
The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions


Abstract
:1. Introduction
1.1. Antimicrobial Use and Antimicrobial Resistance in Aquaculture
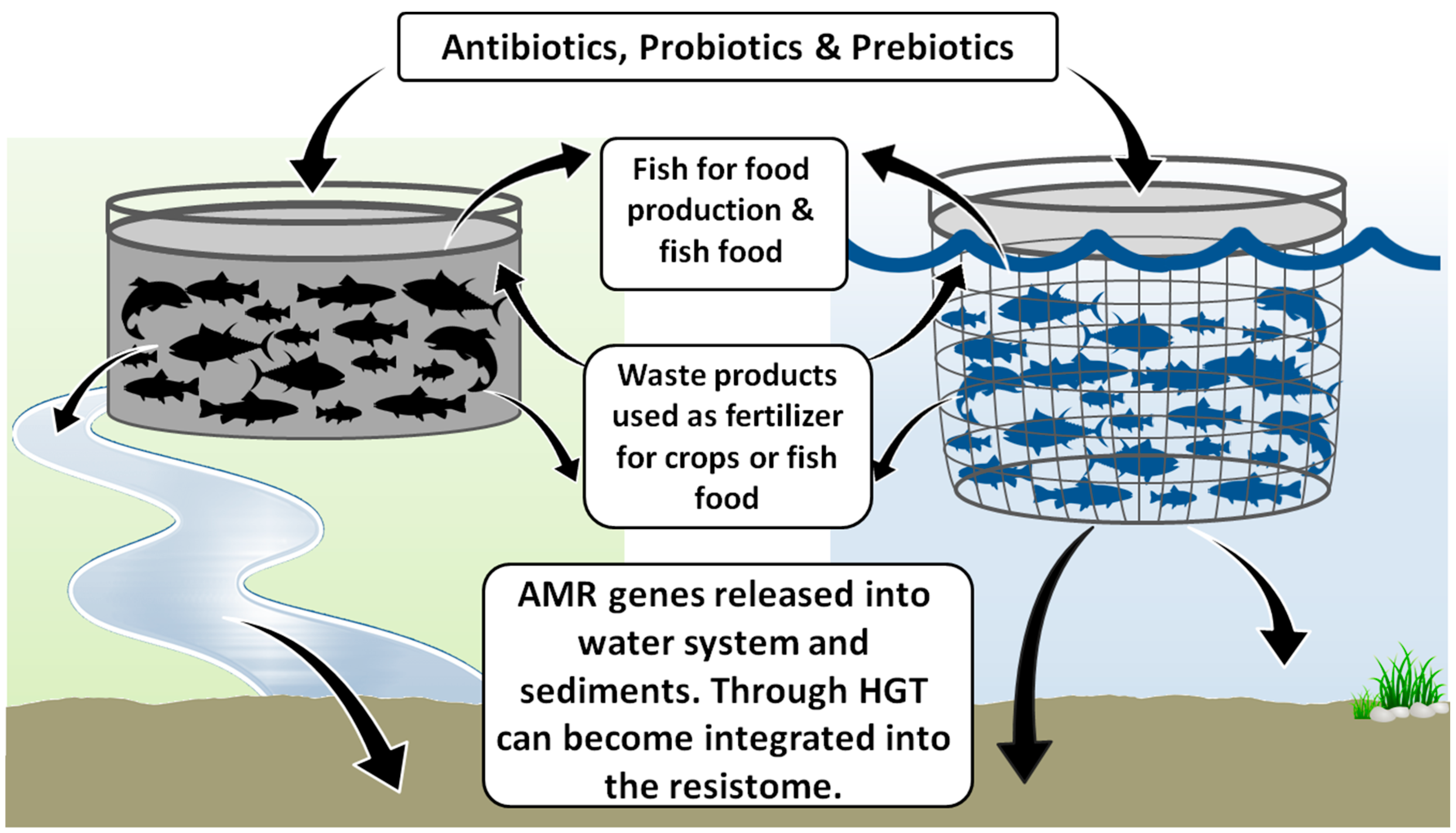
1.2. Increased Transfer of AMR Directly to the Environment through Open Systems
1.3. AMR in Closed Aquaculture Systems
1.4. Integrated Fish Farming and “Waste as Feed”
1.5. Probiotic Application
2. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2016. [Google Scholar]
- Pauly, D.; Zeller, D. Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 2016, 7, 10244. [Google Scholar] [CrossRef] [PubMed]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Chan, K.G.; Lee, L.H.; Goh, B.H. Streptomyces bacteria as potential probiotics in aquaculture. Front. Microbiol. 2016, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S. Recent advances in the role of probiotics and prebiotics in carp aquaculture: A review. Aquaculture 2016, 454, 243–251. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, X.; Yang, Y.; Han, D.; Jin, J.; Xie, S. Effect of dietary chitosan on growth performance, haematology, immune response, intestine morphology, intestine microbiota and disease resistance in gibel carp (Carassius auratus gibelio). Aquac. Nutr. 2014, 20, 532–546. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Ramos, F. Analytical strategies for the detection and quantification of antibiotic residues in aquaculture fishes: A review. Trends Food Sci. Technol. 2016, 52, 16–30. [Google Scholar] [CrossRef]
- Smith, P. Antimicrobial resistance in aquaculture. Rev. Sci. Tech. Int. Epiz. 2008, 27, 243–264. [Google Scholar] [CrossRef]
- European Council, 2001a. Directive 2001/82/EC of the European Parliament and of the Council of 6th November 2001 on the Community code relating to veterinary medicinal products. Off. J. Eur. Community L-311 2004, 1–66. [Google Scholar]
