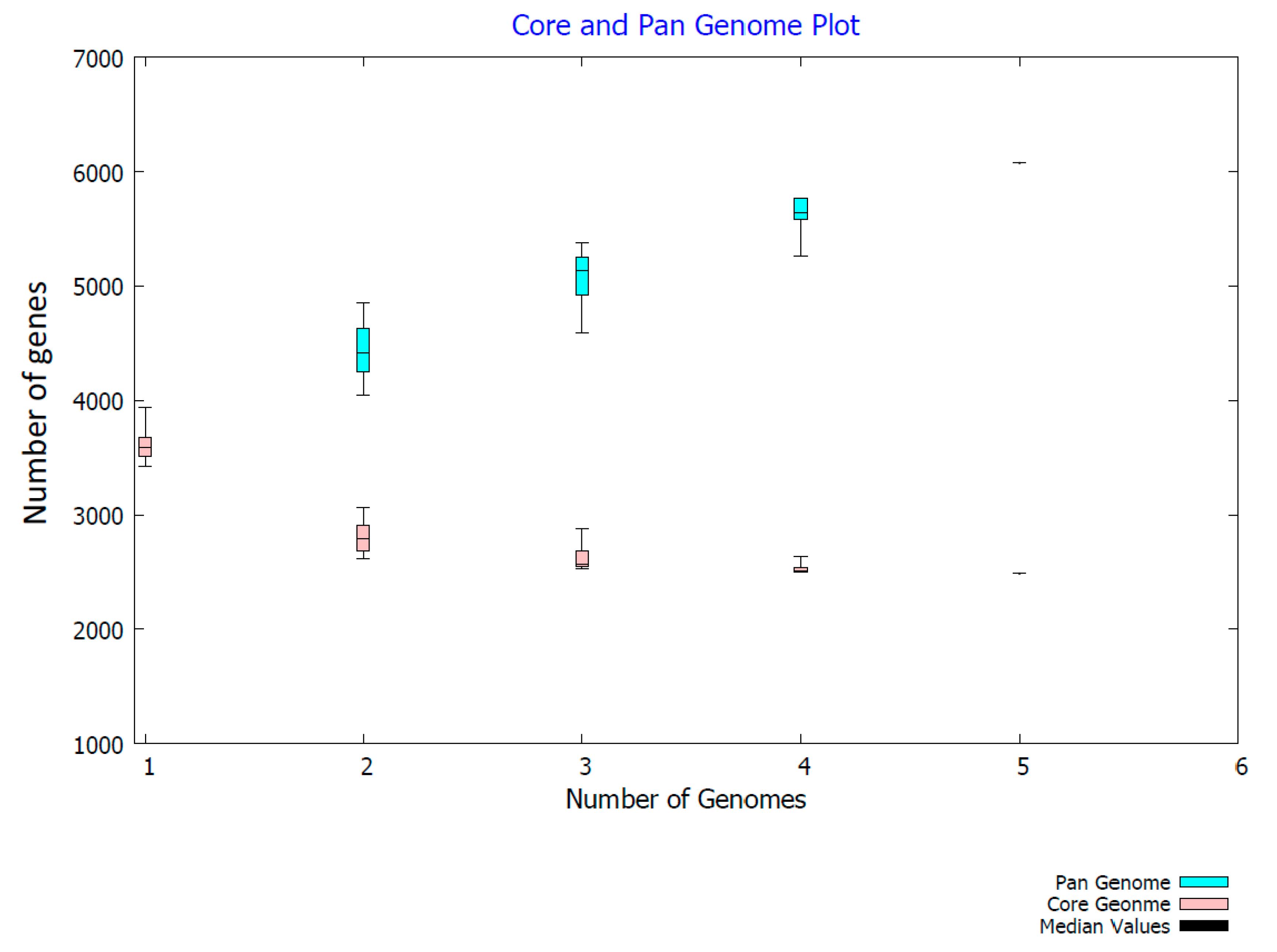
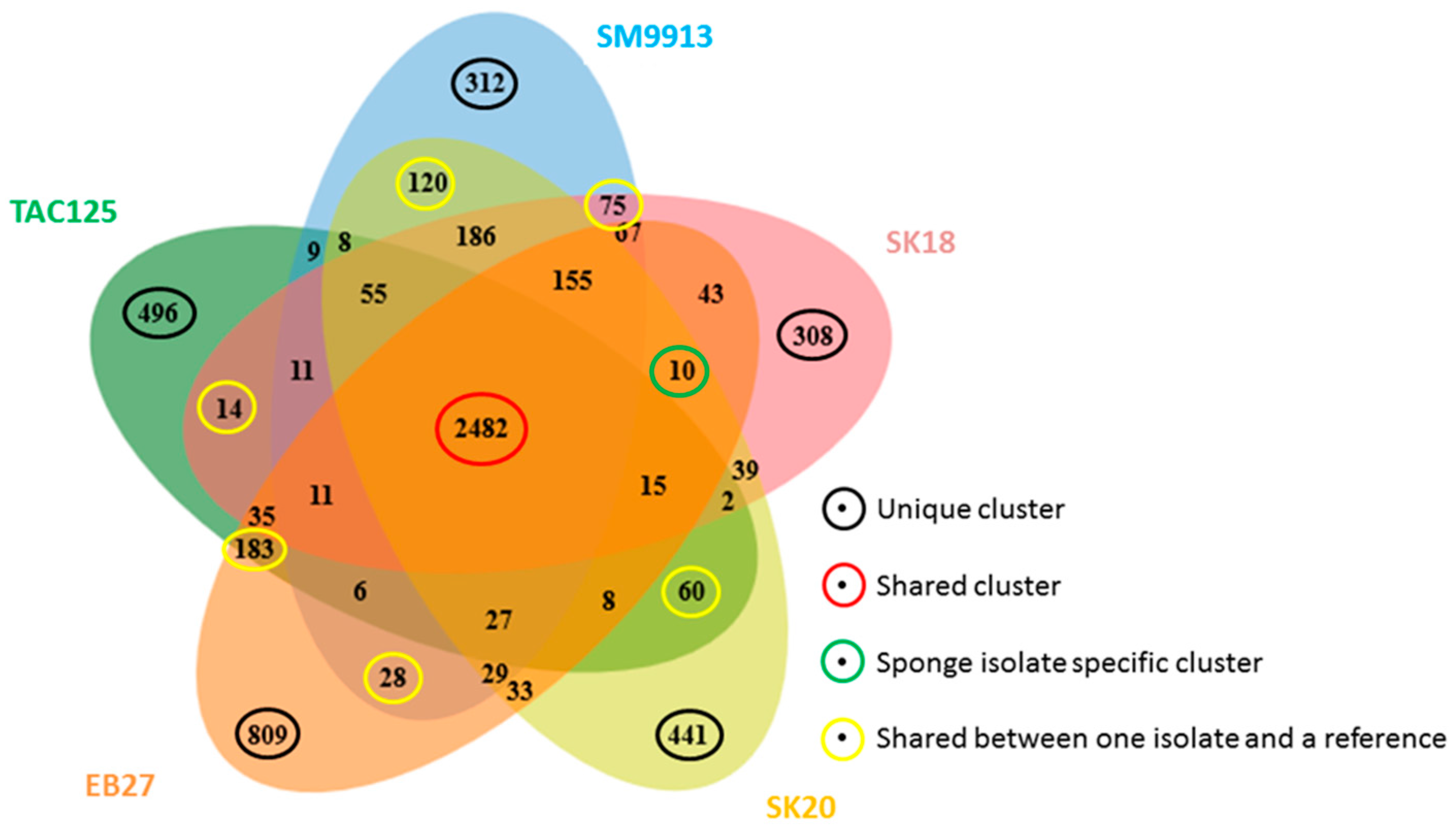
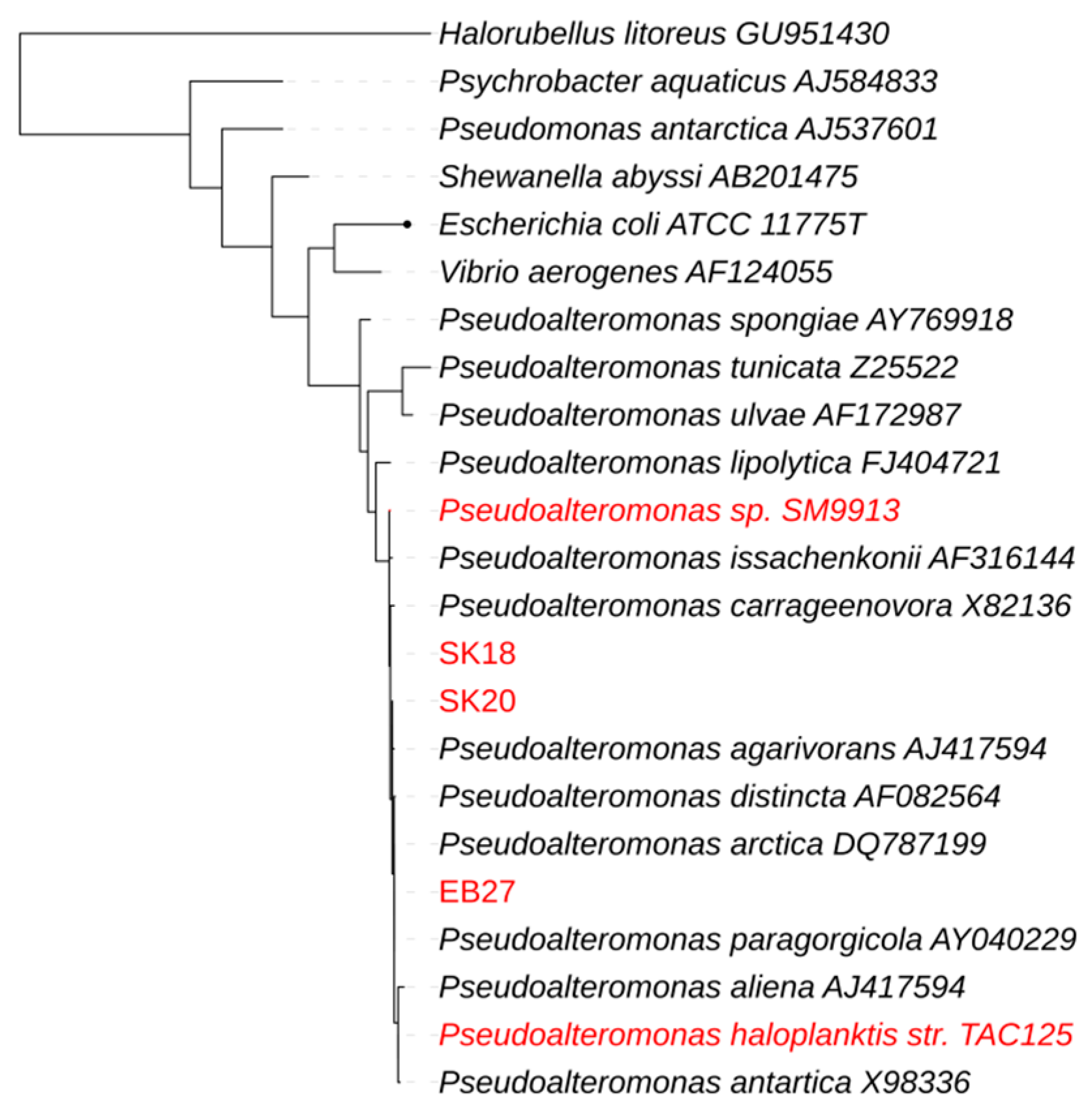
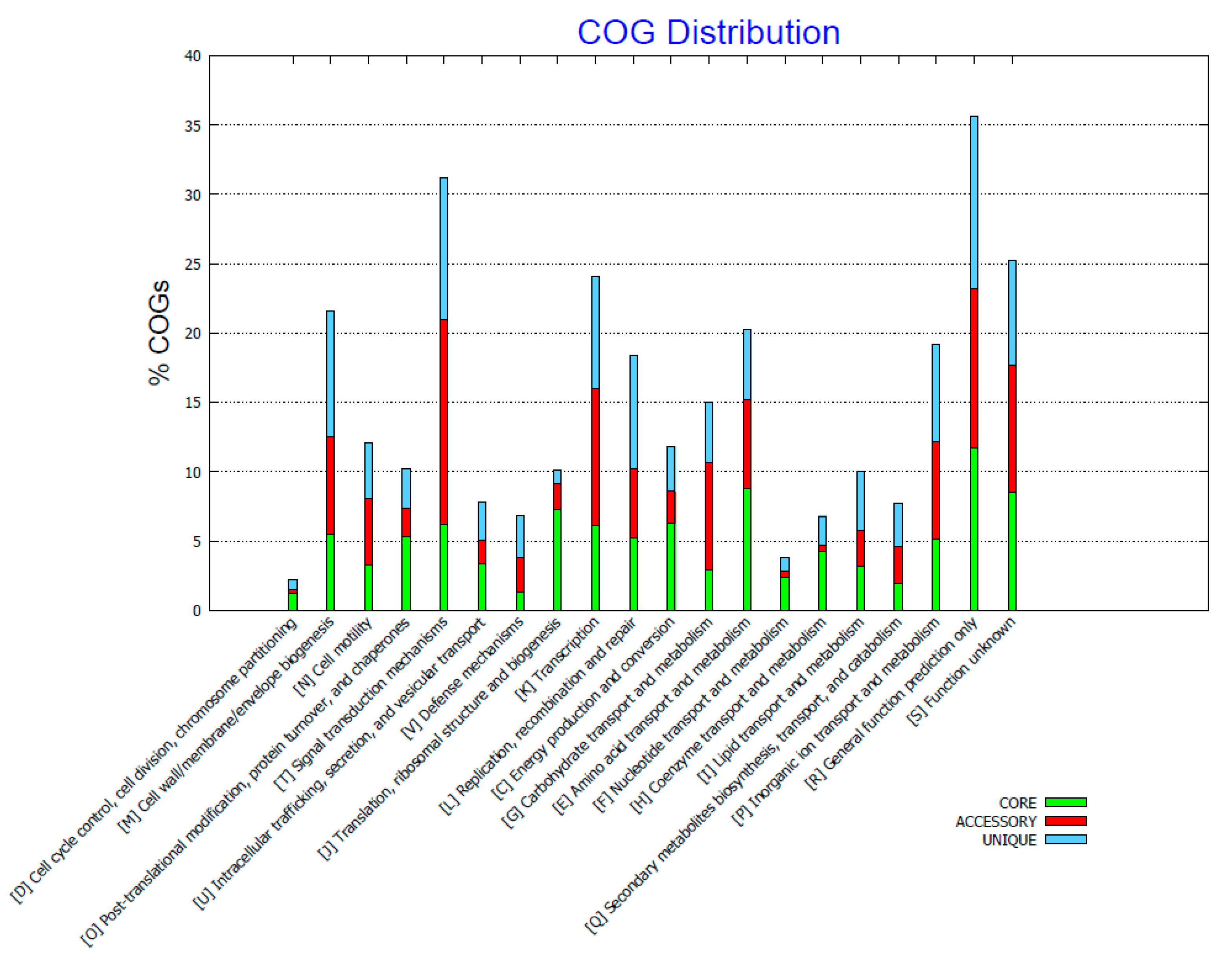
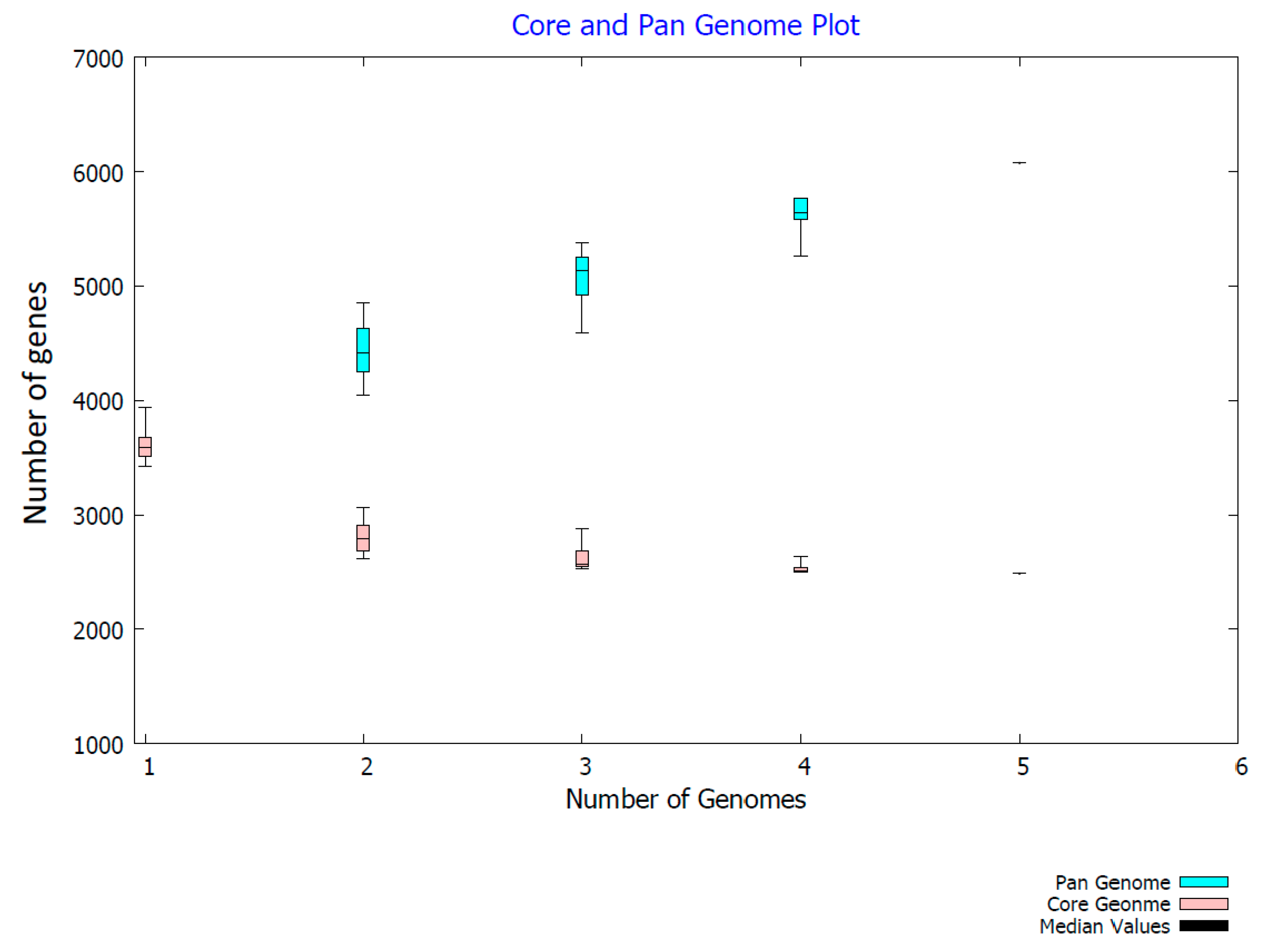
2.3. Genome Comparison
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sponge Collection and Isolation of Microorganisms
5.2. Enzyme Activity Plate Screenings
5.3. Enzyme Assays and Growth Characterization
5.4. Genomic DNA Isolation and Sequencing
5.5. Cloning, Expression, and Purification of the β-Glucosidase
5.6. Genome Analysis and Comparison
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gauthier, G.; Gauthier, M.; Christen, R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. Int. J. Syst. Bacteriol. 1995, 45, 755–761. [Google Scholar] [PubMed]
- Egan, S.; James, S.; Holmström, C.; Kjelleberg, S. Correlation between pigmentation and antifouling compounds produced by Pseudoalteromonas tunicata. Environ. Microbiol. 2002, 4, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar. Drugs 2007, 5, 220–241. [Google Scholar] [CrossRef] [PubMed]
- Fehér, D.; Barlow, R.; McAtee, J.; Hemscheidt, T.K. Highly brominated antimicrobial metabolites from a marine Pseudoalteromonas sp. J. Nat. Prod. 2010, 73, 1963–1966. [Google Scholar] [CrossRef] [PubMed]
- Holmström, C.; James, S.; Egan, S.; Kjelleberg, S. Inhibition of common fouling organisms by marine bacterial isolates ith special reference to the role of pigmented bacteria. Biofouling 1996, 10, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Cieśliński, H.; Kur, J.; Białkowska, A.; Baran, I.; Makowski, K.; Turkiewicz, M. Cloning, expression, and purification of a recombinant cold-adapted beta-galactosidase from antarctic bacterium Pseudoalteromonas sp. 22b. Protein Expr. Purif. 2005, 39, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Xiong, H.; Xu, Y.; Levin, L.A.; Qian, P.Y. Novel antifoulants: Inhibition of larval attachment by proteases. Mar. Biotechnol. N. Y. 2007, 9, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Kim, J.H.; Cho, K.W. A novel extracellular phospholipase C purified from a marine bacterium, Pseudoalteromonas sp. J937. Biotechnol. Lett. 2009, 31, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Nikapitiya, C.; Lee, Y.; Whang, I.; Kim, S.J.; Kang, D.H.; Lee, J. Cloning, purification and biochemical characterization of beta agarase from the marine bacterium Pseudoalteromonas sp. AG4. J. Ind Microbiol. Biotechnol. 