Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients

 ,
,
Abstract
1. Introduction
2. Mycosporine-Like Amino Acids
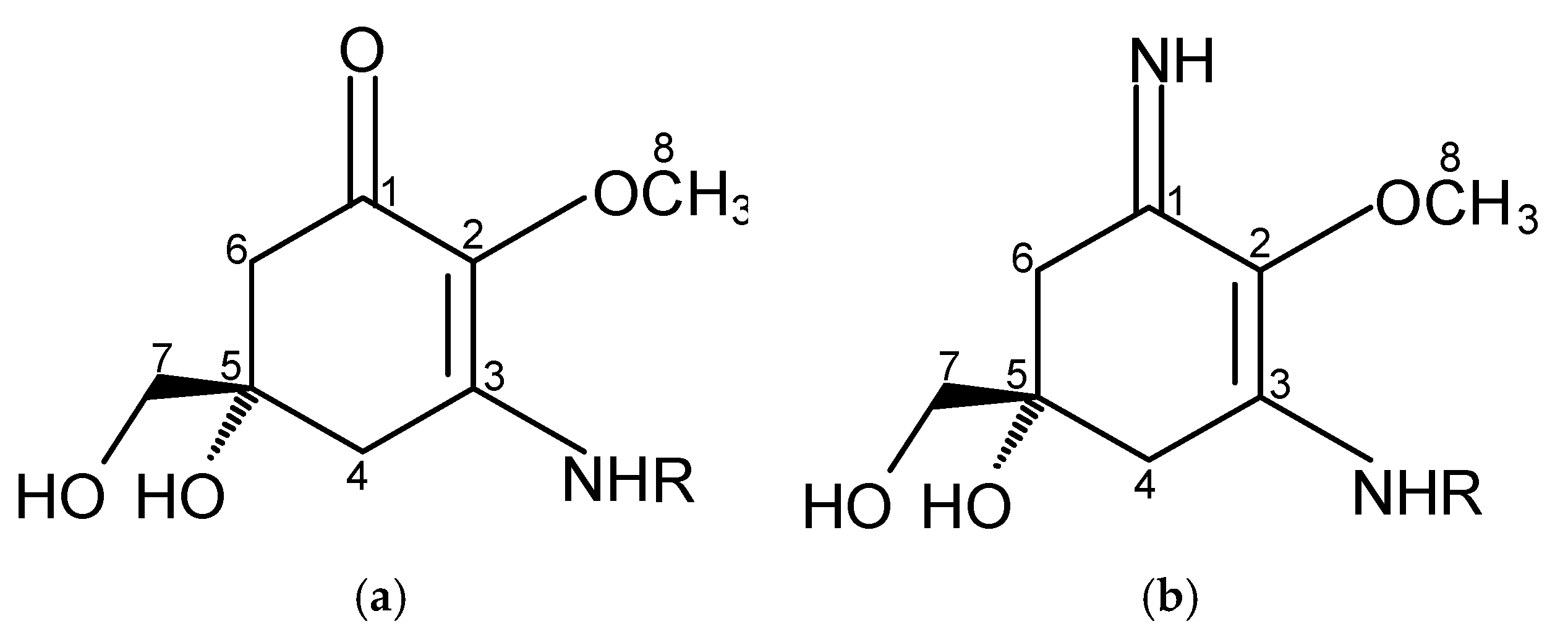
2.1. Structure and Physicochemical Properties
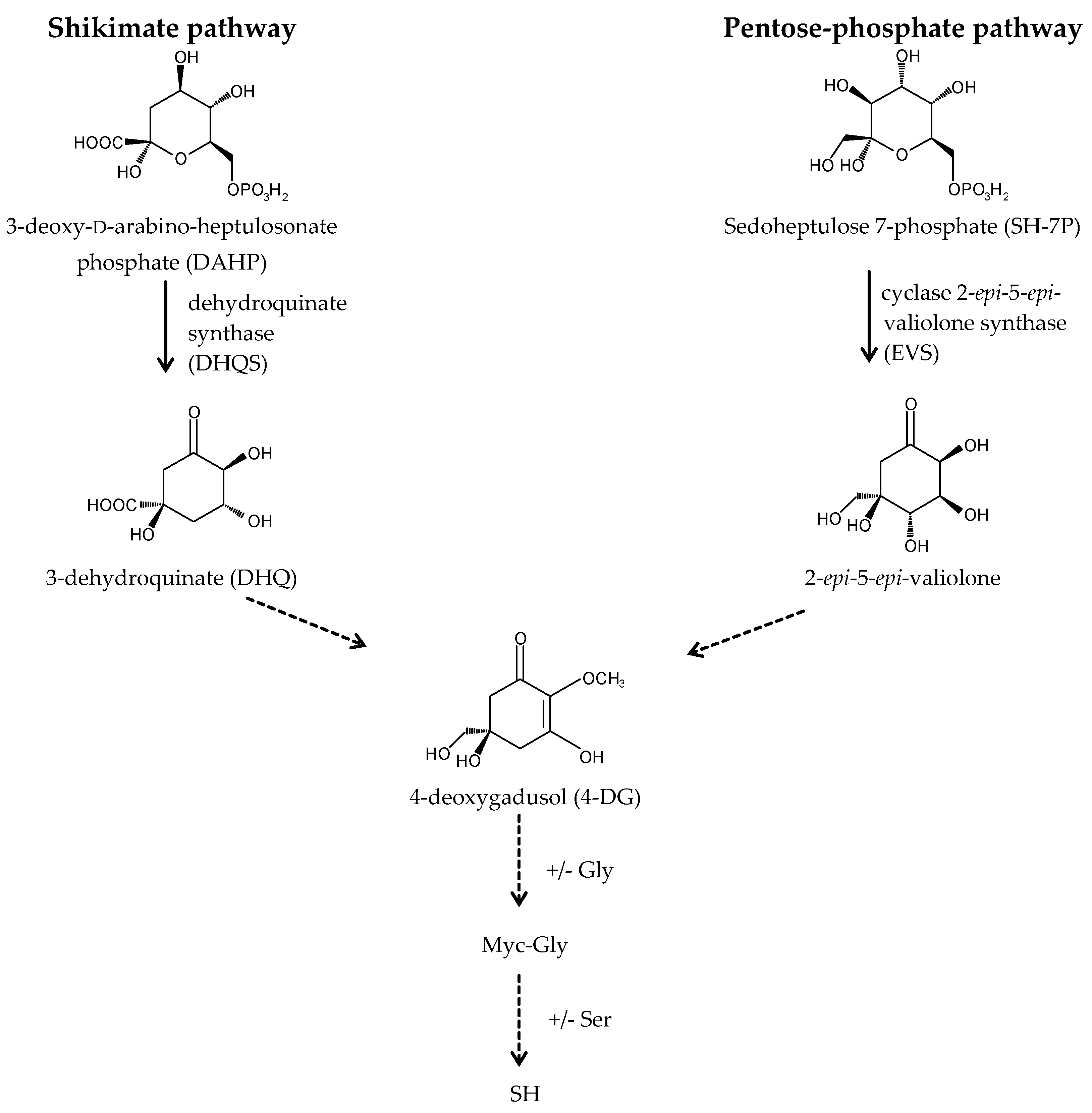
2.2. Biosynthetic Pathways and Their Regulation
2.3. Occurrence and Distribution in the Environment
2.4. Functions
2.4.1. Photoprotection
2.4.2. Biological Antioxidant Molecules
2.4.3. Protection against Abiotic Stress Factors
2.4.4. Nitrogen Storage
2.4.5. Other Functions
3. MAAs: A Commercial Approach
3.1. Harmful Effects of UV on the Human Body
3.2. Internal and External Skin Protection Against UV
3.3. Potential of MAAs in Skin Protection
3.3.1. MAAs as Sunscreens
3.3.2. MAAs as Anti-Cancer Agents
3.3.3. MAAs as Anti-Photoaging Agents
3.3.4. MAAs as Wound Healing Agents
3.3.5. MAAs as Functional Components of UV-Protective Materials
3.3.6. MAAs Commercial Applications
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Karentz, D. Chemical defenses of marine organisms against solar radiation exposure: UV-absorbing mycosporine-like amino acids and scytonemin. In Marine Chemical Ecology; McClintock, J.B., Baker, J., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 2001; pp. 481–520. ISBN 978-0-8493-9064-7. [Google Scholar]
- Afaq, F.; Mukhtar, H. Botanical antioxidants in the prevention of photocarcinogenesis and photoaging. Exp. Dermatol. 2006, 15, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Mckenzie, R.L.; Björn, L.O.; Bais, A.; Ilyasd, M. Changes in biologically active ultraviolet radiation reaching the Earth’s surface. Photochem. Photobiol. Sci. 2003, 2, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.V.; Downs, N. Cloud cover and horizontal plane eye damaging solar UV exposures. Int. J. Biometeorol. 2004, 49, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.A. The diffuse component of erythemal ultraviolet radiation. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2015, 14, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Moseley, H. Non-Ionizing Radiation: Microwaves, Ultraviolet and Laser Radiation; JW Arrowsmith Ltd.: Bristol, UK, 1988; ISBN 0852741669. [Google Scholar]
- Björn, L.O. Stratospheric ozone, ultraviolet radiation, and cryptogams. Biol. Conserv. 2007, 135, 326–333. [Google Scholar] [CrossRef]
- Saika, K.; Sobue, T. Cancer statistics in the world. Gan To Kagaku Ryoho 2013, 40, 2475–2480. [Google Scholar] [PubMed]
- Wang, S.Q.; Setlow, R.; Berwick, M.; Polsky, D.; Marghoob, A.A.; Kopf, A.W.; Bart, R.S. Ultraviolet A and melanoma: A review. J. Am. Acad. Dermatol. 2001, 44, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Eisemann, N.; Waldmann, A.; Geller, A.C.; Weinstock, M.A.; Volkmer, B.; Greinert, R.; Breitbart, E.W.; Katalinic, A. Non-melanoma skin cancer incidence and impact of skin cancer screening on incidence. J. Investig. Dermatol. 2014, 134, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Temina, M.; Tolstikov, A.G.; Dembitsky, V.M. Natural microbial UV radiation filters—Mycosporine-like amino acids. Folia Microbiol. Praha 2004, 49, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, D. Computational exploration of natural sunscreens. Phys. Chem. Chem. Phys. 2011, 13, 5584–5586. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine microbial-derived molecules and their potential use in cosmeceutical and cosmetic products. Mar. Drugs 2017, 15, E118. [Google Scholar] [CrossRef] [PubMed]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef] [PubMed]
- Bandaranayake, W.M. Mycosporines: Are they nature’s sunscreens? Nat. Prod. Rep. 1998, 15, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Richa, R.; Kumari, S. Biotechnological potential of mycosporine-like amino acids and phycobiliproteins of cyanobacterial origin. Biotechnol. Bioinf. Bioeng. 2011, 1, 159–171. [Google Scholar]
- Eom, S.H.; Kim, S.K. Cosmeceutical applications from marine organisms. In Cosmeceuticals and Cosmetic Practice; Farris, P.K., Ed.; John Wiley Sons, Ltd.: Chichester, UK, 2013; pp. 200–208. ISBN 9781118384800. [Google Scholar]
- Sinha, R.P. Biomedical applications of mycosporine-like amino acids. In Marine Microbiology: Bioactive Compounds and Biotechnological Applications; Kim, S.K., Ed.; Wiley-VCH Verlag GmbH Co. KGaA: Weinheim, Germany, 2013; pp. 509–534. ISBN 9783527333271. [Google Scholar]
- Bhatia, S.; Sharma, K.; Sharma, A.; Garg, A.; Kumar, S.; Purohit, A. Mycosporine and mycosporine-like amino acids: A paramount tool against ultra violet irradiation. Pharmacogn. Rev. 2011, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Gröniger, A.; Sinha, R.P.; Klisch, M.; Häder, D.P. Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae—A database. J. Photochem. Photobiol. B Biol. 2000, 58, 115–122. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, acumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Singh, S.P.; Häder, D.P. Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J. Photochem. Photobiol. B Biol. 2007, 89, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, W.C.; Shick, J.M. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: A biochemical and environmental perspective. J. Phycol. 1998, 34, 418–430. [Google Scholar] [CrossRef]
- Shibata, K. Pigments and a UV-absorbing substance in corals and a blue-green alga living in the Great Barrier Reef. Plant Cell Physiol. 1969, 10, 325–335. [Google Scholar] [CrossRef]
- Nakamura, H.; Kobayashi, J.; Hirata, Y. Separation of mycosporine-like amino acids in marine organisms using reversed-phase high-performance liquid chromatography. J. Chromatogr. 1982, 250, 113–118. [Google Scholar] [CrossRef]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like amino acids (MAAs): Chemical structure, biosynthesis and significance as UV-absorbing/screening compounds. Indian J. Exp. Biol. 2008, 46, 7–17. [Google Scholar] [PubMed]
- Favre-Bonvin, J.; Arpin, N.; Brevard, C. Structure de la mycosporine (P 310). Can. J. Chem. 1976, 54, 1105–1113. [Google Scholar] [CrossRef]
- Torres, A.; Hochberg, M.; Pergament, I.; Smoum, R.; Niddam, V.; Dembitsky, V.M.; Temina, M.; Dor, I.; Lev, O.; Srebnik, M.; et al. A new UV-B absorbing mycosporine with photo protective activity from the lichenized ascomycete Collema cristatum. Eur. J. Biochem. 2004, 271, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-like amino acids and their derivatives as natural antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef] [PubMed]
- Klisch, M.; Richter, P.; Puchta, R.; Häder, D.P.; Bauer, W. The stereostructure of porphyra-334: An experimental and calculational NMR investigation. Evidence for an efficient ‘proton sponge’. Helv. Chim. Acta 2007, 90, 488–511. [Google Scholar] [CrossRef]
- Ishihara, K.; Watanabe, R.; Uchida, H.; Suzuki, T.; Yamashita, M.; Takenaka, H.; Nazifi, E.; Matsugo, S.; Yamaba, M.; Sakamoto, T. Novel glycosylated mycosporine-like amino acid, 13-O-(β-galactosyl)-porphyra-334, from the edible cyanobacterium Nostoc sphaericum-protective activity on human keratinocytes from UV light. J. Photochem. Photobiol. B Biol. 2017, 172, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Carignan, M.O.; Cardozo, K.H.M.; Oliveira-Silva, D.; Colepicolo, P.; Carreto, J.I. Palythine-threonine, a major novel mycosporine-like amino acid (MAA) isolated from the hermatypic coral Pocillopora capitata. J. Photochem. Photobiol. B Biol. 2009, 94, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Wu Won, J.J.; Chalker, B.E.; Rideout, J.A. Two new UV-absorbing compounds from Stylophora pistillata: Sulphate esters of mycosporine-like amino acids. Tetrahedron Lett. 1997, 38, 2525–2526. [Google Scholar] [CrossRef]
