
Marine Natural Products from New Caledonia—A Review




Abstract
:
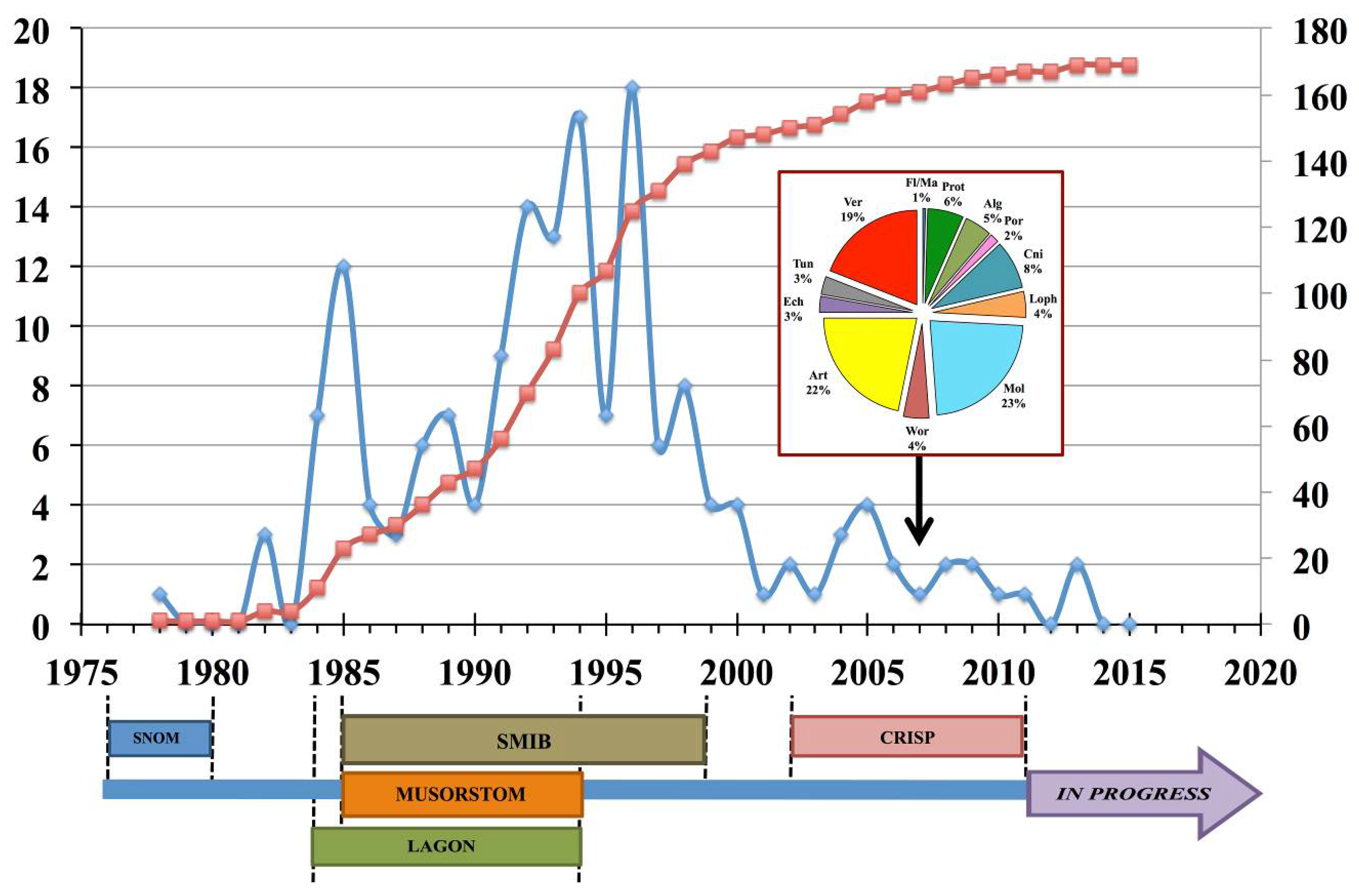
1. Introduction
2. Taxonomy
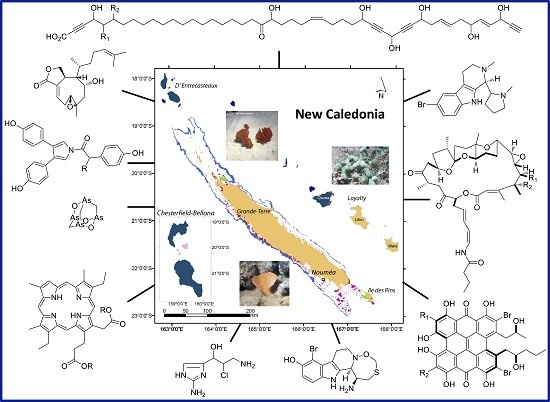
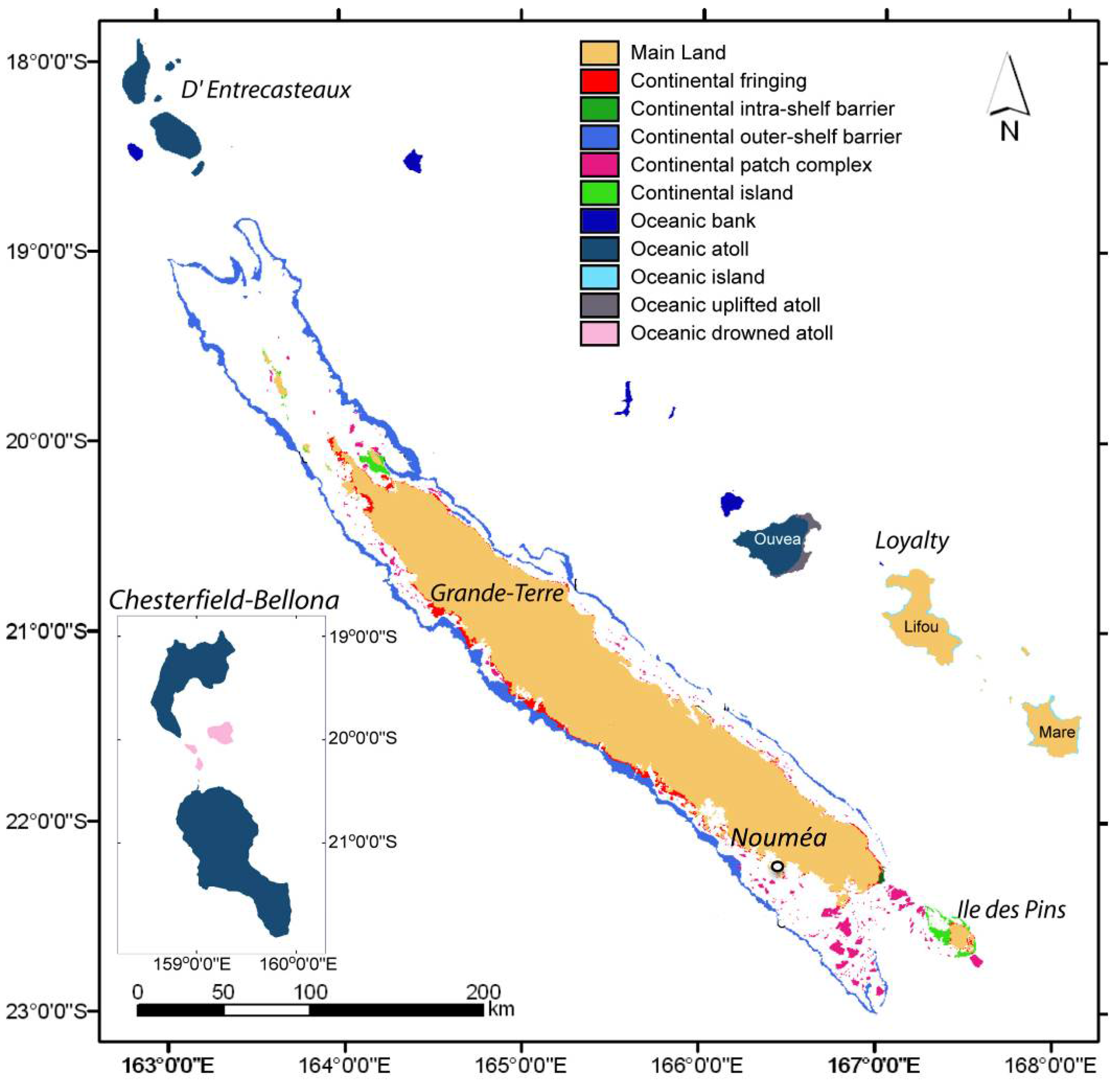
2.1. Sample Collection Sites
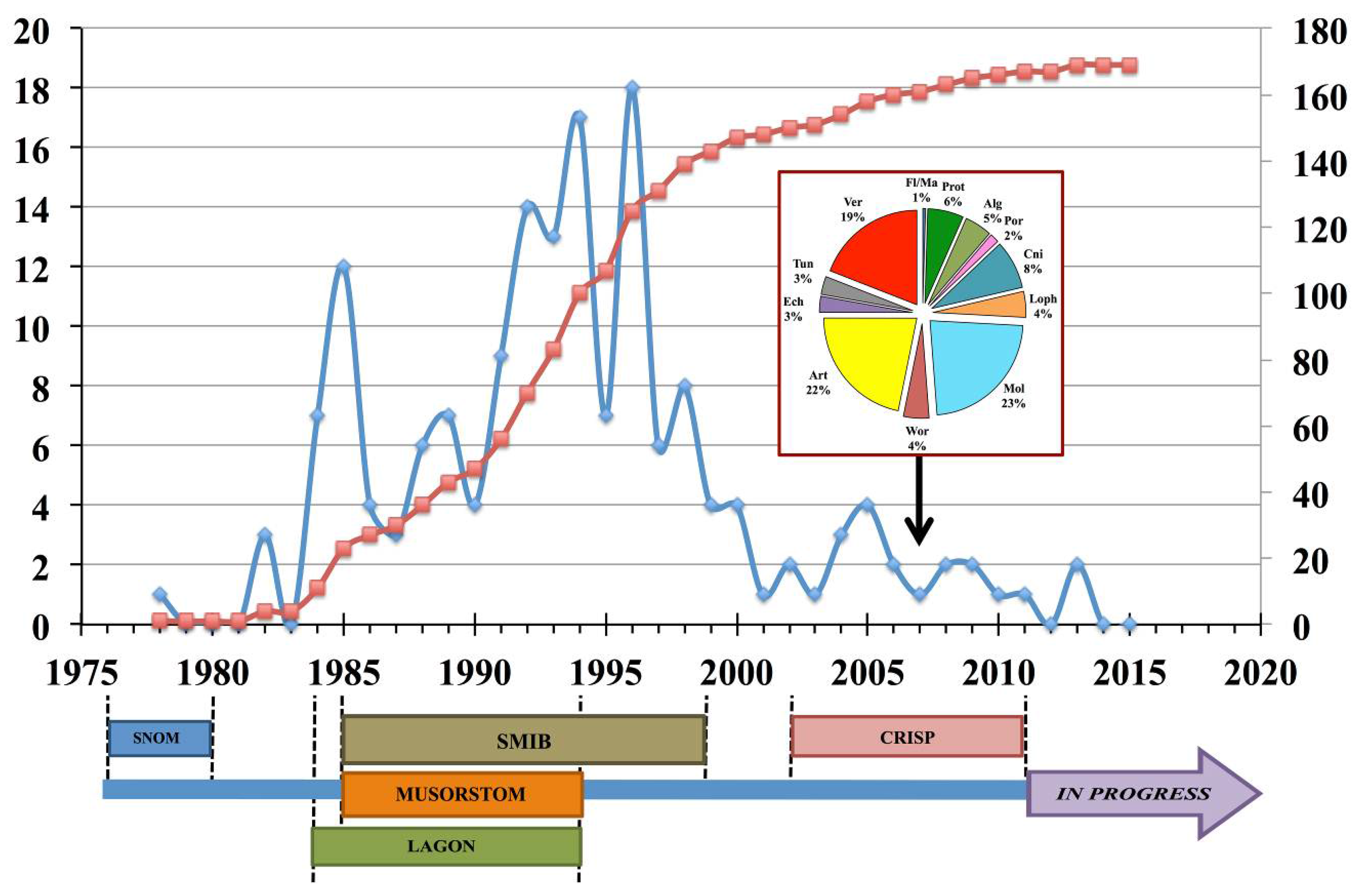
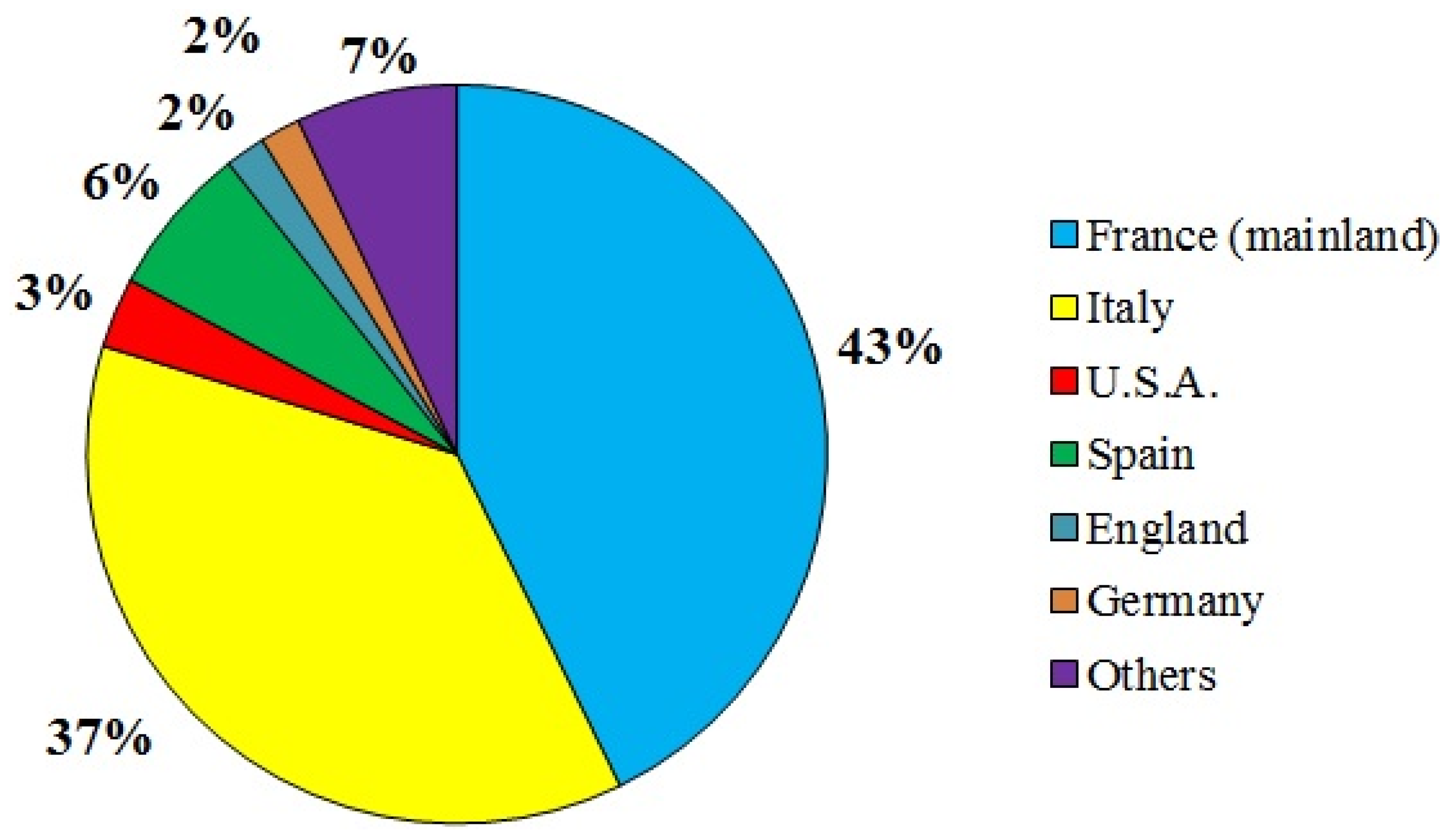
2.2. Biological Material Sampling
3. Chemistry
3.1. Biological Material
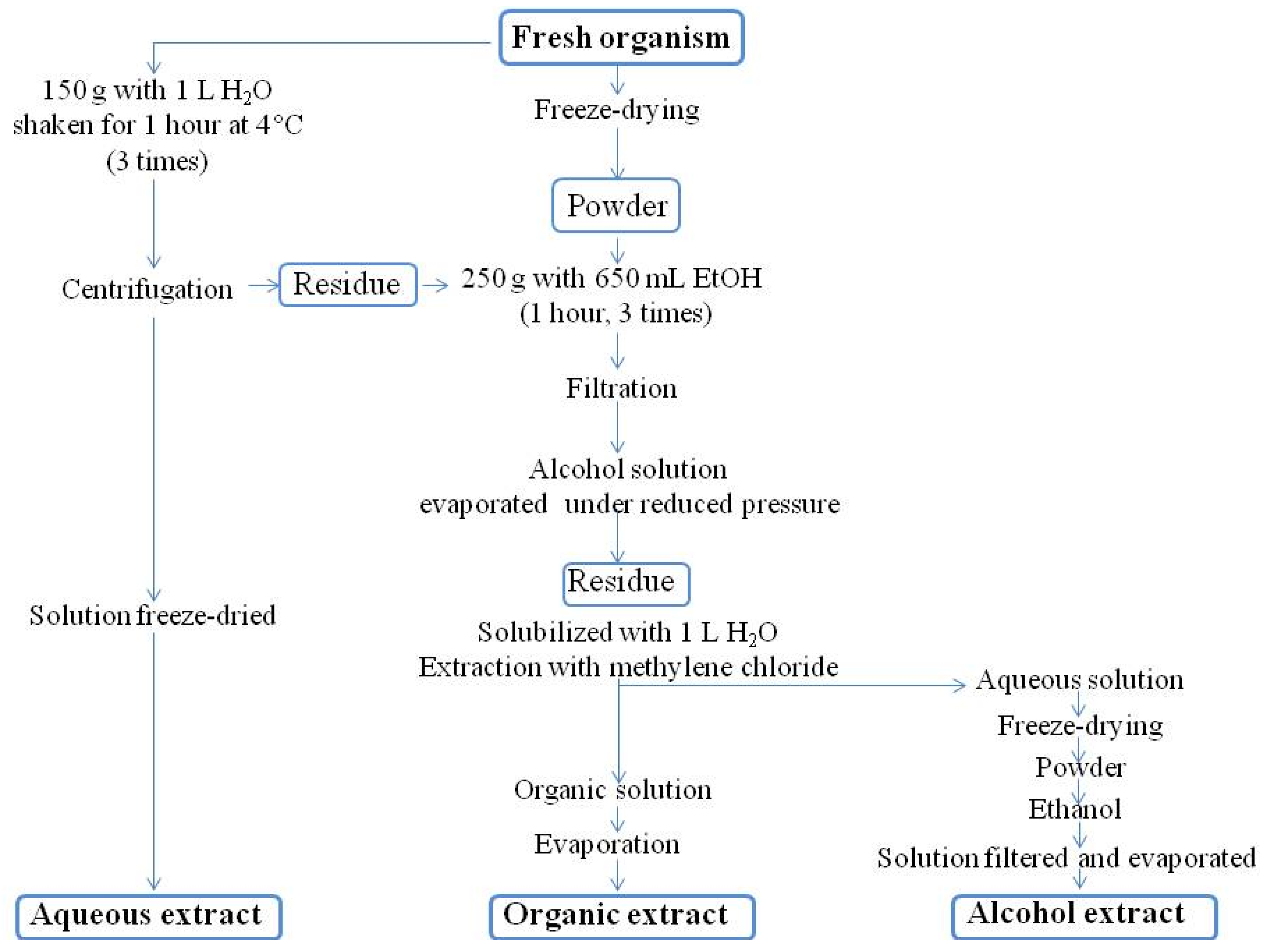
3.2. Conditioning Samples for Chemistry
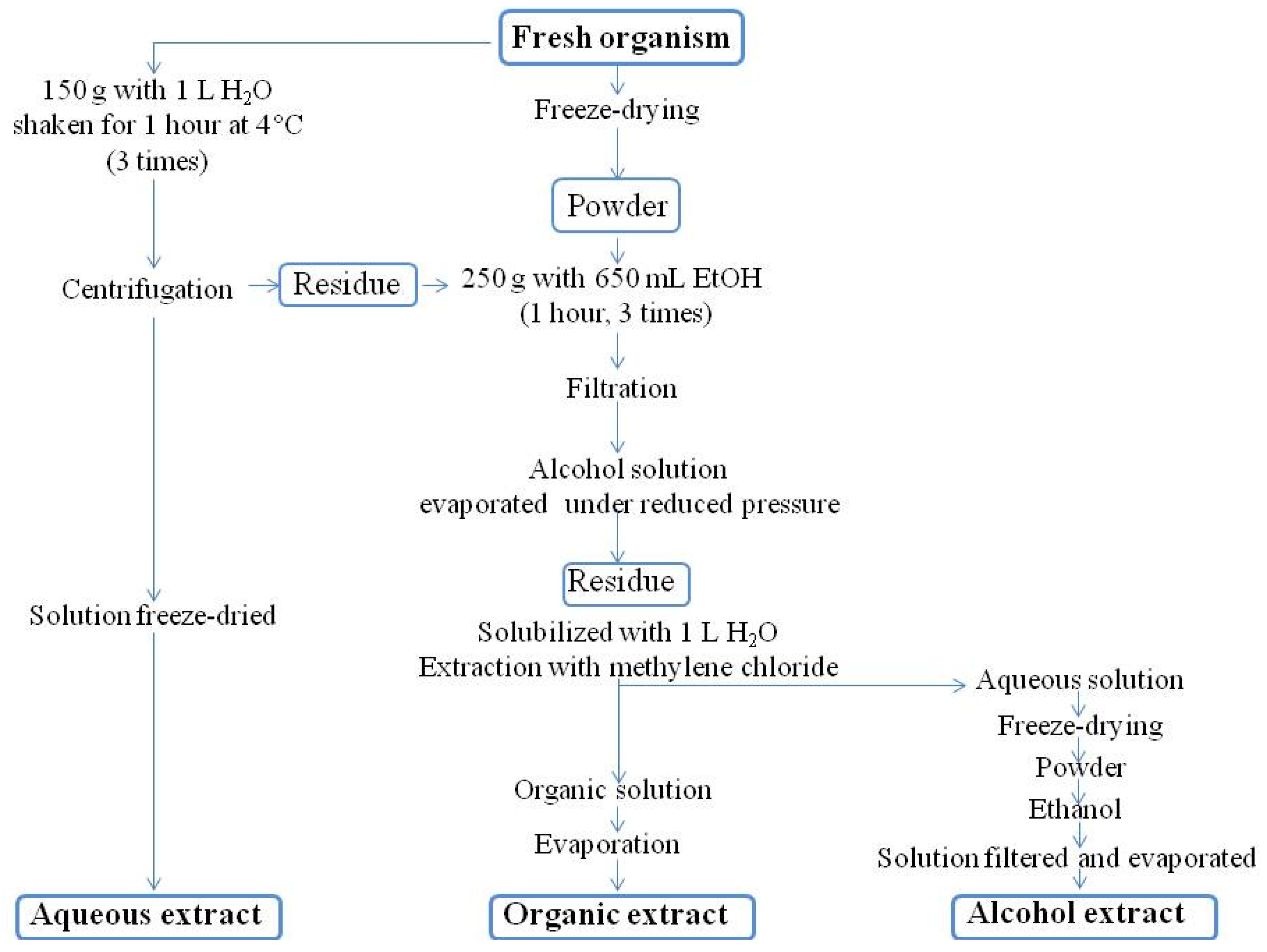
3.3. Extraction
3.3.1. Routine Procedure
3.3.2. Peptide Protease Inhibitors