2010, 37, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.Q.; Chen, X.L.; Hou, X.Y.; He, H.; Zhou, B.C.; Zhang, Y.Z. Molecular analysis of the gene encoding a cold-adapted halophilic subtilase from deep-sea psychrotolerant bacterium Pseudoalteromonas sp. SM9913: Cloning, expression, characterization and function analysis of the C-terminal PPC domains. Extremophiles 2009, 13, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Holmström, C.; James, S.; Neilan, B.A.; White, D.C.; Kjelleberg, S. Pseudoalteromonas tunicata sp. nov., a bacterium that produces antifouling agents. Int. J. Syst. Bacteriol. 1998, 48 Pt 4, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.; Holmström, C.; Kjelleberg, S. Pseudoalteromonas ulvae sp. nov., a bacterium with antifouling activities isolated from the surface of a marine alga. Int. J. Syst. Evol. Microbiol. 2001, 51, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Shevchenko, L.S.; Sawabe, T.; Lysenko, A.M.; Svetashev, V.I.; Gorshkova, N.M.; Satomi, M.; Christen, R.; Mikhailov, V.V. Pseudoalteromonas maricaloris sp. nov., isolated from an australian sponge, and reclassification of [Pseudoalteromonas aurantia] NCIMB 2033 as Pseudoalteromonas flavipulchra sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Kiprianova, E.A.; Mikhailov, V.V.; Levanova, G.F.; Garagulya, A.D.; Gorshkova, N.M.; Vysotskii, M.V.; Nicolau, D.V.; Yumoto, N.; Taguchi, T.; et al. Phenotypic diversity of Pseudoalteromonas citrea from different marine habitats and emendation of the description. Int. J. Syst. Bacteriol. 1998, 48 Pt 1, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Simidu, U.; Kita-Tsukamoto, K.; Yasumoto, T.; Yotsu, M. Taxonomy of four marine bacterial strains that produce tetrodotoxin. Int. J. Syst. Bacteriol. 1990, 40, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Takadera, T.; Adachi, K.; Nishijima, M.; Sano, H. Korormicin, a novel antibiotic specifically active against marine gram-negative bacteria, produced by a marine bacterium. J. Antibiot. Tokyo 1997, 50, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Akagawa-Matsushita, M.; Matsuo, M.; Koga, Y.; Yamasato, K. Alteromonas atlantica sp. nov and Alteromonas carrageenovora sp. nov., bacteria that decompose algal polysaccharides. Int. J. Syst. Evol. Microbiol. 1992, 42, 621–627. [Google Scholar] [CrossRef]
- Bozal, N.; Tudela, E.; Rosselló-Mora, R.; Lalucat, J.; Guinea, J. Pseudoalteromonas antarctica sp. nov., isolated from an antarctic coastal environment. Int. J. Syst. Bacteriol. 1997, 47, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P. Pseudoalteromonas prydzensis sp. nov., a psychrotrophic, halotolerant bacterium form antarctic sea ice. Int. J. Syst. Bacteriol. 1998, 48 Pt 3, 1037–1041. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.L.; Li, Y.; Zhang, Y.J.; Zhou, Z.M.; Zhang, W.X.; Chen, X.L.; Zhang, X.Y.; Zhou, B.C.; Wang, L.; Zhang, Y.Z. Comparative genomics reveals a deep-sea sediment-adapted life style of Pseudoalteromonas sp. SM9913. ISME J. 2011, 5, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.; Levin, L.; Arbizu, P.; Menot, L.; Buhl-Mortensen, P. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef]