- Bohm, G.A.; Pfleiderer, W.; Boger, P.; Scherer, S. Structure of a novel oligosaccharide-mycosporine-amino acid ultraviolet A/B sunscreen pigment from the terrestrial cyanobacterium Nostoc commune. J. Biol. Chem. 1995, 270, 8536–8539. [Google Scholar] [CrossRef] [PubMed]
- Carreto, J.I.; Lutz, V.A.; De Marco, S.G.; Carignan, M.O. Fluence and wavelength dependence of mycosporine-like amino acid synthesis in the dinoflagellate Alexandrium excavatum. In Toxic Marine Phytoplankton; Graneli, E., Edler, L., Sundström, B., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 275–279. [Google Scholar]
- Karsten, U.; Sawall, T.; Wiencke, C. A survey of the distribution of UV-absorbing substances in tropical macroalgae. Phycol. Res. 1998, 46, 271–279. [Google Scholar] [CrossRef]
- Karsten, U.; Wiencke, C. Factors controlling the formation of UV-absorbing mycosporine-like amino acids in the marine red alga Palmaria palmata from Spitsbergen (Norway). J. Plant Physiol. 1999, 155, 407–415. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537–558. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P. Solar ultraviolet radiation-induced dna damage and protection/repair strategies in cyanobacteria. Int. J. Pharma Bio Sci. 2011, 2, 271–288. [Google Scholar]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The photoprotector mechanism of mycosporine-like amino acids. Excited-state properties and photostability of porphyra-334 in aqueous solution. J. Photochem. Photobiol. B Biol. 2000, 56, 139–144. [Google Scholar] [CrossRef]
- Whitehead, K.; Vernet, M. Influence of mycosporine-like amino acids (MAAs) on UV absorption by particulate and dissolved organic matter in La Jolla Bay. Limnol. Oceanogr. 2000, 45, 1788–1796. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O.; Montoya, N.G. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar. Biol. 2005, 146, 237–252. [Google Scholar] [CrossRef]
- Callone, A.I.; Carignan, M.; Montoya, N.G.; Carreto, J.I. Biotransformation of mycosporine like amino acids (MAAs) in the toxic dinoflagellate Alexandrium tamarense. J. Photochem. Photobiol. B Biol. 2006, 84, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. Experimental study of the excited-state properties and photostability of the mycosporine-like amino acid palythine in aqueous solution. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2007, 6, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Klisch, M.; Groniger, A.; Häder, D.P. Mycosporine-like amino acids in the marine red alga Gracilaria cornea - Effects of UV and heat. Environ. Exp. Bot. 2000, 43, 33–43. [Google Scholar] [CrossRef]
- Klisch, M.; Häder, D.P. Mycosporine-like amino acids and marine toxins-The common and the different. Mar. Drugs 2008, 6, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Schürch, C.; Zülli, F. UV-A sunscreen from red algae for protection against premature skin aging. Cosmetics 2004, 2004, 139–143. [Google Scholar]
- Gröniger, A.; Häder, D.P. Stability of mycosporine-like amino acids. Recent J. Photochem. Photobiol. 2000, 4, 247–252. [Google Scholar]
- Hirata, Y.; Uemura, D.; Ueda, K.; Takano, S. Several compounds from Palythoa tuberculosa (Coelenterata). Pure Appl. Chem. 1979, 51, 1875–1883. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, X.; Yuri, T.; Shingo, M.; Hiroo, O. Researches on the stability of porphyra-334 solution and its influence factors. J. Ocean Univ. China 2004, 3, 166–170. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D.; Incharoensakdi, A. Characterization and antioxidant functions of mycosporine-like amino acids in the Cyanobacterium Nostoc sp. R76DM. Algal Res. 2016, 16, 110–118. [Google Scholar] [CrossRef]
- Yoshiki, M.; Tsuge, K.; Tsuruta, Y.; Yoshimura, T.; Koganemaru, K.; Sumi, T.; Matsui, T.; Matsumoto, K. Production of new antioxidant compound from mycosporine-like amino acid, porphyra-334 by heat treatment. Food Chem. 2009, 113, 1127–1132. [Google Scholar] [CrossRef]
- Shick, J.M.; Romaine-Lioud, S.; Ferrier-Pages, C.; Gattuso, J.P. Ultraviolet-B radiation stimulates shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora pistillata despite decreases in its population of symbiotic dinoflagellates. Limnol. Oceanogr. 1999, 44, 1667–1682. [Google Scholar] [CrossRef]
- Portwich, A.; García-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp. strain PCC 6912. Phycologia 2003, 42, 384–392. [Google Scholar] [CrossRef]
- Favre-Bonvin, J.; Arpin, N.; Brevard, C. Biosynthesis of micosporine: Mycosporine-glutaminol in Trichothecium roseum. Phytochemistry 1987, 26, 2509–2514. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Genome mining of mycosporine-like amino acid (MAA) synthesizing and non-synthesizing cyanobacteria: A bioinformatics study. Genomics 2010, 95, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Balskus, E.P.; Walsh, C.T. The genetic and molecular basis for sunscreen biosynthesis in cyanobacteria. Science 2010, 329, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Shinzato, C.; Shoguchi, E.; Kawashima, T.; Hamada, M.; Hisata, K.; Tanaka, M.; Fujie, M.; Fujiwara, M.; Koyanagi, R.; Ikuta, T.; et al. Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 2011, 476, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Tartarotti, B.; Sommaruga, R. Seasonal and ontogenetic changes of mycosporine-like amino acids in planktonic organisms from an alpine lake. Limnol. Oceanogr. 2006, 51, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Incharoensakdi, A. UV radiation-induced biosynthesis, stability and antioxidant activity of mycosporine-like amino acids (MAAs) in a unicellular cyanobacterium Gloeocapsa sp. CU2556. J. Photochem. Photobiol. B Biol. 2014, 130, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Incharoensakdi, A. UV radiation-induced accumulation of photoprotective compounds in the green alga Tetraspora sp. CU2551. Plant Physiol. Biochem. 2013, 70, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Taira, H.; Aoki, S.; Yamanoha, B.; Taguchi, S. Daily variation in cellular content of UV-absorbing compounds mycosporine-like amino acids in the marine dinoflagellate Scrippsiella sweeneyae. J. Photochem. Photobiol. B Biol. 2004, 75, 145–155. [Google Scholar] [CrossRef]
- Klisch, M.; Häder, D.P. Wavelength dependence of mycosporine-like amino acid synthesis in Gyrodinium dorsum. J. Photochem. Photobiol. B. 2002, 66, 60–66. [Google Scholar] [CrossRef]
- Laurion, I.; Roy, S. Growth and photoprotection in three dinoflagellates (including two strains of Alexandrium tamarense) and one diatom exposed to four weeks of natural and enhanced ultraviolet-B radiation. J. Phycol. 2009, 45, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Neale, P.J.; Banaszak, A.T.; Jarriel, C.R. Ultraviolet sunscreens in Gymnodinium sanguineum (Dynophyceae): Mycosporine-like amino acids protect against inhibition of photosynthesis. J. Phycol. 1998, 938, 928–938. [Google Scholar] [CrossRef]
- Riegger, L.; Robinson, D. Photoinduction of UV-absorbing compounds in Antarctic diatoms and Phaeocystis antarctica. Mar. Ecol. Prog. Ser. 1997, 160, 13–25. [Google Scholar] [CrossRef]
- Hannach, G.; Sigleo, A.C. Photoinduction of UV-absorbing compounds in six species of marine phytoplankton. Mar. Ecol. Prog. Ser. 1998, 174, 207–222. [Google Scholar] [CrossRef]
- Hernando, M.; Carreto, J.I.; Carignan, M.O.; Ferreyra, G.A.; Gross, C. Effects of solar radiation on growth and mycosporine-like amino acids content in Thalassiosira sp, an Antarctic diatom. Polar Biol. 2002, 25, 12–20. [Google Scholar] [CrossRef]
- Kräbs, G.; Bischof, K.; Hanelt, D.; Karsten, U.; Wiencke, C. Wavelength-dependent induction of UV-absorbing mycosporine-like amino acids in the red alga Chondrus crispus under natural solar radiation. J. Exp. Mar. Biol. Ecol. 2002, 268, 69–82. [Google Scholar] [CrossRef]