3.4. Separation, Purification
3.5. Structural Analysis
3.6. Chemical Databases
4. Biological Activities
4.1. Preliminary Testing
4.1.1. Brine Shrimp Toxicity Assay
4.1.2. Mosquito Fish Toxicity Assay
4.1.3. Fertilization of Sea Urchin Eggs
4.1.4. Anti-Serpin Activity
4.2. Further Biological Testing
5. Natural Products by Taxon
5.1. Porifera (Sponges)
5.1.1. General Comments
5.1.2. Porifera Success Stories
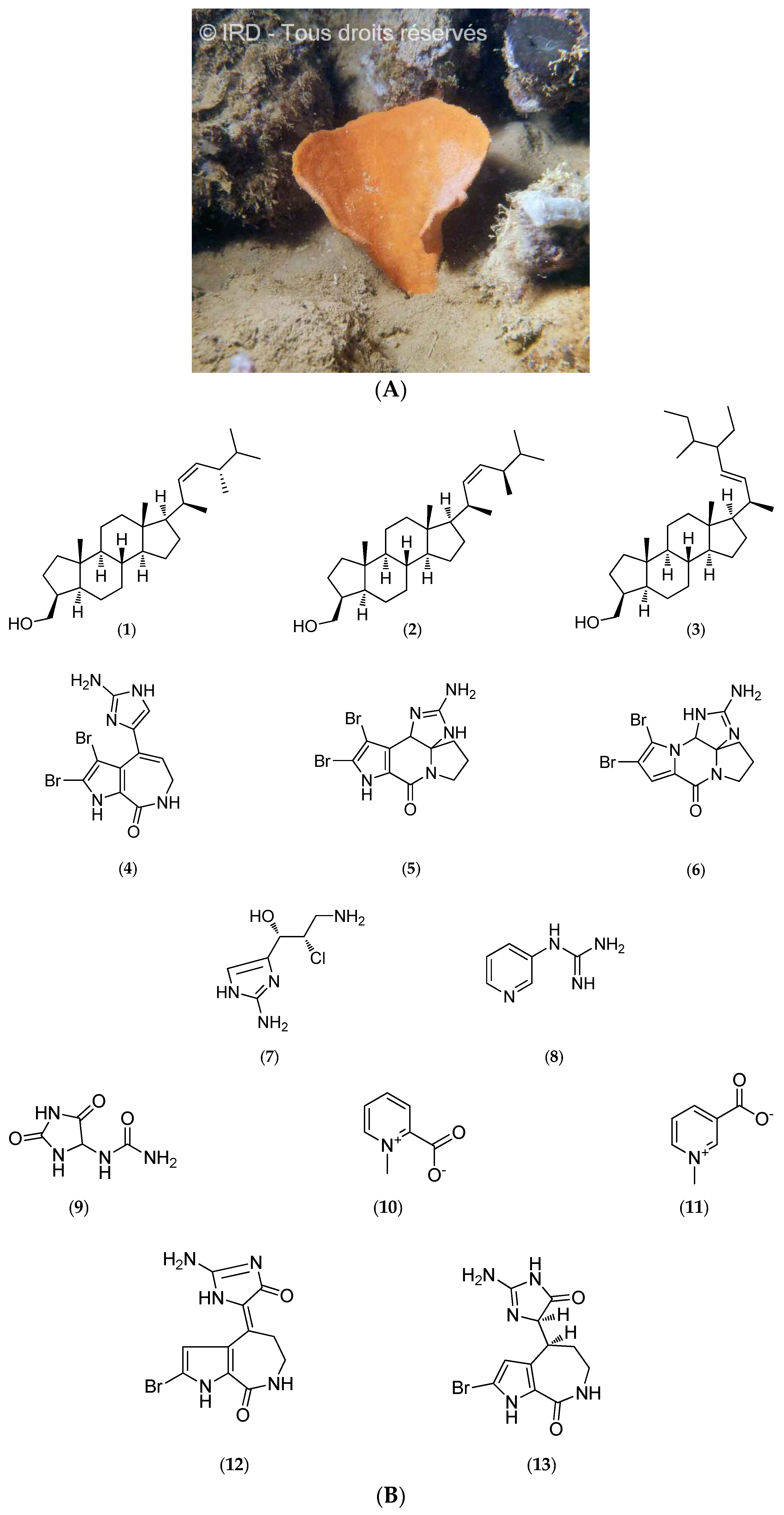
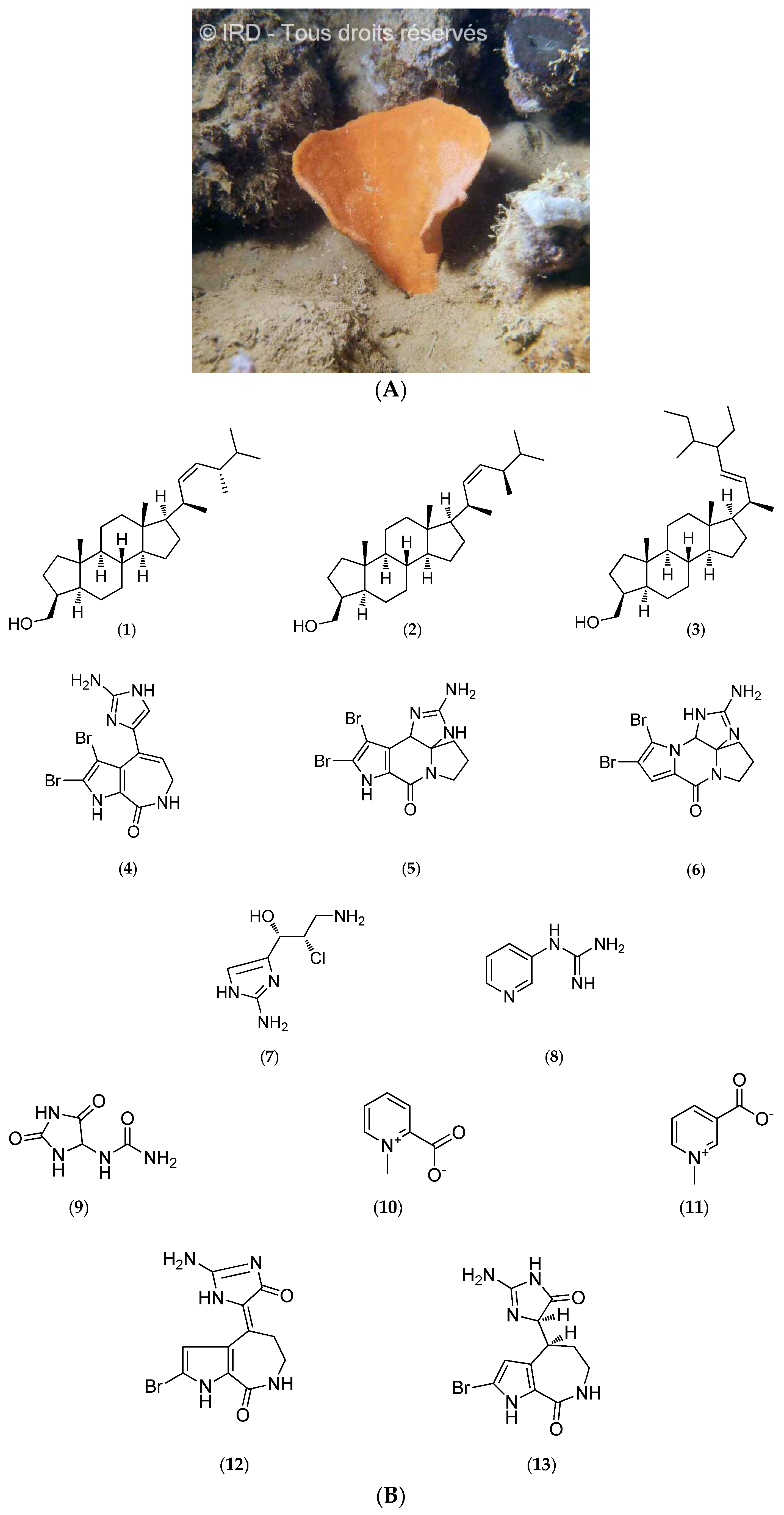
Cymbastela Cantharella
Echinochalina Bargibanti
Dendrilla sp.
Niphates sp.
Corallistes sp.