- Sipkema, D. Marine biotechnology: Diving deeper for drugs. Microb. Biotechnol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Borchert, E.; Jackson, S.A.; O’Gara, F.; Dobson, A.D. Diversity of natural product biosynthetic genes in the microbiome of the deep sea sponges Inflatella pellicula, Poecillastra compressa, and Stelletta normani. Front. Microbiol. 2016, 7, 1027. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.A.; Flemer, B.; McCann, A.; Kennedy, J.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D. Archaea appear to dominate the microbiome of Inflatella pellicula deep sea sponges. PLoS ONE 2013, 8, e84438. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Flemer, B.; Jackson, S.A.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D. Evidence of a putative deep sea specific microbiome in marine sponges. PLoS ONE 2014, 9, e91092. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Pang, K.L.; Luo, Z.H. High fungal diversity and abundance recovered in the deep-sea sediments of the Pacific Ocean. Microb. Ecol. 2014, 68, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Li, X.M.; Li, X.; Xu, G.M.; Liu, Y.; Wang, B.G. Aspewentins D-H, 20-Nor-isopimarane derivatives from the deep sea sediment-derived fungus Aspergillus wentii SD-310. J. Nat. Prod. 2016, 79, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Schmaljohann, R.; Imhoff, J.F. Biscogniauxone, a new isopyrrolonaphthoquinone compound from the fungus Biscogniauxia mediterranea isolated from deep-sea sediments. Mar. Drugs 2016, 14, 204. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Ran, L.Y.; Liu, C.; Chen, X.L.; Zhang, X.Y.; Qin, Q.L.; Zhou, B.C.; Zhang, Y.Z. Culture condition optimization and pilot scale production of the M12 metalloprotease Myroilysin produced by the deep-sea bacterium myroides profundi D25. Molecules 2015, 20, 11891–11901. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, Y.; Tang, B.L.; Zhong, S.; Shi, M.; Xie, B.B.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Chen, X.L. Pilot-scale production and thermostability improvement of the M23 protease pseudoalterin from the deep sea bacterium Pseudoalteromonas sp. CF6-2. Molecules 2016, 21, 1567. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Siddiqui, K.S.; Andrews, D.; Sowers, K.R. Low-temperature extremophiles and their applications. Curr. Opin. Biotechnol. 2002, 13, 253–261. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, molecular mechanisms, and industrial applications of cold-active enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef] [PubMed]