- Franklin, L.A.; Kräbs, G.; Kuhlenkamp, R. Blue light and UV-A radiation control the synthesis of mycosporine-like amino acids in Chondrus crispus (Florideophyceae). J. Phycol. 2001, 37, 257–270. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Effects of abiotic stressors on synthesis of the mycosporine-like amino acid shinorine in the cyanobacterium Anabaena variabilis PCC 7937. Photochem. Photobiol. 2008, 84, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Ambasht, N.K.; Sinha, J.P.; Klisch, M.; Häder, D.P. UV-B-induced synthesis of mycosporine-like amino acids in three strains of Nodularia (cyanobacteria). J. Photochem. Photobiol. B Biol. 2003, 71, 51–58. [Google Scholar] [CrossRef]
- Lesser, M.P. Depth-dependent photoacclimatization to solar ultraviolet radiation in the Caribbean coral Montastrea faveolata. Mar. Ecol. Prog. Ser. 2000, 192, 137–151. [Google Scholar] [CrossRef]
- Singh, S.P.; Sinha, R.P.; Klisch, M.; Häder, D.P. Mycosporine-like amino acids (MAAs) profile of a rice-field cyanobacterium Anabaena doliolum as influenced by PAR and UVR. Planta 2008, 229, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Reef, R.; Kaniewska, P.; Hoegh-Guldberg, O. Coral sekeletons defence against ultraviolet radiation. PLoS ONE 2009, 4, e7995. [Google Scholar] [CrossRef] [PubMed]
- Portwich, A.; García-Pichel, F. Ultraviolet and osmotic stresses induce and regulate the synthesis of mycosporines in the cyanobacterium Chlorogloeopsis PCC 6912. Arch. Microbiol. 1999, 172, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol. Lett. 2007, 269, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shick, J.M.; Dunlap, W.C.; Chalker, B.E.; Banaszak, A.T.; Rosenzweig, T.K. Survey of ultraviolet radiation-absorbing mycosporine-like amino acids in organs of coral reef holothuroids. Mar. Ecol. Prog. Ser. 1992, 90, 139–148. [Google Scholar] [CrossRef]
- Portwich, A.; García-Pichel, F. A novel prokaryotic UVB photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912. Photochem. Photobiol. 2000, 71, 493–498. [Google Scholar] [CrossRef]
- Korbee Peinado, N.; Abdala Díaz, R.T.; Figueroa, F.L.; Helbling, E.W. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J. Phycol. 2004, 40, 248–259. [Google Scholar] [CrossRef]
- Litchman, E.; Neale, P.J.; Banaszak, A.T. Increased sensitivity to ultraviolet radiation in nitrogen-limited dinoflagellates: Photoprotection and repair. Limnol. Oceanogr. 2002, 47, 86–94. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Airs, R.L. Distribution and abundance of MAAs in 33 species of microalgae across 13 classes. Mar. Drugs 2010, 8, 1273–1291. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Sulfur deficiency changes mycosporine-like amino acid (MAA) composition of Anabaena variabilis PCC 7937: A possible role of sulfur in MAA bioconversion. Photochem. Photobiol. 2010, 86, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.L.; Karentz, D. The induction and repair of DNA photodamage in the environment. In Environmental UV Photobiology; Young, A., Moan, J., Björn, L., Nultsch, W., Eds.; Springer: New York, NY, USA, 1993; pp. 345–377. ISBN 9781489924087. [Google Scholar]
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.P. Ultraviolet-absorbing/screening substances in cyanobacteria, phytoplankton and macroalgae. J. Photochem. Photobiol. B Biol. 1998, 47, 83–94. [Google Scholar] [CrossRef]
- Cardozo, K.H.M.; Carvalho, V.M.; Pinto, E.; Colepicolo, P. Fragmentation of mycosporine-like amino acids by hydrogen/deuterium exchange and electrospray ionisation tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Sommaruga, R.; Whitehead, K.; Shick, J.M.; Lobban, C.S. Mycosporine-like amino acids in the zooxanthella-ciliate symbiosis Maristentor dinoferus. Protist 2006, 157, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Enk, C.D.; Hochberg, M.; Srebnik, M. Porphyra-334, a potential natural source for UVA protective sunscreens. Photochem. Photobiol. Sci. 2006, 5, 432–435. [Google Scholar] [CrossRef] [PubMed]
- McClintock, J.B.; Karentz, D. Mycosporine-like amino acids in 38 species of subtidal marine organisms from McMurdo Sound, Antarctica. Antarct. Sci. 1997, 9, 392–398. [Google Scholar] [CrossRef]
- Volkmann, M.; Gorbushina, A.A.; Kedar, L.; Oren, A. Structure of euhalothece-362, a novel red-shifted mycosporine-like amino acid, from a halophilic cyanobacterium (Euhalothece sp.). FEMS Microbiol. Lett. 2006, 258, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P.; Gómez, I.; Figueroa, F.L.; Ulloa, N.; Morales, V.; Lovengreen, C. Ultraviolet-absorbing mycosporine-like amino acids in red macroalgae from Chile. Bot. Mar. 2004, 47, 21–29. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.J.; Rhee, J.S.; Choi, E.M.; Brown, M.T.; Häder, D.P.; et al. Ultraviolet radiation and cyanobacteria. J. Photochem. Photobiol. B Biol. 2014, 141, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Helbling, E.W.; Menchi, C.F.; Villafañe, V.E. Bioaccumulation and role of UV-absorbing compounds in two marine crustacean species from Patagonia, Argentina. Photochem. Photobiol. Sci. 2002, 1, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.J.; Dunlap, W.C.; Nicol, S.; Ritz, D. Antarctic krill (Euphausia superba) acquire a UV-absorbing mycosporine-like amino acid from dietary algae. J. Exp. Mar. Biol. Ecol. 2000, 255, 93–110. [Google Scholar] [CrossRef]
- Kicklighter, C.E.; Kamio, M.; Nguyen, L.; Germann, M.W.; Derby, C.D. Mycosporine-like amino acids are multifunctional molecules in sea hares and their marine community. Proc. Natl. Acad. Sci. USA 2011, 108, 11494–11499. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.T. The impact of UVB radiation on marine plankton. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1998, 422, 119–129. [Google Scholar] [CrossRef]
- Holzinger, A.; Lutz, C. Algae and UV irradiation: Effects on ultrastructure and related metabolic functions. Micron 2006, 37, 190–207. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.P. Life under solar UV radiation in aquatic organisms. Adv. Space Res. 2002, 30, 1547–1556. [Google Scholar] [CrossRef]
- Stochaj, W.R.; Dunlap, W.C.; Shick, J.M. Two new UV-absorbing mycosporine-like amino acids from the sea anemone Anthopleura elegantissima and the effects of zooxanthellae and spectral irradiance on chemical composition and content. Mar. Biol. 1994, 118, 149–156. [Google Scholar] [CrossRef]
- Osborn, A.R.; Almabruk, K.H.; Holzwarth, G.; Asamizu, S.; LaDu, J.; Kean, K.M.; Karplus, P.A.; Tanguay, R.L.; Bakalinsky, A.T.; Mahmud, T. De novo synthesis of a sunscreen compound in vertebrates. eLife 2015, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Traverso, G. Why some fish don′t tan. Sci. Transl. Med. 2015, 7, 290–293. [Google Scholar] [CrossRef]
- Mason, D.S.; Schafer, F.; Shick, J.M.; Dunlap, W.C. Ultraviolet radiation-absorbing mycosporine-like amino acids (MAAs) are acquired from their diet by medaka fish (Oryzias latipes) but not by SKH-1 hairless mice. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 120, 587–598. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Williams, D.M.; Chalker, B.E.; Banaszak, A.T. Biochemical photoadaptation in vision: UV-absorbing pigments in fish eye tissues. Comp. Biochem. Physiol. B 1989, 93, 601–607. [Google Scholar] [CrossRef]
- Przeslawski, R.; Benkendorff, K.; Davis, A.R. A quantitative survey of mycosporine-like amino acids (MAAs) in intertidal egg masses from temperate rocky shores. J. Chem. Ecol. 2005, 31, 2417–2438. [Google Scholar] [CrossRef] [PubMed]