5.2. Ascidians
5.2.1. General Comments
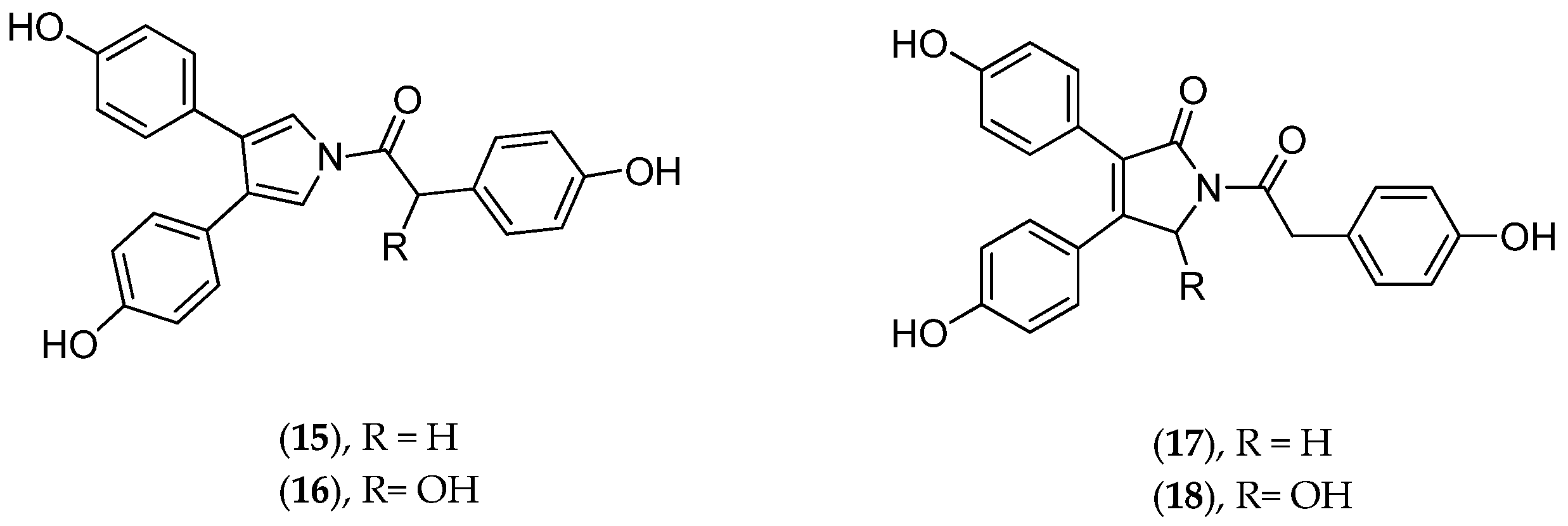
5.2.2. Lissoclinum bistratum
5.3. Cnidaria
5.3.1. Alcyonarians
General Comments
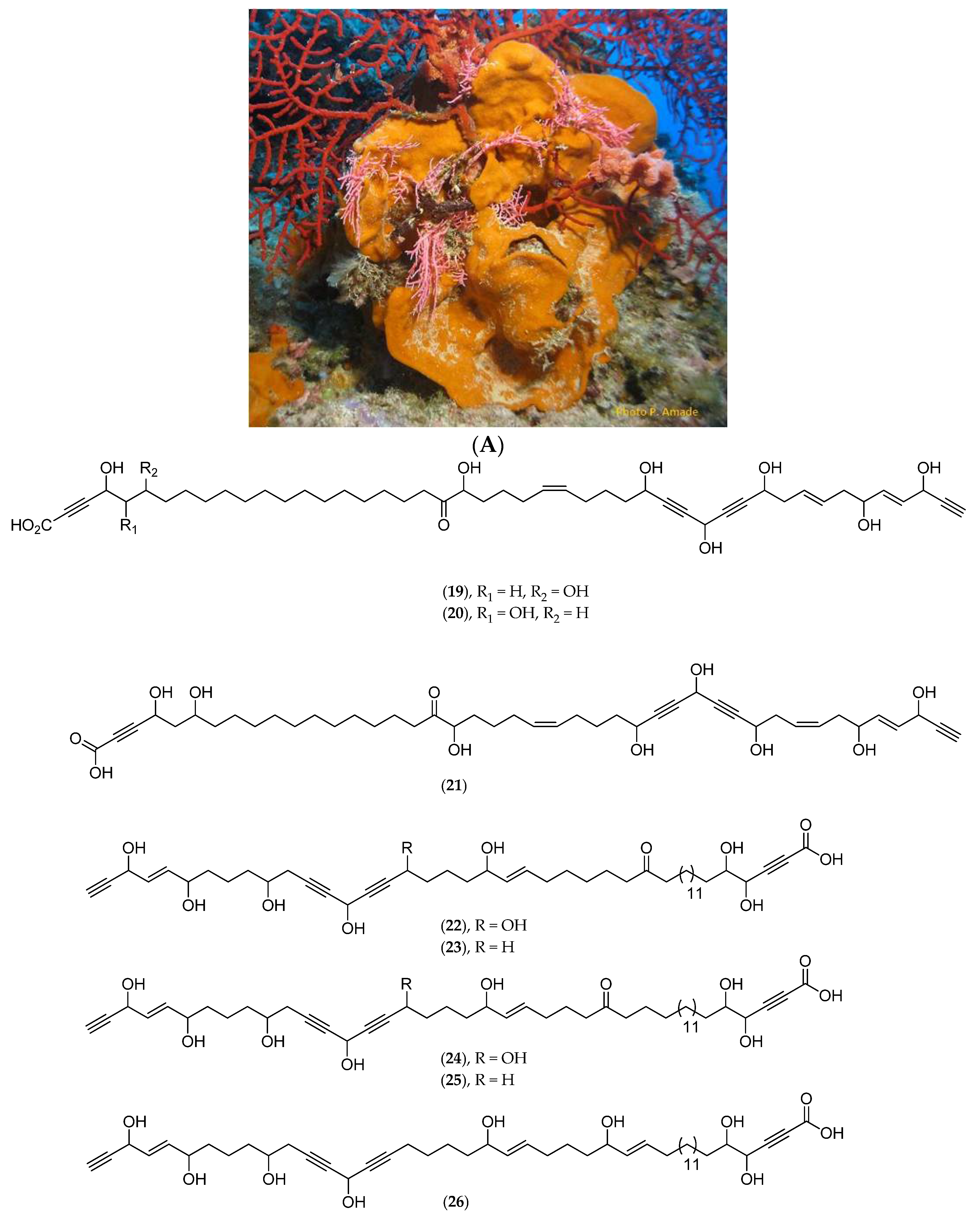
Xenia Garciae
5.3.2. Gorgonians
General Comments
Melithea cf. Stormii
Villogorgia Rubra
5.3.3. Pennatularians
General Comments
Lituaria Australasiae
5.3.4. Scleractinians
5.4. Echinoderms
5.4.1. General Comments
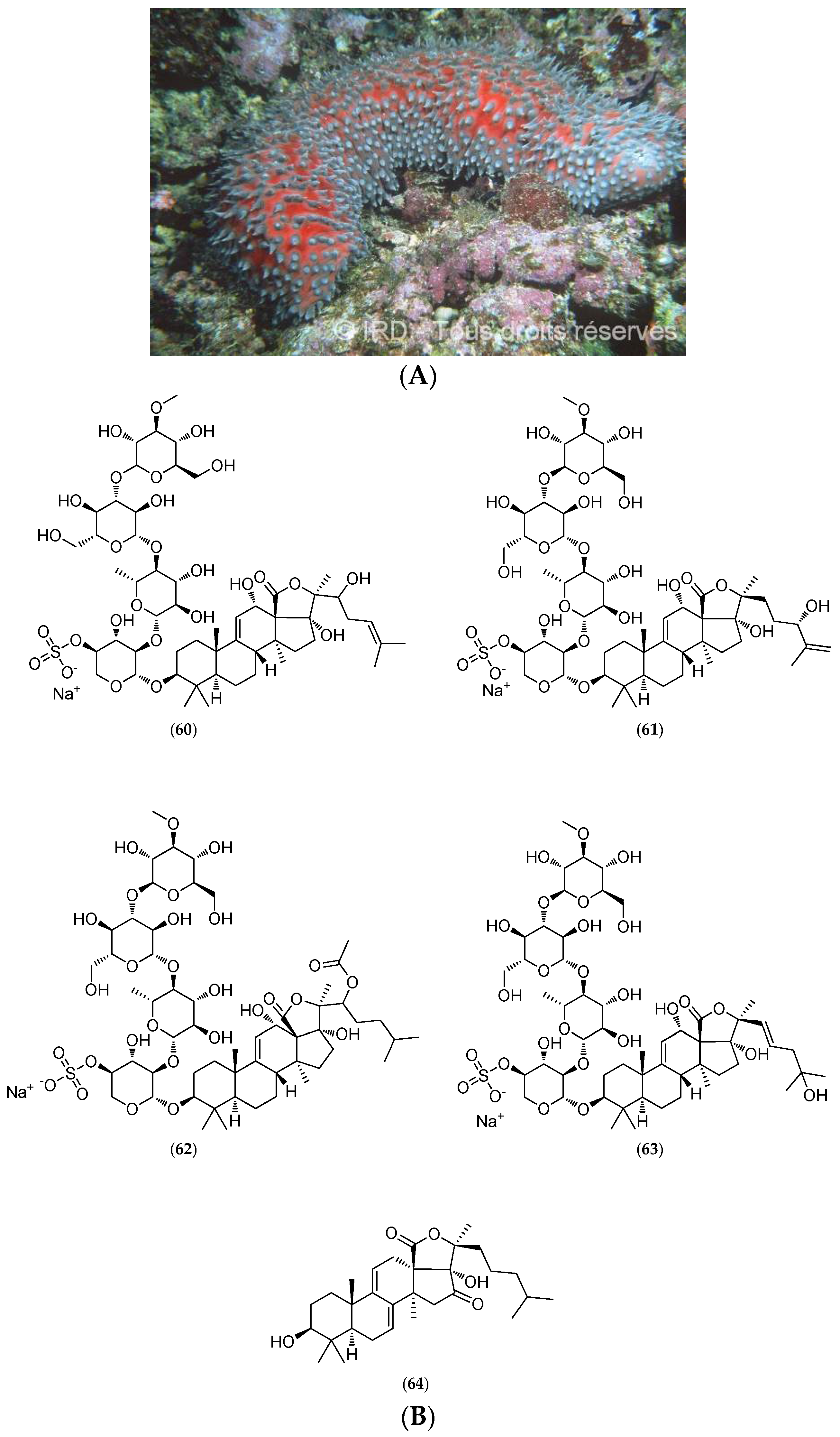
5.4.2. Actinopyga Flammea
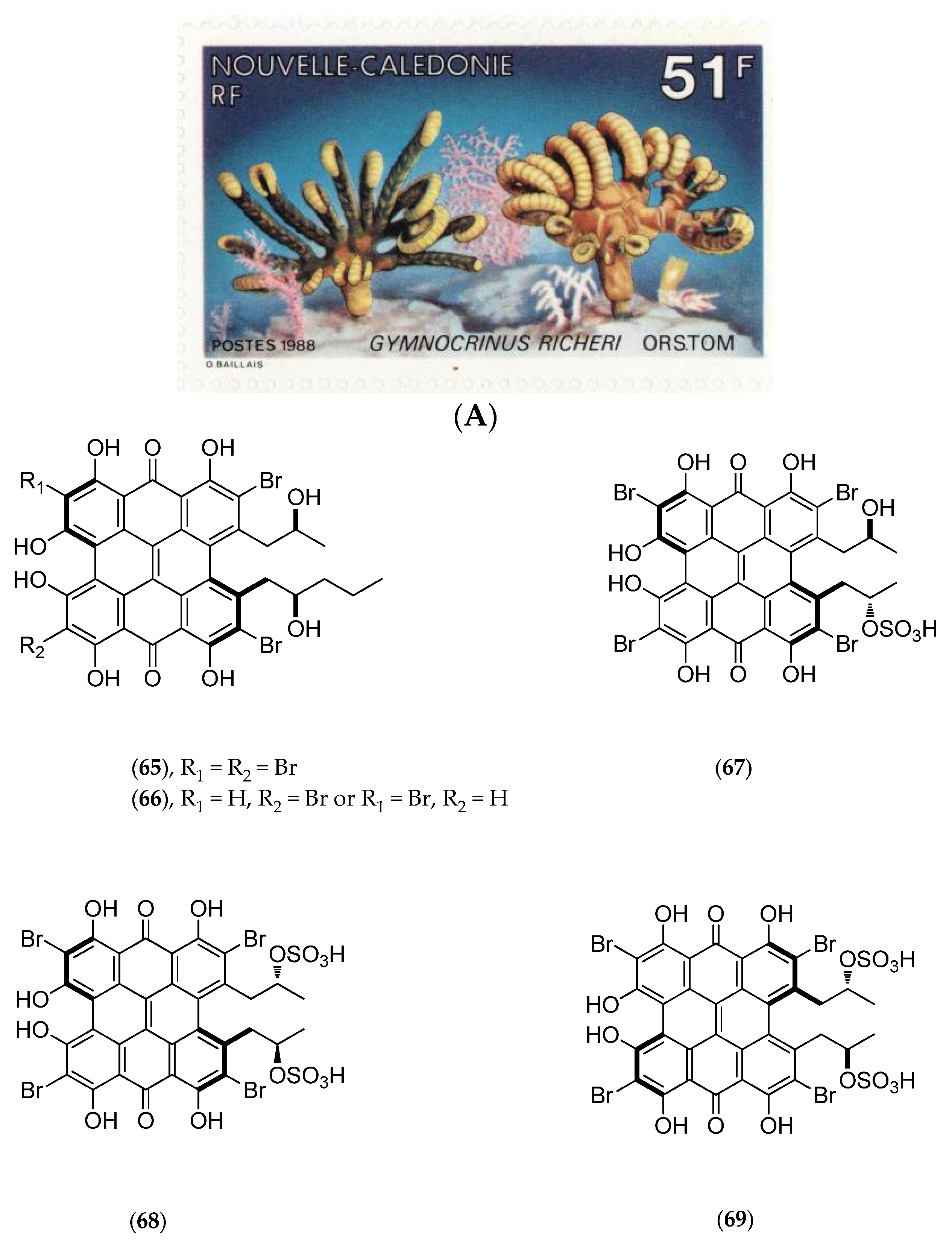
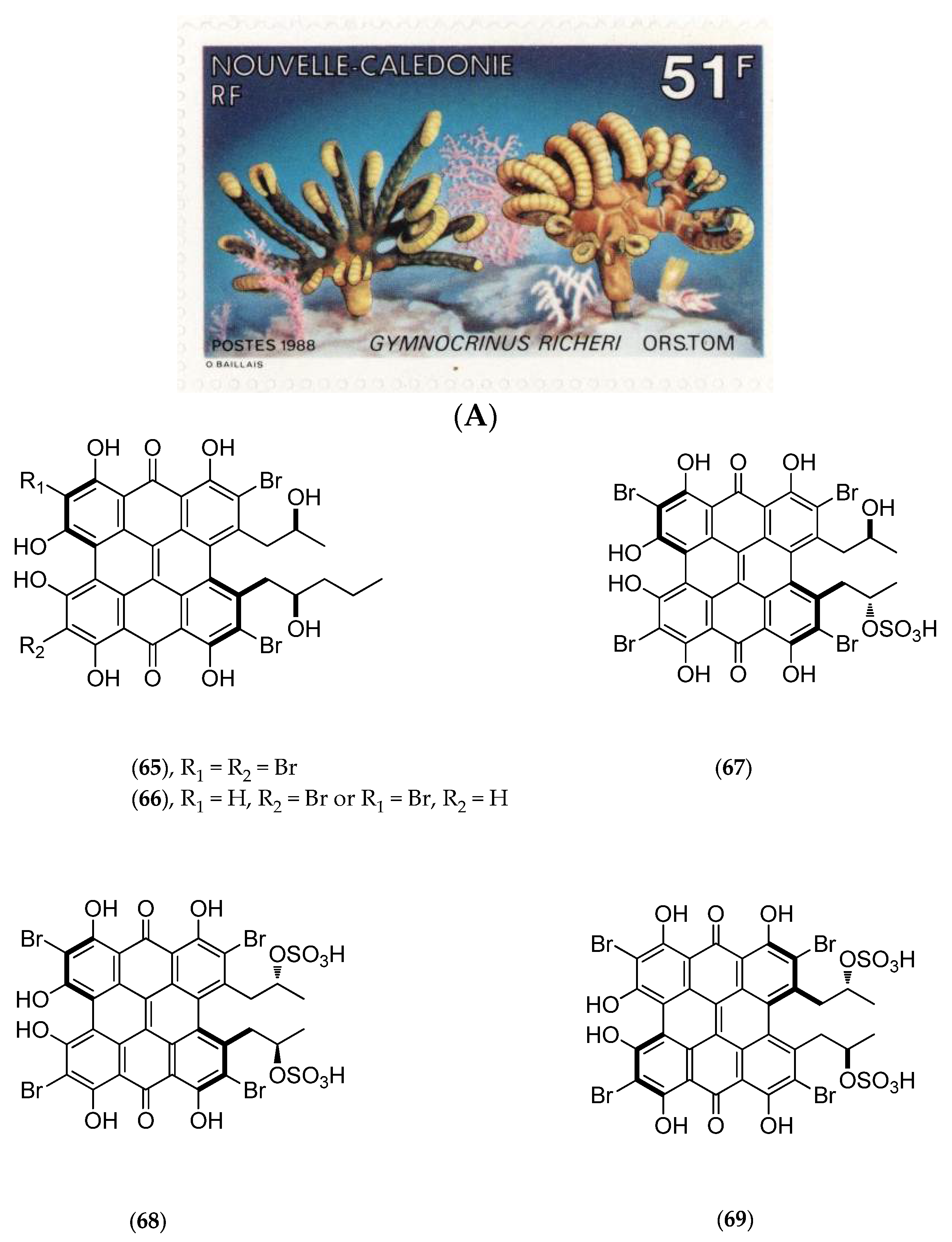
5.4.3. Gymnochrinus Richeri
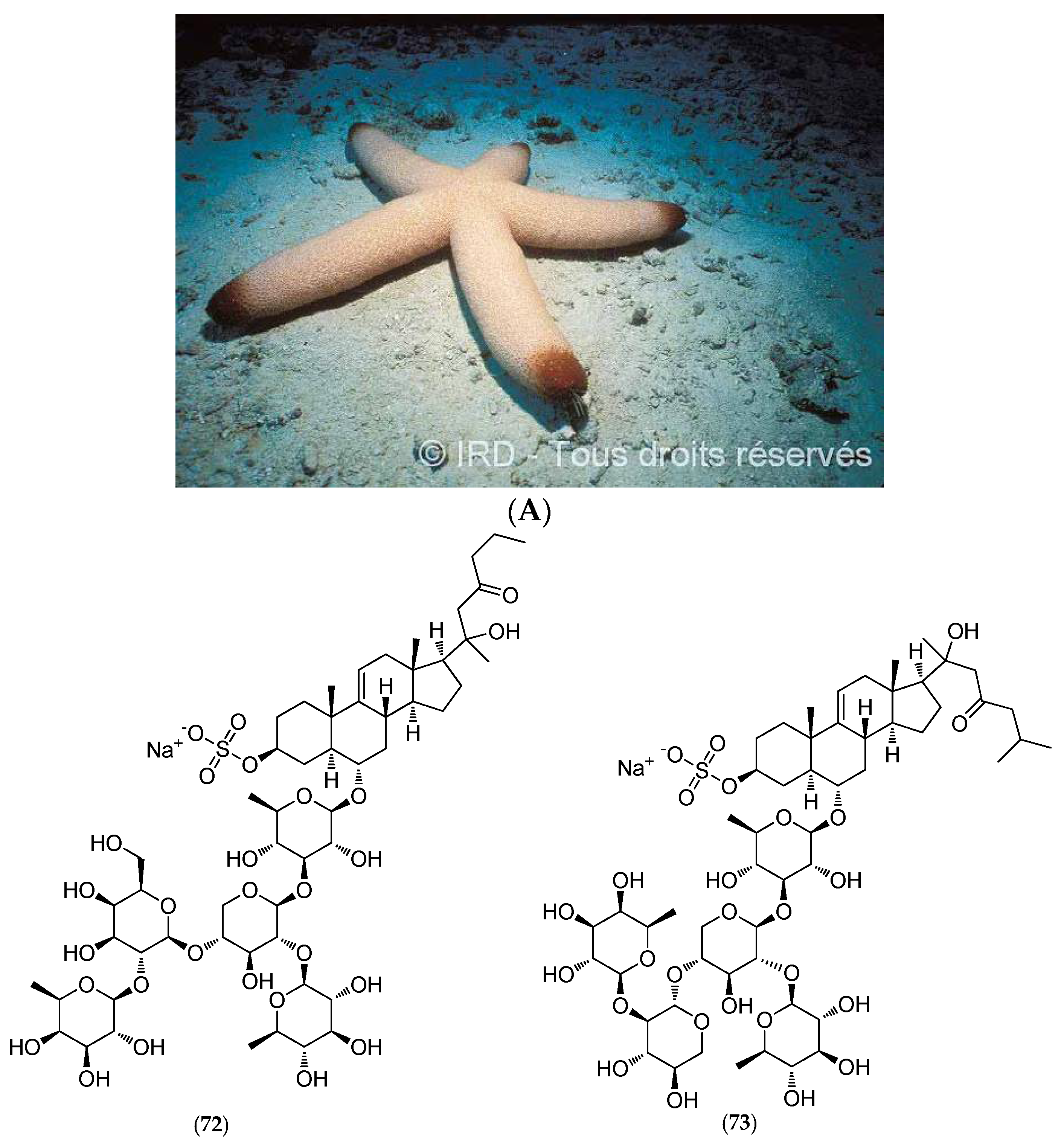
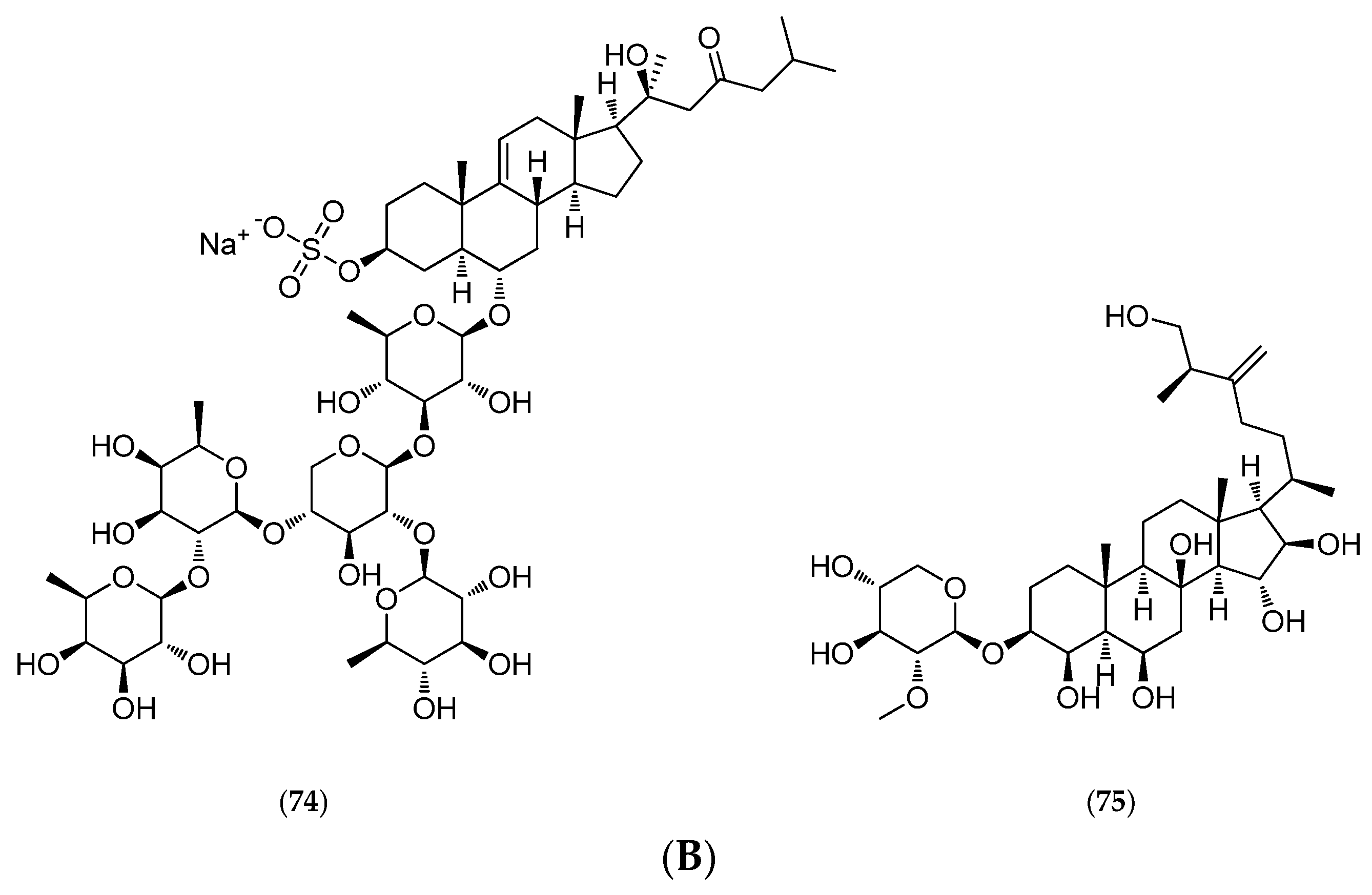
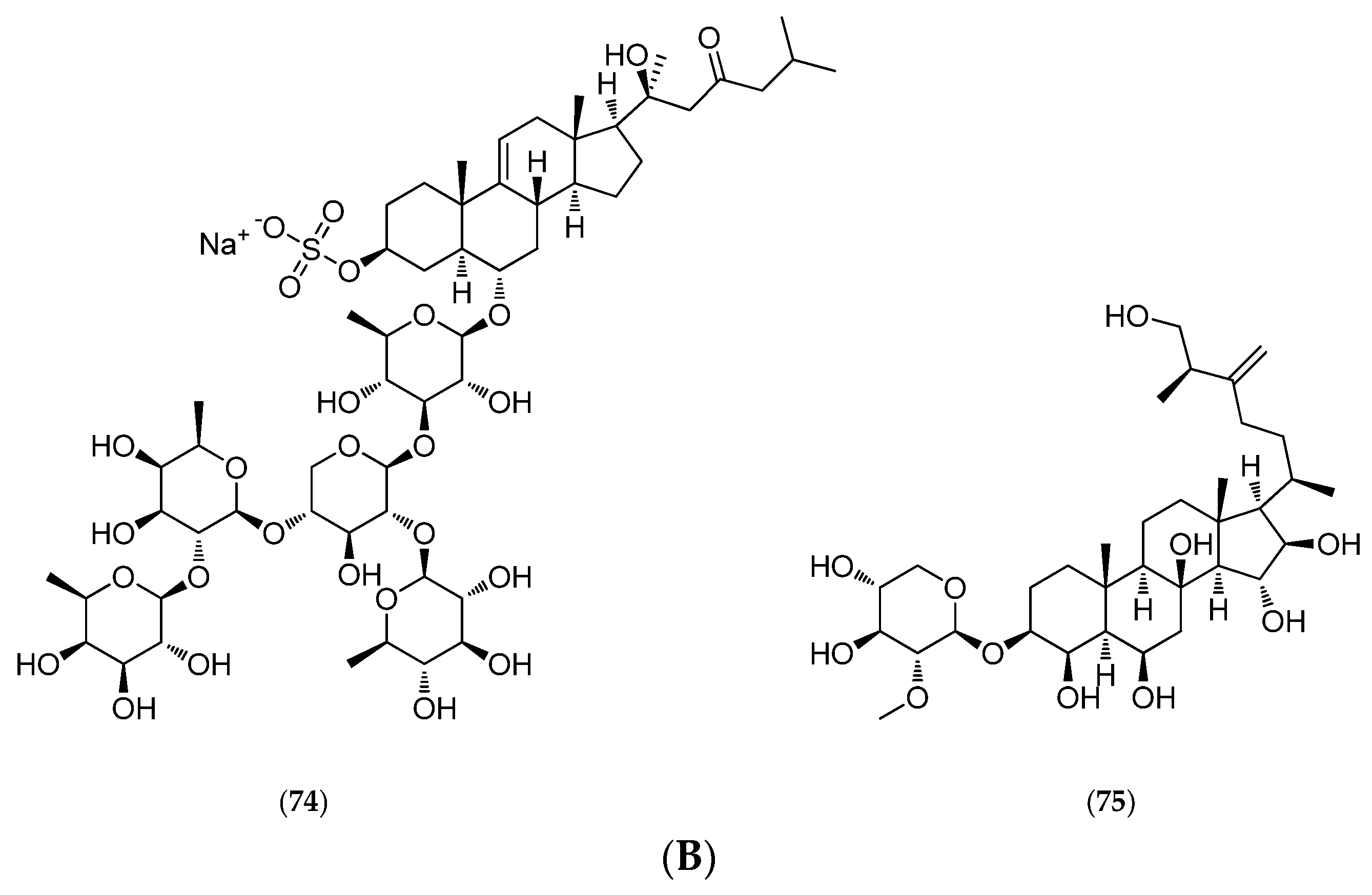
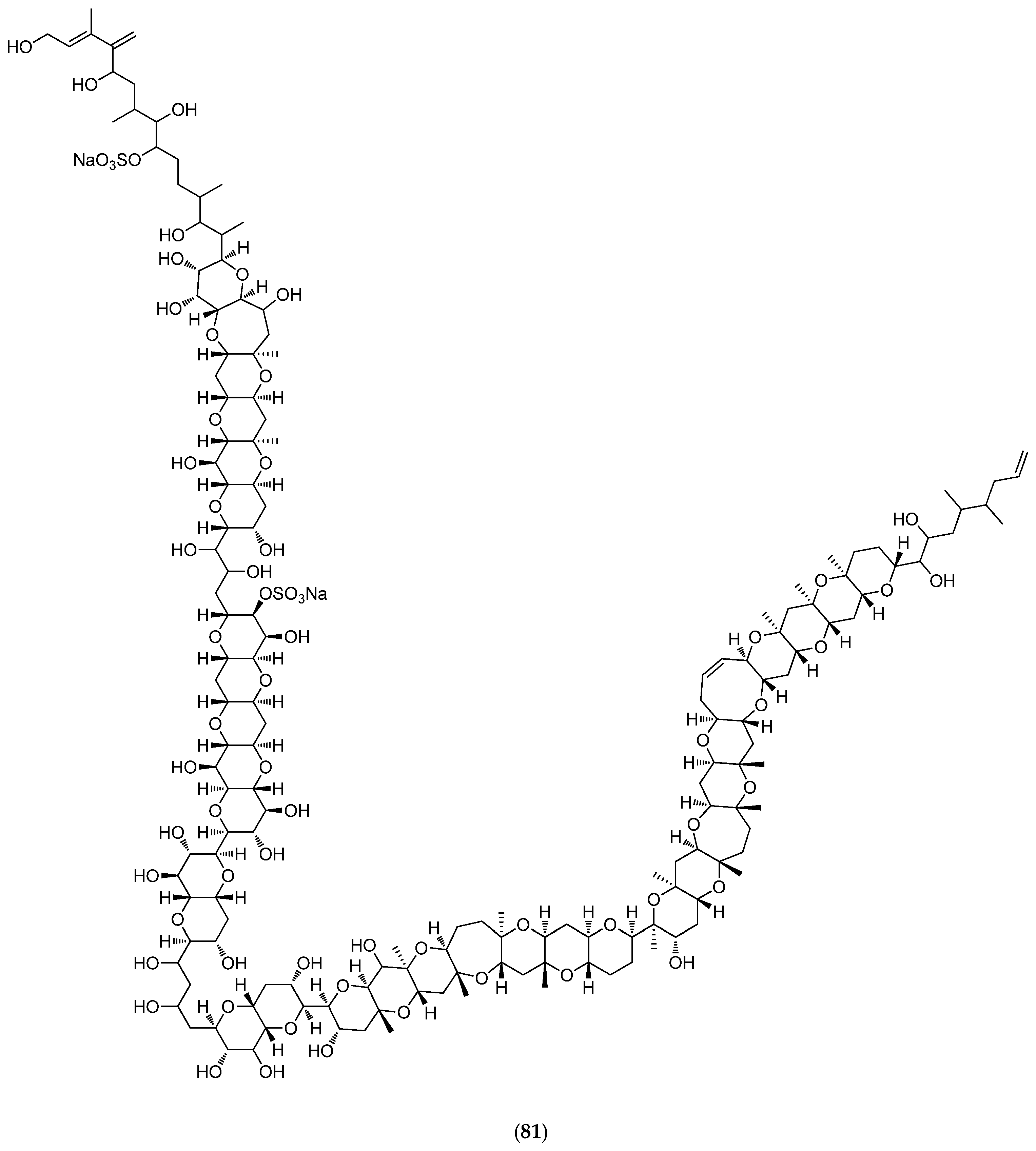
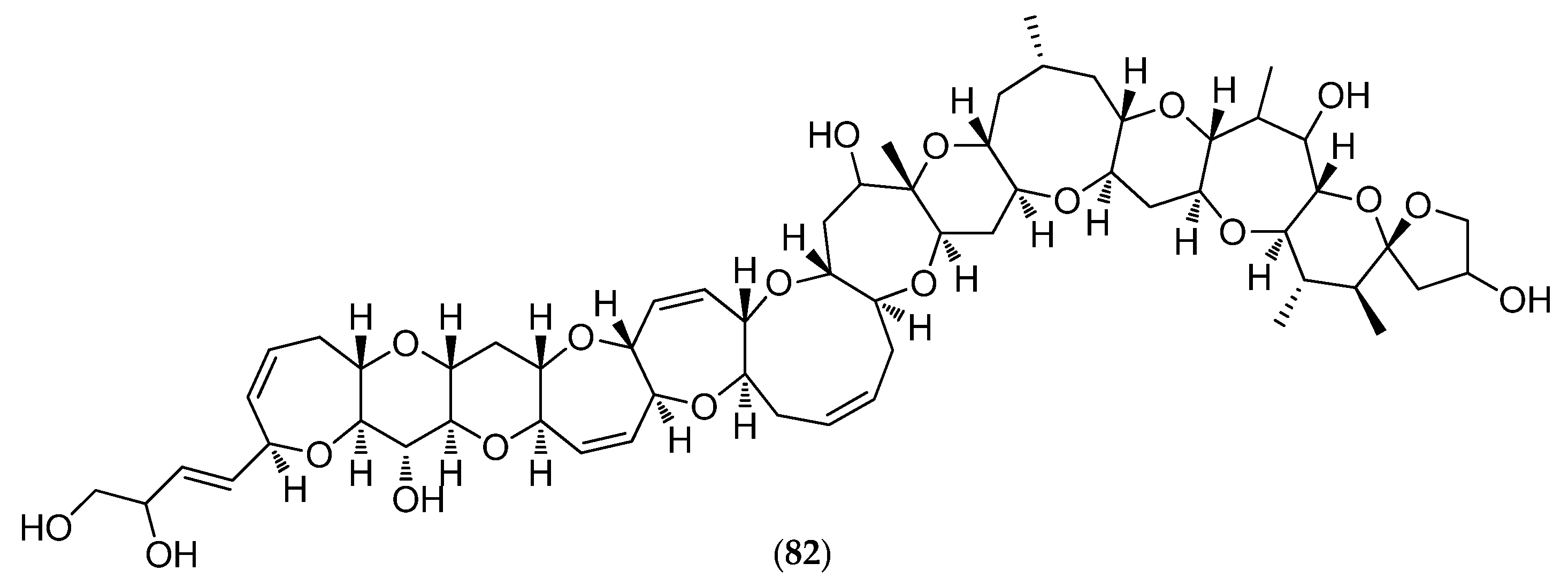
5.4.4. Thromidia Catalai
5.5. Macroalgae
5.6. Microalgae and Cyanobacteria
5.7. Other Biological Sources
5.7.1. Prokaryotes and Fungi
Micrococcus Luteus
Bacteria from Pseudoalteromonas and Vibrio Genus
Acremonium Neocaledoniae
5.7.2. Venomous Cone Snails
5.8. Vertebrates: Venomous Marine Snakes
6. Recent Advances on Selected New Caledonian Marine Natural Products
7. Conclusions
- -drug development, including (i) isolation of active principles; (ii) high-throughput bioactivity screening; (iii) structure-activity investigations and structural elucidation; (iv) cultivation technologies or bioinspired synthesis; (v) clinical trials and beyond;
- -aquaculture focusing on the treatment of locally grown species of prawns and oyster varieties that are sensitive to seasonal blooms of toxigenic bacteria and microalgae.
