Deep Sea Water Improves Abnormalities in Lipid Metabolism through Lipolysis and Fatty Acid Oxidation in High-Fat Diet-Induced Obese Rats


Abstract
:1. Introduction
2. Results
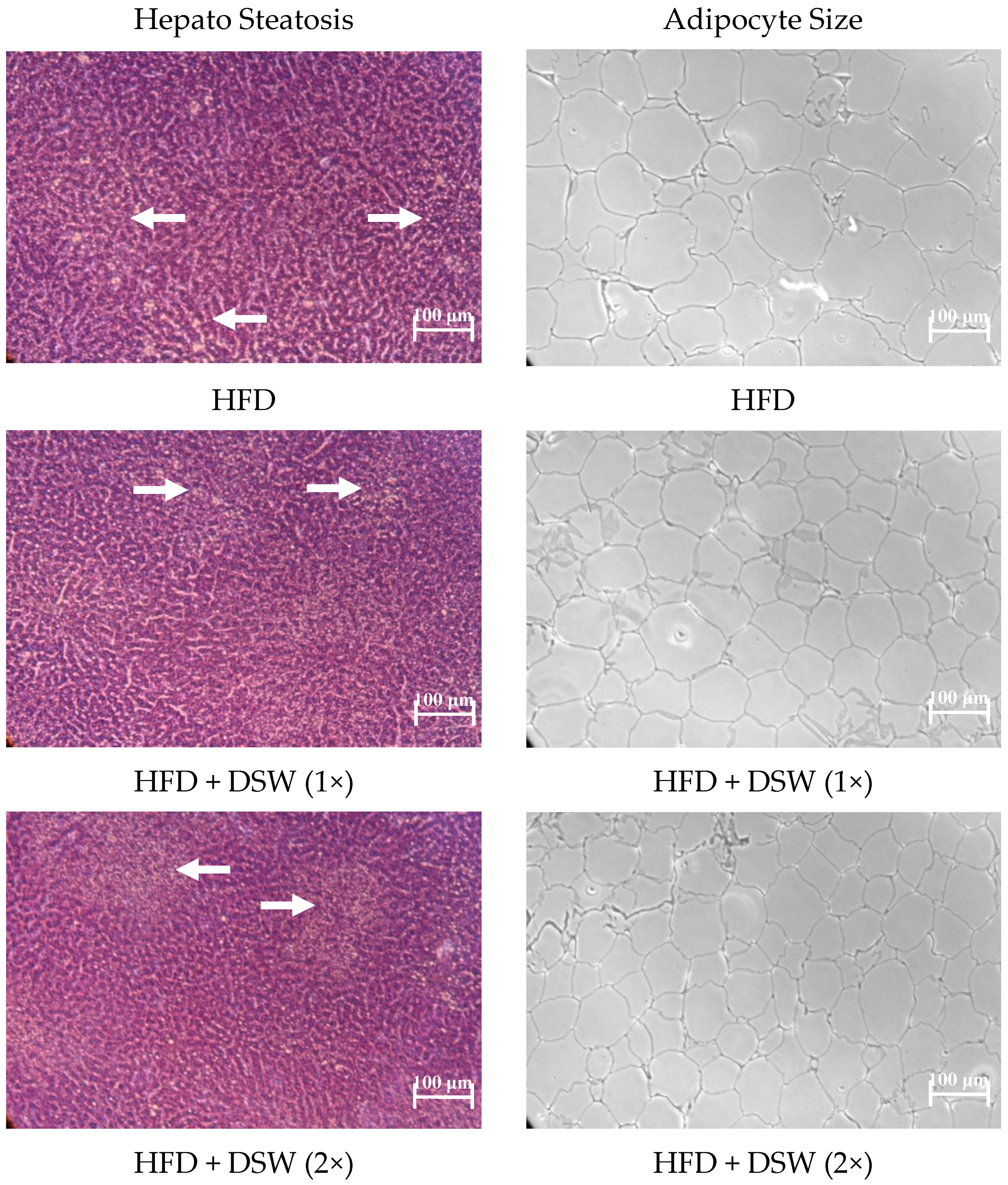
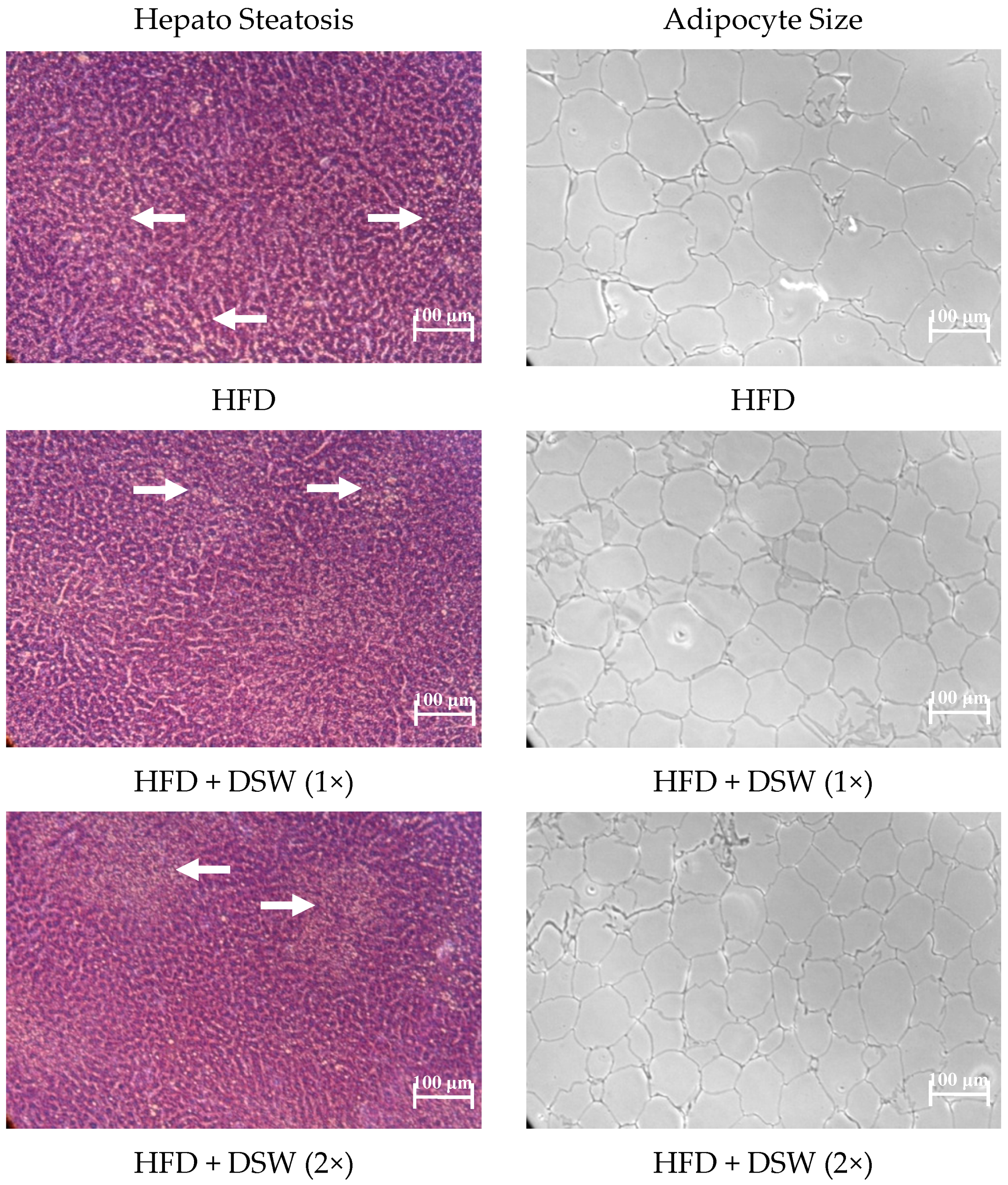
2.1. Effect of DSW on Body Weight, Water Intake, Food Intake, Energy Intake, Organ Size, and Adipose Tissue in Rats with HFD-Induced Obesity
2.2. Effect of DSW on the Serum Biochemical Indicators in Rats with HFD-Induced Obesity
2.3. Effect of DSW on Hepatic Antioxidant Enzyme and Total Hepatic and Fecal Lipids, Triglycerides, Cholesterol, and Mg2+ in Rats with HFD-Induced Obesity
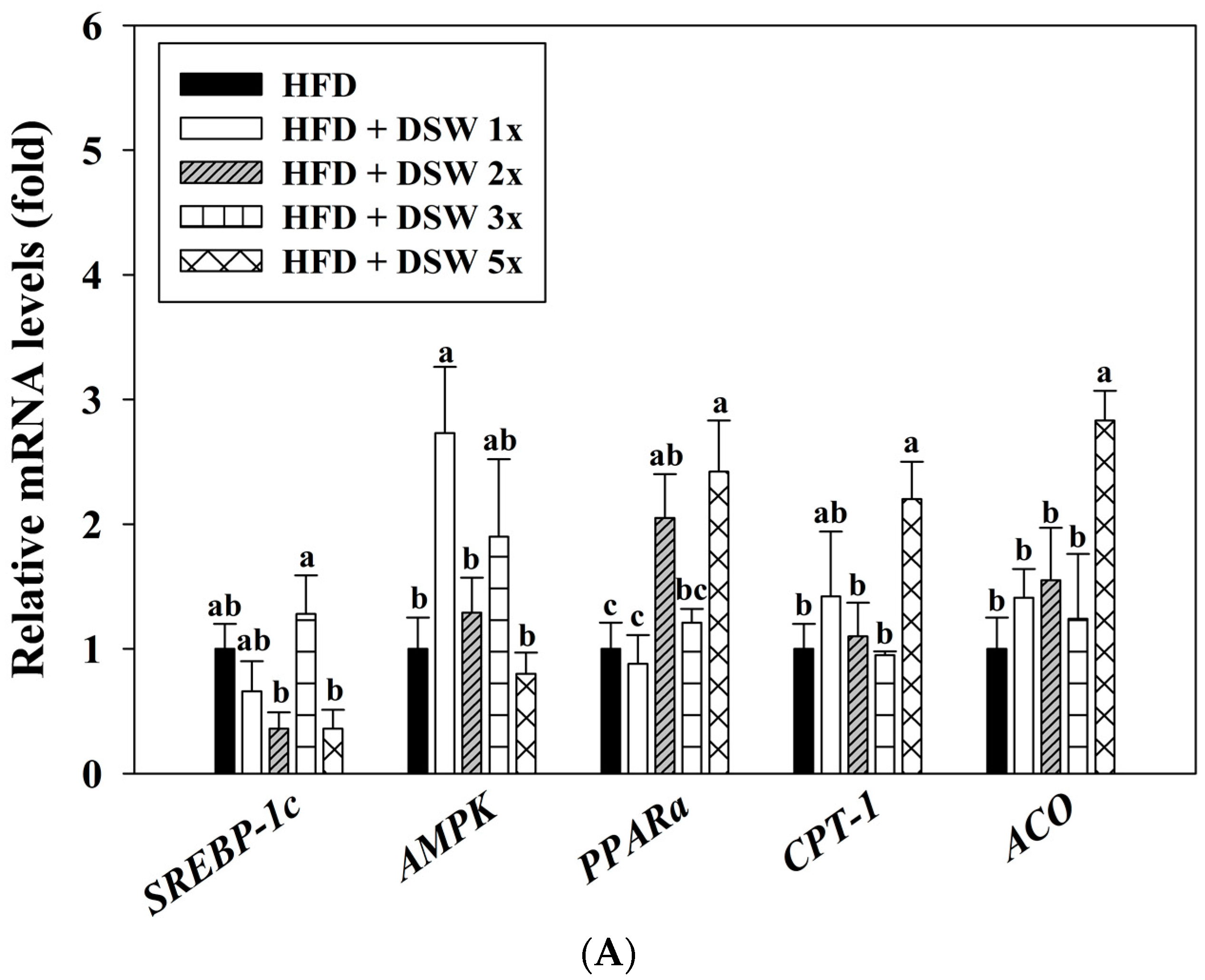
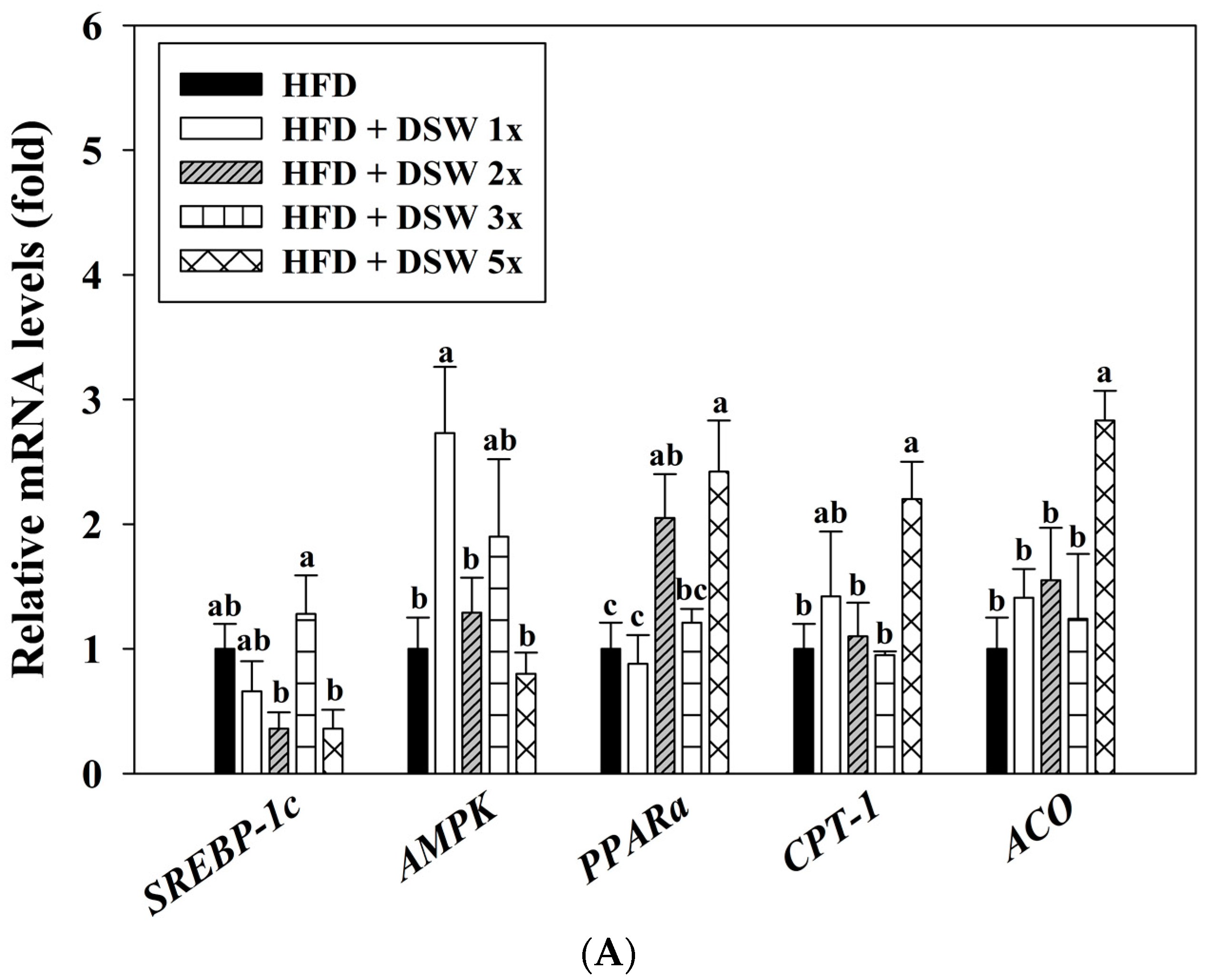
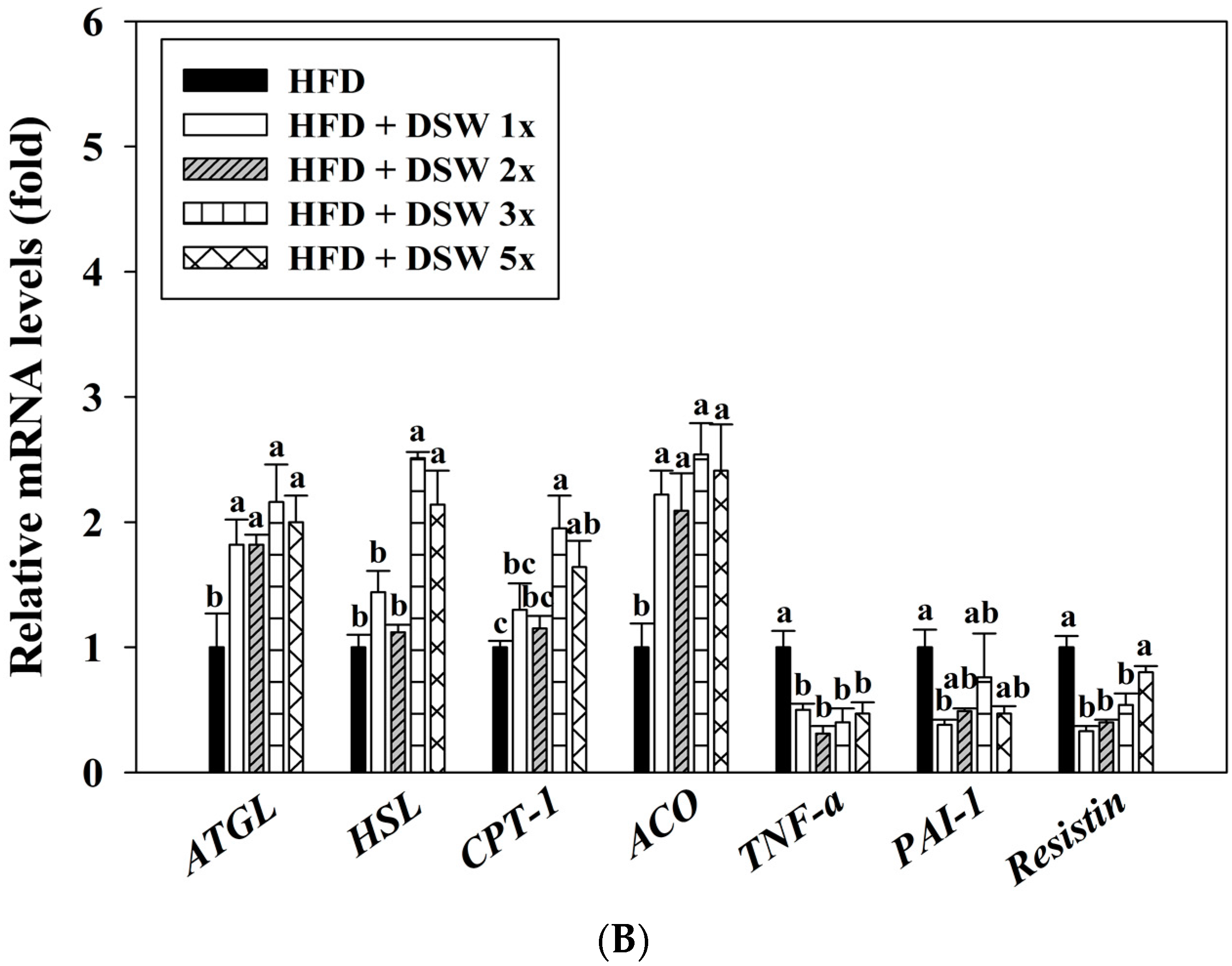
2.4. Effects of DSW on Gene Expressions in Rats with HFD-Induced Obesity
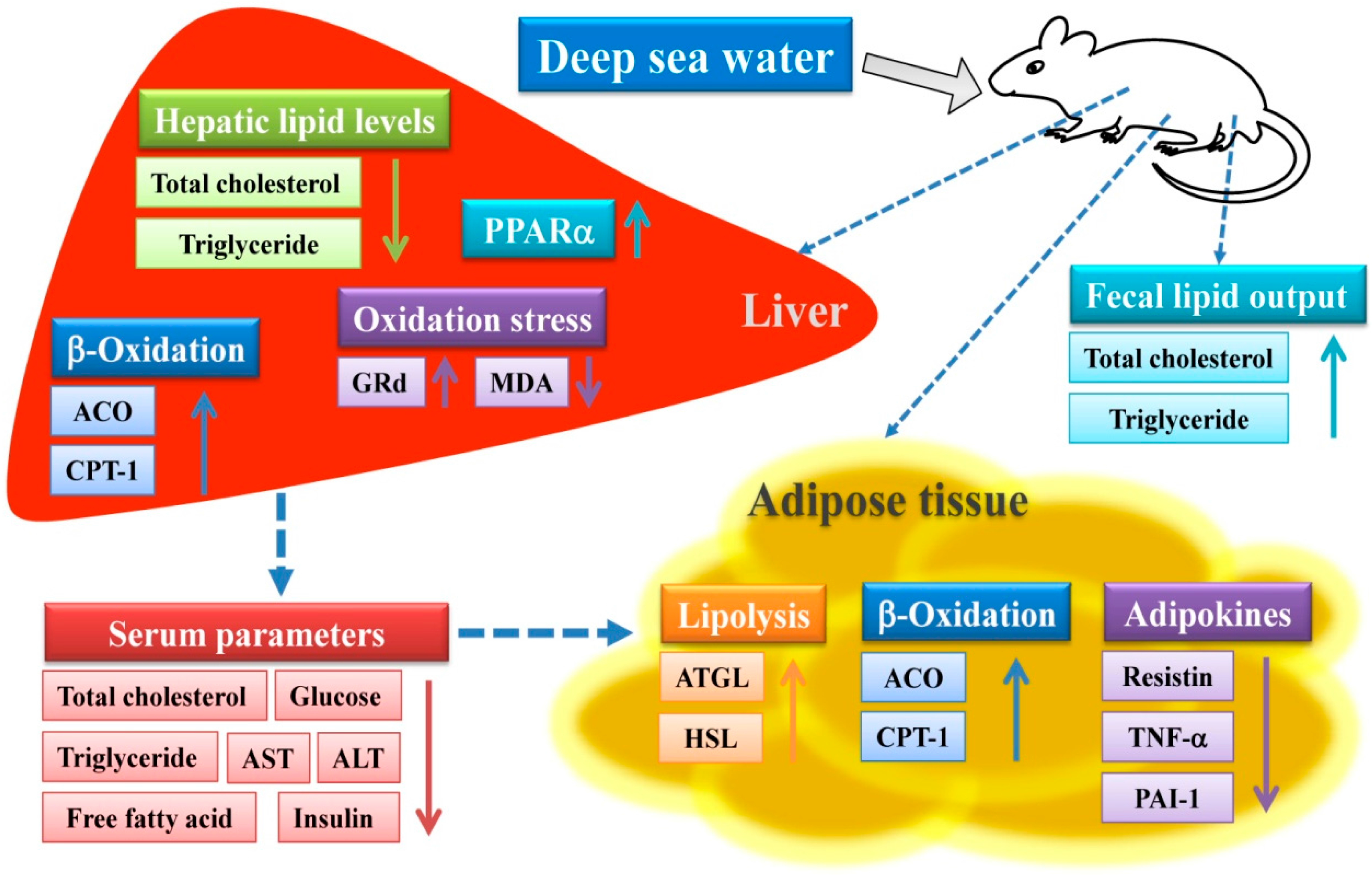
3. Discussion
4. Materials and Methods
4.1. Collection of Deep Sea Water (DSW)
4.2. Animals, Diets, and Experimental Design
4.3. Measurement of Serum Parameters
4.4. Hematoxylin/Eosin (H&E) Staining
4.5. Hepatic and Fecal Lipid Analysis
4.6. Hepatic Lipid Peroxidation Assay
4.7. TEAC Assay
4.8. Determination of Antioxidant Enzymes in the Liver
4.9. RNA Extraction and Real-Time RT-PCR
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACO | acyl-CoA oxidase |
| ALT | alanine transaminase |
| AMPK | AMP-activated protein kinase |
| AST | aspartate aminotransferase |
| ATGL | adipose triglyceride lipase |
| CPT | carnitine palmitoyl transferase |
| GRd | glutathione reductase |
| GST | glutathione S-transferase |
| HDL cholesterol | high-density lipoprotein cholesterol |
| HSL | hormone-sensitive lipase |
| IL | interleukin |
| LDL cholesterol | low-density lipoprotein cholesterol |
| MDA | malondialdehyde |
| PAI | plasminogen activator inhibitor |
| PPAR | peroxisome proliferator-activated receptor |