- Carefoot, T.H.; Karentz, D.; Pennings, S.C.; Young, C.L. Distribution of mycosporine-like amino acids in the sea hare Aplysia dactylomela: Effect of diet on amounts and types sequestered over time in tissues and spawn. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2000, 126, 91–104. [Google Scholar] [CrossRef]
- Oyamada, C.; Kaneniwa, M.; Ebitani, K.; Murata, M.; Ishihara, K. Mycosporine-like amino acids extracted from scallop (Patinopecten yessoensis) ovaries: UV protection and growth stimulation activities on human cells. Mar. Biotechnol. 2008, 10, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Braun, C.; Reef, R.; Siebeck, U.E. Ultraviolet absorbing compounds provide a rapid response mechanism for UV protection in some reef fish. J. Photochem. Photobiol. B Biol. 2016, 160, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The deactivation pathways of the excited-states of the mycosporine-like amino acids shinorine and porphyra-334 in aqueous solution. Photochem. Photobiol. Sci. 2004, 3, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.P.; Sinha, R.P. Solar ultraviolet radiation-induced DNA damage in aquatic organisms: Potential environmental impact. Mutat. Res. 2005, 571, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K.; Hedges, J.I. Photodegradation and photosensitization of mycosporine-like amino acids. J. Photochem. Photobiol. B Biol. 2005, 80, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Shick, J.M.; Lesser, M.P.; Jokiel, P.L. Effects of ultraviolet radiation on corals and other coral reef organisms. Glob. Chang. Biol. 1996, 2, 527–545. [Google Scholar] [CrossRef]
- Misonou, T.; Saitoh, J.; Oshiba, S.; Tokitomo, Y.; Maegawa, M.; Inoue, Y.; Hori, H.; Sakurai, T. UV-absorbing substance in the red alga Porphyra yezoensis (Bangiales, Rhodophyta) block thymine photodimer production. Mar. Biotechnol. 2003, 5, 194–200. [Google Scholar] [CrossRef] [PubMed]
- García-Pichel, F.; Castenholz, R.W. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl. Environ. Microbiol. 1993, 59, 163–169. [Google Scholar] [PubMed]
- García-Pichel, F.; Wingard, C.E.; Castenholz, R.W. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the Cyanobacterium Gloeocapsa sp. Appl. Environ. Microbiol. 1993, 59, 170–176. [Google Scholar] [PubMed]
- Lechowski, Z.; Białczyk, J. Barwniki ekranujące słoneczne promieniowanie ultrafioletowe u roślin i grzybów [Solar ultraviolet radiation screening pigments in plants and fungi]. Wiadomości Bot. 2003, 47, 41–54. [Google Scholar]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K.; Karentz, D.; Hedges, J. Mycosporine-like amino acids (MAAs) in phytoplankton, a herbivorous pteropod (Limacina helicina), and its pteropod predator (Clione antarctica) in McMurdo Bay, Antarctica. Mar. Biol. 2001, 139, 1013–1019. [Google Scholar] [CrossRef]
- Leavitt, P.R.; Vinebrooke, R.D.; Donald, D.B.; Smol, J.P.; Schindler, D.W. Past ultraviolet radiation environments in lakes derived from fossil pigments. Nature 1997, 388, 457–459. [Google Scholar] [CrossRef]
- Suh, H.J.; Lee, H.W.; Jung, J. Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency. Photochem. Photobiol. 2003, 78, 109–113. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Yamamoto, Y. Small-molecule antioxidants in marine organisms: Antioxidant activity of mycosporine-glycine. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 112, 105–114. [Google Scholar] [CrossRef]
- Tao, C.; Sugawara, T.; Maeda, S.; Wang, X.; Hirata, T. Antioxidative activities of a mycosporine-like amino acid, porphyra-334. Fish. Sci. 2008, 74, 1166–1172. [Google Scholar] [CrossRef]
- Suh, S.S.; Hwang, J.; Park, M.; Seo, H.H.; Kim, H.S.; Lee, J.H.; Moh, S.H.; Lee, T.K. Anti-inflammation activities of mycosporine-like amino acids (MAAs) in response to UV radiation suggest potential anti-skin aging activity. Mar. Drugs 2014, 12, 5174–5187. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Sharma, K.; Namdeo, A.G.; Chaugule, B.B.; Kavale, M.; Nanda, S. Broad-spectrum sun-protective action of porphyra-334 derived from Porphyra vietnamensis. Pharmacogn. Res. 2010, 2, 45–49. [Google Scholar] [CrossRef] [PubMed]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L.; de Gálvez, M.V.; Herrera, E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J. Appl. Phycol. 2009, 21, 161–169. [Google Scholar] [CrossRef]
- Kogej, T.; Gostinčar, C.; Volkmann, M.; Gorbushina, A.A.; Gunde-Cimerman, N. Mycosporines in extremophilic fungi-Novel complementary osmolytes? Environ. Chem. 2006, 3, 105–110. [Google Scholar] [CrossRef]
- Jiang, H.; Gao, K.; Helbling, E.W. UV-absorbing compounds in Porphyra haitanensis (Rhodophyta) with special reference to effects of desiccation. J. Appl. Phycol. 2008, 20, 387–395. [Google Scholar] [CrossRef]
- Singh, S.P.; Häder, D.P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (UVR) stress: Mitigation strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Sommaruga, R. The role of solar UV radiation in the ecology of alpine lakes. J. Photochem. Photobiol. B Biol. 2001, 62, 35–42. [Google Scholar] [CrossRef]
- Arpin, N.; Bouillant, M.L. Light and mycosporines. In The Fungal Spore: Morphogenetic Controls; Turian, G., Hohl, H.R., Eds.; Academic Press: London, UK, 1981; pp. 159–173. ISBN 9780127036809. [Google Scholar]
- Bandaranayake, W.M.; Bourne, D.J.; Sim, R.G. Chemical composition during maturing and spawning of the sponge Dysidea herbacea (Porifera: Demospongiae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1997, 118, 851–859. [Google Scholar] [CrossRef]
- Bandaranayake, W.M.; Des Rocher, A. Role of secondary metabolites and pigments in the epidermal tissues, ripe ovaries, viscera, gut contents and diet of the sea cucumber Holothuria atra. Mar. Biol. 1999, 133, 163–169. [Google Scholar] [CrossRef]
- Gao, K.; Wu, Y.; Li, G.; Wu, H.; Villafañe, V.E.; Helbling, E.W. Solar UV radiation drives CO2 fixation in marine phytoplankton: A double-edged sword. Plant Physiol. 2007, 144, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, A.; Psotová, J.; Walterová, D. Natural phenolics in the prevention of UV-induced skin damage. A review. Biomed. Pap. 2003, 147, 137–145. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Schulman, J.M.; Fisher, D.E. Indoor ultraviolet tanning and skin cancer: Health risks and opportunities. Curr. Opin. Oncol. 2009, 21, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Sklar, L.R.; Almutawa, F.; Lim, H.W.; Hamzavi, I. Effects of ultraviolet radiation, visible light, and infrared radiation on erythema and pigmentation: A review. Photochem. Photobiol. Sci. 2013, 12, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kripke, M.L. Immunological unresponsiveness induced by ultraviolet radiation. Immunol. Rev. 1984, 80, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Halliday, G.M. Inflammation, gene mutation and photoimmunosuppression in response to UVR-induced oxidative damage contributes to photocarcinogenesis. Mutat. Res. 2005, 571, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.P.; Lee, T.K. Adverse effects of ultraviolet radiation: A brief review. Prog. Biophys. Mol. Biol. 2006, 92, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.C.; Krutmann, J.; Elmets, C.A.; Bergstresser, P.R.; Cruz, P.D. Ultraviolet B-irradiated antigen-presenting cells display altered accessory signaling for T-cell activation: Relevance to immune responses initiated in skin. J. Investig. Dermatol. 1992, 98, S66–S69. [Google Scholar] [CrossRef]
- Vermeer, M.; Streilein, J.W. Ultraviolet B light-induced alterations in epidermal Langerhans cells are mediated in part by tumor necrosis factor-alpha. Photodermatol. Photoimmunol. Photomed. 1990, 7, 258–265. [Google Scholar] [PubMed]