- European Council, 2001b. EC Enterprise Directorate General. Notice to Applicants: Vol 6B Presentation and Content of the Dossier Part II G and H: Guidance on Data Relating to the Environmental Risk Assessment for Products Containing or Consisting of Genetically Modified Organisms; European Council: Brussels, Belgium, 2001. [Google Scholar]
- European Council, 2001c. EC Health and Consumer Protection Directorate General. Guidelines on the Assessment of Additives in Animal Nutrition: Additives other than Micro-Organisms and Enzymes; European Council: Brussels, Belgium, 2001. [Google Scholar]
- O’Neill, J. Antimicrobials in Agriculture and the Environment: Reducing Unnecessary Use and Waste. The Review on Antimicrobial Resistance. Available online: http://amr-review.org/sites/default/files/Antimicrobials%20in%20agriculture%20and%20the%20environment%20-%20Reducing%20unnecessary%20use%20and%20waste.pdf (accessed on 25 May 2017).
- Norwegian Ministries. Norwegian Government’s National Strategy against Antimicrobial Resistance 2015–2020; Norwegian Ministry of Health and Care Services; Publication number: I-1164. Available online: https://www.regjeringen.no/contentassets/5eaf66ac392143b3b2054aed90b85210/antibiotic-resistance-engelsk-lavopploslig-versjon-for-nett-10-09-15.pdf (accessed on 25 May 2017).
- Chuah, L.; Effarizah, M.E.; Goni, A.M.; Rusul, G. Antibiotic application and emergence of multiple antibiotic resistance (MAR) in global catfish aquaculture. Curr. Environ. Health Rep. 2016, 3, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; Benjamín, P. Plan de Contingencia Sernapesca para la Industria del Salmón. Available online: http://repositorio.uchile.cl/handle/2250/132847 (accessed on 25 May 2017).
