Evidence for a Saponin Biosynthesis Pathway in the Body Wall of the Commercially Significant Sea Cucumber Holothuria scabra

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
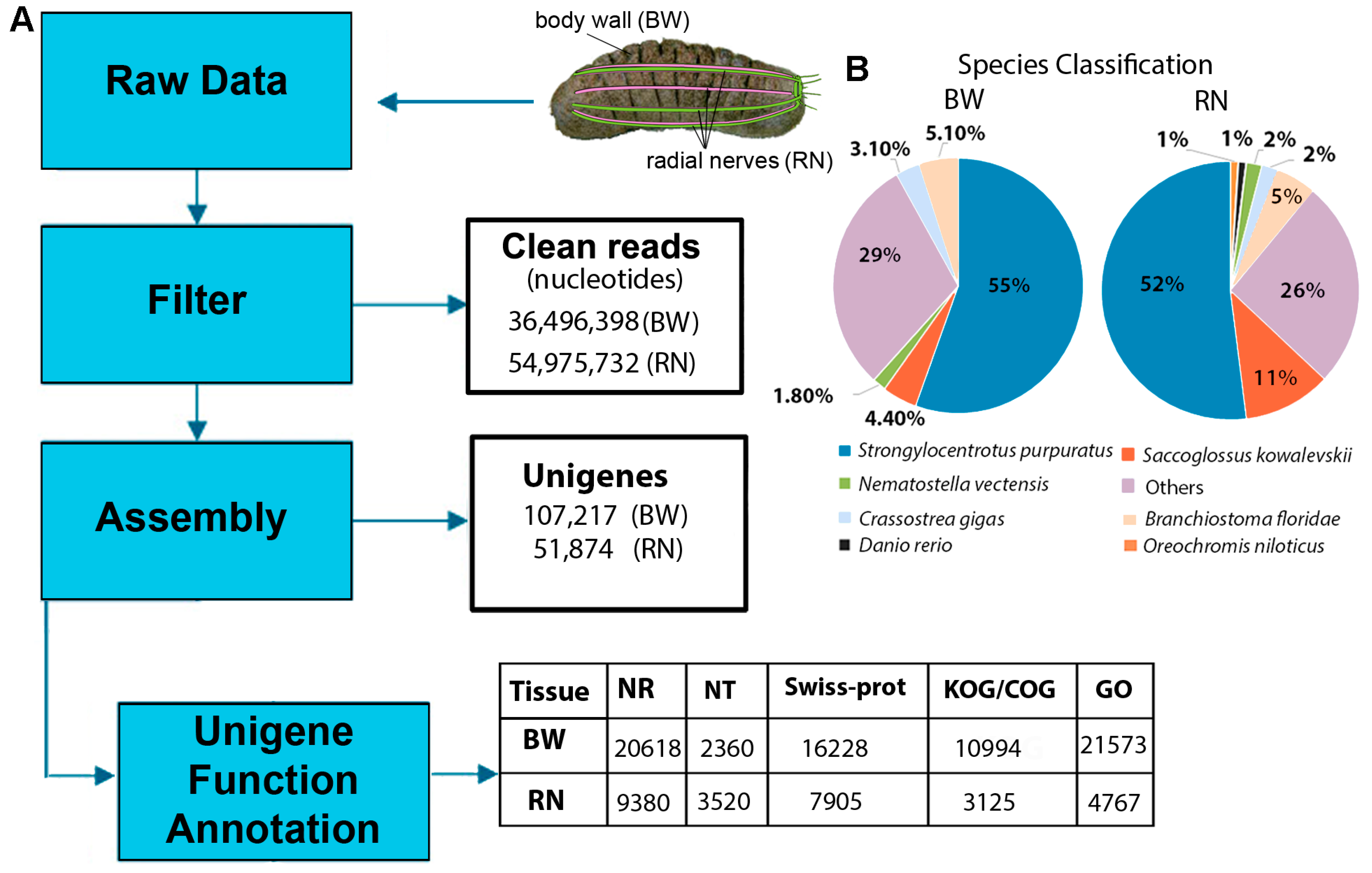
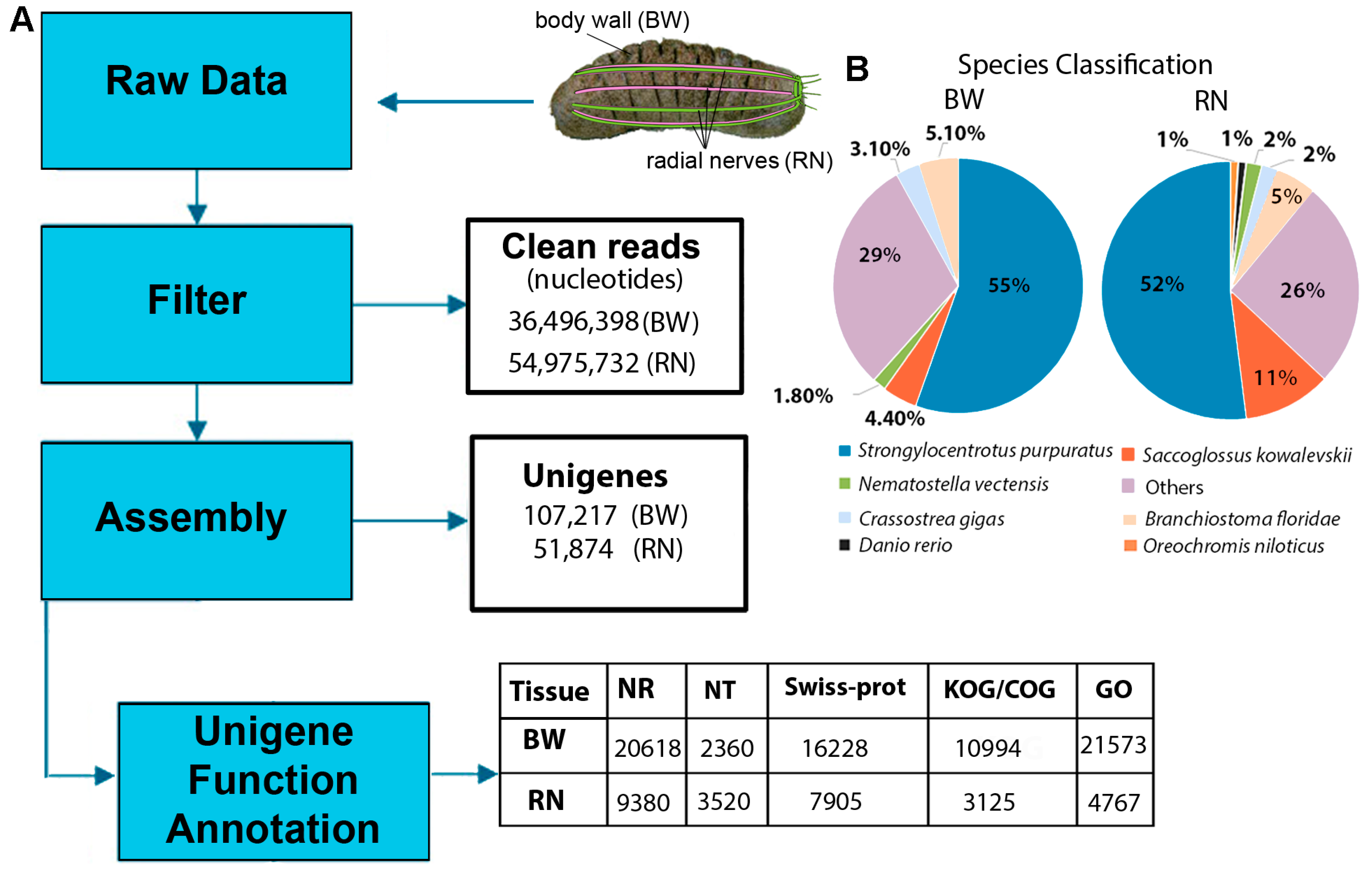
2.1. Body Wall and Radial Nerve Transcriptome Analysis
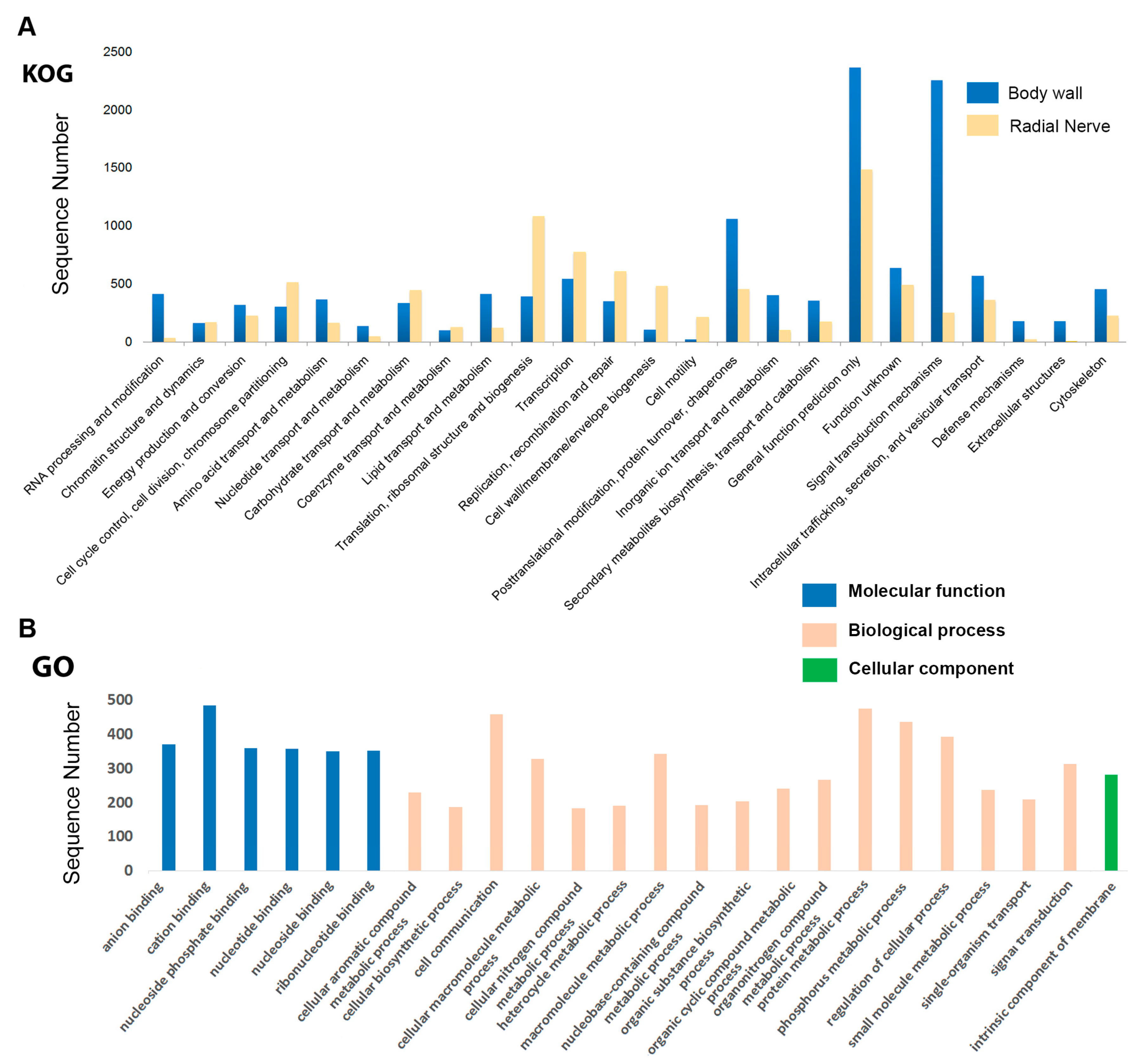
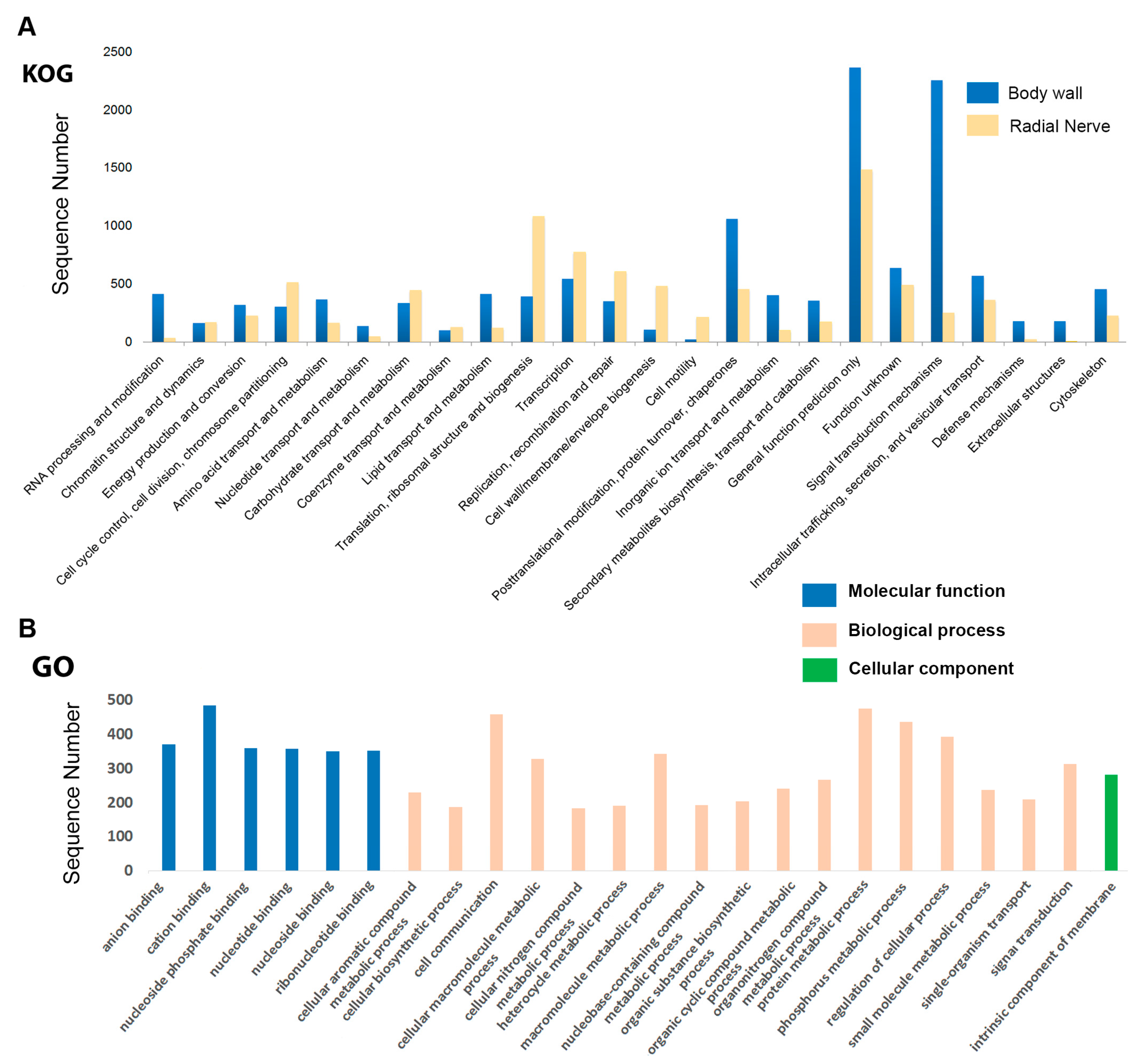
2.2. Functional Annotation of H. scabra Body Wall and Radial Nerve Transcripts
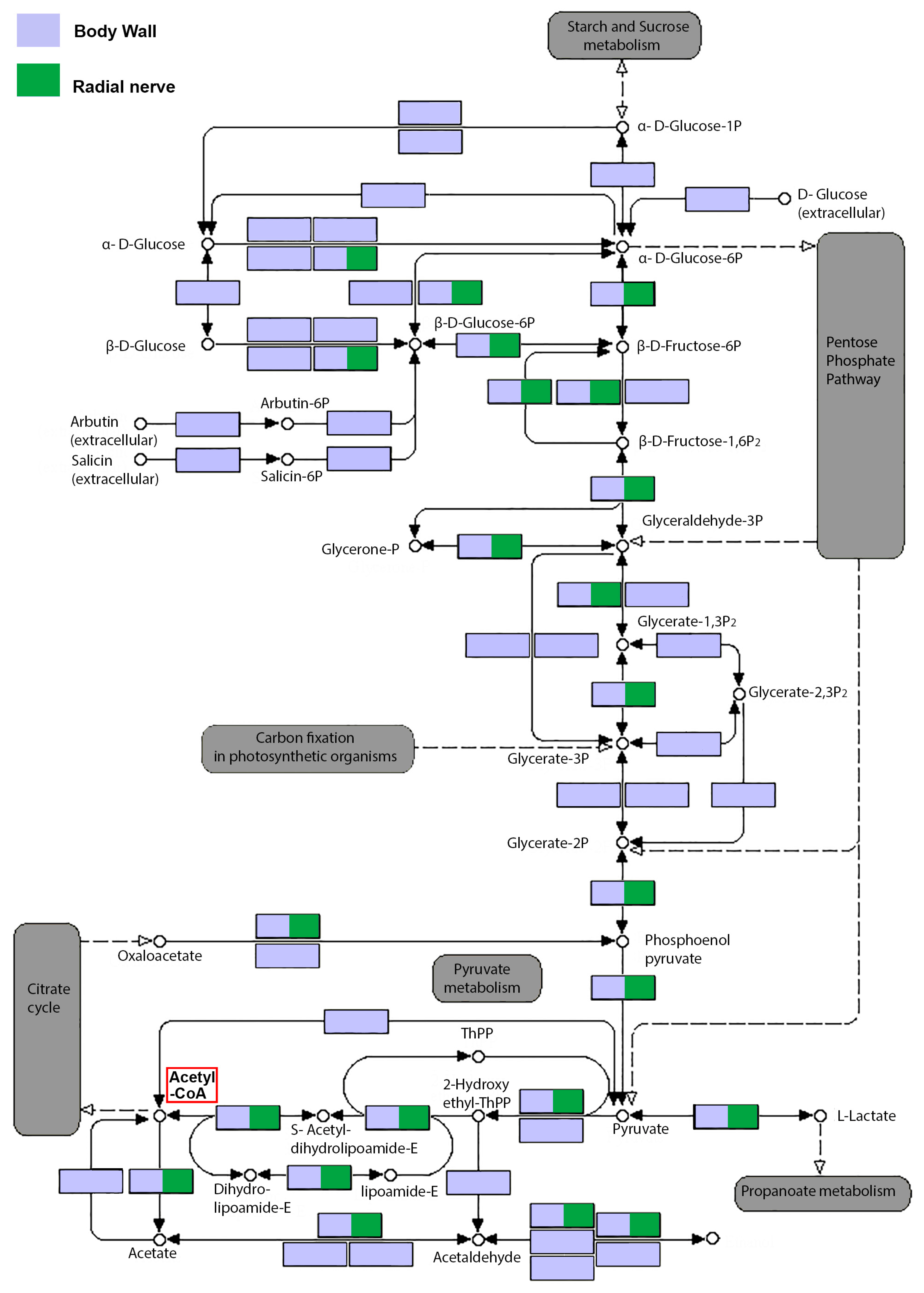
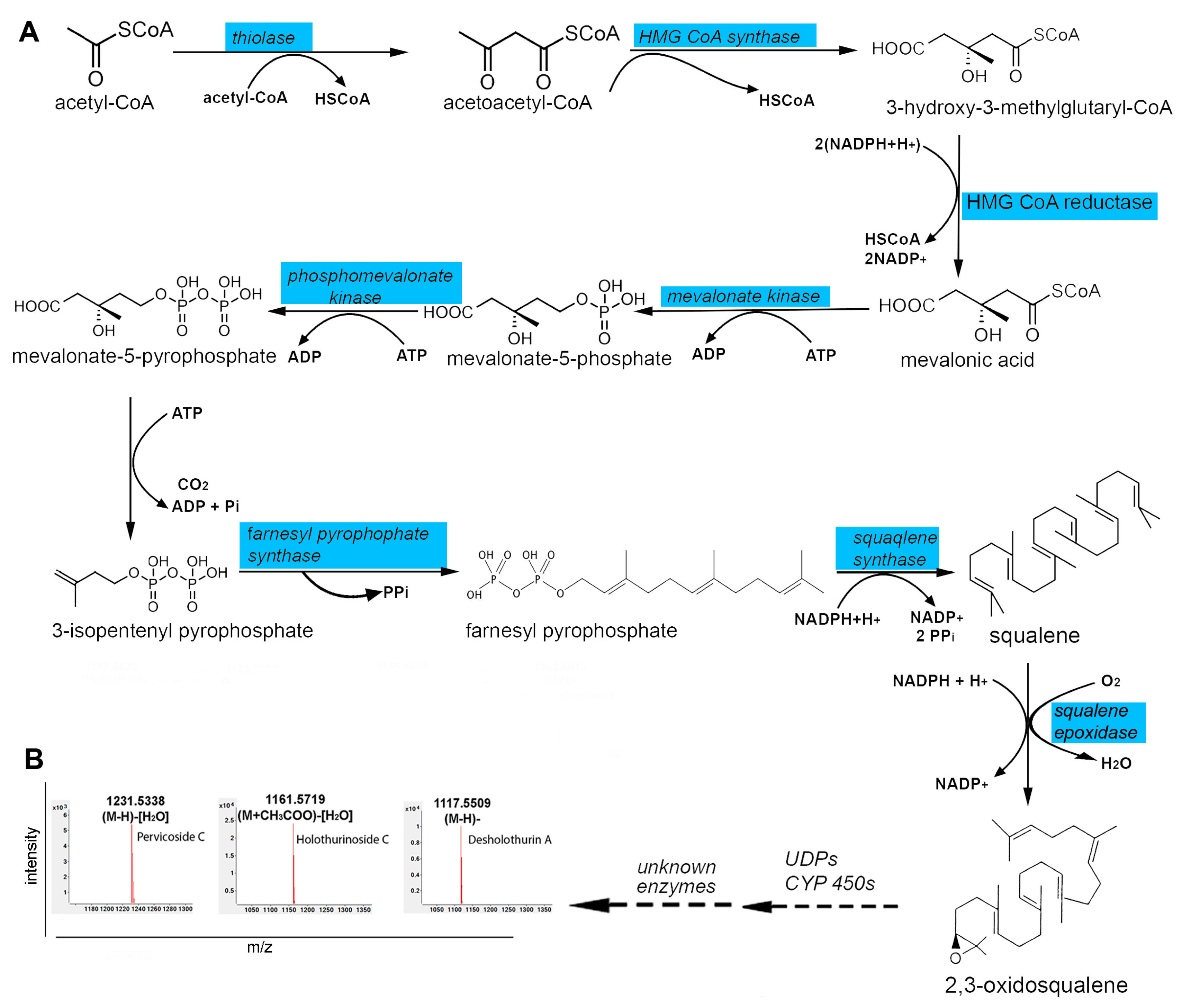
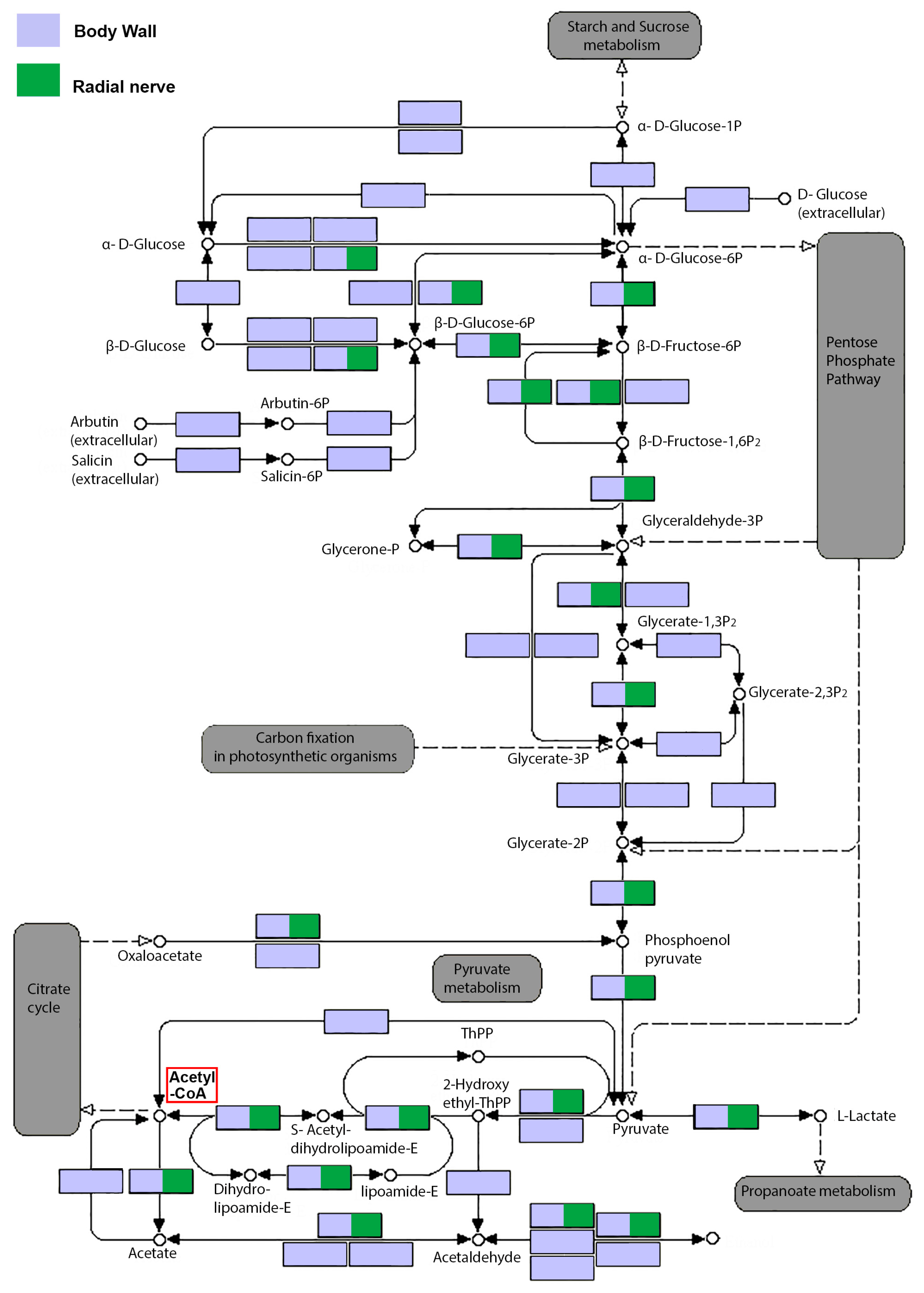
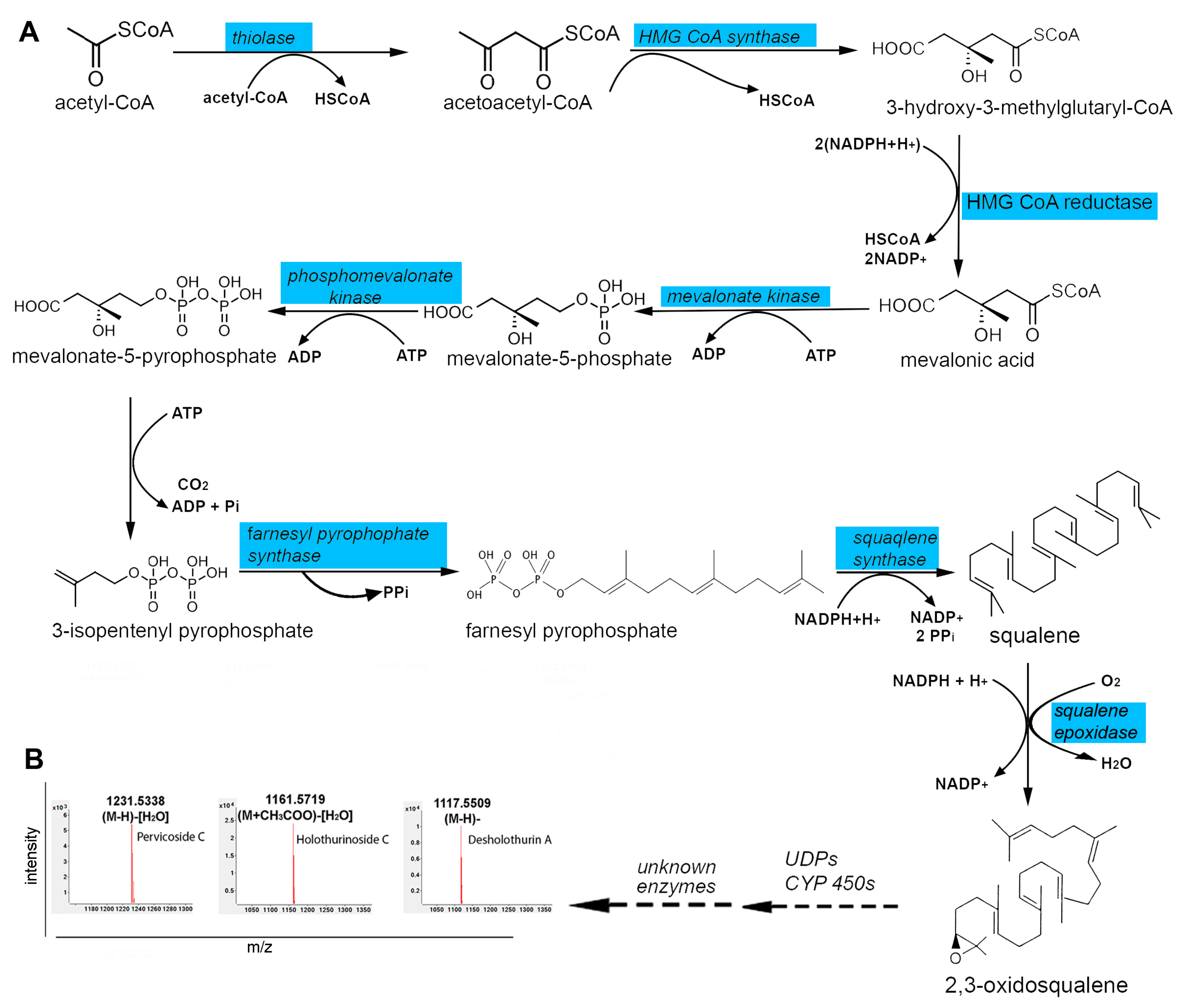
2.3. Secondary Metabolite Biosynthesis Pathways in the Body Wall
3. Materials and Methods
3.1. Animals and Tissue Collection
3.2. Total RNA Isolation, Transcriptome Analysis and Pathway Analysis
3.3. Liquid Chromatography-Mass Spectrometry-Based Metabolomic (LC-MS) Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, Z.; Chen, H.; Wang, W.; Jia, B.; Yang, T.; Zhao, Z.; Ding, J.; Xiao, X. Differentiation of dried sea cucumber products from different geographical areas by surface desorption atmospheric pressure chemical ionization mass spectrometry. J. Agric. Food Chem. 2009, 57, 9356–9364. [Google Scholar] [CrossRef] [PubMed]
- Conand, C. Harvest and trade: Utilization of sea cucumbers; sea cucumber fisheries; current international trade; illegal, unreported and unregulated trade; bycatch; socio-economic characteristics of the trade in sea cucumbers. In The Proceedings of the Technical Workshop on the Conservation of Sea Cucumbers in the Families Holothuridae and Stichopodidae; NOAA Technical Memorandum: Seattle, WA, USA, 2005; pp. 47–69. [Google Scholar]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea cucumbers metabolites as potent anti-cancer agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kumiko, K.; Yasuzawa, T.; Matsui, M.; Suzuki, S.; Kitagawa, I. Marine natural products. XXVII. Distribution of lanostane-type triterpene oligoglycosides in ten kinds of Okinawan sea cucumbers. Chem. Pharm. Bull. 1991, 39, 2282–2287. [Google Scholar] [CrossRef]