| SREBP | sterol regulatory element binding protein |
| TBARS | thiobarbituric acid reactive substances |
| TEAC | Trolox equivalent antioxidant capacity |
| TNF | tumor necrosis factor |
References
- Van Greevenbroek, M.M.J.; Schalkwijk, C.G.; Stehouwer, C.D.A. Dysfunctional adipose tissue and low-grade inflammation in the management of the metabolic syndrome: Current practices and future advances. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N. Innate immune activation in obesity. Mol. Asp. Med. 2013, 34, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Rossen, L.M.; Rossen, E.A. Obesity 101; Springer: New York, NY, USA, 2012. [Google Scholar]
- Berrington de Gonzalez, A.; Hartge, P.; Cerhan, J.R.; Flint, A.J.; Hannan, L.; MacInnis, R.J.; Moore, S.C.; Tobias, G.S.; Anton-Culver, H.; Freeman, L.B.; et al. Body-mass index and mortality among 1.46 million white adults. N. Engl. J. Med. 2010, 363, 2211–2219. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.G.; Apovian, C.M.; Aronne, L.J. New drug targets for the treatment of obesity. Clin. Pharmacol. Ther. 2011, 90, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Trigueros, L.; Peña, S.; Ugidos, A.V.; Sayas-Barberá, E.; Pérez-Álvarez, J.A.; Sendra, E. Food ingredients as anti-obesity agents: A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kim, S.H.; Yoo, Y.G.; Chu, Y.S.; Shon, Y.H.; Nam, K.S.; Yun, J.W. Inhibitory effect of deep-sea water on differentiation of 3T3-L1 adipocytes. Mar. Biotechnol. 2009, 11, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kim, H.A.; Lee, S.H.; Yun, J.W. Anti-obesity and antidiabetic effects of deep sea water on ob/ob mice. Mar. Biotechnol. 2009, 11, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Toyota, T.; Nakashima, T. Comparison of the effects of water-soluble (EDTA) and particle (chelex-100) synthetic ligands on the growth of phytoplankton population in the disphotic zone seawater. J. Phys. Oceanogr. 1998, 54, 19–28. [Google Scholar] [CrossRef]