- Burridge, L.; Weis, J.S.; Cabello, F.; Pizarro, J.; Bostick, K. Chemical use in salmon aquaculture: A review of current practices and possible environmental effects. Aquaculture 2010, 306, 7–23. [Google Scholar] [CrossRef]
- Holmström, K.; Graslund, S.; Wahlstrom, A.; Poungshompoo, S.; Bengtsson, B.-E.; Kautsky, N. Antibiotic use in shrimp farming and implications for environmental impacts and human health. Int. J. Food Sci. Technol. 2003, 38, 255–266. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Tomova, A.; Lopez, A.; Maldonado, M.A.; Henriquez, L.A.; Ivanova, L.; Moy, F.; Godfrey, H.P.; Cabello, F.C. Salmon aquaculture and antimicrobial resistance in the marine environment. PLoS ONE 2012, 7, e42724. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dolz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial use in aquaculture re-examined: Its relevance to antimicrobial resistance and to animal and human health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.S.; Grant, J.; Catchpole, M. The Drugs Don’t Work: A Global Threat; Penguin: London, UK, 2013. [Google Scholar]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 73, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Kolter, R.; van Wezel, G.P. Goodbye to brute force in antibiotic discovery? Nat. Microbiol. 2016, 1, 15020. [Google Scholar] [CrossRef] [PubMed]
- Chamier, B.; Lorenz, M.G.; Wackernagel, W. Natural transformation of Acinetobacter calcoaceticus by plasmid DNA adsorbed on sand and groundwater aquifer material. Appl. Environ. Microbiol. 1993, 59, 1662–1667. [Google Scholar] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, W.; Takagi, T. Rapid pathway evolution facilitated by horizontal gene transfers across prokaryotic lineages. PLoS Genet. 2009, 5, e1000402. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R.I. Horizontal gene exchange in environmental microbiota. Front. Microbiol. 2011, 2, 158. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.G.; Verner-Jeffreys, D.W.; Baker-Austin, C. Aquatic systems: Maintaining, mixing and mobilising antimicrobial resistance? Trends Ecol. Evol. 2011, 26, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.C. Integron involvement in environmental spread of antibiotic resistance. Front. Microbiol. 2012, 3, 119. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef] [PubMed]
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Burgmann, H.; Sorum, H.; Norstrom, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Amos, G.C.; Gozzard, E.; Carter, C.E.; Mead, A.; Bowes, M.J.; Hawkey, P.M.; Zhang, L.; Singer, A.C.; Gaze, W.H.; Wellington, E.M. Validated predictive modelling of the environmental resistome. ISME J. 2015, 9, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Banquero, F.; Martínez, J.L.; Canton, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Muziasari, W.I.; Parnanen, K.; Johnson, T.A.; Lyra, C.; Karkman, A.; Stedtfeld, R.D.; Tamminen, M.; Tiedje, J.M.; Virta, M. Aquaculture changes the profile of antibiotic resistance and mobile genetic element associated genes in Baltic Sea sediments. FEMS Microbiol. Ecol. 2016, 92, flw052. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L. Recent advances on antibiotic resistance genes. In Recent Advances in Marine Biotechnology Vol 10: Molecular Genetics of Marine Organisms; Fingerman, M., Ed.; CRC Press: London, UK, 2003; pp. 13–32. [Google Scholar]
- Tamminen, M.; Karkman, A.; Lohmus, A.; Muziasari, W.I.; Takasu, H.; Wada, S.; Suzuki, S.; Virta, M. Tetracycline resistance genes persist at aquaculture farms in the absence of selection pressure. Environ. Sci. Technol. 2011, 45, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilback, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed]
- Done, H.Y.; Halden, R.U. Reconnaissance of 47 antibiotics and associated microbial risks in seafood sold in the United States. J. Hazard. Mater. 2015, 282, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, L.; Yu, X.; Hu, J.; Chen, Y.; He, G.; Jiang, Q. Antibiotic residues in meat, milk and aquatic products in Shanghai and human exposure assessment. Food Control 2017, 80, 217–225. [Google Scholar] [CrossRef]