- Elyakov, G.; Kuznetsova, T.; Stonik, V.; Levin, V.; Albores, R. Glycosides of marine invertebrates—IV. A comparative study of the glycosides from Cuban sublittoral holothurians. Comp. Biochem. Physiol. Part B Comp. Biochem. 1975, 52, 413–417. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [PubMed]
- Kerr, R.G.; Chen, Z. In vivo and in vitro biosynthesis of saponins in sea cucumbers. J. Nat. Prod. 1995, 58, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Dewi, A.S.; Tarman, K.; Uria, A.R. Marine natural products: Prospects and impacts on the sustainable development in Indonesia. In Proceedings of the Indonesian Student Scientific Meeting (ISSM) on Sustainable Development in Indonesia: An Interdisciplinary Approach, Delft, The Netherlands, 13–15 May 2008; pp. 13–15. [Google Scholar]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev. 2001, 70, 673–715. [Google Scholar] [CrossRef]
- Skropeta, D. Deep-sea natural products. Nat. Prod. Rep. 2008, 25, 1131–1166. [Google Scholar] [CrossRef] [PubMed]
- Vincken, J.-P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Negi, J.; Negi, P.; Pant, G.; Rawat, M.; Negi, S. Naturally occurring saponins: Chemistry and biology. J. Poisonous Med. Plant Res. 2013, 1, 6–11. [Google Scholar]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity ofsaponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Nah, S.-Y. Ginseng ginsenoside pharmacology in the nervous system: Involvement in the regulation of ion channels and receptors. Front. Physiol. 2014, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed]
- Skene, C.D.; Sutton, P. Saponin-adjuvanted particulate vaccines for clinical use. Methods 2006, 40, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Maier, M.S. Biological activities of sulfated glycosides from echinoderms. Stud. Nat. Prod. Chem. 2008, 35, 311–354. [Google Scholar]
- Caulier, G.; Van Dyck, S.; Gerbaux, P.; Eeckhaut, I.; Flammang, P. Review of saponin diversity in sea cucumbers belonging to the family Holothuriidae. Secr. Pac. Community Beche-De-Mer Inf. Bull. 2011, 31, 48–54. [Google Scholar]
