
Biochemical and Structural Insights into a Novel Thermostable β-1,3-Galactosidase from Marinomonas sp. BSi20414

Abstract
:1. Introduction
2. Results
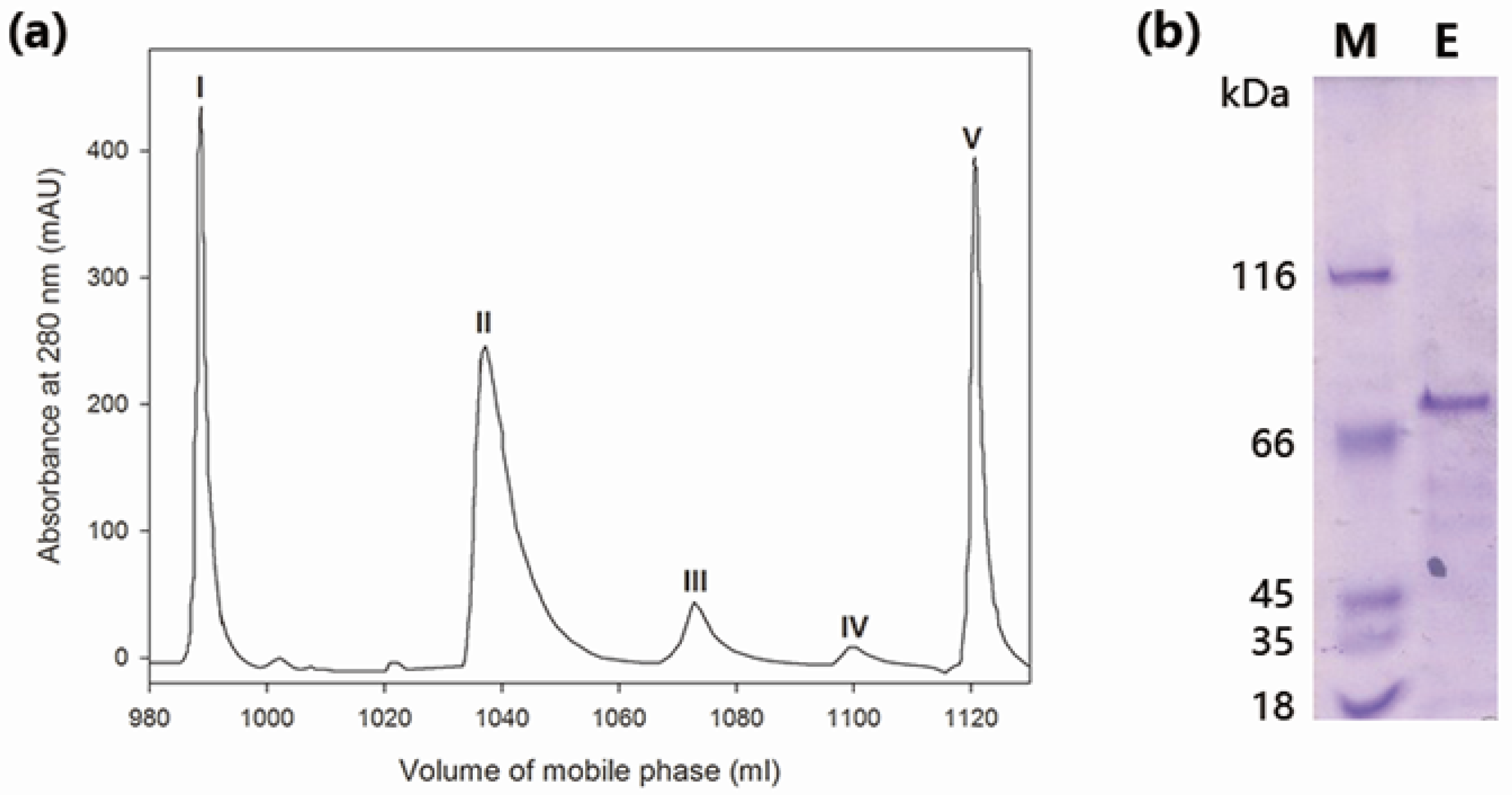
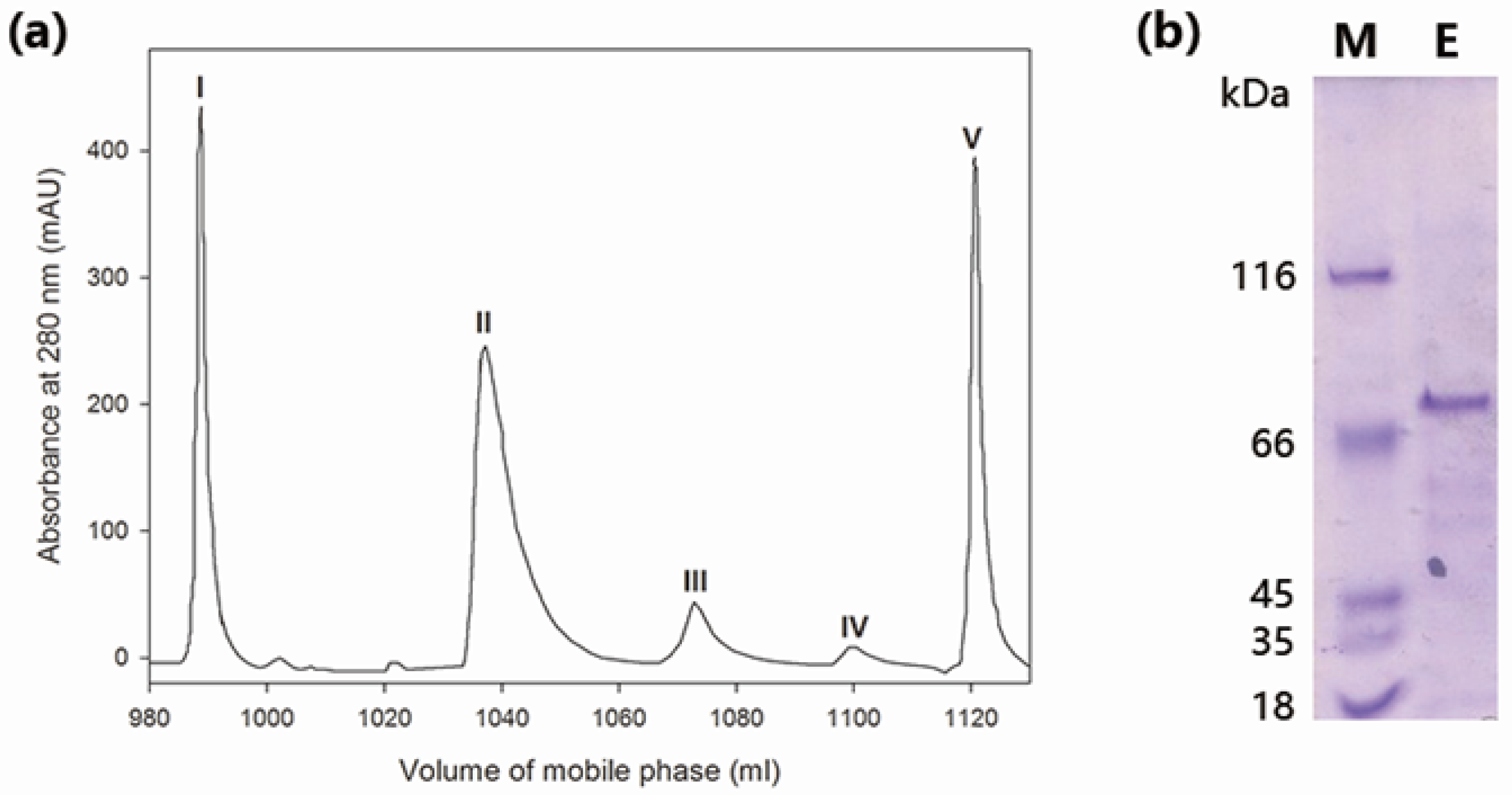
2.1. Purification of Wild-Type MaBGA
2.2. Enzymatic Characterization of MaBGA
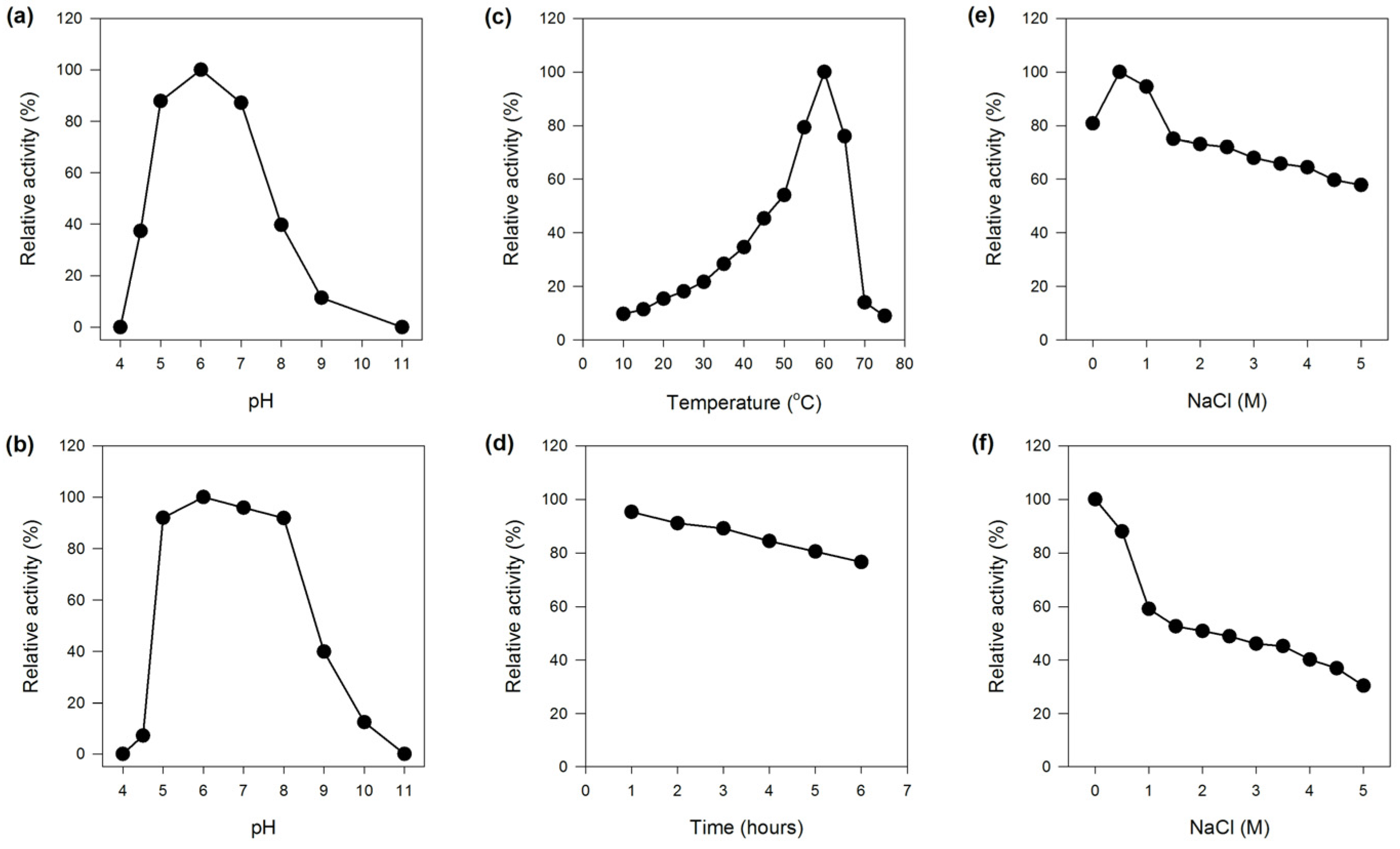
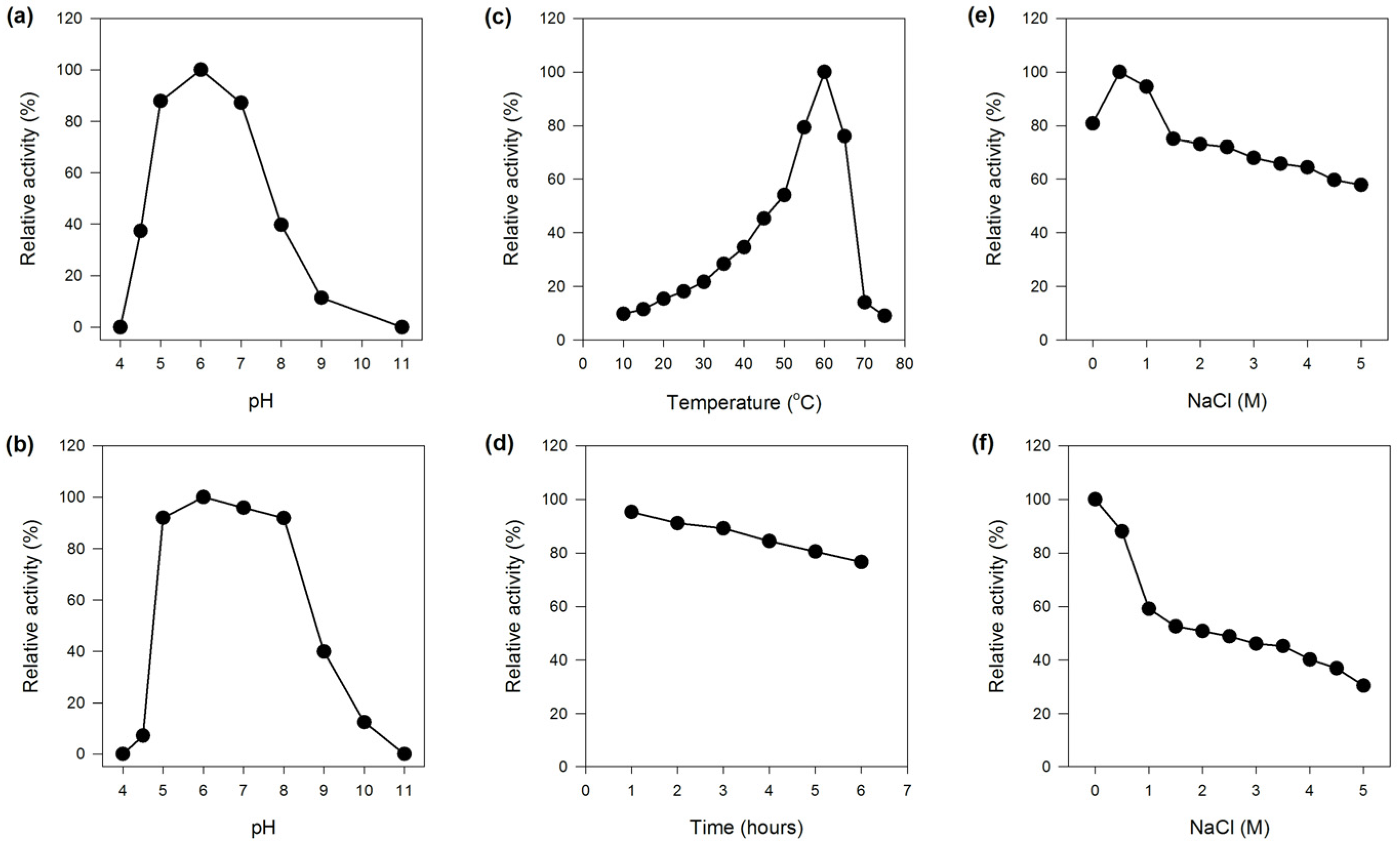
2.2.1. Effect of pH on the Activity and Stability of MaBGA
2.2.2. Effect of Temperature on the Activity and Stability of MaBGA
2.2.3. Effect of NaCl on the Activity and Stability of MaBGA
2.2.4. Effects of Metal Ions and Chemicals on the Activity of MaBGA
2.2.5. Substrate Specificity and Steady-State Kinetic Analysis
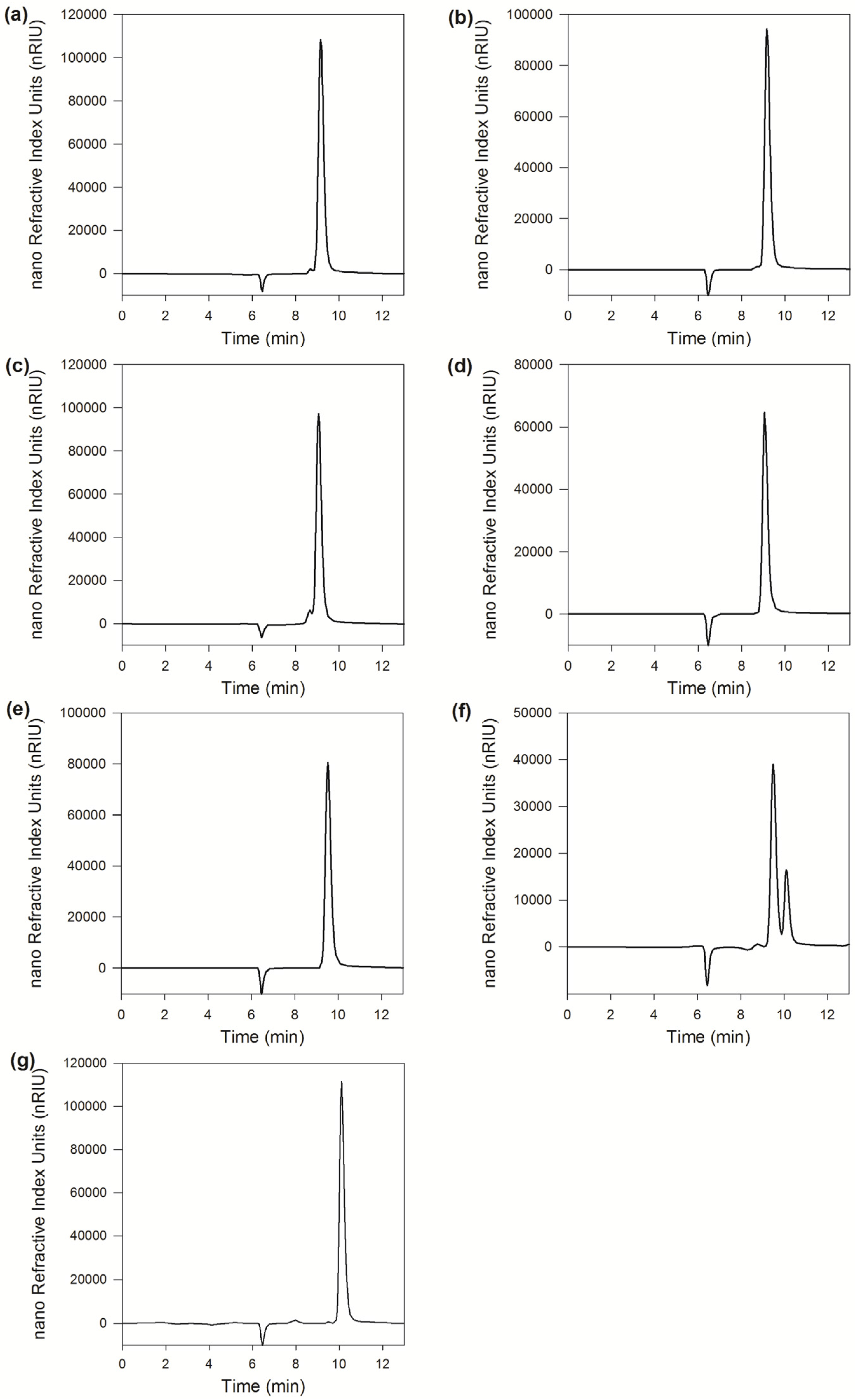
2.2.6. Linkage Selectivity Analysis
2.3. Gene Cloning and Sequence Analysis
2.3.1. Gene Cloning
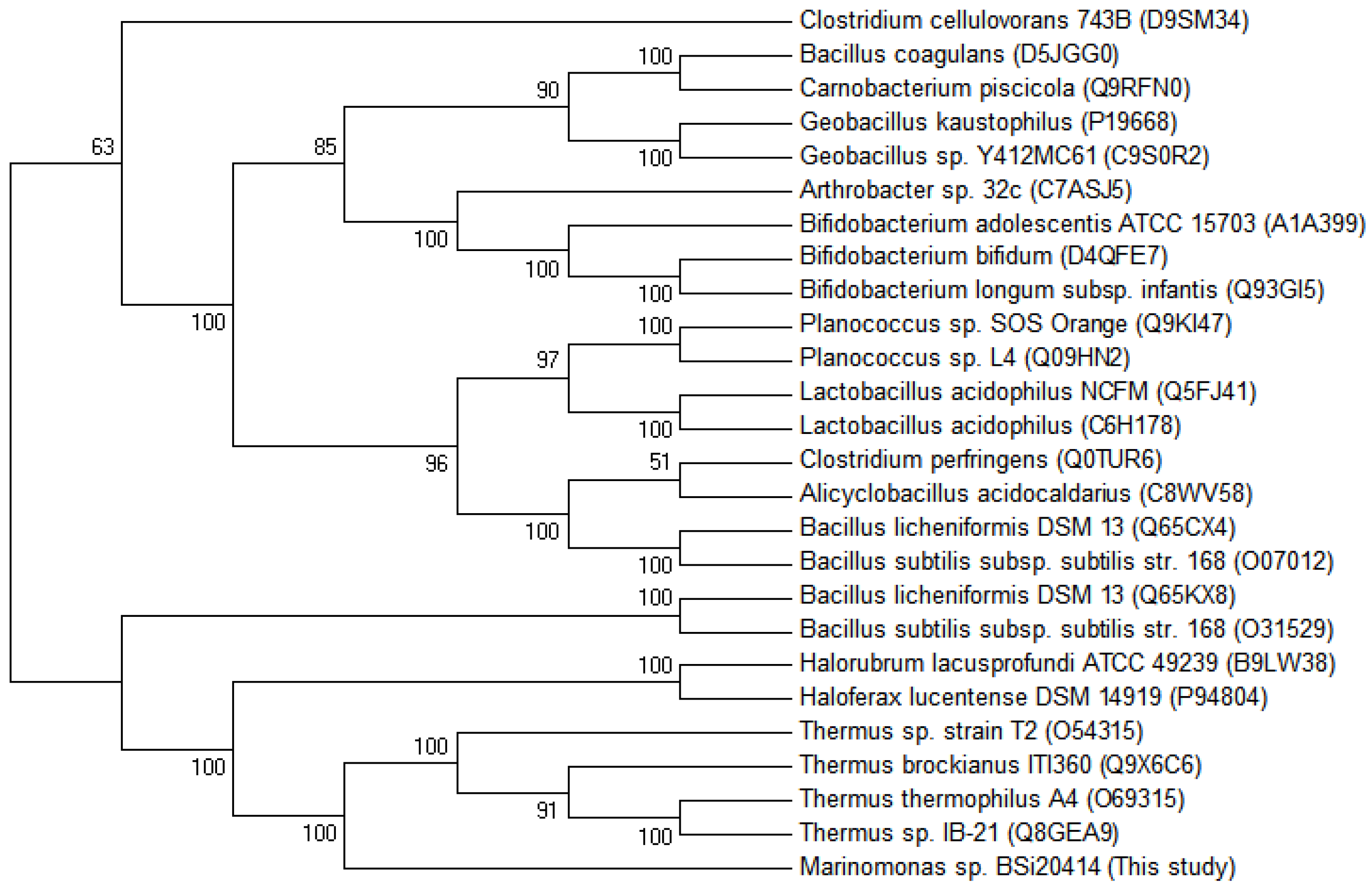
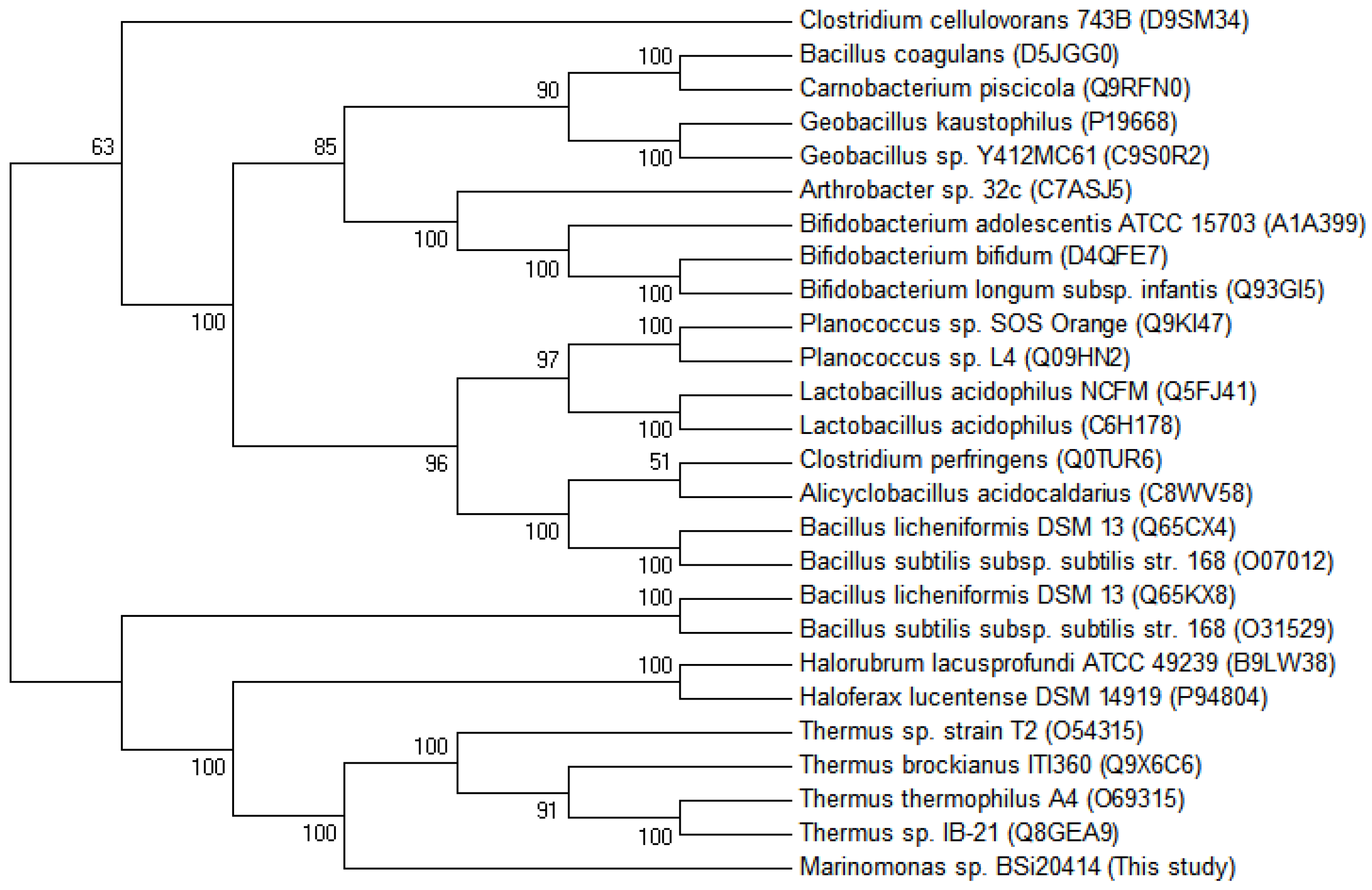
2.3.2. Sequence Analysis
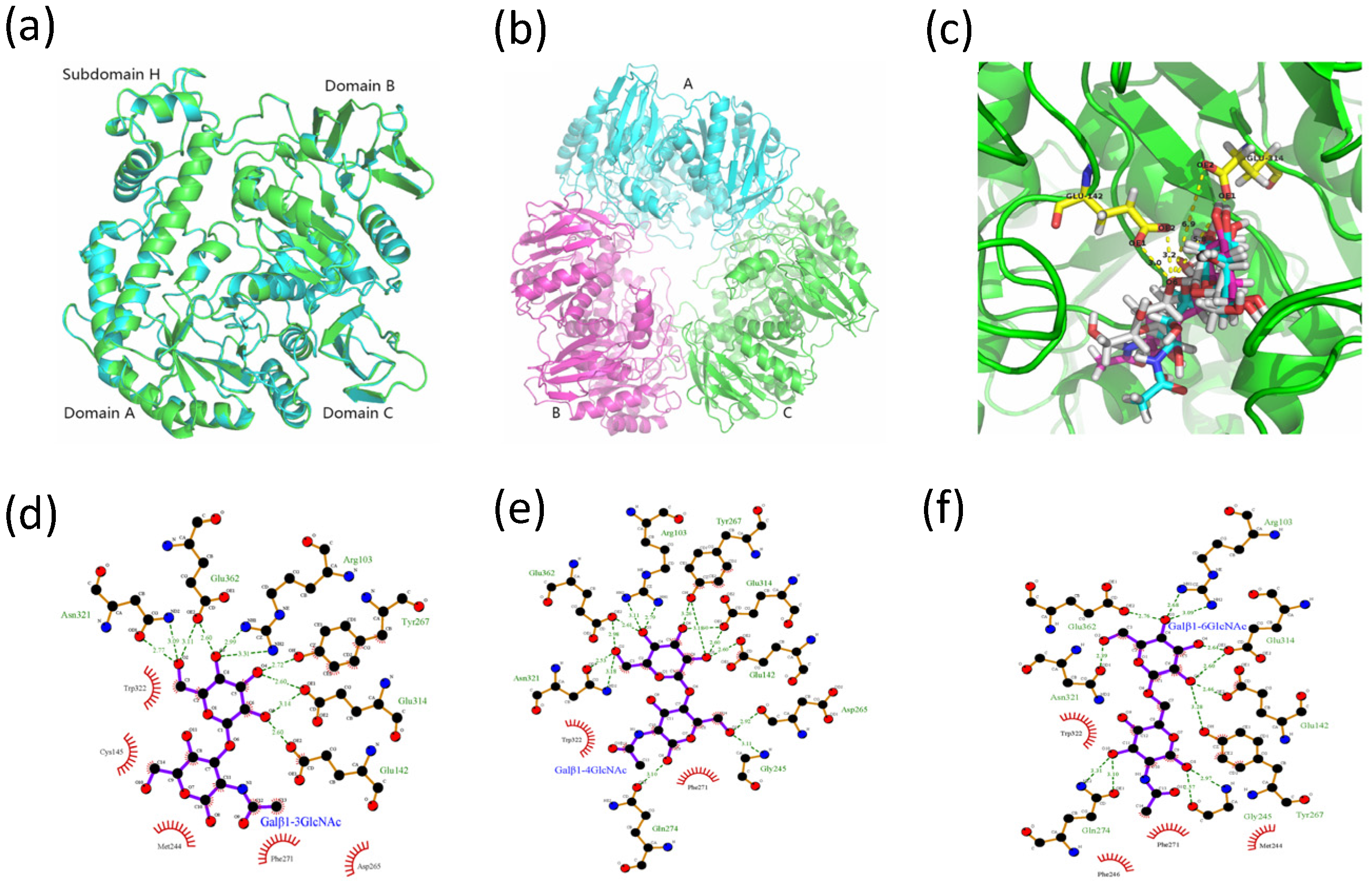
2.4. Structural Analysis of MaBGA
2.4.1. Homology Modelling
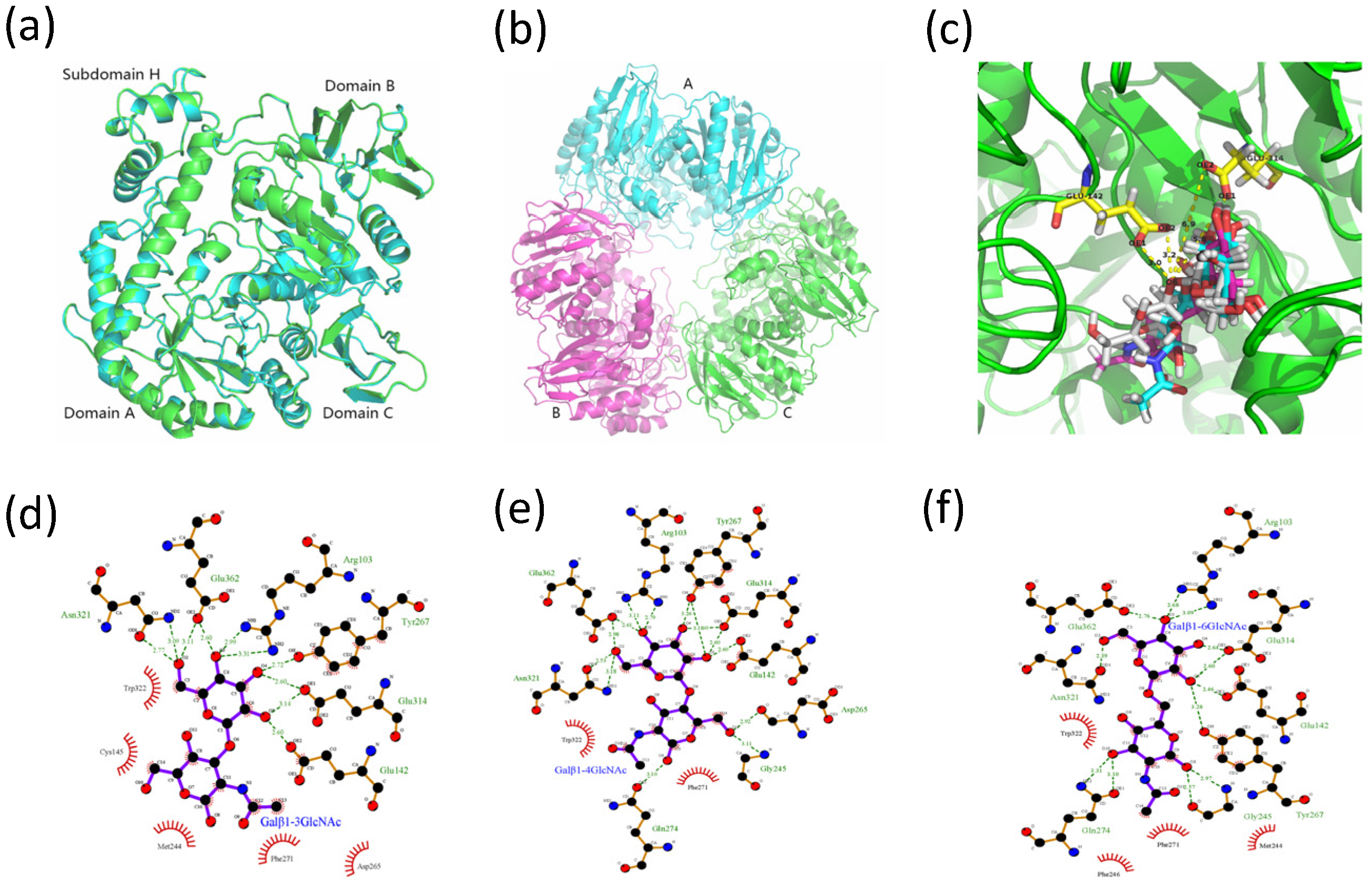
2.4.2. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Culture Conditions
4.2. Purification of Wild-Type MaBGA
4.3. SDS-PAGE Analysis
4.4. β-galactosidase Activity Assay
4.5. Effect of pH on the Activity and Stability of MaBGA
4.6. Effect of Temperature on the Activity and Stability of MaBGA
4.7. Effect of NaCl on the Activity and Stability of MaBGA
4.8. Effect of Metal Ions and Chemicals on the Activity of MaBGA
4.9. Substrate Specificity
4.10. Steady-State Kinetic Analysis
4.11. Linkage Selectivity Analysis
4.12. Gene Cloning and Sequence Analysis