- Schlenk, D.; Gollon, J.L.; Griffin, B.R. Efficacy of copper sulfate for the treatment of Ichthyophthiriasis in Channel catfish. J. Aquat. Anim. Health 1998, 10, 390–396. [Google Scholar] [CrossRef]
- Kundu, G.K.; Alauddin, M.; Akter, M.S.; Khan, M.S.; Islam, M.M.; Mondal, G.; Islam, D.; Mohanta, L.C.; Huque, A. Metal contamination of commercial fish feed and quality aspects of farmed tilapia (Oreochromis niloticus) in Bangladesh. Biores. Commun. 2017, 3, 345–353. [Google Scholar]
- Choi, M.H.; Cech, J.J. Unexpectedly high mercury level in pelleted commercial fish feed. Environ. Toxicol. Chem. 1998, 17, 1979–1981. [Google Scholar] [CrossRef]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Seiler, C.; Berendonk, T.U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stobberingh, E.E. Antibiotic resistance of motile aeromonads in indoor catfish and eel farms in the southern part of The Netherlands. Int. J. Antimicrob. Agents 2008, 31, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, L.; Chenia, H.Y. Characterization of integrons and tetracycline resistance determinants in Aeromonas spp. isolated from South African aquaculture systems. Int. J. Food Microbiol. 2007, 114, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, D.T. Bacterial zoonoses of fishes: A review and appraisal of evidence for linkages between fish and human infections. Vet. J. 2015, 203, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Haenen, O.L.M. Bacterial infections from aquatic species: Potential for and prevention of contact zoonoses. Rev. Sci. Tech. 2013, 32, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Rivas, A.J.; Lemos, M.L.; Osorio, C.R. Photobacterium damselae subsp. damselae, a bacterium pathogenic for marine animals and humans. Front. Microb. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.; Torres, C.; Barros, J.; Somalo, S.; Igrejas, G.; Poeta, P. Gilthead seabream (Sparus aurata) as carriers of SHV-12 and TEM-52 extended –spectrum beta-lactamases-containing Escherichia coli isolates. Foodborne Pathog. Dis. 2011, 8, 1139–1141. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.H.; Park, S.G.; Choi, S.M.; Hwang, Y.O.; Ham, H.J.; Kim, S.U.; Lee, Y.K.; Kim, M.S.; Park, G.Y.; Kim, K.S.; et al. Antimicrobial resistance and resistance genes in Escherichia coli strains isolated from commercial fish and seafood. Int. J. Food Microbiol. 2012, 152, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Chiu, T.H.; Kao, L.Y.; Chen, M.L. Antibiotic resistance and molecular typing of Photobacterium damselae subsp. damselae, isolated from seafood. J. Appl. Microbiol. 2013, 114, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Lekshmi, M.; Parvathi, A.; Nayak, B.B.; Varela, M.F. Antibiotic Resistance in Seafood Borne Pathogens. In Food Borne Pathogens and Antibiotic Resistance; Singh, O.V., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Chen, H.; Liu, S.; Xu, X.R.; Liu, S.S.; Zhou, G.J.; Sun, K.F.; Zhao, J.L.; Ying, G.G. Antibiotics in typical marine aquaculture farms surrounding Hailing Island, South China: Occurrence, bioaccumulation and human dietary exposure. Mar. Pollut. Bull. 2015, 90, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Rigos, G.; Nengas, I.; Alexis, M.; Troisi, G.M. Potential drug (oxytetracycline and oxolinic acid) pollution from Mediterranean sparid fish farms. Aquat. Toxicol. 2004, 69, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Huerta, B.; Marti, E.; Gros, M.; López, P.; Pompêo, M.; Armengol, J.; Barceló, D.; Balcázar, J.L.; Rodríguez-Mozaz, S.; Marcé, R. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs. Sci. Total Environ. 2013, 456–457, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.G.; Weston, D.P.; Miller, V.; Shoemaker, C. Antibacterial residues in marine sediments and invertebrates following chemotherapy in aquaculture. Aquaculture 1996, 145, 55–75. [Google Scholar] [CrossRef]