- Katsuda, S.; Yasukawa, T.; Nakagawa, K.; Miyake, M.; Yamasaki, M.; Katahira, K.; Mohri, M.; Shimizu, T.; Hazama, A. Deep-sea water improves cardiovascular hemodynamics in Kurosawa and Kusanagi-Hypercholesterolemic (KHC) rabbits. Biol. Pharm. Bull. 2008, 31, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.G.; Moon, D.S.; Kim, H.J.; Shon, Y.H. Magnesium and calcium-enriched deep-sea water promotes mitochondrial biogenesis by AMPK-activated signals pathway in 3T3-L1 preadipocytes. Biomed. Pharmacother. 2016, 83, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Lung, T.Y.; Kung, Y.H.; Wang, J.J.; Tsai, T.Y.; Wei, B.L.; Pan, T.M.; Lee, C.L. Enhanced anti-obesity activities of red mold dioscorea when fermented using deep ocean water as the culture water. Mar. Drugs 2013, 11, 3902–3925. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Chang, W.T.; Hsu, C.L. Inhibitory effect of deep sea water on adipogenesis in 3T3-L1 adipocytes. Chung Shan Med. J. 2015, 26, 35–41. [Google Scholar]
- Hsu, C.L.; Chang, Y.Y.; Chiu, C.H.; Yang, K.T.; Wang, Y.; Fu, S.G.; Chen, Y.C. Cardiovascular protection of deep-seawater drinking water in high-fat/cholesterol fed hamsters. Food Chem. 2011, 127, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hata, K.; Hiwatashi, K.; Hori, K.; Suzuki, N.; Itoh, H. Suppression of murine preadipocyte differentiation and reduction of visceral fat accumulation by a Petasitesjaponicus ethanol extract in mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2010, 74, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Yen, G.C. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2007, 55, 8404–8410. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Yen, G.C. Effects of capsaicin on induction of apoptosis and inhibition of adipogenesis in 3T3-L1 cells. J. Agric. Food Chem. 2007, 55, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Lin, Y.J.; Ho, C.T.; Yen, G.C. Inhibitory effects of garcinol and pterostilbene on cell proliferation and adipogenesis in 3T3-L1 cells. Food Funct. 2012, 3, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Yen, G.C. Phenolic compounds: Evidence for inhibitory effects against obesity and their underlying molecular signaling mechanisms. Mol. Nutr. Food. Res. 2008, 52, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Oscai, L.B.; Miller, W.C.; Arnall, D.A. Effects of dietary sugar and dietary fat on food intake and body fat content in rats. Growth 1987, 51, 64–73. [Google Scholar] [PubMed]
- Hsu, C.L.; Yen, G.C. Effect of gallic acid on high fat diet-induced dyslipidemia, hepatosteatosis, and oxidative stress in rats. Br. J. Nutr. 2007, 98, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Milani, R.V. Obesity and cardiovascular disease: The hippocrates paradox. J. Am. Coll. Cardiol. 2003, 42, 677–679. [Google Scholar] [CrossRef]
- Ouchi, Y.; Tabata, R.E.; Stergiopoulos, K.; Sato, F.; Hattori, A.; Orimo, H. Effect of dietary magnesium on development of atherosclerosis in cholesterol-fed rabbits. Arteriosclerosis 1990, 10, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Tzang, B.S.; Yang, T.Y.; Hsiao, Y.C.; Yang, H.C.; Chen, Y.C. Effects of deep-sea water on blood lipids and pressure in high-cholesteroldietary mice. J. Food Biochem. 2011, 35, 241–259. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.G.; Park, J.E.; Shin, E.J.; Shon, Y.H. Effects of balance deep-sea water on adipocyte hypertrophy and liver steatosis in high-fat diet-induced obese mice. Obesity 2014, 22, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Nani, S.Z.M.; Majid, F.A.A.; Jaafar, A.B.; Mahdzir, A.; Musa, M.N. Potential Health Benefits of Deep Sea Water: A Review. Evid. Based Complement. Altern. Med. 2016, 2016, 6520475. [Google Scholar] [CrossRef]