- Georlette, D.; Jónsson, Z.O.; Van Petegem, F.; Chessa, J.; Van Beeumen, J.; Hübscher, U.; Gerday, C. A DNA ligase from the psychrophile Pseudoalteromonas haloplanktis gives insights into the adaptation of proteins to low temperatures. Eur J. Biochem. 2000, 267, 3502–3512. [Google Scholar] [CrossRef] [PubMed]
- Albino, A.; De Angelis, A.; Marco, S.; Severino, V.; Chambery, A.; Di Maro, A.; Desiderio, D.; Raimo, G.; Masullo, M.; De Vendittis, E. The cold-adapted γ-glutamyl-cysteine ligase from the psychrophile Pseudoalteromonas haloplanktis. Biochimie 2014, 104, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Truong, L.V.; Tuyen, H.; Helmke, E.; Binh, L.T.; Schweder, T. Cloning of two pectate lyase genes from the Marine antarctic bacterium Pseudoalteromonas haloplanktis strain ANT/505 and characterization of the enzymes. Extremophiles 2001, 5, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Cotugno, R.; Rosaria Ruocco, M.; Marco, S.; Falasca, P.; Evangelista, G.; Raimo, G.; Chambery, A.; Di Maro, A.; Masullo, M.; De Vendittis, E. Differential cold-adaptation among protein components of the thioredoxin system in the psychrophilic eubacterium Pseudoalteromonas haloplanktis TAC 125. Mol. Biosyst. 2009, 5, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Falasca, P.; Evangelista, G.; Cotugno, R.; Marco, S.; Masullo, M.; De Vendittis, E.; Raimo, G. Properties of the endogenous components of the thioredoxin system in the psychrophilic eubacterium Pseudoalteromonas haloplanktis TAC 125. Extremophiles 2012, 16, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Di Maro, A.; Ruocco, M.R.; Chambery, A.; Parente, A.; Di Martino, M.T.; Parlato, G.; Masullo, M.; De Vendittis, E. Psychrophilic superoxide dismutase from Pseudoalteromonas haloplanktis: Biochemical characterization and identification of a highly reactive cysteine residue. Biochimie 2006, 88, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Ruocco, M.R.; Cecere, F.; Di Maro, A.; Chambery, A.; Michniewicz, A.; Parlato, G.; Masullo, M.; De Vendittis, E. Glutathionylation of the iron superoxide dismutase from the psychrophilic eubacterium Pseudoalteromonas haloplanktis. Biochem. Biophys. Acta 2008, 1784, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Pulicherla, K.K.; Sambasiva Rao, K.R.S. Marine biocatalysts and their stability: Molecular approach. In Marine Enzymes for Biocatalysis—Sources, Biocatalytic Characteristics and Bioprocesses of Marine Enzymes; Trincone, A., Ed.; Woodhead Publishing Series in Biomedicine: Cambridge, UK, 2013; pp. 109–130. [Google Scholar]