- Bose, U.; Centurion, E.; Hodson, M.; Shaw, P.; Storey, K.; Cummins, S. Global metabolite analysis of the land snail Theba pisana hemolymph during active and aestivated states. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 19, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Kruangkum, T.; Wang, T.; Zhao, M.; Ventura, T.; Mitu, S.A.; Hodson, M.P.; Shaw, P.N.; Sobhon, P.; Cummins, S.F. Biomolecular changes that occur in the antennal gland of the giant freshwater prawn (Machrobrachium rosenbergii). PLoS ONE 2017, 12, e0177064. [Google Scholar] [CrossRef] [PubMed]
- Caulier, G.; Mezali, K.; Soualili, D.L.; Decroo, C.; Demeyer, M.; Eeckhaut, I.; Gerbaux, P.; Flammang, P. Chemical characterization of saponins contained in the body wall and the cuvierian tubules of the sea cucumber Holothuria (Platyperona) sanctori (Delle Chiaje, 1823). Biochem. Syst. Ecol. 2016, 68, 119–127. [Google Scholar] [CrossRef]
- Han, H.; Yi, Y.; Xu, Q.; La, M.; Zhang, H. Two new cytotoxic triterpene glycosides from the sea cucumber Holothuria scabra. Planta Med. 2009, 75, 1608–1612. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P. Defensive roles for secondary metabolites from marine sponges and sponge-feeding nudibranchs. Toxicon 1994, 32, 639–655. [Google Scholar] [CrossRef]
- Sun, L.; Chen, M.; Yang, H.; Wang, T.; Liu, B.; Shu, C.; Gardiner, D.M. Large scale gene expression profiling during intestine and body wall regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, C.; Zhu, L.; Su, X.; Li, Y.; Jin, C.; Li, T. De novo assembly of the sea cucumber Apostichopus japonicus hemocytes transcriptome to identify miRNA targets associated with skin ulceration syndrome. PLoS ONE 2013, 8, e73506. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Bao, Z.; Hou, R.; Wang, S.; Su, H.; Yan, J.; Tian, M.; Li, Y.; Wei, W.; Lu, W. Transcriptome sequencing and characterization for the sea cucumber Apostichopus japonicus (Selenka, 1867). PLoS ONE 2012, 7, e33311. [Google Scholar] [CrossRef] [PubMed]
- San Miguel-Ruiz, J.E.; Maldonado-Soto, A.R.; Garcia-Arraras, J.E. Regeneration of the radial nerve cord in the sea cucumber Holothuria glaberrima. BioMed Cent. Dev. Biol. 2009, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szczesniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Present Status and Prospects of Sea Cucumber Industry in China; Food and Agricultural Organization Fisheries and Technical Paper; FAO: Rome, Italy, 2005; pp. 25–38. [Google Scholar]