- Bertinato, J.; Lavergne, C.; Rahimi, S.; Rachid, H.; Vu, N.A.; Plouffe, L.J.; Swist, E. Moderately low magnesium intake impairs growth of lean body mass in obese-prone and obese-resistant rats fed a high-energy diet. Nutrients 2016, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Vaskonen, T.; Mervaala, E.; Krogerus, L.; Karppanen, H. Supplementation of plant sterols and minerals benefits obese Zucker rats fed an atherogenic diet. J. Nutr. 2002, 132, 231–237. [Google Scholar] [PubMed]
- Berezovskiĭ, M.E. Method of staining of semi-thin sections with hematoxylin-eosin. Arkhiv Pathol. 1978, 40, 69–70. [Google Scholar]
- Tzang, B.S.; Yang, S.F.; Fu, S.G.; Yang, H.C.; Sun, H.L.; Chen, Y.C. Effects of dietary flaxseed oil on cholesterol metabolism of hamsters. Food Chem. 2009, 114, 1450–1455. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Arnao, M.B.; Cano, A.; Hernandez-Ruiz, J.; Garcia-Canovas, F.; Acosta, M. Inhibition by L-ascorbic acid and other antioxidants of the 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) oxidation catalyzed by peroxidase: A new approach for determining total antioxidant status of foods. Anal. Biochem. 1996, 236, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Bellomo, G.; Mirabelli, F.; Dimonte, D.; Richelmi, P.; Thor, H.; Orrenius, C. Formation and reduction of glutathione-mixed disulfides during oxidative stress. Biochem. Pharmacol. 1987, 36, 1313–1320. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferase: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Obese Rat * | HFD Supplemented with DSW | ||||
|---|---|---|---|---|---|
| Control | 1× | 2× | 3× | 5× | |
| Initial Weight (g) | 323.49 ± 2.35 a | 323.39 ± 2.36 a | 323.31 ± 2.30 a | 323.88 ± 1.79 a | 323.48 ± 1.97 a |
| Final Weight (g) | 510.35 ± 9.68 a | 478.46 ± 6.72 b | 480.23 ± 12.33 b | 459.61 ± 6.73 b | 457.95 ± 7.30 b |