- Xie, B.B.; Shu, Y.L.; Qin, Q.L.; Rong, J.C.; Zhang, X.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Genome sequence of the cycloprodigiosin-producing bacterial strain Pseudoalteromonas rubra ATCC 29570(T). J. Bacteriol. 2012, 194, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Bosi, E.; Fondi, M.; Orlandini, V.; Perrin, E.; Maida, I.; de Pascale, D.; Tutino, M.L.; Parrilli, E.; Lo Giudice, A.; Filloux, A.; et al. The pan-genome of (Antarctic) Pseudoalteromonas bacteria: Evolutionary and functional insights. BMC Genom. 2017, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Isnansetyo, A.; Kamei, Y. Mc21-A, a bactericidal antibiotic produced by a new marine bacterium, Pseudoalteromonas phenolica sp. nov. O-BC30(T), against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Sannino, F.; Parrilli, E.; Apuzzo, G.A.; de Pascale, D.; Tedesco, P.; Maida, I.; Perrin, E.; Fondi, M.; Fani, R.; Marino, G.; et al. Pseudoalteromonas haloplanktis produces methylamine, a volatile compound active against Burkholderia cepacia complex strains. New Biotechnol. 2017, 35, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Médigue, C.; Krin, E.; Pascal, G.; Barbe, V.; Bernsel, A.; Bertin, P.N.; Cheung, F.; Cruveiller, S.; D’Amico, S.; Duilio, A.; et al. Coping with cold: The genome of the versatile marine Antarctica bacterium Pseudoalteromonas haloplanktis TAC125. Genome Res. 2005, 15, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Nordberg, H.; Shabalov, I.; Aerts, A.; Cantor, M.; Goodstein, D.; Kuo, A.; Minovitsky, S.; Nikitin, R.; Ohm, R.A.; et al. The genome portal of the department of energy joint genome institute. Nucleic Acids Res. 2012, 40, D26–D32. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the department of energy joint genome institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. Bpga—An ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. Rnammer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Reynolds, D.; Liu, M.; Stark, M.; Kjelleberg, S.; Webster, N.S.; Thomas, T. Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc. Natl. Acad. Sci. USA 2012, 109, E1878–E1887. [Google Scholar] [CrossRef] [PubMed]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (itol): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef] [PubMed]