- Chen, J. Overview of sea cucumber farming and sea ranching practices in China. Secr. Pac. Community Beche-De-Mer Inf. Bull. 2003, 18, 18–23. [Google Scholar]
- Mohammadizadeh, F.; Ehsanpor, M.; Afkhami, M.; Mokhlesi, A.; Khazaali, A.; Montazeri, S. Antibacterial, antifungal and cytotoxic effects of a sea cucumber Holothuria leucospilota, from the north coast of the Persian Gulf. J. Mar. Biol. Assoc. UK 2013, 93, 1401–1405. [Google Scholar] [CrossRef]
- Omran, N.E.-S.E.-S. Nutritional value of some Egyptian sea cucumbers. Afr. J. Biotechnol. 2013, 12, 5466–5472. [Google Scholar]
- Sroyraya, M.; Hanna, P.J.; Siangcham, T.; Tinikul, R.; Jattujan, P.; Poomtong, T.; Sobhon, P. Nutritional components of the sea cucumber Holothuria scabra. Funct. Foods Health Dis. 2017, 7, 168–181. [Google Scholar]
- Adeniyi, S.; Orjiekwe, C.; Ehiagbonare, J.; Josiah, S. Nutritional composition of three different fishes (Clarias gariepinus, Malapterurus electricus and Tilapia guineensis). Pak. J. Nutr. 2012, 11, 793. [Google Scholar] [CrossRef]
- Chukwu, O. Influences of drying methods on nutritional properties of tilapia fish (Oreochromis nilotieus). World J. Agric. Sci. 2009, 5, 256–258. [Google Scholar]
- Waagbo, R.; Sandnes, K.; Torrissen, O.J.; Sandvin, A.; Lie, O. Chemical and sensory evaluation of fillets from Atlantic salmon (Salmo salar) fed three levels of n-3 polyunsaturated fatty acids at two levels of vitamin E. Food Chem. 1993, 46, 361–366. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, X.; Liu, J.; Storey, K.B. High-throughput sequencing reveals differential expression of miRNAs in intestine from sea cucumber during aestivation. PLoS ONE 2013, 8, e76120. [Google Scholar] [CrossRef] [PubMed]
- Binyon, J. Physiology of Echinoderms: International Series of Monographs in Pure and Applied Biology Zoology; Pergamon: New York, NY, USA, 2013; Volume 49. [Google Scholar]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Each Organ Has a Unique Metabolic Profile Available Biochemistry, 5th ed.; W H Freeman Co. Ltd.: New York, NY, USA, 2002; pp. 851–854. [Google Scholar]