- Chung, H.T.; Burnham, D.K.; Robertson, B.; Roberts, L.K.; Daynes, R.A. Involvement of prostaglandins in the immune alterations caused by the exposure of mice to ultraviolet radiation. J. Immunol. 1986, 137, 2478–2484. [Google Scholar] [PubMed]
- Capote, R.; Alonso-Lebrero, J.L.; García-Pichel, F.; Brieva, A.; Pivel, J.P.; González, S. Polypodium leucotomos extract inhibits trans-urocanic acid photoisomerization and photodecomposition. J. Photochem. Photobiol. B Biol. 2006, 82, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Park, K.; Kweon, G.R.; Jang, B.C.; Baek, W.K.; Suh, M.H.; Kim, C.W.; Lee, K.S.; Suh, S.I. Curcumin inhibits the expression of COX-2 in UVB-irradiated human keratinocytes (HaCaT) by inhibiting activation of AP-1: p38 MAP kinase and JNK as potential upstream targets. Exp. Mol. Med. 2005, 37, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Park, H.M.; Moon, E.; Kim, A.J.; Kim, M.H.; Lee, S.; Lee, J.B.; Park, Y.K.; Jung, H.S.; Kim, Y.B.; Kim, S.Y. Extract of Punica granatum inhibits skin photoaging induced by UVB irradiation. Int. J. Dermatol. 2010, 49, 276–282. [Google Scholar] [CrossRef] [PubMed]
- De Gruijl, F.R. Photocarcinogenesis: UVA vs. UVB radiation. Ski. Pharmacol. Appl. Ski. Physiol. 2002, 15, 316–320. [Google Scholar] [CrossRef]
- Besaratinia, A.; Synold, T.W.; Chen, H.H.; Chang, C.; Xi, B.; Riggs, A.D.; Pfeifer, G.P. DNA lesions induced by UV A1 and B radiation in human cells: Comparative analyses in the overall genome and in the p53 tumor suppressor gene. Proc. Natl. Acad. Sci. USA 2005, 102, 10058–10063. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, G.P.; You, Y.H.; Besaratinia, A. Mutations induced by ultraviolet light. Mutat. Res. 2005, 571, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Setlow, R.B.; Carrier, W.L. Pyrimidine dimers in ultraviolet-irradiated DNA’s. J. Mol. Biol. 1966, 17, 237–254. [Google Scholar] [CrossRef]
- Häder, D.P.; Worrest, R.C.; Kumar, H.D.; Smith, R.C. Effects of increased solar ultraviolet radiation on aquatic ecosystems. Ambio 1995, 24, 174–180. [Google Scholar]
- Sinha, R.P.; Lebert, M.; Kumar, A.; Kumar, H.D.; Häder, D.P. Spectroscopic and biochemical analyses of UV effects on phycobiliproteins of Anabaena sp. and Nostoc carmium. Bot. Acta 1995, 108, 87–92. [Google Scholar] [CrossRef]
- Sinha, R.P.; Singh, N.; Kumar, A.; Kumar, H.D.; Häder, M.; Häder, D.P. Effects of UV irradiation on certain physiological and biochemical processes in cyanobacteria. J. Photochem. Photobiol. B Biol. 1996, 32, 107–113. [Google Scholar] [CrossRef]
- Helbling, E.W.; Villafrañe, V.; Ferrario, M.; Holm-Hansen, O. Impact of natural ultraviolet radiation on rates of photosynthesis and on spedcific marine phytoplankton species. Mar. Ecol. Prog. Ser. 1992, 80, 89–100. [Google Scholar] [CrossRef]
- Mitchell, D.L. The relative cytotoxicity of (6–4) photoproducts and cyclobutane dimers in mammalian cells. Photochem. Photobiol. 1988, 48, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.L.; Nairn, R.S. The biology of the (6–4) photoproduct. Photochem. Photobiol. 1989, 49, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.P.; Kumar, H.D.; Smith, R.C.; Worrest, R.C. Effects of solar UV radiation on aquatic ecosystems and interactions with climate change. Photochem. Photobiol. Sci. 2007, 6, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K. Silymarin and skin cancer prevention: Anti-inflammatory, antioxidant and immunomodulatory effects. Int. J. Oncol. 2005, 26, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Pustišek, N.; Šitum, M. UV-radiation, apoptosis and skin. Coll. Antropol. 2011, 35, 339–341. [Google Scholar] [PubMed]
- Trautinger, F. Mechanisms of photodamage of the skin and its functional consequences for skin ageing. Clin. Exp. Dermatol. 2001, 26, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Ehrhart, J.C.; Gosselet, F.P.; Culerrier, R.M.; Sarasin, A. UVB-induced mutations in human key gatekeeper genes governing signalling pathways and consequences for skin tumourigenesis. Photochem. Photobiol. Sci. 2003, 2, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Moan, J.; Porojnicu, A.C.; Dahlback, A. Ultraviolet radiation and malignant melanoma. Adv. Exp. Med. Biol. 2008, 624, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Agar, N.S.; Halliday, G.M.; Barnetson, R.S.; Ananthaswamy, H.N.; Wheeler, M.; Jones, A.M. The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: A role for UVA in human skin carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4954–4959. [Google Scholar] [CrossRef] [PubMed]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Endogenous UVA-photosensitizers: Mediators of skin photodamage and novel targets for skin photoprotection. Photochem. Photobiol. Sci. 2006, 5, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Erden Inal, M.; Kahraman, A.; Köken, T. Beneficial effects of quercetin on oxidative stress induced by ultraviolet A. Clin. Exp. Dermatol. 2001, 26, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Thiele, J.; Elsner, P. Oxidants and antioxidants in cutaneous biology. In Current Problem in Dermatology; Burg, G., Ed.; Karger: Basel, Switzerland, 2001; ISBN 9783805571326. [Google Scholar]
- Lesser, M.P.; Farrell, J.H. Exposure to solar radiation increases damage to both host tissues and algal symbionts of corals during thermal stress. Coral Reefs 2004, 23, 367–377. [Google Scholar] [CrossRef]
- Tyrrell, R.M. UVA (320–380 nm) as an oxidative stress. In Oxidative Stress–Oxidants and Antioxidants; Sies, H., Ed.; Academic Press: London, UK, 1991; pp. 57–84. ISBN 9780126427622. [Google Scholar]
- Dykens, J.A.; Shick, J.M.; Benoit, C.; Buettner, G.R.; Winston, G.W. Oxygen radical production in the sea anemone Anthopleura elegantissima and its endosymbiotic algae. J. Exp. Biol. 1992, 168, 219–241. [Google Scholar]
- Shick, J.M.; Lesser, M.P.; Stochaj, W.R. Ultraviolet radiation and photooxidative stress in zooxanthellate Anthozoa: The sea anemone Phyllodiscus semoni and the octocoral Clavularia sp. Symbiosis 1991, 10, 145–173. [Google Scholar]
- Lesser, M.P.; Stochaj, W.R. Photoadaptation and protection against active forms of oxygen in the symbiotic procaryote Prochloron sp. and its ascidian host. Appl. Environ. Microbiol. 1990, 56, 1530–1535. [Google Scholar] [PubMed]
- Lesser, M.P. Acclimation of phytoplankton to UV-B radiation: Oxidative stress and photoinhibition of photosynthesis are not prevented by UV-absorbing compounds in the dinoflagellate Prorocentrum micans. Mar. Ecol. Prog. Ser. 1996, 132, 287–297. [Google Scholar] [CrossRef]
- Pinnell, S.R. Cutaneous photodamage, oxidative stress, and topical antioxidant protection. J. Am. Acad. Dermatol. 2003, 48, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Assefa, Z.; van Laethem, A.; Garmyn, M.; Agostinis, P. Ultraviolet radiation-induced apoptosis in keratinocytes: On the role of cytosolic factors. Biochim. Biophys. Acta Rev. Cancer 2005, 1755, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Mukhtar, H. Photochemoprevention by botanical antioxidants. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Adhami, V.M.; Syed, D.N.; Khan, N.; Afaq, F. Phytochemicals for prevention of solar ultraviolet radiation-induced damages. Photochem. Photobiol. 2008, 84, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J. The role of UVA rays in skin aging. Eur. J. Dermatol. 2001, 11, 170–171. [Google Scholar] [PubMed]
- Ichihashi, M.; Ando, H.; Yoshida, M.; Niki, Y.; Matsui, M. Photoaging of the skin. Anti Aging Med. 2009, 6, 46–59. [Google Scholar] [CrossRef]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Mukhtar, H. Effects of solar radiation on cutaneous detoxification pathways. J. Photochem. Photobiol. B Biol. 2001, 63, 61–69. [Google Scholar] [CrossRef]