- Medini, D.; Donati, C.; Tettelin, H.; Masignani, V.; Rappuoli, R. The microbial pan-genome. Curr. Opin. Genet. Dev. 2005, 15, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Medema, M.H.; Kazempour, D.; Fischbach, M.A.; Breitling, R.; Takano, E.; Weber, T. Antismash 2.0—A versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013, 41, W204–W212. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. Antismash: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. Antismash 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Thomas, T. Evolution and function of eukaryotic-like proteins from sponge symbionts. Mol. Ecol. 2016, 25, 5242–5253. [Google Scholar] [CrossRef] [PubMed]
- Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.; Dandekar, T.; Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 2011, 5, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Kato, J.; Nakashima, K.; Kuroda, A.; Ikeda, T.; Takiguchi, N.; Ohtake, H. Cloning and characterization of extracellular metal protease gene of the algicidal marine bacterium Pseudoalteromonas sp. Strain A28. Biosci. Biotechnol. Biochem. 2002, 66, 1366–1369. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.H.; Wright, M.E.; Silverman, M. Variable expression of extracellular polysaccharide in the marine bacterium Pseudomonas atlantica is controlled by genome rearrangement. Proc. Natl. Acad. Sci. USA 1988, 85, 3923–3927. [Google Scholar] [CrossRef] [PubMed]
- Longeon, A.; Peduzzi, J.; Barthélemy, M.; Corre, S.; Nicolas, J.L.; Guyot, M. Purification and partial identification of novel antimicrobial protein from marine bacterium Pseudoalteromonas species strain X153. Mar. Biotechnol. N. Y. 2004, 6, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Enomoto, K. Characterization of a gene cluster and its putative promoter region for violacein biosynthesis in Pseudoalteromonas sp. 520P1. Appl. Microbiol. Biotechnol. 2011, 90, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli physiology in luria-bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [PubMed]
- Papa, R.; Rippa, V.; Sannia, G.; Marino, G.; Duilio, A. An effective cold inducible expression system developed in Pseudoalteromonas haloplanktis tac125. J. Biotechnol. 2007, 127, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.; Lübeck, M.; Lübeck, P.S.; Ahring, B.K. Fungal beta-glucosidases: A bottleneck in industrial use of lignocellulosic materials. Biomolecules 2013, 3, 612–631. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Verma, A.K.; Kumar, V. Catalytic properties, functional attributes and industrial applications of β-glucosidases. 3 Biotech 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Yokouchi, H.; Suzuki, N.; Ohata, H.; Matsunaga, T. Saccharification of marine microalgae using marine bacteria for ethanol production. Appl. Biochem. Biotechnol. 2003, 105, 247–254. [Google Scholar] [CrossRef]