- Caulier, G.; Flammang, P.; Rakotorisoa, P.; Gerbaux, P.; Demeyer, M.; Eeckhaut, I. Preservation of the bioactive saponins of Holothuria scabra through the processing of trepang. Cah. Biol. Mar. 2013, 54, 685–690. [Google Scholar]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Elucidation of molecular diversity and body distribution of saponins in the sea cucumber Holothuria forskali (Echinodermata) by mass spectrometry. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Kalinin, V.I.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Stonik, V.A. Triterpene glycosides from sea cucucmbers (holothurioidea, echinodermata). Biological activities and functions. Stud. Nat. Prod. Chem. 2008, 35, 135–196. [Google Scholar]
- Van Dyck, S.; Caulier, G.; Todesco, M.; Gerbaux, P.; Fournier, I.; Wisztorski, M.; Flammang, P. The triterpene glycosides of Holothuria forskali: Usefulness and efficiency as a chemical defense mechanism against predatory fish. J. Exp. Biol. 2011, 214, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Q.; Li, L.; Yi, Y.H.; Yuan, W.H.; Liu, B.S.; Weng, Y.Y.; Zhang, S.L.; Sun, P.; Wang, Z.L. Two new cytotoxic nonsulfated pentasaccharide holostane (= 20-hydroxylanostan-18-oic acid γ-lactone) glycosides from the sea cucumber Holothuria grisea. Helv. Chim. Acta 2008, 91, 1453–1460. [Google Scholar] [CrossRef]
- Dang, N.H.; Van Thanh, N.; Van Kiem, P.; Huong, L.M.; Van Minh, C.; Kim, Y.H. Two new triterpene glycosides from the Vietnamese sea cucumber Holothuria scabra. Arch. Pharm. Res. 2007, 30, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. Metabolite profiling of polar steroid constituents in the far eastern starfish Aphelasterias japonica using LC–ESI MS/MS. Metabolomics 2014, 10, 1153–3882. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. LC-ESI MS/MS profiling of polar steroid metabolites of the far eastern starfish Patiria (= Asterina) pectinifera. Metabolomics 2016, 12, 21. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Dmitrenok, P.S.; Stonik, V.A. LC–MS-based metabolome analysis on steroid metabolites from the starfish Patiria (= Asterina) pectinifera in conditions of active feeding and stresses. Metabolomics 2016, 12, 106. [Google Scholar] [CrossRef]