- Pathak, M.A. Functions of melanin and protection by melanin. In Melanin: Its Role in Human Photoprotection; Zeise, L., Chedekel, M.R., Fitzpatrick, T.B., Eds.; Valdemar Publishing Company: Overland Park, KS, USA, 1995; pp. 125–134. ISBN 9780963210531. [Google Scholar]
- González, S.; Fernández-Lorente, M.; Gilaberte-Calzada, Y. The latest on skin photoprotection. Clin. Dermatol. 2008, 26, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.J. Sunscreens: Development: Evaluation, and Regulatory Aspects; Marcel Dekker: New York, NY, USA, 1996; ISBN 9780824793067. [Google Scholar]
- Osterwalder, U.; Sohn, M.; Herzog, B. Global state of sunscreens. Photodermatol. Photoimmunol. Photomed. 2014, 30, 62–80. [Google Scholar] [CrossRef] [PubMed]
- Schlossmann, D.; Shao, Y. Inorganic ultraviolet filters. In Sunscreens–Regulations and Commercial Development; Shaath, N.A., Ed.; Taylor and Francis Group: New York, NY, USA, 2005; pp. 239–279. ISBN 9780824757946. [Google Scholar]
- Dromgoole, S.H.; Maibach, H.I. Sunscreening agent intolerance: Contact and photocontact sensitization and contact urticaria. J. Am. Acad. Dermatol. 1990, 22, 1068–1078. [Google Scholar] [CrossRef]
- Nałęcz-Jawecki, G.; Zawadzki, T.; Skrzypczak, A. Sunscreens and the environment. Biul. Wydz. Farm. Warsz. Uniw. Med. 2012, 5, 32–39. [Google Scholar]
- Morabito, K.; Shapley, N.C.; Steeley, K.G.; Tripathi, A. Review of sunscreen and the emergence of non-conventional absorbers and their applications in ultraviolet protection. Int. J. Cosmet. Sci. 2011, 33, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Giokas, D.L.; Salvador, A.; Chisvert, A. UV filters: From sunscreens to human body and the environment. Trends Food Sci. Technol. 2007, 26, 360–374. [Google Scholar] [CrossRef]
- Klann, A.; Levy, G.; Lutz, I.; Müller, C.; Kloas, W.; Hildebrandt, J.P. Estrogen-like effects of ultraviolet screen 3-(4-methylbenzylidene)-camphor (Eusolex 6300) on cell proliferation and gene induction in mammalian and amphibian cells. Environ. Res. 2005, 97, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Raikou, V.; Protopapa, E.; Kefala, V. Photo-protection from marine organisms. Rev. Clin. Pharmacol. Pharmacokinet. Int. Ed. 2011, 25, 131–136. [Google Scholar]
- Senevirathne, W.S.M.; Kim, S.K. Cosmeceuticals from algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Dominguez, H., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition: Belihuloya, Sri Lanka, 2013; pp. 694–713. ISBN 9780857095121. [Google Scholar]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Bioactive components from seaweeds: Cosmetic applications and future development. In Advances in Botanical Research; Bourgougnon, N., Ed.; Academic Press: London, UK, 2014; Volume 71, pp. 345–378. ISBN 9780124080621. [Google Scholar]
- Schmid, D.; Schürch, C.; Zülli, F.; Nissem, H.P.; Prieur, H. Mycosporine-like amino acids: Natural UV-screening compounds from red algae to protect the skin against photoaging. SÖFW J. 2003, 129, 38–42. [Google Scholar]
- Ryu, J.; Park, S.J.; Kim, I.H.; Choi, Y.H.; Nam, T.J. Protective effect of porphyra-334 on UVA-induced photoaging in human skin fibroblasts. Int. J. Mol. Med. 2014, 34, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; You, D.H.; Han, T.; Choi, E.M. Modulation of viability and apoptosis of UVB-exposed human keratinocyte HaCaT cells by aqueous methanol extract of laver (Porphyra yezoensis). J. Photochem. Photobiol. B Biol. 2015, 141, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.S.; Oh, S.K.; Lee, S.G.; Kim, I.C.; Kim, S. Porphyra-334, a mycosporine-like amino acid, attenuates UV-induced apoptosis in HaCaT cells. Acta Pharm. 2017, 67, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.S.; Lee, S.; Youn, U.; Han, S.; Kim, I.C.; Kim, S. Comprehensive expression profiling and functional network analysis of porphyra-334, one mycosporine-like amino acid (MAA), in human keratinocyte exposed with UV-radiation. Mar. Drugs 2017, 15, 196. [Google Scholar] [CrossRef] [PubMed]
- De la Coba, F.; Aguilera, J.; de Gálvez, M.V.; Álvarez, M.; Gallego, E.; Figueroa, F.L.; Herrera, E. Prevention of the ultraviolet effects on clinical and histopathological changes, as well as the heat shock protein-70 expression in mouse skin by topical application of algal UV-absorbing compounds. J. Dermatol. Sci. 2009, 55, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Tosato, M.G.; Orallo, D.E.; Ali, S.M.; Churio, M.S.; Martin, A.A.; Dicelio, L. Confocal Raman spectroscopy: In vivo biochemical changes in the human skin by topical formulations under UV radiation. J. Photochem. Photobiol. B Biol. 2015, 153, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.V.; Westcott, N.D.; Hu, C.; Kitts, D.D. Mycosporine-like amino acid composition of the edible red alga, Palmaria palmata (dulse) harvested from the west and east coasts of Grand Manan Island, New Brunswick. Food Chem. 2009, 112, 321–328. [Google Scholar] [CrossRef]
- Athukorala, Y.; Trang, S.; Kwok, C.; Yuan, Y.V. Antiproliferative and antioxidant activities and mycosporine-like amino acid profiles of wild-harvested and cultivated edible Canadian marine red macroalgae. Molecules 2016, 21, 119. [Google Scholar] [CrossRef] [PubMed]
- Scoglio, S.; Lo Curcio, V.; Catalani, S.; Palma, F.; Battistelli, S.; Benedetti, S. Inhibitory effects of Aphanizomenon flos-aquae constituents on human UDP-glucose dehydrogenase activity. J. Enzyme Inhib. Med. Chem. 2016, 6366, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Schürch, C.; Zülli, F. Mycosporine-like amino acids from red algae protect against premature skin-aging. Euro Cosmet. 2006, 9, 1–4. [Google Scholar]
- Hartmann, A.; Gostner, J.; Fuchs, J.E.; Chaita, E.; Aligiannis, N.; Skaltsounis, L.; Ganzera, M. Inhibition of collagenase by mycosporine-like amino acids from marine sources. Planta Med. 2015, 81, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, W.C.; Chalker, B.E.; Bandaranayake, W.M. Nature’s sunscreen from the Great Barrier Reef, Australia. Int. J. Cosmet. Sci. 1998, 51, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.C.M.; Alonso-Varona, A.; Palomares, T.; Zubillaga, V.; Labidi, J.; Bulone, V. Exploiting mycosporines as natural molecular sunscreens for the fabrication of UV-absorbing green materials. ACS Appl. Mater. Interfaces 2015, 7, 16558–16564. [Google Scholar] [CrossRef] [PubMed]
- Andre, G.; Pellegrini, M.; Pellegrini, L. Algal Extracts Containing Amino Acid Analogs of Mycosporin Are Useful as Dermatological Protecting Agents against Ultraviolet Radiation. Patent No. FR2803201, 6 July 2001. [Google Scholar]
- Colabella, F.; Moliné, M.; Libkind, D. UV sunscreens of microbial origin: Mycosporines and mycosporine-like aminoacids. Recent Pat. Biotechnol. 2015, 8, 179–193. [Google Scholar] [CrossRef]
- Barceló-Villalobos, M.; Figueroa, F.L.; Korbee, N.; Álvarez-Gómez, F.; Abreu, M.H. Production of mycosporine-like amino acids from Gracilaria vermiculophylla (Rhodophyta) cultured through one year in an Integrated Multi-Trophic Aquaculture (IMTA) system. Mar. Biotechnol. 2017, 19, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, P.J.; Fitzmaurice, N.; Rigg, D.J.; Thang, S.H.; Bird, G. UV-Absorbing Compounds and Compositions. Patent No. WO1990009995, 7 September 1990. [Google Scholar]
- Bird, G.; Fitzmaurice, N.; Dunlap, W.C.; Chalker, B.E.; Banadaranayake, W.M. Sunscreen Compositions and Compounds for Use Therein. Patent No. WO1988002251, 7 April 1988. [Google Scholar]