- El-Naggar, M.M.; Abdul-Raouf, U.M.; Ibrahim, H.A.; El-Sayed, W.M. Saccharification of ulva lactuca via Pseudoalteromonas piscicida for biofuel production. J. Energy Natl. Resour. 2014, 3, 77–84. [Google Scholar]
- Offret, C.; Desriac, F.; Le Chevalier, P.; Mounier, J.; Jégou, C.; Fleury, Y. Spotlight on antimicrobial metabolites from the marine bacteria Pseudoalteromonas: Chemodiversity and ecological significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef] [PubMed]
- Kamke, J.; Bayer, K.; Woyke, T.; Hentschel, U. Exploring symbioses by single-cell genomics. Biol. Bull. 2012, 223, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Shikuma, N.J.; Pilhofer, M.; Weiss, G.L.; Hadfield, M.G.; Jensen, G.J.; Newman, D.K. Marine tubeworm metamorphosis induced by arrays of bacterial phage tail-like structures. Science 2014, 343, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Shirazian, P.; Asad, S.; Amoozegar, M. The potential of halotolerant bacteria for the production of antineoplastic enzymes: l-asparaginase and l-glutaminase. Excli J. 2016, 15, 268–279. [Google Scholar] [PubMed]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Mohd Omar, S.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, Y.; Mishra, S.; Bisaria, V.S. Microbial β-glucosidases: Cloning, properties and applications. Crit. Rev. Biotechnol. 2002, 22, 375–407. [Google Scholar]
- Market Reports Hub. Available online: http://www.marketreportshub.com/proteases-industry-research-marketsandmarkets.html (accessed on 8 June 2017).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Spades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. Checkm: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The rast server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. Rasttk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The seed and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | Sponge | Depth (m) | Cellulase | Lipase | Protease | β-Glc/Gal |
|---|---|---|---|---|---|---|
| EB27 | Poecillastra compressa | 1480 | XX | X | XXX | XXX (Glc) |
| SK18 | Sericolophus hawaiicus | 2129 | - | X | XXX | - |
| SK20 | Inflatella pellicula | 2900 | - | X | - | XXX (Gal) |
| ID | 4 °C mu; tgen (min) | 23 °C mu; tgen (min) | 28 °C mu; tgen (min) | 37 °C mu; tgen (min) |
|---|---|---|---|---|
| EB27 | 0.54 ± 0.15; 88.3 ± 20.01 | 1.03 ± 0.05; 40.6 ± 2.1 | 0.98 ± 0.04; 42.46 ± 1.84 | 0.82 ± 0.09; 51.39 ± 6.75 |
| SK18 | 0.28 ± 0.06; 159.36 ± 12.75 | 1.76 ± 0.095; 23.8 ± 1.24 | 2.08 ± 0.013; 20 ± 0.13 | 1.46 ± 0.16; 29.04 ± 2.9 |