- Liu, B.S.; Yi, Y.H.; Li, L.; Sun, P.; Han, H.; Sun, G.Q.; Wang, X.H.; Wang, Z.L. Argusides D and E, two new cytotoxic triterpene glycosides from the sea cucumber Bohadschia argus Jaeger. Chem. Biodivers. 2008, 5, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, I.; Kobayashi, M.; Hori, M.; Kyogoku, Y. Structures of four new triterpenoidal oligoglycosides, bivittoside A, B, C, and D, from the sea cucumber Bohadschia bivittata Mitsukuri. Chem. Pharm. Bull. 1981, 29, 282–285. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Yurchenko, E.A.; Dolmatov, I.Y.; Savchenko, A.M.; Kalinin, V.I. Triterpene glycosides from the sea cucumber Cladolabes schmeltzii. II. Structure and biological action of cladolosides A1-A6. Nat. Prod. Commun. 2014, 9, 1421–1428. [Google Scholar] [PubMed]
- Elbandy, M.; Rho, J.R.; Afifi, R. Analysis of saponins as bioactive zoochemicals from the marine functional food sea cucumber Bohadschia cousteaui. Eur. Food Res. Technol 2014, 238, 937–955. [Google Scholar] [CrossRef]
- Bahrami, Y.; Zhang, W.; Franco, C. Discovery of novel saponins from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 2633–2667. [Google Scholar] [CrossRef] [PubMed]
- Muniain, C.; Centurion, R.; Careaga, V.; Maier, M. Chemical ecology and bioactivity of triterpene glycosides from the sea cucumber Psolus patagonicus (Dendrochirotida: Psolidae). J. Mar. Biol. Assoc. UK 2008, 88, 817–823. [Google Scholar] [CrossRef]
- Hu, S.; Xu, L.; Shi, D.; Wang, J.; Wang, Y.; Lou, Q.; Xue, C. Eicosapentaenoic acid-enriched phosphatidylcholine isolated from Cucumaria frondosa exhibits anti-hyperglycemic effects via activating phosphoinositide 3-kinase/protein kinase B signal pathway. J. Biosci. Bioeng. 2014, 117, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, S.; Okano, K.; Muragaki, H. Structure of glycoside B2, a steroidal saponin in the ovary of the starfish, Asterias amurensis. Tetrahedron Lett. 1979, 20, 1769–1772. [Google Scholar] [CrossRef]
- Kitagawa, I.; Sugawara, T.; Yosioka, I.; Kuriyama, K. Saponin and Sapogenol. XIV. Antifugal glycosides from the sea cucumber Stichopus japonicus Selenka.(1). Structure of Stichopogenin A4, the genuine aglycone of Holotoxin A. Chem. Pharm. Bull. 1976, 24, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Riguera, R. Lefevreiosides: Four novel triterpenoid glycosides from the sea cucumber Cucumaria lefevrei. J. Chem. Res. Synop. 1989, 11, 342–343. [Google Scholar]