- Andreguetti, D.; Stein, E.M.; Pereira, C.M.P.; Pinto, E.; Colepicolo, P. Antioxidant properties and UV absorbance pattern of mycosporine-like amino acids analogs synthesized in an environmentally friendly manner. J. Biochem. Mol. Toxicol. 2013, 27, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae (AFA klamath), Active Compounds, and Their Uses. Patent No. EP2032122, 13 April 2016. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae and Nutritional, Cosmetic and Pharmaceutical Compositions Containing the Same. Patent No. US8337858, 25 December 2012. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae and Nutritional, Cosmetic and Pharmaceutical Compositions Containing the Same. Patent No. US20100021493, 28 January 2010. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae and Nutritional, Cosmetic and Pharmaceutical Compositions Containing the Same. Patent No. WO2008000431, 3 January 2008. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae and Nutritional, Cosmetic and Pharmaceutical Compositions Containing the Same. Patent No. CA2656160, 3 January 2008. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon flos aquae and Nutritional, Cosmetic and Pharmaceutical Compositions Containing the Same. Patent No. CN101489527, 9 April 2014. [Google Scholar]
- Qvit-Raz, N.; Altman, T. Topical Composition Comprising Transformed Bacteria Expressing a Compound of Interest. Patent No. US20140044677, 13 February 2014. [Google Scholar]
- Qvit-Raz, N.; Altman, T. Topical Composition Comprising Transformed Bacteria Expressing a Compound of Interest. Patent No. US20160000701, 7 January 2016. [Google Scholar]
- Qvit-Raz, N. Topical Formulations for UV Protection. Patent No. WO2015195546, 23 December 2015. [Google Scholar]
- Ikeda, H.; Yamamoto, S.; Matsumoto, J.; Sota, M. Method for Producing Mycosporine-Like Amino Acid Using Microbes. Patent No. WO2015174427, 19 November 2015. [Google Scholar]
- York, M.; Ryan, J. Synthesis of UV Absorbing Compounds. Patent No. US20150299124, 22 October 2015. [Google Scholar]
- Ryan, J.; York, M. Synthesis of UV Absorbing Compounds. Patent No. WO2014082124, 5 June 2014. [Google Scholar]
- York, M.; Ryan, J.; Savage, G.P.; Meyer, A.G.; Jarvis, K. UV Absorbing Compounds, Compositions Comprising Same and Uses Thereof. Patent No. WO2015006803, 22 January 2015. [Google Scholar]
- York, M.; Ryan, J.; Savage, G.P.; Meyer, A.G.; Jarvis, K. UV Absorbing Compounds, Compositions Comprising Same and Uses Thereof. Patent No. US20160244409, 25 August 2016. [Google Scholar]
- Abou-Khalil, E.; Raeppel, S.; Raeppel, F. Imino Compounds as Protecting Agents Against Ultraviolet Radiations. Patent No. WO2013181741, 12 December 2013. [Google Scholar]
- Abou-Khalil, E.; Raeppel, S.; Raeppel, F. Imino Compounds as Protecting Agents Against Ultraviolet Radiations. Patent No. US20150152046, 4 June 2015. [Google Scholar]
- Ishihara, K.; Watanabe, R.; Suzuki, T.; Sakamoto, T.; Matsusato, S.; Wada, N.; Takenaka, H.; Yamaguchi, Y. Mycosporin-Like Amino Acids, Production Method Thereof, UV Protecting Agents and Antioxidants. Patent No. JP2014227339, 8 December 2014. [Google Scholar]
- Zhang, C.H.; Xu, J.C.; Xu, Z.H.; Gao, X. Preparation Method for Laver Mycosporine-Like Amino Acids Porphyra-334. Patent No. CN102659621, 12 September 2012. [Google Scholar]
- Su, Z.H.; Lin, J.; Wang, F. Beaty Product Containing Desert Algae Radiation-Proof Ingredient and Natural Medical Whitening Ingredient and Preparation Method Thereof. Patent No. CN102764206, 7 November 2012. [Google Scholar]
- Han, T.J.; Park, J.H. Method for Preparing UV Screening Nontoxic Extract from Red Algae, and Nontoxic Sunscreen Using Same. Patent No. WO2011096628, 11 August 2011. [Google Scholar]
- Han, T.J.; Park, J.H. Method for Preparing UV Screening Nontoxic Extract from Red Algae, and Nontoxic Sunscreen Using Same. Patent No. CN102740869, 17 October 2012. [Google Scholar]
- O’Connor, C.; Skill, S.; Llewellyn, C. Topical Composition. Patent No. WO2011158041, 22 December 2011. [Google Scholar]
- Wolf, F. Cosmetic Sunscreen Composition. Patent No. GB2472021, 26 January 2011. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Benedetti, Y.; Delgado-Esteban, M. Aphanizomenon Flos Aquae Preparation, Extracts and Purified Components Thereof for the Treatment of Neurological, Neurodegenerative and Mood Disorders. Patent No. EP2046354, 10 August 2011. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Benedetti, Y.; Delgado-Esteban, M. Aphanizomenon Flos Aquae Preparation, Extracts and Purified Components Thereof for the Treatment of Neurological, Neurodegenerative and Mood Disorders. Patent No. US20090311286, 17 December 2011. [Google Scholar]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Benedetti, Y.; Delgado-Esteban, M. Aphanizomenon Flos Aquae Preparation, Extracts and Purified Components Thereof for the Treatment of Neurological, Neurodegenerative and Mood Disorders. Patent No. WO2008000430, 3 January 2008. [Google Scholar]
- Han, T.J.; Park, J.H. Method for Manufacturing Non-Toxic Extract for Blocking UV from Red Algae. Patent No. KR100969325, 9 July 2010. [Google Scholar]
- Sakakibara, M.; Torii, M.; Miyamoto, M. Mycosporin-Like Amino Acid Derivative Having Glycosyl Group and Method for Producing the Same. Patent No. JP2009120562, 4 June 2009. [Google Scholar]
- Figueroa, F.L.; Aguilera, J.; de la Coba, F.; Korbee Peinado, N. Sunscreen Composition with Extract of Algae and Lichens. Patent No. ES2317741, 10 February 2010. [Google Scholar]
- Ewart, H.S.; Zhang, J.; Barrow, J.C. Compositions Comprising Porphyra and Methods of Making and Using Thereof. Patent No. WO2007144779, 21 December 2007. [Google Scholar]
- Ewart, H.S.; Zhang, J.; Barrow, J.C. Compositions Comprising Porphyra and Methods of Making and Using Thereof. Patent No. US20070220806, 27 September 2007. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L. Uso de Aminoácido Tipo Micosporina (Shinorine) en Productos Para Prevención y Tratamiento de Eritema Actínico, Fotocarcinogénesis y Fotoenvejecimiento. Patent No. ES2301426, 1 May 2008. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L. Uso de Aminoácido Tipo Micosporina (Porfira 334) En Productos Para Prevención de Procesos Cancerígenos. Patent No. ES2301293, 1 May 2009. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Lopez Figueroa, F. Use of a Mycosporin-Type Amino Acid (Porphyra 334) as An Antioxidant. Patent No. WO2007026035, 8 March 2007. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L. Use of a Mycosporin-Type Amino Acid (M-gly) As an Antioxidant. Patent No. WO2007026036, 8 March 2007. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L. Use of a Mixture of Mycosporin-Type Amino Acids (Asterin 330 + Palythine) As an Antioxidant. Patent No. WO2007026037, 8 March 2007. [Google Scholar]
- De la Coba, F.; Aguilera, J.; Figueroa, F.L. Use of a Mycosporin-Type Amino Acid (Shinorine) As an Antioxidant. Patent No. WO2007026038, 8 March 2007. [Google Scholar]
- Ishihara, K.; Oyamada, C.; Kaneniwa, M. Fibroblast Growth Promoter. Patent No. JP2007016004, 25 January 2007. [Google Scholar]
- Kunshang, G.; Ping, L.; Juntian, X.; Zhihui, C.; Yuming, Q. Cosmetic Including Natural Substance Having Sun-Screening Function. Patent No. CN101061995, 31 October 2007. [Google Scholar]
- Banowski, B.; Hoffmann, D.; Wadle, A.; Siegert, P.; Saettler, A.; Gerke, T. Beta-Glucuronidase Inhibitors for Use in Deodorants and Antiperspirants. Patent No. US20040234466, 25 November 2004. [Google Scholar]
- Banowski, B.; Hoffmann, D.; Wadle, A.; Siegert, P.; Saettler, A.; Gerke, T. Beta-Glucuronidase Inhibitors for Use in Deodorants and Antiperspirants. Patent No. US7294330, 13 November 2007. [Google Scholar]