4.13. Homology Modelling and Molecular Docking Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Juers, D.H.; Matthews, B.W.; Huber, R.E. LacZ β-galactosidase: Structure and function of an enzyme of historical and molecular biological importance. Protein Sci. 2012, 21, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Guimaraes, P.M.; Domingues, L. Recombinant microbial systems for improved β-galactosidase production and biotechnological applications. Biotechnol. Adv. 2011, 29, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Park, A.R.; Oh, D.K. Galacto-oligosaccharide production using microbial β-galactosidase: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B.; Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 1997, 7, 637–644. [Google Scholar] [CrossRef]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (cazy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 1995, 3, 853–859. [Google Scholar] [CrossRef]
- Adam, A.C.; Rubio-Texeira, M.; Polaina, J. Lactose: The milk sugar from a biotechnological perspective. BFSN 2005, 44, 553–557. [Google Scholar] [CrossRef]
- Singh, A.K.; Pluvinage, B.; Higgins, M.A.; Dalia, A.B.; Woodiga, S.A.; Flynn, M.; Lloyd, A.R.; Weiser, J.N.; Stubbs, K.A.; Boraston, A.B.; et al. Unravelling the multiple functions of the architecturally intricate streptococcus pneumoniae β-galactosidase, BgaA. PLoS Pathog. 2014, 10, e1004364. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhang, F.; Zhang, H.; Hao, L.; Gong, X.; Geng, M.; Cao, M.; Zheng, F.; Zhu, J.; Pan, X.; et al. The β-galactosidase (BgaC) of the zoonotic pathogen streptococcus suis is a surface protein without the involvement of bacterial virulence. Sci. Rep. 2014, 4, 4140. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Wang, L.; Jiang, Y.L.; Bai, X.H.; Chu, J.; Li, Q.; Yu, G.; Liang, Q.L.; Zhou, C.Z.; Chen, Y. Structural insights into the substrate specificity of streptococcus pneumoniae β(1,3)-galactosidase BgaC. J. Biol. Chem. 2012, 287, 22910–22918. [Google Scholar] [CrossRef] [PubMed]
- Solomon, H.V.; Tabachnikov, O.; Lansky, S.; Salama, R.; Feinberg, H.; Shoham, Y.; Shoham, G. Structure-function relationships in Gan42B, an intracellular GH42 β-galactosidase from geobacillus stearothermophilus. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Solomon, H.V.; Tabachnikov, O.; Feinberg, H.; Govada, L.; Chayen, N.E.; Shoham, Y.; Shoham, G. Crystallization and preliminary crystallographic analysis of ganb, a GH42 intracellular β-galactosidase from geobacillus stearothermophilus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.K.; Cho, K.K.; Ahn, J.K.; Bok, J.D.; Kang, S.H.; Woo, J.H.; Lee, H.G.; You, S.K.; Choi, Y.J. Three forms of thermostable lactose-hydrolase from Thermus sp. IB-21: Cloning, expression, and enzyme characterization. J. Biotechnol. 2005, 116, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Vian, A.; Carrascosa, A.V.; Garcia, J.L.; Cortes, E. Structure of the β-galactosidase gene from thermus sp. Strain T2: Expression in escherichia coli and purification in a single step of an active fusion protein. Appl. Environ. Microbiol. 1998, 64, 2187–2191. [Google Scholar] [PubMed]