Acknowledgments
Conflicts of Interest
References
- Pelletier, B. Geology of the New Caledonia Region and its Implications for the Study of the New Caledonian Biodiversity. In Compendium of Marine Species from New Caledonia, 2nd ed.; Doc. Sci. Tech. II7; Payri, C.E., de Forges, B.R., Eds.; IRD Nouméa: Nouvelle-Calédonie, France, 2007; pp. 19–32. [Google Scholar]
- Andréfouët, S.; Cabioch, G.; Flamand, B.; Pelletier, B. A reappraisal of the diversity of geomorphological and genetic processes of New Caledonian coral reefs: A synthesis from optical remote sensing, coring and acoustic multibeam observations. Coral Reefs 2009, 28, 691–707. [Google Scholar] [CrossRef]
- Leal, M.C.; Puga, J.; Serôdio, J.; Gomes, N.C.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades—Where and what are we bioprospecting. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Andréfouët, S.; Cabioch, G.; Flamand, B.; Pelletier, B. The diversity of New Caledonia Coral Reef Geomorphology and Genetic Processes: A Synthesis from Optical Remote Sensing, Coring and Acoustic Multi-beam Observations. In Compendium of Marine Species from New Caledonia, 2nd ed.; Payri, C.E., de Forges, B.R., Eds.; IRD Editions: Nouméa, Nouvelle-Calédonie, 2007; pp. 33–49. [Google Scholar]
- Payri, C.; de Forges Richer, B. Compendium of Marine Species of New Caledonia, 2nd ed.; IRD Editions: Nouméa, Nouvelle-Calédonie, 2007. [Google Scholar]
- Natural History Museum. Available online: http://www.nhm.ac.uk/nature-online/science-of-natural-history/expeditions-collecting/hms-challenger-expedition/ (accessed on 21 May 2015).
- Armbrust, E.V.; Palumbi, S.R. Uncovering hidden worlds of ocean biodiversity. Science 2015, 348, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Catala, R. Contribution à l’étude écologique des îlots coralliens du Pacifique Sud. Premiers éléments d’écologie terrestre et marine des îlots voisins du littoral de la Nouvelle-Calédonie. Bull. Biol. Fr. Belg. 1950, 84, 234–310. (In French) [Google Scholar] [PubMed]
- Catala-Stucki, R. Fluorescence effects from corals irradiated with ultra-violet rays. Nature 1959, 183, 949. [Google Scholar] [CrossRef]
- Salvat, B. Prospections faunistiques en Nouvelle-Calédonie dans le cadre de la mission d’Etudes des Récifs Coralliens. Cahtay. Pac. 1964, 6, 77–119. (In French) [Google Scholar]
- Plessis, Y. Un voyage en Nouvelle-Calédonie dans le cadre de la mission Singer-Polignac. Cahtay. Pac. 1962, 4, 81–83. [Google Scholar]
- Foundation Singer-Polignac. Expédition Française sur les récifs Coralliens de la Nouvelle-Calédonie, Organisée Sous l'égide de la Fondation Singer-Polignac, 1960–1962; Editions de la Fondation Singer-Polignac: Paris, France, 1961. (In French) [Google Scholar]
- Dahl, A.L. Surface area in ecological analysis: Quantification of benthic coral reef algae. Mar. Biol. 1973, 23, 239–249. [Google Scholar] [CrossRef]
- Dahl, A.L. Coral Reef Monitoring Handbook; South Pacific Commission: Noumea, New Caledonia, 1981. [Google Scholar]
- Laurent, D.; Pietra, F. Natural-Product Diversity of the New Caledonian Marine Ecosystem Compared to Other Ecosystems: A Pharmacologically Oriented View. Chem. Biodivers. 2004, 1, 539–594. [Google Scholar] [CrossRef] [PubMed]
- Lévi, C.; Laboute, P.; Bargibant, G.; Menou, J.L. Sponges of the New Caledonian Lagoon; ORSTOM Editions (IRD): Nouméa, Nouvelle-Calédonie, 1998. [Google Scholar]
- Guille, A.; Laboute, P.; Menou, J.L. Guide des étoiles de mer, Oursins et Autres échinodermes du lagon de Nouvelle-Calédonie; ORSTOM Editions (IRD): Nouméa, Nouvelle-Calédonie, 1986. (In French) [Google Scholar]
- Grasshoff, M.; Bargibant, G. Coral Reef Gorgonians of New Caledonia; IRD Editions: Nouméa, Nouvelle-Calédonie, 2001. [Google Scholar]
- Monniot, C.; Monniot, F.; Laboute, P. Coral Reef Ascidians of New Caledonia; IRD Editions: Nouméa, Nouvelle-Calédonie, 1991. [Google Scholar]
- Ineich, Y.; Laboute, P. Sea Snakes of New Caledonia; IRD Editions/Muséum National d’Histoire Naturelle: Paris, France, 2002. [Google Scholar]
- Laboute, P.; Grandperrin, R. Poissons de Nouvelle-Calédonie; Editions Catherine Ledru: Nouméa, Nouvelle-Calédonie, 2000. (In French) [Google Scholar]
- Moretti, C.; Debitus, C.; Fournet, A.; Sauvain, M.; Bourdy, G.; Laurent, D. Diversité biologique tropicale et innovation thérapeutique. Les recherches menées par l’ORSTOM. Ann. Soc. Belg. Med. Trop. 1993, 73, 169–178. (In French) [Google Scholar] [PubMed]
- Debitus, C.; Guézennec, J. Pharmacologie des organismes marins des milieux récifaux. In Valorisation et économie des Ressources Marines; Monaco, A., Prouzet, P., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 113–148. (In French) [Google Scholar]
- LagPlon Database. Available online: http://lagplon.ird.nc/index.xhtml (accessed on 18 June 2015).
- Cantharella Database. Available online: http://cantharella.ird.nc (accessed on 19 June 2015).
- Kerr, T.J.; McHale, B.B. Applications in General Microbiology: A Laboratory Manual, 6th ed.; Hunter Textbooks: Winston-Salem, NC, USA, 2001; pp. 202–203. [Google Scholar]
- Solis, P.N.; Wright, C.W.; Anderson, M.M.; Gupta, M.P.; Phillipson, J.D. A microwell cytotoxicity assay using. Artemia Salina Plant Med. 1993, 59, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Coll, J.C.; La Barre, S.; Sammarco, P.W.; Williams, W.T.; Bakus, G.J. Chemical defenses in soft corals (Coelenterata: Octocorallia) of the Great Barrier Reef. Part I: A study of comparative toxicities. Mar. Ecol. Prog. Ser. 1982, 8, 271–278. [Google Scholar] [CrossRef]
- La Barre, S.; Laurent, D.; Sammarco, P.; Williams, W.T.; Coll, J.C. Comparative ichthyotoxicity of shallow and deep-water sponges of New Caledonia. In Proceedings of the 6th International Symposium on Coral Reefs, Townsville, Australia, 8–12 August 1988; Volume 3, pp. 55–59.
- Kobayashi, N. Comparative sensitivity of various developmental stages of sea urchins to some chemicals. Mar. Biol. 1980, 58, 163–171. [Google Scholar] [CrossRef]
- Brun, L.-O.; Urbain, R.; Wacapo, E.; Debitus, C. A Method for the Evaluation of Marine extracts Toxicity for the Coffee Berry Borer: Hypothenemus Hampei. In Proceeding of the Third Pacific-Asia Symposium on Biologically Active Natural Products, Noumea, New Caledonia, 26–30 August 1991; p. 31.
- Stone, B.F.; Haydock, K.P. Method derived from Stone and Haydock. A method for measuring the acaricide-susceptibility of the cattle tick Boophilus microplus (Can.). Bull. Entomol. Res. 1962, 53, 563–578. [Google Scholar] [CrossRef]
- König, G.M.; Coll, J.C.; Bowden, B.F.; Gulbis, J.M.; MacKay, M.F.; La Barre, S.C.; Laurent, D. The structure determination of a Xenicane diterpene from Xenia garciae. J. Nat. Prod. 1989, 52, 294–299. [Google Scholar] [CrossRef]
- Vieira, C.; Thomas, O.P.; Culioli, G.; Genta-Jouve, G.; Houlbreque, F.; Gaubert, J.; De Clerk, O.; Payri, C.E. Allelopathic interactions between the brown algal genus Lobophora (Dictyotales, Phaeophyceae) and scleractinian corals. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- La Barre, S.; (CNRS, Nouméa, New Caledonia). Personal communication, 1989.