- Baschong, W. Amino-Benzophenone UV Filter Formulations for the Prevention of Tanning. Patent No. GB2412866, 12 October 2005. [Google Scholar]
- Enk, D.C.; Srebnik, M.; Lev, O.; Hochberg, M.; Dor, I.; Torres, A.; Dembitsky, V.M. The Utilization of Natural Pigments From Lichens, Cyanobacteria, Fungi and Plants for Sun Protection. Patent No. WO2003020236, 13 March 2003. [Google Scholar]
- Enk, D.C.; Srebnik, M.; Lev, O.; Hochberg, M.; Dor, I.; Torres, A.; Dembitsky, V.M. The Utilization of Natural Pigments from Lichens, Cyanobacteria, Fungi and Plants for Sun Protection. Patent No. EP1424990, 9 June 2004. [Google Scholar]
- Enk, D.C.; Srebnik, M.; Lev, O.; Hochberg, M.; Dor, I.; Torres, A.; Dembitsky, V.M. The Utilization of Natural Pigments from Lichens, Cyanobacteria, Fungi and Plants for Sun Protection. Patent No. US20050129630, 16 June 2005. [Google Scholar]
- Schmid, D.; Schürch, C.; Zülli, F. Cosmetic Skin Care Products and Cosmetic Agents for Protecting Skin Against Premature Aging. Patent No. EP1473028, 3 November 2004. [Google Scholar]
- Krol, M.; Huner, N.; Ivanov, A.; Sarhan, F. Solar Radiation Protection Composition. Patent No. WO2000024369, 4 May 2000. [Google Scholar]
- Huner, N.; Krol, M.; Ivanov, A.; Sarhan, F. Solar Radiation Protection Composition. Patent No. EP1123083, 16 August 2001. [Google Scholar]
- Hunar, N.; Krol, M.; Ivanov, A.; Sarhan, F. Solar Radiation Protection Composition. Patent No. US6787147, 7 September 2004. [Google Scholar]
- Llewellyn, C.; Galley, E. Personal Care Compositions. Patent No. WO2002039974, 23 May 2002. [Google Scholar]
- Llewellyn, C.; Galley, E. Personal Care Compositions. Patent No. EP 1341514, 10 September 2003. [Google Scholar]
- Enk, D.C.; Hochberg, M.; Torres, A.; Lev, O.; Dor, I.; Dembitsky, V.M.; Srebnik, M. Natural UV Filters Derived from Pigments of Lichens. Patent No. IL0200725, 2002. [Google Scholar]
- Sirop, J.C.; Pradines, R.D. Topical Cosmetic Composition, Useful for Protecting Skin and Hair Against Sunlight, Contains an Extract From the Red Alga Polysiphonia lanosa. Patent No. FR2803200, 6 July 2001. [Google Scholar]
- Nakamura, E.; Kobayashi, J.; Abe, R. Mycosporine-Like Amino Acid. Patent No. JPS59137450, 1984. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Patent Title | Patent Number | References |
|---|---|---|
| Extracts of Aphanizomenon flos aquae and nutritional, cosmetic and pharmaceutical compositions containing the same | CN 101489527 | [217,218,219,220,221,222] |
| WO 2008000431 | ||
| CA 2656160 | ||
| MX 2009000137 | ||
| KR 1020090048399 | ||
| US 20100021493 | ||
| US 8337858 | ||
| EP 2032122 | ||
| Topical composition comprising transformed bacteria expressing a compound of interest | WO 2014025938 | [223,224] |
| US 20140044677 | ||
| US 20140044653 | ||
| US 20160000701 | ||
| US 9234204 | ||
| Topical formulations for UV protection | WO 2015195546 | [225] |
| Method for producing mycosporine-like amino acid using microbes | WO 2015174427 | [226] |
| Synthesis of UV absorbing compounds | WO 2014082124 | [227,228] |
| US 20150299124 | ||
| UV absorbing compounds, compositions comprising same and uses thereof | WO 2015006803 | [229,230] |
| US 20160244409 | ||
| Imino compounds as protecting agents against ultraviolet radiations | WO 2013181741 | [231,232] |
| US 20150152046 | ||
| Mycosporin-like amino acids, production method thereof, UV protecting agents and antioxidants | JP 2014227339 | [233] |
| Preparation method for laver mycosporine-like amino acids phorphyra-334 | CN 102659621 | [234] |
| Beaty product containing desert algae radiation-proof ingredient and natural medical whitening ingredient and preparation method thereof | CN 102764206 | [235] |
| Method for preparing UV screening nontoxic extract from red algae, and nontoxic sunscreen using same | WO 2011096628 | [236,237] |
| CN 102740869 | ||
| Topical composition | WO 2011158041 | [238] |
| Cosmetic sunscreen composition | GB 2472021 | [239] |
| Aphanizomenon flos aquae preparation, extracts and purified components thereof for the treatment of neurological, neurodegenerative and mood disorders | WO 2008000430 | [240,241,242] |
| US 20090311286 | ||
| EP 2046354 | ||
| Method for manufacturing non-toxic extract for blocking UV from red algae | KR 100969325 | [243] |
| Mycosporin-like amino acid derivative having glycosyl group and method for producing the same | JP 2009120562 | [244] |
| Sunscreen composition with extract of algae and lichens | ES 2317741 | [245] |
| Compositions comprising Porphyra and methods of making and using thereof | WO 2007144779 | [246,247] |
| US 20070220806 | ||
| EP 2001311 | ||
| Uso de aminoácido tipo micosporina (shinorine) en productos para prevención y tratamiento de eritema actínico, fotocarcinogénesis y fotoenvejecimiento | ES 2301426 | [248] |
| Uso de aminoácido tipo micosporina (porfira 334) en productos para prevención de procesos cancerígenos | ES 2301293 | [249] |
| Use of a mycosporin-type amino acid (porphyra 334) as an antioxidant | WO 2007026035 | [250] |
| Use of a mycosporin-type amino acid (M-gly) as an antioxidant | WO 2007026036 | [251] |
| Use of a mixture of mycosporin-type amino acids (asterin 330 + palythine) as an antioxidant | WO 2007026037 | [252] |
| Use of a mycosporin-type amino acid (shinorine) as an antioxidant | WO 2007026038 | [253] |
| Fibroblast growth promoter | JP 2007016004 | [254] |
| Cosmetic including natural substance having sun-screening function | CN 101061995 | [255] |
| Beta-glucuronidase inhibitors for use in deodorants and antiperspirants | US 20040234466 | [256,257] |
| US 7294330 | ||
| Amino-benzophenone UV filter formulations for the prevention of tanning | GB 2412866 | [258] |
| The utilization of natural pigments from lichens, cyanobacteria, fungi and plants for sun protection | WO 2003020236 | [259,260,261] |
| AU 2002329025 | ||
| EP 1424990 | ||
| US 20050129630 | ||
| Cosmetic skin care products and cosmetic agents for protecting skin against premature aging | EP 1473028 | [262] |
| Solar radiation protection composition | WO 2000024369 | [263,264,265] |
| EP 1123083 | ||
| US 6787147 | ||
| Personal care compositions | WO 2002039974 | [266,267] |
| EP 1341514 | ||
| Natural UV filters derived from pigments of lichens | IL 0200725 | [268] |
| Algal extracts containing amino acid analogs of mycosporin are useful as dermatological protecting agents against ultraviolet radiation | FR 2803201 | [211] |
| Topical cosmetic composition, useful for protecting skin and hair against sunlight, contains an extract from the red alga Polysiphonia lanosa | FR 2803200 | [269] |
| UV-absorbing compounds and compositions | WO 1990009995 | [214] |
| Sunscreen compositions and compounds for use therein | WO 1988002251 | [215] |
| Mycosporine-like amino acid | JPS 59137450 | [270] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrapusta, E.; Kaminski, A.; Duchnik, K.; Bober, B.; Adamski, M.; Bialczyk, J. Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Mar. Drugs 2017, 15, 326. https://doi.org/10.3390/md15100326
Chrapusta E, Kaminski A, Duchnik K, Bober B, Adamski M, Bialczyk J. Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Marine Drugs. 2017; 15(10):326. https://doi.org/10.3390/md15100326
Chicago/Turabian StyleChrapusta, Ewelina, Ariel Kaminski, Kornelia Duchnik, Beata Bober, Michal Adamski, and Jan Bialczyk. 2017. "Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients" Marine Drugs 15, no. 10: 326. https://doi.org/10.3390/md15100326
APA StyleChrapusta, E., Kaminski, A., Duchnik, K., Bober, B., Adamski, M., & Bialczyk, J. (2017). Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Marine Drugs, 15(10), 326. https://doi.org/10.3390/md15100326