- Rico, A.; Oliveira, R.; McDonough, S.; Matser, A.; Khatikarn, J.; Satapornvanit, K.; Nogueira, A.J.; Soares, A.M.; Domingues, I.; Van den Brink, P.J. Use, fate and ecological risks of antibiotics applied in tilapia cage farming in Thailand. Environ. Pollut. 2014, 191, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Mao, D.; Luo, Y.; Wang, L.; Xu, B.; Xu, L. Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res. 2012, 46, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Sun, Y.; Zhang, T.; Ding, X.; Li, Y.; Wang, M.; Zeng, Z. Antibiotics, antibiotic resistance genes, and bacterial community composition in fresh water aquaculture environment in China. Microb. Ecol. 2015, 70, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Coyne, R.; Hiney, M.; O’Connor, B.; Kerry, J.; Cazabon, D.; Smith, P. Concentration and persistence of oxytetracycline in sediments under a marine salmon farm. Aquaculture 1994, 123, 31–42. [Google Scholar] [CrossRef]
- Chen, B.; Liang, X.; Huang, X.; Zhang, T.; Li, X. Differentiating anthropogenic impacts on ARGs in the Pearl River Estuary by using suitable gene indicators. Water Res. 2013, 47, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Luo, Y.; Mathieu, J.; Wang, Q.; Feng, L.; Mu, Q.; Alvarez, P.J.J. Persistence of extracellular DNA in river sediment facilities antibiotic resistance gene propagation. Environ. Sci. Technol. 2013, 48, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, N.; Laffite, A.; Graham, N.D.; Meijer, M.; Prabakar, K.; Mubedi, J.I.; Elongo, V.; Mpiana, P.T.; Ibelings, B.W.; Wildi, W.; et al. Accumulation of clinically relevant antibiotic-resistance genes, bacterial load, and metals in freshwater lake sediments in central Europe. Environ. Sci. Technol. 2015, 49, 6528–6537. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Kim, S.C.; Carlson, K.H.; Pruden, A. Effect of river landscape on the sediment concentrations of antibiotics and corresponding antibiotic resistance genes (ARG). Water Res. 2006, 40, 2427–2435. [Google Scholar] [CrossRef] [PubMed]
- Czekalski, N.; Gascón Díez, E.; Bürgmann, H. Wastewater as a point source of antibiotic-resistance genes in the sediment of a freshwater lake. ISME J. 2014, 8, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, C.; Shu, C.; Liu, L.; Geng, J.; Hu, S.; Feng, J. Marine sediment bacteria harbor antibiotic resistance genes highly similar to those found in human pathogens. Microb. Ecol. 2013, 65, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Antibiotics in the aquatic environment-a review-part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Tello, A.; Keen, P.L. Mechanisms of antimicrobial resistance in finfish aquaculture environments. Front. Microbiol. 2013, 4, 233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Zhao, Y.; Li, B.; Huang, C.L.; Zhang, S.Y.; Yu, S.; Chen, Y.S.; Zhang, T.; Gillings, M.R.; Su, J.Q. Continental-scale pollution of estuaries with antibiotic resistance genes. Nat. Microbiol. 2017, 2, 16270. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martínez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. PNAS 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [PubMed]
- Tomova, A.; Ivanova, L.; Buschmann, A.H.; Rioseco, M.L.; Kalsi, R.K.; Godfrey, H.P.; Cabello, F.C. Antimicrobial resistance genes in marine bacteria and human uropathogenic Escherichia coli from a region of intensive aquaculture. Environ. Microbiol. Rep. 2015, 7, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Bostock, J.; McAndrew, B.; Richards, R.; Jauncey, K.; Telfer, T.; Lorenzen, K.; Little, D.; Ross, L.; Handisyde, N.; Gatward, I.; et al. Aquaculture: Global status and trends. Philos. Trans. R. Soc. B 2010, 365, 2897–2912. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Andersen, J.S.; Kaewmak, T.; Somsiri, T.; Dalsgaard, A. Impact of integrated fish farming on antimicrobial resistance in a pond environment. Appl. Environ. Microbiol. 2002, 68, 6036–6042. [Google Scholar] [CrossRef] [PubMed]
- Cripps, S.J.; Bergheim, A. Solids management and removal for intensive land-based aquaculture production systems. Aquac. Eng. 2000, 22, 33–56. [Google Scholar] [CrossRef]
- Turcios, A.E.; Papenbrock, J. Sustainable treatment of aquaculture effluents-what can we learn from the past for the future? Sustainability 2014, 6, 836–856. [Google Scholar] [CrossRef]