- Zhang, J.-J.; Zhu, Q.-K.; Wu, J.; Zhang, H.-W. New cytotoxic triterpene glycoside from the East China Sea cucumber Holothuria nobilis. Nat. Prod. Commun. 2015, 10, 247–248. [Google Scholar] [PubMed]
- Zhang, S.-Y.; Yi, Y.-H.; Tang, H.-F. Bioactive triterpene glycosides from the sea cucumber Holothuria fuscocinerea. J. Nat. Prod. 2006, 69, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-L.; Li, L.; Yi, Y.-H.; Sun, P. Philinopsides E and F, two new sulfated triterpene glycosides from the sea cucumber Pentacta quadrangularis. Nat. Prod. Res. 2006, 20, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Hewavitharana, A.K.; Ng, Y.K.; Shaw, P.N.; Fuerst, J.A.; Hodson, M.P. LC-MS-Based metabolomics study of marine bacterial secondary metabolite and antibiotic production in Salinispora arenicola. Mar. Drugs 2015, 13, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Hewavitharana, A.K.; Vidgen, M.E.; Ng, Y.K.; Shaw, P.N.; Fuerst, J.A.; Hodson, M.P. Discovering the recondite secondary metabolome spectrum of Salinispora species: A study of inter-species diversity. PLoS ONE 2014, 9, e91488. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sea Cucumber Metabolites | m/z ± ppm | Relative Abundance in Eluate * | Previously Reported Species | Source | References |
|---|---|---|---|---|---|
| Holothurinoside Z | 1101.5488 ± 0.30 | 322,380 | Holothuria forskali | Body wall and cuvierian tubules | [47] |
| Arguside D | 1157.5522 ± 2.1 | 161,629 | Bohadschia argus Jaeger | Body wall, cuvierian tubules | [50] |
| Bivittoside A | 1133.5854±0.02 | 318,206 | Bohadschia bivittata | Conditioned water and cuvierian tubules | [51] |
| Cladoloside A2 | 1155.5696 ± 0.25 | 179,083 | Cladolabes schmeltzii | Whole animal | [52] |
| Cousteside E | 1263.6013 ± 0.39 | 254,678 | Bohadschia cousteaui | Body wall | [53] |
| Cousteside I | 1265.6275 ± 1.93 | 279,592 | Bohadschia cousteaui | Body wall | [53] |
| Desholothurin A | 1117.5509 ± 0.31 | 334,366 | H. lessoni, H. forskali, H. nobilis, A. agassizi, B. argus, B. cousteaui, H. leucospilota, P. graeffei, H. scabra | Body wall | [4,19,53,54] |