- Green, N.M.; Work, E. Pancreatic trypsin inhibitor. 2. Reaction with trypsin. Biochem. J. 1953, 54, 347–352. [Google Scholar] [CrossRef] [PubMed]
- La Barre, S.; Longeon, A.; Barthélémy, M.; Guyot, M.; Le Caer, J.P.; Bargibant, G. Characterization of a novel elastase inhibitor from a fan coral. C. R. Acad. Sci. III 1996, 319, 365–370. [Google Scholar] [PubMed]
- Faulkner, D.J.; Unson, M.D.; Bewley, C.A. The chemistry of some sponges and their symbionts. Pure Appl. Chem. 1994, 66, 1983–1990. [Google Scholar] [CrossRef]
- Kobayashi, J.; Ishibashi, M. Bioactive metabolites of symbiotic marine microorganisms. Chem. Rev. 1993, 93, 1753–1769. [Google Scholar] [CrossRef]
- Kornprobst, J.M. Porifera (Sponges). In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 2, pp. 607–1086. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; Debitus, C.; Ribes, O.; Pusset, J.; Leroy, S.; Pietra, F. Agelastatin A, a new skeleton cytotoxic alkaloid of the oroidin family. Isolation from the axinellid sponge Agelas dendromorpha of the Coral Sea. J. Chem. Soc. Chem. Commun. 1993, 16, 1305–1306. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Chiasera, G.; Pietra, F. Conformational preferences and absolute configuration of agelastatin A, a cytotoxic alkaloid of the axinellid sponge Agelas dendromorpha from the Coral Sea, via combined molecular modelling, NMR, and exciton splitting for diamide and hydroxyamide derivatives. Helv. Chim. Acta 1994, 77, 1895–1902. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; Ripamonti, M.; Debitus, C.; Waikedre, J.; Pietra, F. The active centres of agelastatin A, a strongly cytotoxic alkaloid of the Coral Sea axinellid sponge Agelas dendromorpha, as determined by comparative bioassays with semisynthetic derivatives. Helv. Chim. Acta 1996, 79, 727–735. [Google Scholar] [CrossRef]
- Meijer, L.; Thunnissen, A.M.; White, A.W.; Garnier, M.; Nikolic, M.; Tsai, L.H.; Walter, J.; Cleverley, K.E.; Salinas, P.C.; Wu, Y.Z.; et al. Inhibition of cyclin-dependent kinases, GSK-3β and CK1 by hymenialdisine, a marine sponge constituent. Chem. Biol. 2000, 7, 51–63. [Google Scholar] [CrossRef]
- Vassas, A.; Bourdy, G.; Paillard, J.J.; Lavayre, J.; Pais, M.; Quirion, J.C.; Debitus, C. Naturally occurring somatostatin and vasoactive intestinal peptide inhibitors. Isolation of alkaloids from two marine sponges. Plant. Med. 1996, 62, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; D’Auria, M.V.; Paloma, L.G.; Casapullo, A.; Minale, L.; Debitus, C.; Henin, Y. Callipeltin A. an anti-HIV cyclic depsipeptide from the New Caledonian Lithistida sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 6202–6209. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Zampella, A.; Paloma, L.G.; Minale, L.; Debitus, C.; Roussakis, C.; Le Bert, V. Callipeltins B and C; bioactive peptides from a marine Lithistida sponge Callipelta sp. Tetrahedron 1996, 52, 9589–9596. [Google Scholar]
- Zampella, A.; D’Auria, M.V.; Minale, L.; Debitus, C.; Roussakis, C. Callipeltoside A: A cytotoxic aminodeoxy sugar-containing macrolide of a new type from the marine Lithistida sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 11085–11088. [Google Scholar] [CrossRef]
- Zampella, A.; D’Auria, M.V.; Minale, L.; Debitus, C. Callipeltosides B and C, two novel cytotoxic glycoside macrolides from a marine lithistida sponge Callipelta sp. Tetrahedron 1997, 53, 3243–3248. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Debitus, C.; Ribes, O.; Richer de Forges, B.; Pietra, F. Corallistin A, a second example of a free porphyrin from a living organism. Isolation from the demosponge Corallistes sp. of the Coral Sea and inhibition of abnormal cells. Helv. Chim. Acta 1989, 72, 1451–1454. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Debitus, C.; Ribes, O.; Pietra, F. On the Novel Free Porphyrins Corallistin B, C, D, and E: Isolation from the demosponge Corallistes sp. of the Coral Sea and Reactivity of Their Nickel 55 (II) Complexes toward Formylating Reagents. Helv. Chim. Acta 1993, 76, 1489–1496. [Google Scholar] [CrossRef]
- Loukaci, A.; Le Saout, I.; Samadi, M.; Leclerc, S.; Damiens, E.; Meijer, L.; Debitus, C.; Guyot, M. Coscinosulfate, a CDC25 phosphatase inhibitor from the sponge Coscinoderma mathewsi. Bioorg. Med. Chem. 2001, 9, 3049–3054. [Google Scholar] [CrossRef]
- De Nanteuil, G.; Ahond, A.; Guilhem, J.; Poupat, C.; Tran Huu Dau, E.; Potier, P.; Pusset, M.; Pusset, J.; Laboute, P. Invertébrés marins du lagon néo-calédonien-V. Isolement et identification des métabolites d’une nouvelle espèce de spongiaire, Pseudaxinyssa cantharella. Tetrahedron 1985, 41, 6019–6033. (In French) [Google Scholar] [CrossRef]
- Ahond, A.; Laboute, P.; Laurent, D.; Potier, P.; Poupat, C.; Pusset, J.; Pusset, M.; Thoison, O. New Biologically Active Substance Called Girolline, Extracted from the Sponge Pseudaxinyssa cantharella Process for its Preparation and Pharmaceutical Compositions Containing It. U.S. Patent 4,801,602, 31 January 1989. [Google Scholar]
- Ahond, A.; Bedoya Zurita, M.; Colin, M.; Fizames, C.; Laboute, P.; Lavelle, F.; Laurent, D.; Poupat, C.; Pusset, J.; Pusset, M.; et al. La girolline, nouvelle substance antitumorale extraite de l’éponge, Pseudaxinyssa cantharella n. sp. (Axinellidae). C. R. Acad. Sci. Ser. 1988, 307, 145–148. (In French) [Google Scholar]
- Chiaroni, A.; Riche, C.; Ahond, A.; Poupat, C.; Pusset, M.; Potier, P. Structure cristalline et configuration absolue de la girolline. C. R. Acad. Sci. Ser. 1991, 312, 49–53. [Google Scholar]
- Bedoya Zurita, M.; Ahond, A.; Poupat, C.; Potier, P. Première synthèse totale de la girolline. Tetrahedron 1989, 45, 6713–6720. (In French) [Google Scholar] [CrossRef]
- Marchais, S.; Al Mourabit, A.; Ahond, A.; Poupat, C.; Potier, P. A short synthesis of the marine bioactive metabolite (+/−) girolline. Tetrahedron Lett. 1998, 39, 8085–8088. [Google Scholar] [CrossRef]
- Colson, G.; Rabault, B.; Lavelle, F.; Zerial, A. Mode of action of the antitumor compound girodazole (RP 49532A, NSC 627434). Biochem. Pharmacol. 1992, 43, 1717–1723. [Google Scholar] [CrossRef]
- Catimel, G.; Coquard, R.; Guastalla, J.P.; Merrouche, Y.; Le Bail, N.; Alakl, M.K.; Dumortier, A.; Foy, M.; Clavel, M. Phase I study of RP 49532A, a new protein-synthesis inhibitor, in patients with advanced refractory solid tumors. Cancer Chemother. Pharm. 1995, 35, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Diop, D.; Chauvin, C.; Salhi, S.; Poupat, C.; Ahond, A.; Jean-Jean, O. Girolline interferes with cell-cycle progression, but not with translation. C. R. Biol. 2007, 330, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Benoit-Vical, F.; Saléry, M.; Soh, P.N.; Ahond, A.; Poupat, C. Girolline: A potential lead structure for antiplasmodial drug research. PlantMed 2008, 74, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Al Mourabit, A.; Pusset, M.; Chtourou, M.; Gaigne, C.; Ahond, A.; Poupat, C.; Potier, P. Pyraxinine, a novel nitrogenous compound from the marine sponge Cymbastela cantharella. J. Nat. Prod. 1997, 60, 290–291. [Google Scholar] [CrossRef]
- Sauleau, P.; Retailleau, P.; Nogues, S.; Carletti, I.; Marcourt, L.; Raux, R.; Al Mourabit, A.; Debitus, C. Dihydrohymenialdisines, new pyrrole-2-aminoimidazole alkaloids from the marine sponge Cymbastela cantharella. Tetrahedron Lett. 2011, 52, 2676–2678. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Debitus, C.; Waikedre, J.; Pietra, F. Relative contributions to antitumoral activity of lipophilic vs. polar reactive moieties in marine terpenoids. Tetrahedron Lett. 1997, 38, 6285–6288. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Deharo, E.; Debitus, C.; Munoz, V.; Pietra, F. New Types of Potentially Antimalarial Agents: Epidioxy-substituted norditerpene and norsesterpenes from the marine sponge Diacarnus Levii. Helv. Chim. Acta 1998, 81, 1285–1292. [Google Scholar] [CrossRef]
- Mancini, I.; Guella, G.; Debitus, C.; Waikedre, J.; Pietra, F. From inactive nortopsentin D, a novel bis (indole) alkaloid isolated from the axinellid sponge Dragmacidon sp. from deep waters south of New Caledonia, to a strongly cytotoxic derivative. Helv. Chim. Acta 1996, 79, 2075–2082. [Google Scholar] [CrossRef]
- Mancini, I.; Guella, G.; Frostin, M.; Hnawia, E.; Laurent, D.; Debitus, C.; Pietra, F. On the first polyarsenic organic compound from nature: Arsenicin A from the New Caledonian marine sponge Echinochalina bargibanti. Chem. Eur. J. 2006, 12, 8989–8994. [Google Scholar] [CrossRef] [PubMed]
- Dopeso, J.; Quiñoá, E.; Riguera, R.; Debitus, C.; Bergquist, P.R. Euryspongiols: Ten new highly hydroxylated 9, 11-secosteroids with antihistaminic activity from the sponge Euryspongia sp. Stereochemistry and reduction. Tetrahedron 1994, 50, 3813–3828. [Google Scholar] [CrossRef]
- Zampella, A.; Sepe, V.; Luciano, P.; Bellotta, F.; Monti, M.C.; D’Auria, M.V.; Jepsen, T.; Petek, S.; Adeline, M.T.; Laprévôte, O.; et al. Homophymine A, an anti-HIV cyclodepsipeptide from the sponge Homophymia sp. J. Org. Chem. 2008, 73, 5319–5327. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; Sepe, V.; Bellotta, F.; Luciano, P.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Petek, S.; Poupat, C.; Ahond, A. Homophymines B–E and A1–E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009, 7, 4037–4044. [Google Scholar] [CrossRef] [PubMed]
- Bourguet-Kondracki, M.L.; Martin, M.T.; Debitus, C.; Guyot, M. 12-epi-heteronemin: New sesterterpene from the marine sponge Hyrtios erecta. Tetrahedron Lett. 1994, 35, 109–110. [Google Scholar] [CrossRef]
- Ledroit, V.; Debitus, C.; Ausseil, F.; Raux, R.; Menou, J.L.; Hill, B. Heteronemin as a protein farnesyl transferase inhibitor. Pharm. Biol. 2004, 42, 454–456. [Google Scholar] [CrossRef]
- Bourguet-Kondracki, M.L.; Debitus, C.; Guyot, M. Biologically active sesterterpenes from a New Caledonian marine sponge Hyrtios sp. J. Chem. Res. 1996, 192–193. [Google Scholar]
- Bourguet-Kondracki, M.L.; Debitus, C.; Guyot, M. Dipuupehedione, a cytotoxic new red dimer from a New Caledonian marine sponge Hyrtios sp. Tetrahedron Lett. 1996, 37, 3861–3864. [Google Scholar] [CrossRef]
- Bourguet-Kondracki, M.L.; Lacombe, F.; Guyot, M. Methanol adduct of puupehenone, a biologically active derivative from the marine sponge Hyrtios species. J. Nat. Prod. 1999, 62, 1304–1305. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R.; Debitus, C.; Bourdy, G.; Vassas, A.; Lavayre, J. Bioactive prenylhydroquinone sulfates and a novel C31 furanoterpene alcohol sulfate from the marine sponge, Ircinia sp. J. Nat. Prod. 1995, 58, 1444–1449. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Guerriero, A.; Debitus, C.; Pietra, F. Leucascandrolide A, a new type of macrolide: The first powerfully bioactive metabolite of calcareous sponges (Leucascandra caveolata, a new genus from the Coral Sea). Helv. Chim. Acta 1996, 79, 51–60. [Google Scholar] [CrossRef]
- D’Ambrosio, M.; Tatò, M.; Pocsfalvi, G.; Debitus, C.; Pietra, F. Leucascandrolide B, a New 16-Membered, Extensively Methyl-Branched Polyoxygenated Macrolide from the Calcareous Sponge Leucascandra caveolata from Northeastern Waters of New Caledonia. Helv. Chim. Acta 1999, 82, 347–353. [Google Scholar] [CrossRef]
- Bewley, C.A.; Debitus, C.; Faulkner, D.J. Microsclerodermins A and B. Antifungal cyclic peptides from the Lithistid sponge Microscleroderma sp. J. Am. Chem. Soc. 1994, 116, 7631–7636. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Zampella, A.; Verbist, J.F.; Roussakis, C.; Debitus, C. Three new potent cytotoxic macrolides closely related to sphinxolide from the New Caledonian sponge Neosiphonia superstes. Tetrahedron 1993, 49, 8657–8664. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Zampella, A.; Debitus, C.; Perez, J. Neosiphoniamolide A, a novel cyclodepsipeptide, with antifungal activity from the marine sponge Neosiphonia superstes. J. Nat. Prod. 1995, 58, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Carbonelli, S.; Zampella, A.; Randazzo, A.; Debitus, C.; Gomez-Paloma, L. Sphinxolides E–G and reidispongiolide C: Four new cytotoxic macrolides from the New Caledonian lithistida sponges N. superstes and R. coerulea. Tetrahedron 1999, 55, 14665–14674. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Debitus, C.; Paloma, L.G.; Minale, L.; Zampella, A. Superstolide A: A potent cytotoxic macrolide of a new type from the New Caledonian deep water marine sponge Neosiphonia superstes. J. Am. Chem. Soc. 1994, 116, 6658–6663. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Zampella, A.; Debitus, C. A novel cytotoxic macrolide, superstolide B, related to superstolide A, from the New Caledonian marine sponge Neosiphonia superstes. J. Nat. Prod. 1994, 57, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Legrave, N.; Hamrouni-Buonomo, S.; Dufies, M.; Guérineau, V.; Vacelet, J.; Auberger, P.; Amade, P.; Mehiri, M. Nepheliosyne B, a new polyacetylenic acid from the New Caledonian marine sponge Niphates sp. Mar. Drugs 2013, 11, 2282–2292. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R.; Calignano, A.; Debitus, C. (±)-Gelliusines A and B, two diastereomeric brominated tris-indole alkaloids from a deep water New Caledonian marine sponge (Gellius or Orina sp.). J. Nat. Prod. 1994, 57, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, G.; Bruno, I.; Riccio, R.; Lavayre, J.; Bourdy, G. Further brominated bis- and tris-indole alkaloids from the deep-water New Caledonian marine sponge Orina sp. J. Nat. Prod. 1995, 58, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- Paloma, L.G.; Randazzo, A.; Minale, L.; Debitus, C.; Roussakis, C. New cytotoxic sesterterpenes from the New Caledonian marine sponge Petrosaspongia nigra (Bergquist). Tetrahedron 1997, 53, 10451–10458. [Google Scholar] [CrossRef]
- Randazzo, A.; Debitus, C.; Minale, L.; Garcia Pastor, P.; Alcaraz, M.J.; Payá, M.; Gomez-Paloma, L. Petrosaspongiolides M–R: New Potent and Selective Phospholipase A2 Inhibitors from the New Caledonian Marine Sponge Petrosaspongia nigra. J. Nat. Prod. 1998, 61, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Kourany-Lefoll, E.; Pais, M.; Sévenet, T.; Guittet, E.; Montagnac, A.; Fontaine, C.; Guénard, D.; Adeline, M.T.; Debitus, C. Phloeodictines A and B: New antibacterial and cytotoxic bicyclic amidinium salts from the New Caledonian sponge, Phloeodictyon sp. J. Org. Chem. 1992, 57, 3832–3835. [Google Scholar] [CrossRef]
- Kourany-Lefoll, E.; Laprévote, O.; Sévenet, T.; Montagnac, A.; Pais, M.; Debitus, C. Phloeodictines A1–A7 and C1–C2, antibiotic and cytotoxic guanidine alkaloids from the New Caledonian sponge, Phloeodictyon sp. Tetrahedron 1994, 50, 3415–3426. [Google Scholar] [CrossRef]
- Chevallier, C.; Laprévote, O.; Bignon, J.; Debitus, C.; Guénard, D.; Sévenet, T. Isolation of cytotoxic chondropsins, macrolide lactams from the New Caledonian marine sponge Psammoclemma sp. and electrospray ion trap multiple stage MS study of these macrolides. Nat. Prod. Res. 2004, 18, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Rubnov, S.; Chevallier, C.; Thoison, O.; Debitus, C.; Laprevote, O.; Guénard, D.; Sévenet, T. Echinosulfonic acid D: An ESI MSn evaluation of a new cytotoxic alkaloid from the New Caledonian sponge Psammoclemma sp. Nat. Prod. Res. 2005, 19, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, M.S.; Carroll, A.R.; Addepalli, R.; Avery, V.M.; Hooper, J.N.; Quinn, R.J. Psammaplysenes C and D, cytotoxic alkaloids from Psammoclemma sp. J. Nat. Prod. 2007, 70, 1827–1829. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Zampella, A.; Verbist, J.F.; Roussakis, C.; Debitus, C.; Patissou, J. Reidispongiolide A and B, two new potent cytotoxic macrolides from the New Caledonian sponge Reidispongia coerulea. Tetrahedron 1994, 50, 4829–4834. [Google Scholar] [CrossRef]