- Ohtsu, N.; Motoshima, H.; Goto, K.; Tsukasaki, F.; Matsuzawa, H. Thermostable β-galactosidase from an extreme thermophile, Thermus sp. A4: Enzyme purification and characterization, and gene cloning and sequencing. Biosci. Biotechnol. Biochem. 1998, 62, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Karan, R.; Capes, M.D.; DasSarma, P.; DasSarma, S. Cloning, overexpression, purification, and characterization of a polyextremophilic β-galactosidase from the antarctic haloarchaeon halorubrum lacusprofundi. BMC Biotechnol. 2013, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.L.; Scopes, R.K.; Moritz, R.L.; Simpson, R.J.; Englert, C.; Pfeifer, F.; Dyall-Smith, M.L. Purification and analysis of an extremely halophilic β-galactosidase from haloferax alicantei. Biochim. Biophys. Acta 1997, 1337, 276–286. [Google Scholar] [CrossRef]
- Maksimainen, M.; Paavilainen, S.; Hakulinen, N.; Rouvinen, J. Structural analysis, enzymatic characterization, and catalytic mechanisms of β-galactosidase from bacillus circulans sp. Alkalophilus. FEBS J. 2012, 279, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Golyshina, O.; Beloqui, A.; Golyshin, P.N. Mining enzymes from extreme environments. Curr. Opin. Microbiol. 2007, 10, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kwon, K.K.; Kang, S.G.; Cha, S.-S.; Kim, S.-J.; Lee, J.-H. Approaches for novel enzyme discovery from marine environments. Curr. Opin. Biotechnol. 2010, 21, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Beloqui, A.; Timmis, K.N.; Golyshin, P.N. Metagenomics for mining new genetic resources of microbial communities. J. Mol. Microbiol. Biotechnol. 2008, 16, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wang, Y.; Sun, K.; Yu, Y.; Chen, B. Preliminary studies on the screening, identification and optimum fermentative conditions of a strain Marinomonas sp. Bsi20414 isolated from arctic sea ice producing β-galactosidase. Chin. J. Polar Res. 2011, 108–114. [Google Scholar]