- Gravel, V.; Dorais, M.; Dey, D.; Vandenberg, G. Fish effluents promote root growth and suppress fungal diseases in tomato transplants. Can. J. Plant Sci. 2015, 95, 427–436. [Google Scholar] [CrossRef]
- Martínez, J.L. Antibiotics and antibiotic resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Wellington, E.M.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Gelfand, I.; Barak, Y.; Even-Chen, Z.; Cytryn, E.; Van Rijn, J.; Krom, M.D.; Neori, A. A novel zero discharge intensive seawater recirculating system for the culture of marine fish. J. World Aquac. Soc. 2003, 34, 344–358. [Google Scholar] [CrossRef]
- Zohar, Y.; Tal, Y.; Schreier, H.J.; Steven, C.R.; Stubblefield, J.; Place, A.R. Commercially feasible urban recirculating aquaculture: Addressing the marine sector. In Urban Aquaculture; Costa-Pierce, B., Desbonnet, A., Edwards, P., Baker, D., Eds.; CABI Publishing: Cambridge, MA, USA, 2005; pp. 159–171. [Google Scholar]
- Tal, Y.; Schreier, H.J.; Sowers, K.R.; Stubblefield, J.D.; Place, A.R.; Zohar, Y. Environmentally sustainable land-based marine aquaculture. Aquaculture 2009, 286, 28–35. [Google Scholar] [CrossRef]
- Schreier, H.J.; Mirzoyan, N.; Saito, K. Microbial diversity of biological filters in recirculating aquaculture systems. Curr. Opin. Biotechnol. 2010, 21, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, S.; Ye, C.; Lin, W.; Zhang, M.; Chen, L.; Li, J.; Yu, X. Biofilm process in treating mariculture wastewater may be a reservoir of antibiotic resistance genes. Mar. Pollut. Bull. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Blancheton, J.P.; Attramadal, K.J.K.; Michaud, L.; d’Orbcastel, E.R.; Vadstein, O. Insight into bacterial population in aquaculture systems and its implication. Aquac. Eng. 2013, 53, 30–39. [Google Scholar] [CrossRef]
- Bebak-Williams, J.; Bullock, G.; Carson, M.C. Oxytetracycline residues in a freshwater recirculating system. Aquaculture 2002, 205, 221–230. [Google Scholar] [CrossRef]
- Saavedra, M.J.; Guedes-Novais, S.; Alves, A.; Rema, P.; Tacão, M.; Correia, A.; Martínez-Murcia, A. Resistance to β-lactam antibiotics in Aeromonas hydrophila isolated from rainbow trout (Onchorhynchus mykiss). Int. Microbiol. 2010, 7, 207–211. [Google Scholar]
- Krkošek, M. Host density thresholds and disease control for fisheries and aquaculture. Aquac. Environ. Interact. 2010, 1, 21–32. [Google Scholar] [CrossRef]
- Shah, S.Q.; Colquhoun, D.J.; Nikuli, H.L.; Sorum, H. Prevalence of antibiotic resistance genes in the bacterial flora of integrated fish farming environments of Pakistan and Tanzania. Environ. Sci. Technol. 2012, 46, 8672–8679. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Nonaka, L.; Suzuki, S. Occurrence of tetracycline resistance genes tet (M) and tet (S) in bacteria from marine aquaculture sites. FEMS Microb. Lett. 2004, 237, 147–156. [Google Scholar] [CrossRef]
- Shah, S.Q.; Cabello, F.C.; L’Abée-Lund, T.M.; Tomova, A.; Godfrey, H.P.; Buschmann, A.H.; Sørum, H. Antimicrobial resistance and antimicrobial resistance genes in marine bacteria from salmon aquaculture and non-aquaculture sites. Environ. Microb. 2014, 16, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Blanco, A.; Lemos, M.L.; Osorio, C.R. Integrating conjugative elements as vectors of antibiotic, mercury, and quaternary ammonium compound resistance in marine aquaculture environments. Antimicrob. Agents Chemother. 2012, 56, 2619–2626. [Google Scholar]
- Nguyen, H.N.K. Molecular characterization of antibiotic resistance in Pseudomonas and Aeromonas isolates from catfish of the Mekong Delta, Vietnam. Vet. Microbiol. 2014, 171, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Rzeczycka, M.; Ostrowski, R.; Popowska, M. Diversity of antibiotic resistance among bacteria isolated from sediments and water of carp farms located in a Polish nature reserve. Pol. J. Environ. Stud. 2017, 26, 239–252. [Google Scholar] [CrossRef]
- Jiang, H.X.; Tang, D.; Liu, Y.H.; Zhang, X.H.; Zeng, Z.L.; Xu, L.; Hawkey, P.M. Prevalence and characteristics of β-lactamase and plasmid-mediated quinolone resistance genes in Escherichia coli isolated from farmed fish in China. J. Antimicrob. Chemother. 2012, 67, 2350–2353. [Google Scholar] [CrossRef] [PubMed]