- Bourguet-Kondracki, M.L.; Longeon, A.; Debitus, C.; Guyot, M. New cytotoxic isomalabaricane-type sesterterpenes from the New Caledonian marine sponge Rhabdastrella globostellata. Tetrahedron Lett. 2000, 41, 3087–3090. [Google Scholar] [CrossRef]
- Debitus, C.; Guella, G.; Mancini, I.; Waikedre, J.; Guemas, J.P.; Nicolas, J.L.; Pietra, F. Quinolones from a bacterium and tyrosine metabolites from its host sponge, Suberea creba from the Coral Sea. J. Mar. Biotechnol. 1998, 6, 136–141. [Google Scholar] [PubMed]
- Quirion, J.C.; Sévenet, T.; Husson, H.P.; Weniger, B.; Debitus, C. Two new alkaloids from Xestospongia sp., a New Caledonian sponge. J. Nat. Prod. 1992, 55, 1505–1508. [Google Scholar] [CrossRef] [PubMed]
- De Nanteuil, G.; Ahond, A.; Poupat, C.; Potier, P.; Pusset, M.; Pusset, J.; Laboute, P. Invertebres marins du lagon neo-caledonien—VI : Isolement et identification de onze sterols de type hydroxymethyl-3β nor-A cholestane du spongiaire, Pseudaxinyssa cantharella. Tetrahedron 1985, 41, 6035–6039. [Google Scholar] [CrossRef]
- Lu, D.; Rae, A.D.; Salem, G.; Weir, M.L.; Willis, A.C.; Wild, S.B. Arsenicin A, a natural polyarsenical: Synthesis and crystal structure. Organometallics 2010, 29, 32–33. [Google Scholar] [CrossRef]
- Lu, D.; Coote, M.L.; Ho, J.; Kilah, N.L.; Lin, C.Y.; Salem, G.; Weir, M.L.; Willis, A.C.; Wild, S.B. Resolution and Improved Synthesis of (±)-Arsenicin A: A Natural Adamantane-Type Tetraarsenical Possessing Strong Anti-Acute Promelocytic Leukemia Cell Line Activity. Organometallics 2012, 31, 1808–1816. [Google Scholar] [CrossRef]
- Valentin, B.B.; Vinod, V.; Beulah, M.C. Biopotential of secondary metabolites isolated from marine sponge Dendrilla nigra. Asian Pac. J. Trop. Dis. 2011, 1, 299–303. [Google Scholar] [CrossRef]
- Selvin, J.; Lipton, A.P. Dendrilla nigra, a marine sponge, as potential source of antibacterial substances for managing shrimp diseases. Aquaculture 2004, 236, 277–283. [Google Scholar] [CrossRef]
- Liu, R.; Liu, Y.; Zhou, Y.D.; Nagle, D.G. Molecular-targeted antitumor agents. 15. Neolamellarins from the marine sponge Dendrilla nigra inhibit hypoxia-inducible factor-1 activation and secreted vascular endothelial growth factor production in breast tumor cells. J. Nat. Prod. 2007, 70, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.V.; Ravinder, K.; Narasimhulu, M.; Sridevi, A.; Satyanarayana, N.; Kondapi, A.K.; Venkateswarlu, Y. New anticancer bastadin alkaloids from the sponge Dendrilla cactos. Bioorg. Med. Chem. 2006, 14, 4452–4457. [Google Scholar] [CrossRef] [PubMed]
- Witowski, C.G. Investigation of Bioactive Metabolites from the Antarctic Sponge Dendrilla Membranosa and Marine Microorganisms. Ph.D. thesis, University of South Florida, Tampa, FL, USA, 10 April 2015. [Google Scholar]
- Selvin, J.; Gandhimathi, R.; Kiran, G.S.; Priya, S.S.; Ravji, T.R.; Hema, T.A. Culturable heterotrophic bacteria from the marine sponge Dendrilla nigra: Isolation and phylogenetic diversity of actinobacteria. Helgol. Mar. Res. 2009, 63, 239–247. [Google Scholar] [CrossRef]
- Ersmark, K.; Del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Yon-Hin, P.; Scott, A.I. Total synthesis of corallistin A. Tetrahedron Lett. 1991, 32, 4231–4234. [Google Scholar] [CrossRef]
- Donia, M.S.; Fricke, W.F.; Partensky, F.; Cox, J.; Elshahawi, S.I.; White, J.R.; Phillippy, A.M.; Schatz, M.C.; Piel, J.; Haygoood, M.G.; et al. Complex microbiome underlying secondary and primary metabolism in the tunicate-Prochloron symbiosis. Proc. Natl. Acad. Sci. USA 2011, 108, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.M. Ascidians (Tunicates). In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 3, pp. 1609–1731. [Google Scholar]
- Adesanya, S.A.; Chbani, M.; Pais, M.; Debitus, C. Brominated β-carbolines from the marine tunicate Eudistoma album. J. Nat. Prod. 1992, 55, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Debitus, C.; Laurent, D.; Pais, M. Alcaloides d’une ascidie néocalédonienne, Eudistoma fragum. J. Nat. Prod. 1988, 51, 799–801. (In French) [Google Scholar] [CrossRef]
- Gouiffes, D.; Juge, M.; Grimaud, N.; Welin, L.; Sauviat, M.P.; Barbin, Y.; Laurent, D.; Roussakis, C.; Hénichart, J.P.; Verbist, J.F. Bistramide A, a new toxin from the Urochordata Lissoclinum bistratum Sluiter: Isolation and preliminary characterization. Toxicon 1988, 26, 1129–1136. [Google Scholar] [CrossRef]
- Roussakis, C.; Robillard, N.; Riou, D.; Biard, J.F.; Pradal, G.; Piloquet, P.; Debitus, C.; Verbist, J.F. Effects of bistramide A on a non-small-cell bronchial carcinoma line. Cancer Chemother. Pharmacol. 1991, 28, 283–292. [Google Scholar] [PubMed]
- Biard, J.F.; Roussakis, C.; Kornprobst, J.M.; Gouiffes-Barbin, D.; Verbist, J.F.; Cotelle, P.; Foster, M.P.; Ireland, C.M.; Debitus, C. Bistramides A, B, C, D, and K: A new class of bioactive cyclic polyethers from Lissoclinum bistratum. J. Nat. Prod. 1994, 57, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Sauviat, M.P.; Gouiffes-Barbin, D.; Ecault, E.; Verbist, J.F. Blockade of sodium channels by bistramide A in voltage-clamped frog skeletal muscle fibres. Biochim. BBA-Biomembr. 1992, 1103, 109–114. [Google Scholar] [CrossRef]
- Sauviat, M.P.; Chesnais, J.M.; Choukri, N.; Diacono, J.; Biard, J.F.; Verbist, J.F. The polyether bistramide A affects the calcium sensitivity of the contractile proteins in frog atrial heart muscle. Cell Calcium 1993, 14, 301–309. [Google Scholar] [CrossRef]
- Pusset, J.; Maillere, B.; Debitus, C. Evidence that bistramide A, from the ascidian Lissoclinum bistratum Sluiter, has immunomodulating properties in vitro. J. Nat. Toxins 1996, 5, 1–6. [Google Scholar]
- Malochet-Grivois, C.; Cotelle, P.; Biard, J.F.; Hénichart, J.P.; Debitus, C.; Roussakis, C.; Verbist, J.F. Dichlorolissoclimide, a new cytotoxic labdane derivative from Lissoclinum voeltzkowi Michaelson (Urochordata). Tetrahedron Lett. 1991, 32, 6701–6702. [Google Scholar] [CrossRef]
- Malochet-Grivois, C.; Roussakis, C.; Robillard, N.; Biard, J.F.; Riou, D.; Debitus, C.; Verbist, J.F. Effects in vitro of two marine substances, chlorolissoclimide and dichlorolissoclimide, on a non-small-cell bronchopulmonary carcinoma line (NSCLC-N6). Anticancer Drug Des. 1992, 7, 493–502. [Google Scholar] [PubMed]
- Biard, J.F.; Malochet-Grivois, C.; Roussakis, C.; Cotelle, P.; Hénichart, J.P.; Debitus, C.; Verbist, J.F. Lissoclimides, cytotoxic diterpenes from Lissoclinum voeltzkowi Michaelsen. Nat. Prod. Lett. 1994, 4, 43–50. [Google Scholar] [CrossRef]
- Chbani, M.; Pais, M.; Delauneux, J.M.; Debitus, C. Brominated indole alkaloids from the marine tunicate Pseudodistoma arborescens. J. Nat. Prod. 1993, 56, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gouiffes, D.; Moreau, S.; Helbecque, N.; Bernier, J.L.; Hénichart, J.P.; Barbin, Y.; Laurent, D.; Verbist, J.F. Proton nuclear magnetic study of bistramide A, a new cytotoxic drug isolated from Lissoclinum bistratum Sluiter. Tetrahedron 1988, 44, 451–459. [Google Scholar] [CrossRef]
- Foster, M.P.; Mayne, C.L.; Dunkel, R.; Pugmire, R.J.; Grant, D.M.; Kornprobst, J.M.; Verbist, J.F.; Biard, J.F.; Ireland, C.M. Revised structure of bistramide A (bistratene A): Application of a new program for the automated analysis of 2D INADEQUATE spectra. J. Am. Chem. Soc. 1992, 114, 1110–1111. [Google Scholar] [CrossRef]
- Espada, A.; Jiménez, C.; Debitus, C.; Riguera, R. Villagorgin A and B. New type of indole alkaloids with acetylcholine antagonist activity from the gorgonian Villagorgia rubra. Tetrahedron Lett. 1993, 34, 7773–7776. [Google Scholar] [CrossRef]
- Vidal, J.P.; Escale, R.; Girard, J.P.; Rossi, J.C.; Chantraine, J.M.; Aumelas, A. Lituarines A, B, and C: A new class of macrocyclic lactones from the New Caledonian sea pen Lituaria australasiae. J. Org. Chem. 1992, 57, 5857–5860. [Google Scholar] [CrossRef]
- Clastres, A.; Ahond, A.; Poupat, C.; Potier, P.; Kan, S.K. Invertébrés Marins Du Lagon Néo-Calédonien, II. Étude Structurale De Trois Nouveaux Diterpènes Isolés Du Pennatulaire Pteroides laboutei. J. Nat. Prod. 1984, 47, 155–161. (In French) [Google Scholar] [CrossRef]
- La Barre, S.; Coll, J.C.; Sammarco, P.W. Defensive strategies of soft corals (Coelenterata: Octocorallia) of the Great Barrier Reef. II—The relationship between toxicity and feeding deterrence. Biol. Bull. 1986, 171, 565–576. [Google Scholar] [CrossRef]
- La Barre, S.; Coll, J.C.; Sammarco, P.W. Competitive strategies of soft corals (Coelenterata: Octocorallia): III. Spacing and aggressive interactions between alcyonaceans. Mar. Ecol. Prog. Ser. 1986, 28, 147–156. [Google Scholar] [CrossRef]
- Coll, J.C.; Bowden, B.F.; Heaton, A.; Scheuer, P.J.; Li, M.K.W.; Clardy, J.; Schulte, G.K.; Finer-Moore, J.S. Structures and possible functions of epoxypu-kalide and pukalide: Diterpenes associated with eggs of sinularian soft corals (Cnidaria, Anthozoa, Octocorallia, Alcyonacea, Alcyoniidae). J. Chem. Ecol. 1989, 15, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.M. Cnidaria and Ctenophora. In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 3, pp. 1087–1286. [Google Scholar]
- Kornprobst, J.M. Phaeophyceae (Brown Algae). In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 1, pp. 435–436. [Google Scholar]
- Tschesche, H; Kolkenbrock, H.; Bode, W. The covalent structure of the elastase inhibitor from Anemonia sulcata—A “non classical” Kazal-type protein. Biol. Chem. Hoppe-Seyler 1989, 368, 1297–1304. [Google Scholar]
- Cha, S. Tight-binding inhibitors-kinetic behaviour. Biochem. Pharmacol. 1975, 24, 2177–2185. [Google Scholar] [CrossRef]
- Williams, G.C. Living genera of sea pens (Coelenterata: Octocorallia: Pennatulacea): Illustrated key and synopses. Zool. J. Linn. Soc. 1995, 113, 93–140. [Google Scholar] [CrossRef]
- Lang, J.C. Interspecific aggression by scleractinian corals. II. Why the race is not only to the swift. Bull. Mar. Sci. 1973, 23, 261–279. [Google Scholar]
- Salih, A.; Larkum, A.; Cox, G.; Kühl, M.; Hoegh-Guldberg, O. Fluorescent pigments in corals are photoprotective. Nature 2000, 408, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.M. Echinodermata. In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 3, pp. 1499–1599. [Google Scholar]
- De Riccardis, F.; Iorizzi, M.; Minale, L.; Riccio, R.; de Forges, B. R.; Debitus, C. The gymnochromes: Novel marine brominated phenanthroperylenequinone pigments from the stalked crinoid Gymnocrinus richeri. J. Org. Chem. 1991, 56, 6781–6787. [Google Scholar] [CrossRef]
- Laille, M.; Gerald, F.; Debitus, C. In vitro antiviral activity on dengue virus of marine natural products. Cell. Mol. Life Sci. 1998, 54, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Palagiano, E.; De Marino, S.; Minale, L.; Riccio, R.; Zollo, F.; Iorizzi, M.; Carré, J.B.; Debitus, C.; Lucarain, L.; Provost, J. Ptilomycalin A, crambescidin 800 and related new highly cytotoxic guanidine alkaloids from the starfishes Fromia monilis and Celerina heffernani. Tetrahedron 1995, 51, 3675–3682. [Google Scholar] [CrossRef]
- Bruno, I.; Minale, L.; Riccio, R.; La Barre, S.; Laurent, D. Isolation and structure of new polyhydroxylated sterols from a deep-water starfish of the genus Rosaster. Gazz. Chim. Ital. 1990, 120, 449–451. [Google Scholar]
- Bhatnagar, S.; Dudouet, B.; Ahond, A.; Poupat, C.; Thoison, O.; Clastres, A.; Laurent, D.; Potier, P. Invertébrés marins du lagon néocalédonien. IV: Saponines et sapogénines d’une holothurie, Actinopyga flammea. Bull. Soc. Chim. Fr. 1985, 1, 124–129. (In French) [Google Scholar]
- De Riccardis, F.; Giovannitti, B.; Iorizzi, M.; Minale, L.; Riccio, R.; Debitus, C.; Richer de Forges, B. Sterol composition of the “living fossil” crinoid Gymnocrinus richeri. Comp. Biochem. Physiol. B Comp. Biochem. 1991, 100, 647–651. [Google Scholar] [CrossRef]
- Sauviat, M.P.; Benoit, A.G.; Debitus, C.; Pouny, I.; Laurent, D. Alterations of transmembrane currents in frog atrial heart muscle induced by photoexcited gymnochrome A purified from the crinoid, Gymnochrinus richeri. Photochem. Photobiol. 2001, 74, 115–119. [Google Scholar] [CrossRef]
- Laurent, D.; Baumann, F.; Benoit, A.G.; Mortelecqe, A.; Nitatpattana, N.; Desvignes, I.; Debitus, C.; Laille, M.; Gonzalez, J.P.; Chungue, E. Structure-activity relationships of dengue antiviral polycyclic quinones. South. Asian J. Trop. Med. Public Health 2005, 36, 901–905. [Google Scholar]
- Riccio, R.; Greco, O.S.; Minale, L.; La Barre, S.; Laurent, D. Starfish Saponins, Part 36. Steroidal Oligoglycosides from the Pacific Starfish Thromidia catalai. J. Nat. Prod. 1988, 51, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.M. Photosynthetic eukaryotes. In Encyclopedia of Marine Natural Products, Greatly Enlarged Edition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 1, pp. 285–504. [Google Scholar]
- Garrigue, C.; Tsuda, R.T. Catalog of marine benthic algae from New Caledonia. Micronesica 1988, 21, 53–70. [Google Scholar]
- Garrigue, C. Biomass and production of two Halimeda species in the Southwest New Caledonian lagoon. Oceano. Acta 1991, 14, 581–588. [Google Scholar]
- Laurent, D.; Garrigue, C.; Bargibant, G.; Menou, J.L.; Tirard, P. Répartition bathymétrique des Caulerpes (Chlorophyces) et corrélation avec la présence de caulerpine. In Proceedings of the 5th International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985.
- Mattio, L.; Dirberg, G.; Payri, C.E.; Andréfouët, S. Diversity, biomass and distribution pattern of Sargassum beds in the South West lagoon of New Caledonia (South Pacific). In Proceedings Nineteenth International Seaweed Symposium, Kobe, Japan, 26–31 March 2007; Springer: Dordrecht, The Netherlands, 2007; pp. 361–373. [Google Scholar]
- Mattio, L.; Payri, C.E.; Verlaque, M. Taxonomic revision and geographic distribution of the subgenus Sargassum (Fucales, Phaeophyceae) in the Western and central pacific islands based on morphological and molecular analyses. J. Phycol. 2009, 45, 1213–1227. [Google Scholar] [CrossRef]
- Vidal, J.P.; Laurent, D.; Kabore, S.A.; Rechencq, E.; Boucard, M.; Girard, J.P.; Escale, R.; Rossi, J.C. Caulerpin, caulerpicin, Caulerpa scalpelliformis: Comparative acute toxicity study. Bot. Mar. 1984, 27, 533–538. [Google Scholar] [CrossRef]
- Amade, P.; Mehiri, M.; Lewis, R.J. Outstanding Marine Biotoxins: Stx, Ttx, and Ctx; Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some chemical properties of maitotoxin, a putative calcium channel agonist isolated from a marine dinoflagellate. J. Biochem. 1988, 104, 184–187. [Google Scholar] [PubMed]
- Wang, K.K.; Nath, R.; Raser, K.J.; Hajimohammadreza, I. Maitotoxin induces calpain activation in sh-sy5y neuroblastoma cells and cerebrocortical cultures. Arch. Biochem. Biophys. 1996, 331, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Roué, M.; Gugger, M.; Golubic, S.; Amzil, Z.; Aráoz, R.; Turquet, J.; Chinain, M.; Laurent, D. Marine cyanotoxins potentially harmul to human health. Chapter 1. In Outstanding Marine Molecules—Chemistry, Biology, Analysis, 1st ed.; La Barre, S., Kornprobst, J.M., Eds.; Wiley-VCH Verlag GmbH & Co KgaA: Weinheim, Germany, 2014; pp. 3–22. [Google Scholar]
- Kumar-Roine, S.; Taiana Darius, H.; Matsui, M.; Fabre, N.; Haddad, M.; Chinain, M.; Pauillac, S.; Laurent, D. A review of traditional remedies of ciguatera fish poisoning in the pacific. Phytother. Res. 2011, 25, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Abida, H.; Ruchaud, S.; Rios, L.; Humeau, A.; Probert, I.; De Vargas, C.; Bach, S.; Bowler, C. Bioprospecting marine plankton. Mar. Drugs 2013, 11, 4594–4611. [Google Scholar] [CrossRef] [PubMed]
- Physiology and Biotechnology of Algae laboratory. AMICAL. Available online: http://wwz.ifremer.fr/pba_eng/Projects/AMICAL (accessed on 26 May 2015).