- Hidaka, M.; Fushinobu, S.; Ohtsu, N.; Motoshima, H.; Matsuzawa, H.; Shoun, H.; Wakagi, T. Trimeric crystal structure of the glycoside hydrolase family 42 β-galactosidase from thermus thermophilus a4 and the structure of its complex with galactose. J. Mol. Biol. 2002, 322, 79–91. [Google Scholar] [CrossRef]
- Fridjonsson, O.; Watzlawick, H.; Gehweiler, A.; Rohrhirsch, T.; Mattes, R. Cloning of the gene encoding a novel thermostable α-galactosidase from thermus brockianus ITI360. Appl. Environ. Microbiol. 1999, 65, 3955–3963. [Google Scholar] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. Swiss-model: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Chivian, D.; Baker, D. Protein structure prediction and analysis using the robetta server. Nucleic Acids Res. 2004, 32, W526–W531. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Sali, A. Comparative protein structure modeling using Modeller. Curr. Protoc. Bioinform. 2014. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-tasser suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Wiederstein, M.; Sippl, M.J. Prosa-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. Procheck: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; van der Spoel, D. Gromacs 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and autodocktools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Depeint, F.; Tzortzis, G.; Vulevic, J.; I’Anson, K.; Gibson, G.R. Prebiotic evaluation of a novel galactooligosaccharide mixture produced by the enzymatic activity of bifidobacterium bifidum ncimb 41171, in healthy humans: A randomized, double-blind, crossover, placebo-controlled intervention study. Am. J. Clin. Nutr. 2008, 87, 785–791. [Google Scholar] [PubMed]
- Legler, G. Glycoside hydrolases: Mechanistic information from studies with reversible and irreversible inhibitors. Adv. Carbohydr. Chem. Biochem. 1990, 48, 319–384. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.G.; Whittier, R.F. Thermal asymmetric interlaced pcr: Automatable amplification and sequencing of insert end fragments from p1 and yac clones for chromosome walking. Genomics 1995, 25, 674–681. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Gouet, P.; Robert, X.; Courcelle, E. Espript/endscript: Extracting and rendering sequence and 3d information from atomic structures of proteins. Nucleic Acids Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Recovery (%) | Purification Fold |
|---|---|---|---|---|---|
| Cell lysate | 162.54 | 1818.18 | 11.19 | 100 | 1 |
| Ammonium sulfate precipitation | 29.9 | 927.27 | 31.01 | 51 | 2.77 |
| HiTrap DEAE FF | 1.01 | 91.80 | 90.89 | 9.9 | 8.12 |
| Metal Ions | Relative Activity (%) | Chemicals | Relative Activity (%) |
|---|---|---|---|
| K+ | 96 | EDTA | 98 |
| Na+ | 95 | l-Cysteine | 110 |
| Fe2+ | 211 | l-Glutathion | 103 |
| Mn2+ | 98 | Dithiotreitol | 106 |
| Mg2+ | 89 | ||
| Co2+ | 88 | ||
| Ni2+ | 76 | ||
| Zn2+ | 74 |
| Substrate | Relative Activity (%) |
|---|---|
| o-nitrophenyl-β-d-galactopyranoside | 100 |
| p-nitrophenyl-β-d-galactopyranoside | 422 |