- Little, D.C.; Edwards, P. Integrated Livestock-Fish Farming Systems; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2003. [Google Scholar]
- Hoa, P.T.P.; Managaki, S.; Nakada, N.; Takada, H.; Shimizu, A.; Anh, D.H.; Viet, P.H.; Suzuki, S. Antibiotic contamination and occurrence of antibiotic-resistant bacteria in aquatic environments of northern Vietnam. Sci. Total Environ. 2011, 409, 2894–2901. [Google Scholar] [CrossRef] [PubMed]
- Neela, F.A.; Banu, M.N.A.; Rahman, M.A.; Rahman, M.H.; Alam, M.F. Occurrence of antibiotic resistant bacteria in pond water associated with integrated poultry-fish farming in Bangladesh. Sains Malays. 2014, 44, 371–377. [Google Scholar] [CrossRef]
- White, P. Environmental consequences of poor feed quality and feed management. In On-Farm Feeding and Feed Management in Aquaculture; Hasan, M.R., New, M.B., Eds.; FAO Fisheries and Aquaculture Technical Paper No. 583; FAO: Rome, Italy, 2013; pp. 553–564. [Google Scholar]
- Cao, L.; Naylor, R.; Henriksson, P.; Leadbitter, D.; Metian, M.; Troell., M.; Zhang, W. China’s aquaculture and the world’s wild fisheries. Science 2015, 347, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.B. Probiotics, the other half of the antibiotic story. Anim. Nutr. Health 1974, 29, 4–8. [Google Scholar]
- Food and Agricultural Organization of the United Nations and World Health Organization. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Hill, C.; Garner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Kozasa, M. Toyocerin (Bacillus toyoi) as growth promotor for animal feeding. Microbiol. Aliment. Nutr. 1986, 4, 121–135. [Google Scholar]
- Lazado, C.C.; Marlowe, C.; Caipang, A.; Estante, E.G. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions. Fish Shellfish Immunol. 2015, 45, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Lazado, C.C.; Safari, R.; Yeganeh, S.; Zorriehzahra, M.J. Aqualase®, a yeast-based in-feed probiotic, modulates intestinal microbiota, immunity and growth of rainbow trout Oncorhynchus mykiss. Aquac. Res. 2017, 48, 1815–1826. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Ruiz-Zarzuela, I.; Blas, I.; Balcázar, J.L. Probiotics in aquaculture: A current assessment. Rev. Aquac. 2014, 6, 133–146. [Google Scholar] [CrossRef]
- Iwashita, M.K.; Nakandakare, I.B.; Terhune, J.S.; Wood, T.; Ranzani-Paiva, M.J. Dietary supplementation with Bacillus subtilis, Saccharomyces cerevisiae and Aspergillus oryzae enhance immunity and disease resistance against Aeromonas hydrophila and Streptococcus iniae infection in juvenile tilapia Oreochromis niloticus. Fish Shellfish Immunol. 2015, 43, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Newaj-Fyzul, A.; Austin, B. Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J. Fish Dis. 2015, 38, 937–955. [Google Scholar] [CrossRef] [PubMed]
- Resende, J.A.; Borges, M.L.; Pacheco, K.D.; Ribeiro, I.H.; Cesar, D.E.; Silva, V.L.; Diniz, C.G.; Apolônio, A.C.M. Antibiotic resistance in potentially bacteriocinogenic probiotic bacteria in aquaculture environments. Aquac. Res. 2016, 1–7. [Google Scholar] [CrossRef]
- Gillor, O.; Etzion, A.; Riley, M.A. The dual role of bacteriocins as anti- and probiotics. Appl. Microbiol. Biotechnol. 2008, 81, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Imperial, I.C.V.J; Ibana, J.A. Addressing the antibiotic resistance problem with probiotics: Reducing the risk of its double-edged sword effect. Front. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; Del Campo, R.; Hernández, P.E.; Herranz, C.; Cintas, L.M. Antimicrobial activity, antibiotic susceptibility and virulence factors of lactic acid bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. The Review on Antimicrobial Resistance. Available online: http://amr-review.org/Publications.html (accessed on 25 May 2017).
- O’Neill, J. Antimicrobial Resistance Review: Government Response London: Wellcome Trust and Department of Health. Available online: http://amr-review.org/Publications.html (accessed on 25 May 2017).