| SK20 | 0.29 ± 0.02; 144.92 ± 31.2 | 0.99 ± 0.08; 42.36 ± 3.56 | 1.41 ± 0.01; 29.5 ± 0.24 | 0.9 ± 0.14; 48.66 ± 7.29 |
| ID | Genome Size (Mb) | GC Content | N50 (kb) | Contigs | CDS | No. of RNAs | Coverage | Genome Completness (%) |
|---|---|---|---|---|---|---|---|---|
| TAC125 | 3.85 | 40.1% | n/a | n/a | 3473 | 134 | n/a | 100 |
| SM9913 | 4.04 | 40.3% | n/a | n/a | 3699 | 87 | n/a | 100 |
| EB27 | 4.56 | 39.1% | 216.9 | 114 | 4012 | 136 | 196× | 99.72 |
| SK18 | 3.98 | 40.2% | 156.5 | 115 | 3582 | 110 | 213× | 99.75 |
| SK20 | 4.15 | 40.3% | 98.5 | 213 | 3811 | 139 | 230× | 99.66 |
| ID | Lipase/Est. | β-Galactosidase | Protease | β-Glucosidase | Cellulase |
|---|---|---|---|---|---|
| TAC125 | 49 | 0 | 35 | 0 | 2 |
| SM9913 | 67 | 0 | 42 | 2 | 3 |
| EB27 | 69 | 1 | 48 | 4 | 5 |
| SK18 | 63 | 0 | 39 | 2 | 3 |
| SK20 | 56 | 1 | 40 | 2 | 3 |
| ID | Bacteriocin | Arylpolyene | Siderophore |
|---|---|---|---|
| TAC125 | 1 | 1 | - |
| SM9913 | 1 | - | 1 |
| EB27 | 2 | 1 | - |
| SK18 | 1 | 1 | 1 |
| SK20 | 1 | - | - |
| ID | Ankyrin Repeats | Tetratricopeptide Repeats | Nitrite Reductase | Proteases | Sulfatases | Peptidases |
|---|---|---|---|---|---|---|
| TAC125 | 2 | 2 | 1 | 35 | 0 | 58 |
| SM9913 | 1 | 2 | 0 | 42 | 1 | 63 |
| EB27 | 2 | 2 | 3 | 48 | 0 | 65 |
| SK18 | 1 | 2 | 0 | 39 | 1 | 63 |
| SK20 | 1 | 2 | 0 | 40 | 1 | 58 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borchert, E.; Knobloch, S.; Dwyer, E.; Flynn, S.; Jackson, S.A.; Jóhannsson, R.; Marteinsson, V.T.; O’Gara, F.; Dobson, A.D.W. Biotechnological Potential of Cold Adapted Pseudoalteromonas spp. Isolated from ‘Deep Sea’ Sponges. Mar. Drugs 2017, 15, 184. https://doi.org/10.3390/md15060184
Borchert E, Knobloch S, Dwyer E, Flynn S, Jackson SA, Jóhannsson R, Marteinsson VT, O’Gara F, Dobson ADW. Biotechnological Potential of Cold Adapted Pseudoalteromonas spp. Isolated from ‘Deep Sea’ Sponges. Marine Drugs. 2017; 15(6):184. https://doi.org/10.3390/md15060184
Chicago/Turabian StyleBorchert, Erik, Stephen Knobloch, Emilie Dwyer, Sinéad Flynn, Stephen A. Jackson, Ragnar Jóhannsson, Viggó T. Marteinsson, Fergal O’Gara, and Alan D. W. Dobson. 2017. "Biotechnological Potential of Cold Adapted Pseudoalteromonas spp. Isolated from ‘Deep Sea’ Sponges" Marine Drugs 15, no. 6: 184. https://doi.org/10.3390/md15060184
APA StyleBorchert, E., Knobloch, S., Dwyer, E., Flynn, S., Jackson, S. A., Jóhannsson, R., Marteinsson, V. T., O’Gara, F., & Dobson, A. D. W. (2017). Biotechnological Potential of Cold Adapted Pseudoalteromonas spp. Isolated from ‘Deep Sea’ Sponges. Marine Drugs, 15(6), 184. https://doi.org/10.3390/md15060184