| Water Intake (mL/day) | 44.32 ± 0.82 c | 48.76 ± 1.70 bc | 47.92 ± 2.42 bc | 59.49 ± 4.81 a | 53.93 ± 1.40 ab |
| Food Intake (g/rat/day) | 26.08 ± 0.89 a | 25.22 ± 0.74 a | 25.49 ± 0.69 a | 24.80 ± 0.36 a | 24.50 ± 0.76 a |
| Energy Intake (kcal/rat/day) | 129.59 ± 4.40 a | 125.27 ± 3.69 a | 126.62 ± 3.43 a | 123.22 ± 1.77 a | 121.70 ± 3.79 a |
| Heart (mg/g rat) | 2.72 ± 0.08 a | 2.83 ± 0.07 a | 2.79 ± 0.11 a | 2.91 ± 0.04 a | 2.79 ± 0.07 a |
| Liver (mg/g rat) | 27.19 ± 1.09 a | 25.86 ± 0.25 a | 24.09 ± 0.32 b | 24.00 ± 0.41 b | 23.91 ± 0.38 b |
| Spleen (mg/g rat) | 1.84 ± 0.06 a | 1.84 ± 0.06 a | 1.82 ± 0.07 a | 1.89 ± 0.08 a | 1.91 ± 0.12 a |
| Lung (mg/g rat) | 3.29 ± 0.12 a | 3.33 ± 0.14 a | 3.20 ± 0.09 a | 3.27 ± 0.08 a | 3.37 ± 0.09 a |
| Kidney (mg/g rat) | 6.22 ± 0.16 a | 6.09 ± 0.15 a | 6.28 ± 0.17 a | 6.20 ± 0.12 a | 6.17 ± 0.13 a |
| Perirenal Fat (mg/g rat) | 41.64 ± 2.87 a | 30.99 ± 1.55 bc | 34.00 ± 2.30 b | 24.42 ± 2.26 c | 28.04 ± 2.10 bc |
| Epididymal Fat (mg/g rat) | 26.09 ± 0.93 a | 23.34 ± 1.07 ab | 23.06 ± 1.90 abc | 20.57 ± 1.37 bc | 18.98 ± 1.43 c |
| Biochemical Indicators * | HFD Supplemented with DSW | ||||
|---|---|---|---|---|---|
| Control | 1× | 2× | 3× | 5× | |
| Glucose (mg/dL) | 242.75 ± 11.73 a | 228.88 ± 9.82 ab | 205.38 ± 10.58 bc | 185.63 ± 14.68 cd | 168.38 ± 9.85 d |
| Triglycerides (mg/dL) | 116.63 ± 9.63 a | 79.13 ± 4.42 bc | 85.88 ± 4.99 b | 68.75 ± 4.43 bc | 67.25 ± 3.44 c |
| Total Cholesterol (mg/dL) | 60.54 ± 4.72 a | 47.00 ± 2.97 b | 46.86 ± 4.53 b | 52.73 ± 3.28 ab | 45.03 ± 2.78 b |
| HDL-Cholesterol (mg/dL) | 29.60 ± 1.39 a | 30.16 ± 2.12 a | 30.22 ± 1.55 a | 28.88 ± 1.38 ab | 24.48 ± 1.55 b |
| LDL-Cholesterol (mg/dL) | 8.51 ± 0.68 ab | 7.26 ± 0.30 b | 7.05 ± 0.34 b | 9.44 ± 0.75 a | 7.35 ± 0.58 b |
| AST (U/L) | 69.00 ± 2.95 a | 64.50 ± 3.26 ab | 61.38 ± 1.93 ab | 60.25 ± 2.62 ab | 57.00 ± 3.22 b |