- Gudding, R.; Van Muiswinkel, W.B. A history of fish vaccination: Science-based disease prevention in aquaculture. Fish Shellfish Immunol. 2013, 35, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Olsen, R.E.; Gifstad, T.Ø.; Dalmo, R.A.; Amlund, H.; Hemre, G.I.; Bakke, A.M. Prebiotics in aquaculture: A review. Aquac. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Cunha, A.; Calado, R.; Gomes, N.C.; Almeida, A. Influence of environmental variables in the efficiency of phage therapy in aquaculture. Microb. Biotechnol. 2014, 7, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, M.; Quan, C.S.; Fan, S.D. Mechanisms of quorum sensing and strategies for quorum sensing disruption in aquaculture pathogens. J. Fish Dis. 2015, 38, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Summerfelt, S.T. Ozonation and UV irradiation—An introduction and examples of current applications. Aquac. Eng. 2003, 28, 21–36. [Google Scholar] [CrossRef]
- Powell, A.; Scolding, J.W.S. Direct application of ozone in aquaculture systems. Rev. Aquac. 2016, 1–15. [Google Scholar] [CrossRef]


{kind=link}
| Antibiotic Class | Antibiotic Resistance Gene | Aquaculture System or Fish Species | Reference |
|---|---|---|---|
| β-Lactam (e.g., Ampicillin, Amoxicillin) | blaTEM-52, blaSHV-12 | Gilthead Seabream | Sousa et al. [55] |
| blaTEM | Fish farms, Pakistan and Tanzania | Shah et al. [96] | |
| Tetracycline (tetracycline, oxytetracycline, chlortetracycline) | tetM, tetO, tetT, tetQ | Fish farms, Tianjin, and Guangdong, China | Gao et al. [64], Xiong et al. [65] |
| tetM, tetS | Japanese and Korean coastal farms | Kim et al. [97] | |
| tetA, tetG | Chilean salmon | Shah et al. [98] | |
| Fish farms, Pakistan and Tanzania | Shah et al. [96] | ||
| tetA | Marine aquaculture, Spain and Portugal | Rodriguez-Blanco et al. [99] | |
| tetA, tetB, tetK | Salmon aquaculture, Chile | Buschmann et al. [19] | |
| Sulfonamide, sulfamethizole | sul1, sul2, sul3 | Fish farms, Tianjin, China; farmed freshwater fish, Guangdong, China; Gilthead seabream | Sousa et al. [55], Gao et al. [64], Xiong et al. [65] |
| sul1, sul2 | Chilean salmon; fish farms, Tanzania and Pakistan | Shah et al. [96], Shah et al. [98] | |
| Aminoglycoside (Streptomycin, spectinomycin, neomycin) | aadA strA-strB | Chilean salmon, fish farms, Tanzania and Pakistan; catfish farm, Vietnam; carp farms, Poland | Shah et al. [96], Shah et al. [98], Nguyen et al. [100], Piotrowska et al. [101] |
| aad1 | Gilthead Seabream | Sousa et al. [55] | |
| Amphenicol (chloramphenicol, florfenicol) | cmlA | Gilthead Seabream | Sousa et al. [55] |
| cat-1 | Fish farms, Tanzania and Pakistan | Shah et al. [96] | |
| floR | Salmon aquaculture, Chile | Buschmann et al. [19] | |
| catB | Catfish farm, Vietnam | Nguyen, et al. [100] | |
| Quinolones (oxolinic acid, ciproflaxin) | qepA, oqxAB, qnrS, aac(6′)-Ib, qnrB, qnrD | Farmed freshwater fish, Guangdong, China | Shah et al. [98], Jiang et al. [102] |
| qnrA, qnrB, qnrS | Salmon aquaculture, Chile | Buschmann et al. [19] | |
| Macrolides (erythromycin) | mefA | Fish farms, Tanzania | Shah et al. [96] |
| ermC, ermE, ermX, ermC | Carp farms, Poland | Piotrowska et al. [101] | |
| Trimethoprim | dfrA1, dfrA5, dftA12 | Chilean salmon; fish farms, Tanzania and Pakistan | Shah et al. [96], Shah et al. [98] |
| dfrA12 | Catfish farm, Vietnam | Nguyen et al. [100] | |
| Quinoxoline 1, 4-di-N-oxides (carbadox, olaquindox, mequindox) | oqxA | Salmon aquaculture, Chile | Buschmann et al. [19] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Mar. Drugs 2017, 15, 158. https://doi.org/10.3390/md15060158
Watts JEM, Schreier HJ, Lanska L, Hale MS. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Marine Drugs. 2017; 15(6):158. https://doi.org/10.3390/md15060158
Chicago/Turabian StyleWatts, Joy E. M., Harold J. Schreier, Lauma Lanska, and Michelle S. Hale. 2017. "The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions" Marine Drugs 15, no. 6: 158. https://doi.org/10.3390/md15060158
APA StyleWatts, J. E. M., Schreier, H. J., Lanska, L., & Hale, M. S. (2017). The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Marine Drugs, 15(6), 158. https://doi.org/10.3390/md15060158