| p-nitrophenyl-β-d-glucopyranoside | <1 |
| p-nitrophenyl-β-d-xylopyranoside | <1 |
| p-nitrophenyl-β-d-lactopyranoside | <1 |
| p-nitrophenyl-β-d-glucuronide | <1 |
| p-nitrophenyl-a-d-galactopyranoside | <1 |
| p-nitrophenyl-β-l-arabinopyranoside | <1 |
| p-nitrophenyl-β-d-cellobioside | <1 |
| Model | Z-Score 1 | Ramachandran Plot 2 | |||
|---|---|---|---|---|---|
| Most Favored (%) | Additional Allowed (%) | Generously Allowed (%) | Disallowed (%) | ||
| Template (4oif) | −12.19 | 88.3 | 10.7 | 0.6 | 0.3 |
| Robetta | −10.19 | 87.9 | 10.2 | 1.6 | 0.4 |
| Template (1kwk) | −12.24 | 90.6 | 8.2 | 0.7 | 0.4 |
| SWISS-MODEL | −9.93 | 89.7 | 8.1 | 1.5 | 0.7 |
| MODELLER | −10.08 | 91.9 | 6.8 | 0.4 | 0.9 |
| Template (1kwg) | −12.27 | 91.2 | 7.9 | 0.6 | 0.4 |
| I-TASSER | −10.14 | 79.1 | 15.8 | 3.2 | 1.9 |
| Primers | Sequence (5′ to 3′) | Purpose |
|---|---|---|
| F1 | GCNTGGGGNAAYGTNTTYT | Degenerated PCR |
| F2 | TNTGGACNTGGGARGCNTT | Degenerated PCR |
| F3 | GGARCARCARCCNGGNCCNGT | Degenerated PCR |
| R1 | CCARCANGCRTCRTARTCRAA | Degenerated PCR |
| R2 | RAANGCYTCCCANGTCCA | Degenerated PCR |
| R3 | GGRTTRTGNGGNGCCARTT | Degenerated PCR |
| A208 | TGGATHATGGAGGAGCCC | Degenerated PCR |
| A76 | CGGGACCTGGTGCAYAAYTAY | Degenerated PCR |
| A195 | CAYAAYTAYATGGGCTTCTTC | Degenerated PCR |
| B1 | CAGACCCAGAACGAGTAYKGN | Degenerated PCR |
| C1 | GCACCACAAGTACCACCARGA | Degenerated PCR |
| C2 | GTYCTRDWNCTGCACCGGCCG | Degenerated PCR |
| U1 | CCGTAAAGAATCCCATGAGT | DNA Walking (1st-round upstream) |
| D1 | GGACATTTTGCGTGCG | DNA Walking (1st-round downstream) |
| U2 | AACGCTGAAAGTCCAACCCGAT | DNA Walking (2nd-round upstream) |