| ALT (U/L) | 22.88 ± 1.36 a | 18.50 ± 1.28 b | 18.34 ± 0.75 b | 18.50 ± 1.25 b | 16.50 ± 0.71 b |
| Uric Acid (mg/dL) | 4.39 ± 0.20 a | 4.07 ± 0.25 a | 4.12 ± 0.32 a | 3.64 ± 0.28 a | 3.93 ± 0.14 a |
| Creatinine (mg/dL) | 0.76 ± 0.02 a | 0.79 ± 0.01 a | 0.75 ± 0.02 a | 0.75 ± 0.02 a | 0.74 ± 0.02 a |
| Ketone Body (mmol/L) | 1.94 ± 0.10 bc | 2.08 ± 0.14 bc | 1.76 ± 0.06 c | 1.84 ± 0.05 c | 2.25 ± 0.13 a |
| Free Fatty Acid (mmol/dL) | 0.99 ± 0.03 a | 0.79 ± 0.02 c | 0.93 ± 0.04 ab | 0.84 ± 0.05 bc | 0.74 ± 0.04 c |
| Insulin (μg/L) | 1.04 ± 0.12 a | 0.62 ± 0.05 b | 0.71 ± 0.07 b | 0.57 ± 0.07 b | 0.55 ± 0.02 b |
| Mg2+ (mmol/L) | 2.61 ± 0.10 b | 2.92 ± 0.08 ab | 3.00 ± 0.12 a | 2.97 ± 0.10 a | 3.01 ± 0.13 a |
| Na+ (mmol/L) | 165.63 ± 3.39 a | 162.50 ± 2.42 a | 166.50 ± 4.96 a | 164.50 ± 3.71 a | 160.13 ± 3.37 a |
| K+ (mmol/L) | 6.41 ± 0.26 b | 6.69 ± 0.18 ab | 6.73 ± 0.18 ab | 6.80 ± 0.28 ab | 7.18 ± 0.24 a |
| Cl− (mmol/L) | 103.00 ± 1.88 a | 103.75 ± 1.41 a | 103.75 ± 2.31 a | 106.00 ± 2.56 a | 104.25 ± 1.58 a |
| TEAC (nmol/mL) | 242.75 ± 11.73 a | 228.88 ± 9.82 ab | 205.38 ± 10.58 bc | 185.63 ± 14.68 cd | 168.38 ± 9.85 d |
| Obese Rat * | HFD Supplemented with DSW | ||||
|---|---|---|---|---|---|
| Control | 1× | 2× | 3× | 5× | |
| Hepatic | |||||
| TEAC (nmol/mg protein) | 1.02 ± 0.01 a | 1.05 ± 0.01 a | 1.03 ± 0.04 a | 1.02 ± 0.01 a | 0.99 ± 0.03 a |
| TBARS (nmol MDA eq./mg protein) | 0.41 ± 0.08 a | 0.15 ± 0.04 b | 0.32 ± 0.06 ab | 0.12 ± 0.04 b | 0.37 ± 0.12 a |
| GRd (nmol/mg protein) | 9.76 ± 1.03 b | 11.17 ± 0.77 b | 9.89 ± 0.48 b | 14.34 ± 0.78 a | 16.30 ± 1.57 a |
| GST (nmol/mg protein) | 18.13 ± 1.48 ab | 14.84 ± 1.15 b | 17.39 ± 1.91 ab | 18.56 ± 1.82 ab | 21.25 ± 1.35 a |
| Total Lipids (mg/g tissue) | 66.63 ± 2.76 a | 63.93 ± 2.42 ab | 62.31 ± 3.04 ab | 52.11 ± 1.80 c | 56.92 ± 2.78 bc |