| Holothurinoside C | 1161.5719 ± 1.58 | 273,822 | H. lessoni, H. forskali, A. agassizi, H. scabra | Body wall and cuvierian tubules | [4,19,54] |
| Ds-patagonicoside A | 1103.5637 ± 0.86 | 170,738 | Psolus patagonicus | Whole animal | [55] |
| Eicosapentanoic acid | 301.2189 ± 1.92 | 76,358 | Cucumaria frondosa | Body wall | [56] |
| Glycoside B2 | 1225.5305 ± 1.3 | 276,012 | Asterias amurensis | Ovary | [57] |
| Holotoxin A4 | 469.3312 ± 0.11 | 379,504 | Stichopus japonicas | Whole animal | [58] |
| Lefevreioside D | 1199.5249 ± 0.15 | 352,784 | Cucumaria (Aslia) lefevrei | Whole animal | [59] |
| Nobiliside E | 1198.5034 ± 1.52 | 531,373 | Holothuria nobilis | Whole animal | [60] |
| Pervicoside C | 1231.5338 ± 5.04 | 77,368 | Holothuria fuscocinerea, H scabra | Whole animal | [4,61] |
| Philinopside E | 1155.4523 ± 1.23 | 207,304 | Pentacta quadrangularis | Body wall | [62] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitu, S.A.; Bose, U.; Suwansa-ard, S.; Turner, L.H.; Zhao, M.; Elizur, A.; Ogbourne, S.M.; Shaw, P.N.; Cummins, S.F. Evidence for a Saponin Biosynthesis Pathway in the Body Wall of the Commercially Significant Sea Cucumber Holothuria scabra. Mar. Drugs 2017, 15, 349. https://doi.org/10.3390/md15110349
Mitu SA, Bose U, Suwansa-ard S, Turner LH, Zhao M, Elizur A, Ogbourne SM, Shaw PN, Cummins SF. Evidence for a Saponin Biosynthesis Pathway in the Body Wall of the Commercially Significant Sea Cucumber Holothuria scabra. Marine Drugs. 2017; 15(11):349. https://doi.org/10.3390/md15110349
Chicago/Turabian StyleMitu, Shahida Akter, Utpal Bose, Saowaros Suwansa-ard, Luke H. Turner, Min Zhao, Abigail Elizur, Steven M. Ogbourne, Paul Nicholas Shaw, and Scott F. Cummins. 2017. "Evidence for a Saponin Biosynthesis Pathway in the Body Wall of the Commercially Significant Sea Cucumber Holothuria scabra" Marine Drugs 15, no. 11: 349. https://doi.org/10.3390/md15110349
APA StyleMitu, S. A., Bose, U., Suwansa-ard, S., Turner, L. H., Zhao, M., Elizur, A., Ogbourne, S. M., Shaw, P. N., & Cummins, S. F. (2017). Evidence for a Saponin Biosynthesis Pathway in the Body Wall of the Commercially Significant Sea Cucumber Holothuria scabra. Marine Drugs, 15(11), 349. https://doi.org/10.3390/md15110349