| D2 | GGACACTTATCCGCTGGGTTT | DNA Walking (2nd-round downstream) |
| U3 | GATTGGCTTCGGTCACGGT | DNA Walking (3rd-round upstream) |
| D3 | CCCGATTTTGGTGCTTTTCA | DNA Walking (3rd-round downstream) |
| DW-ACP 1 | ACP-AGGTC | DNA Walking (1st-round) |
| DW-ACP 2 | ACP-TGGTC | DNA Walking (1st-round) |
| DW-ACP 3 | ACP-GGGTC | DNA Walking (1st-round) |
| DW-ACP 4 | ACP-CGGTC | DNA Walking (1st-round) |
| DW-ACP N | ACPN-GGTC | DNA Walking (2nd-round) |
| Uni-primer | TCACAGAAGTATGCCAAGCGA | DNA Walking (3rd-round) |
| MaBGA-F | CGGAATTCAAGTTAGGTGTATGTTACTACCCAG | Full-length amplification |
| MaBGA-R | GTTCGCGCTCGAGGATTTCTTGCCAAATGGC | Full-length amplification |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, H.; Zeng, Q.; Zhou, L.; Yu, Y.; Chen, B. Biochemical and Structural Insights into a Novel Thermostable β-1,3-Galactosidase from Marinomonas sp. BSi20414. Mar. Drugs 2017, 15, 13. https://doi.org/10.3390/md15010013
Ding H, Zeng Q, Zhou L, Yu Y, Chen B. Biochemical and Structural Insights into a Novel Thermostable β-1,3-Galactosidase from Marinomonas sp. BSi20414. Marine Drugs. 2017; 15(1):13. https://doi.org/10.3390/md15010013
Chicago/Turabian StyleDing, Haitao, Qian Zeng, Lili Zhou, Yong Yu, and Bo Chen. 2017. "Biochemical and Structural Insights into a Novel Thermostable β-1,3-Galactosidase from Marinomonas sp. BSi20414" Marine Drugs 15, no. 1: 13. https://doi.org/10.3390/md15010013
APA StyleDing, H., Zeng, Q., Zhou, L., Yu, Y., & Chen, B. (2017). Biochemical and Structural Insights into a Novel Thermostable β-1,3-Galactosidase from Marinomonas sp. BSi20414. Marine Drugs, 15(1), 13. https://doi.org/10.3390/md15010013