| Triglycerides (mg/g tissue) | 22.96 ± 1.82 a | 23.17 ± 1.44 a | 17.24 ± 1.85 b | 17.72 ± 1.82 b | 17.91 ± 0.83 b |
| Cholesterol (mg/g tissue) | 9.55 ± 0.98 a | 7.98 ± 0.66 ab | 7.47 ± 1.05 ab | 6.50 ± 0.68 b | 6.77 ± 0.85 b |
| Magnesium (mg/g tissue) | 57.40 ± 7.95 b | 74.69 ± 8.19 ab | 66.92 ± 6.97 ab | 75.42 ± 5.78 ab | 85.81 ± 5.49 a |
| Fecal | |||||
| Total Lipids (mg/g dried fecal) | 51.52 ± 1.69 b | 61.46 ± 5.67 a | 64.04 ± 2.29 a | 68.22 ± 1.50 a | 63.62 ± 3.08 a |
| Triglycerides (mg/g dried fecal) | 7.93 ± 0.30 b | 8.70 ± 0.72 b | 12.34 ± 1.00 a | 11.60 ± 0.54 a | 13.09 ± 1.23 a |
| Cholesterol (mg/g dried fecal) | 6.70 ± 0.29 b | 6.48 ± 0.31 b | 8.30 ± 0.27 a | 7.95 ± 0.47 a | 8.33 ± 0.41 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-T.; Lu, T.-Y.; Cheng, M.-C.; Lu, H.-C.; Wu, M.-F.; Hsu, C.-L. Deep Sea Water Improves Abnormalities in Lipid Metabolism through Lipolysis and Fatty Acid Oxidation in High-Fat Diet-Induced Obese Rats. Mar. Drugs 2017, 15, 386. https://doi.org/10.3390/md15120386
Chang W-T, Lu T-Y, Cheng M-C, Lu H-C, Wu M-F, Hsu C-L. Deep Sea Water Improves Abnormalities in Lipid Metabolism through Lipolysis and Fatty Acid Oxidation in High-Fat Diet-Induced Obese Rats. Marine Drugs. 2017; 15(12):386. https://doi.org/10.3390/md15120386
Chicago/Turabian StyleChang, Wei-Tang, Tsung-Yueh Lu, Ming-Ching Cheng, Hsun-Chi Lu, Mei-Fang Wu, and Chin-Lin Hsu. 2017. "Deep Sea Water Improves Abnormalities in Lipid Metabolism through Lipolysis and Fatty Acid Oxidation in High-Fat Diet-Induced Obese Rats" Marine Drugs 15, no. 12: 386. https://doi.org/10.3390/md15120386
APA StyleChang, W.-T., Lu, T.-Y., Cheng, M.-C., Lu, H.-C., Wu, M.-F., & Hsu, C.-L. (2017). Deep Sea Water Improves Abnormalities in Lipid Metabolism through Lipolysis and Fatty Acid Oxidation in High-Fat Diet-Induced Obese Rats. Marine Drugs, 15(12), 386. https://doi.org/10.3390/md15120386