- ADECAL. Available online: http://www.adecal.nc/en/2013-10-25-03-02-53/adecal (accessed on 26 May 2015).
- Bultel-Poncé, V.; Debitus, C.; Blond, A.; Cerceau, C.; Guyot, M. Lutoside: An acyl-1-(acyl-6′-mannobiosyl)-3-glycerol isolated from the sponge-associated bacterium Micrococcus luteus. Tetrahedron Lett. 1997, 38, 5805–5808. [Google Scholar] [CrossRef]
- Bultel-Poncé, V.; Berge, J.P.; Debitus, C.; Nicolas, J.L.; Guyot, M. Metabolites from the sponge-associated bacterium Pseudomonas species. Mar. Biotechnol. 1999, 1, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Chalkiadakis, E.; Dufourcq, R.; Schmitt, S.; Brandily, C.; Kervarec, N.; Coatanea, D.; Amir, H.; Loubersac, L.; Chanteau, S.; Guezennec, J.; et al. Partial characterization of an exopolysaccharide secreted by a marine bacterium, Vibrio neocaledonicus sp. nov, from New Caledonia. J. Appl. Microbiol. 2013, 114, 1702–1712. [Google Scholar] [CrossRef] [PubMed]
- Dufourcq, R.; Chalkiadakis, E.; Fauchon, M.; Deslandes, E.; Kerjean, V.; Chanteau, S.; Petit, E.; Guezennec, J.; Dupont-Rouzeyrol, M. Isolation and partial characterization of bacteria (Pseudoalteromonas sp.) with potential antibacterial activity from a marine costal environment from New Caledonia. Lett. Appl. Microbiol. 2014, 58, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Bettucci, L.; Pietra, F.; Laurent, D.; Roquebert, M.F. Acremonium neocaledoniae, a new species from wood in the Southwestern Lagoon of New Caledonia. Mycotaxon 2000, 75, 349–356. [Google Scholar]
- Laurent, D.; Guella, G.; Roquebert, M.F.; Farinole, F.; Mancini, I.; Pietra, F. Cytotoxins, mycotoxins and drugs from a new deuteromycete, Acremonium neocaledoniae, from the southwestern lagoon of New Caledonia. Plant. Med. 2000, 66, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Craik, J. Conotoxins and Other Conopeptides. Chapter 14. In Outstanding Marine Molecules—Chemistry, Biology, Analysis, 1st ed.; La Barre, S., Kornprobst, J.M., Eds.; Wiley-VCH Verlag GmbH & Co KgaA: Weinheim, Germany, 2014; pp. 319–332. [Google Scholar]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1743–1748. [Google Scholar] [CrossRef] [PubMed]
- Brauer, A.; Kurz, A.; Sockwell, T.; Baden-Tillson, H.; Heidler, J.; Wittig, I.; Kauferstein, S.; Mebs, D.; Stöcklin, R.; Remm, M. The mitochondrial genome of the venomous cone snail Conus consors. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Conus consors. Available online: http://www.biolib.cz/en/taxonimages/id582835/?type=1 (accessed on 28 March 2015).
- Gail, R.; Rageau, J. First findings on a venomous marine fish from New Caledonia, the stonefish Synanceia verrucos. Bull. Soc. Path. Exot. Film. 1956, 49, 846–854. [Google Scholar]
- Servent, D.; Winckler-Dietrich, V.; Hu, H.-Y.; Kessler, P.; Drevet, P.; Bertrand, D.; Ménez, A. Only snake curaremimetic toxins with a fifth disulfide bond have high affinity for the neuronal α7 nicotinic receptor. J. Biol. Chem. 1997, 272, 24279–24286. [Google Scholar] [CrossRef] [PubMed]
- Obara, K.; Fuse, N.; Tsuchiya, T.; Nonomura, Y.; Ménez, A.; Tamiya, T. Sequence analysis of a cDNA encoding erabutoxin b from the sea-snake Laticauda semifasciata. Nucl. Acids Res. 1989, 17, 10490. [Google Scholar] [CrossRef] [PubMed]
- Tamiya, N.; Yagi, T. Studies on sea snake venom. Proc Jpn. Acad. Ser. B. 2011, 87, 41–52. [Google Scholar] [CrossRef]
- Ducancel, F.; Guignery-Frelat, G.; Boulain, J.C.; Ménez, A. Nucleotide sequencing and structure analysis of cDNAs encoding short-chain neurotoxins from venom glands of a sea snake (Aipisurus laevis). Toxicon 1990, 28, 119–123. [Google Scholar] [CrossRef]
- Han, S.; Siegel, D.S.; Morrison, K.C.; Hergenrother, P.J.; Movassaghi, M. Synthesis and anticancer activity of all known (−)-Agelastatin alkaloids. J. Org. Chem. 2013, 78, 11970–11984. [Google Scholar] [CrossRef] [PubMed]
- Stout, E.P.; Choi, M.Y.; Castro, J.E.; Molinski, T.F. Potent Fluorinated Agelastatin Analogues for Chronic Lymphocytic Leukemia: Design, Synthesis, and Pharmacokinetic Studies. J. Med. Chem. 2014, 57, 5085–5093. [Google Scholar] [CrossRef] [PubMed]
- Cassiano, C.; Monti, M.C.; Festa, C.; Zampella, A.; Riccio, R.; Casapullo, A. Chemical proteomics reveals heat shock protein 60 to be the main cellular target of the marine bioactive sesterterpene suvanine. ChemBioChem 2012, 13, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Di Leva, F.S.; Festa, C.; D’Amore, C.; De Marino, S.; Renga, B.; D’Auria, M.V.; Novellino, E.; Limongelli, V.; Zampella, A.; Fiorucci, S. Binding mechanism of the farnesoid X receptor marine antagonist suvanine reveals a strategy to forestall drug modulation on nuclear receptors. Design, synthesis, and biological evaluation of novel ligands. J. Med. Chem. 2013, 56, 4701–4717. [Google Scholar] [CrossRef] [PubMed]
- Furuta, A.; Salam, K.A.; Hermawan, I.; Akimitsu, N.; Tanaka, J.; Tani, H.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; et al. Identification and biochemical characterization of halisulfate 3 and suvanine as novel inhibitors of hepatitis C virus NS3 helicase from a marine sponge. Mar. Drugs 2014, 12, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Karthikeyan, C.; Hari Narayana Moorthy, N.S.; Kumar Waiker, D.; Kumar Jain, A.; Trivedi, P. Human CDC2-like kinase 1 (CLK1): A novel target for Alzheimer’s disease. Curr. Drug Targets 2014, 15, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Shin, U.; Williams, D.E.; Kozakov, D.; Hall, D.R.; Beglov, D.; Vajda, S.; Andersen, R.J.; Pelletier, J. Stimulators of translation identified during a small molecule screening campaign. Anal. Biochem. 2014, 447, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Saleem, R.S.Z.; Tepe, J.J. A concise total synthesis of hymenialdisine. Tetrahedron Lett. 2015, 56, 3011–3013. [Google Scholar] [CrossRef]
- Lu, D.; Arulmozhiraja, S.; Coote, M.L.; Rae, A.D.; Salem, G.; Willis, A.C.; Wild, S.B.; Benhenda, S.; Lallemand Breitenbach, V.; de Thé, H.; Zhai, X.; et al. Sulfur Derivatives of the Natural Polyarsenical Arsenicin A: Biologically Active, Organometallic Arsenic–Sulfur Cages Related to the Minerals Realgar and Uzonite. Organometallics 2015, 34, 829–840. [Google Scholar] [CrossRef]
- Kopf, S.; Viola, K.; Atanasov, A.G.; Jarukamjorn, K.; Rarova, L.; Kretschy, N.; Teichmann, M.; Vonach, C.; Saiko, P.; Giessrigl, B.; et al. In vitro characterisation of the anti-intravasative properties of the marine product heteronemin. Arch. Toxicol. 2013, 87, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Cassiano, C.; Esposito, R.; Tosco, A.; Zampella, A.; D’Auria, M.V.; Riccio, R.; Casapullo, A.; Monti, M.C. Heteronemin, a marine sponge terpenoid, targets TDP-43, a key factor in several neurodegenerative disorders. Chem. Commun. 2014, 50, 406–408. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Sung, P.J.; Chang, Y.L.; Pan, S.L.; Teng, C.M. Heteronemin, a Spongean Sesterterpene, Induces Cell Apoptosis and Autophagy in Human Renal Carcinoma Cells. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.Y.; Zhang, R.T.; Ma, Y.M.; Gu, M.; Liu, G.; Li, J.; Nan, F.J. Design, Synthesis, and Biological Evaluation of Ring-Opened Bengamide Analogues. ChemMedChem 2011, 6, 1555–1558. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Lu, J.P.; Ye, Q.Z. Structural analysis of bengamide derivatives as inhibitors of methionine aminopeptidases. J. Med. Chem. 2012, 55, 8021–8027. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.P.; Yuan, X.H.; Ye, Q.Z. Structural analysis of inhibition of Mycobacterium tuberculosis methionine aminopeptidase by bengamide derivatives. Eur. J. Med. Chem. 2012, 47, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Martín-Gálvez, F.; García-Ruiz, C.; Sánchez-Ruiz, A.; Valeriote, F.A.; Sarabia, F. An array of bengamide E analogues modified at the terminal olefinic position: Synthesis and antitumor properties. ChemMedChem 2013, 8, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Sarabia, F.; Martin-Galvez, F.; García-Ruiz, C.; Sánchez-Ruiz, A.; Vivar-García, C. Epi-, epoxy-, and C2-modified bengamides: Synthesis and biological evaluation. J. Org. Chem. 2013, 78, 5239–5253. [Google Scholar] [CrossRef] [PubMed]
- LaBarbera, D.V.; Modzelewska, K.; Glazar, A.I.; Gray, P.D.; Kaur, M.; Liu, T.; Grossman, D.; Harper, M.K.; Kuwada, S.K.; Moghal, N.; et al. The marine alkaloid naamidine A promotes caspase-dependent apoptosis in tumor cells. Anticancer Drugs 2009, 20, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Copp, B.R.; Fairchild, C.R.; Cornell, L.; Casazza, A.M.; Robinson, S.; Ireland, C.M. Naamidine A is an antagonist of the epidermal growth factor receptor and an in vivo active antitumor agent. J. Med. Chem. 1998, 41, 3909–3911. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, E.A.; Maers, K.; Roberts, J.; Kemami-Wangun, H.V.; Harmody, D.; Wright, A.E. The marine natural product microsclerodermin A is a novel inhibitor of the nuclear factor kappa B and induces apoptosis in pancreatic cancer cells. Investig. New Drugs 2015, 33, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Andrés, R.M.; Montesinos, M.C.; Navalón, P.; Payá, M.; Terencio, M.C. NF-κB and STAT3 inhibition as a therapeutic strategy in psoriasis: In vitro and in vivo effects of BTH. J. Investig. Dermatol. 2013, 133, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.C.; Margarucci, L.; Riccio, R.; Bonfili, L.; Mozzicafreddo, M.; Eleuteri, A.M.; Casapullo, A. Mechanistic insights on petrosaspongiolide M inhibitory effects on immunoproteasome and autophagy. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2014, 1844, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; Rodríguez-Nieto, S.; García-Caballero, M.; Medina, M.Á.; Quesada, A.R. The antiangiogenic compound aeroplysinin-1 induces apoptosis in endothelial cells by activating the mitochondrial pathway. Mar. Drugs 2012, 10, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Stuhldreier, F.; Kassel, S.; Schumacher, L.; Wesselborg, S.; Proksch, P.; Fritz, G. Pleiotropic effects of spongean alkaloids on mechanisms of cell death, cell cycle progression and DNA damage response (DDR) of acute myeloid leukemia (AML) cells. Cancer Lett. 2015, 361, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Mijares, M.R.; Ochoa, M.; Barroeta, A.; Martinez, G.P.; Suarez, A.I.; Compagnone, R.S.; Chirinos, P.; Avila, R.; De Sanctis, J.B. Cytotoxic effects of Fisturalin-3 and 11-Deoxyfisturalin-3 on Jurkat and U937 cell lines. Biomed. Pap. 2013, 157, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.A.; Liu, S.; Chen, Z.; Skau, C.; Pytynia, M.; Kovar, D.R.; Chmura, S.J.; Kozmin, S.A. Rationally simplified bistramide analog reversibly targets actin polymerization and inhibits cancer progression in vitro and in vivo. J. Am. Chem. Soc. 2010, 132, 7288–7290. [Google Scholar] [CrossRef] [PubMed]
- Herkommer, D.; Dreisigacker, S.; Sergeev, G.; Sasse, F.; Gohlke, H.; Menche, D. Design, Synthesis, and Biological Evaluation of Simplified Side Chain Hybrids of the Potent Actin Binding Polyketides Rhizopodin and Bistramide. ChemMedChem 2015, 10, 470–489. [Google Scholar] [CrossRef] [PubMed]
- Kicha, A.A.; Kalinovsky, A.I.; Malyarenko, T.V.; Ivanchina, N.V.; Dmitrenok, P.S.; Menchinskaya, E.S.; Yurchenko, E.A.; Pislyagin, E.A.; Aminin, D.L.; Huong, T.T.T.; et al. Cyclic Steroid Glycosides from the Starfish Echinaster luzonicus: Structures and Immunomodulatory Activities. J. Nat. Prod. 2015, 78, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Elmallah, M.I.; Micheau, O. Marine drugs regulating apoptosis induced by tumor necrosis factor-related apoptosis-inducing ligand (trail). Mar. Drugs 2015, 13, 6884–6909. [Google Scholar] [CrossRef] [PubMed]
- FDA Approves New Therapy for Certain Types of Advanced Soft Tissue Sarcoma. Available online: http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm468832.htm (accessed on 23 October 2015).
- Mason, C.K.; McFarlane, S.; Johnston, P.G.; Crowe, P.; Erwin, P.J.; Domostoj, M.M.; Campbell, F.C.; Manaviazar, S.; Hale, K.J.; El-Tanani, M. Agelastatin A: A novel inhibitor of osteopontin-mediated adhesion, invasion, and colony formation. Mol. Cancer Ther. 2008, 7, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Hur, W.; Cho, C.Y.; Liu, Y.; Adrian, F.J.; Lozach, O.; Bach, S.; Mayer, T.; Fabbro, D.; Meijer, L. Synthesis and target identification of hymenialdisine analogs. Chem. Biol. 2004, 11, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.; Alonso, D.; Martin-Aparicio, E.; Fuertes, A.; Perez-Puerto, M.J.; Castro, A.; Morales, S.; Navarro, M.L.; del Monte-Millan, M.; Medina, M. Glycogen synthase kinase-3 (GSK-3) inhibitory activity and structure-activity relationship (sar) studies of the manzamine alkaloids. Potential for alzheimer’s disease. J. Nat. Prod. 2007, 70, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Marine Pharmaceuticals: The Clinical Pipeline. Available online: http://marinepharmacology.midwestern.edu/clinPipeline.htm (accessed on 21 February 2016).
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Kwon, H.J.; Jung, H.S. Osteogenic potency of nacre on human mesenchymal stem cells. Mol. Cells 2015, 38, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Lopez, E.; Vidal, B.; Berland, S.; Camprasse, S.; Camprasse, G.; Silve, C. Demonstration of the capacity of nacre to induce bone formation by human osteoblasts maintained in vitro. Tissue Cell 1992, 24, 667–679. [Google Scholar] [CrossRef]
- Pernice, M; Wetzel, S.; Gros, O.; Boucher-Rodoni, N.; Dubilier, N. Enigmatic dual symbiosis in the excretory organ of Nautilus macromphalus (Cephalopoda: Nautiloidea). Proc. R. Soc. B 2007, 274, 1143–1152. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Model | Species | Type of Activity Tested | Reference |
|---|---|---|---|
| Bacteria | Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus faecalis (now Enterococcus faecalis), Vibrio anguillarum | Antibacterial | [26] |
| Fungi | Candida albicans, Candida tropicalis, Helminthosporium graminearum, Helminthosporium turcicum, Penicillium italicum, Phytophthora parasitica, Pyricularia oryzae | Antifungal | [26] |
| Brine shrimp larvae | Artemia salina | Cytotoxicity | [27] |
| Fish | Gambusia affinis | Neuro/Cytotoxicity | [28,29] |
| Urchin eggs | Echinometra mathaei | Cytotoxicity | [30] |
| Insect | Hypothenemus hampei | Insecticide | [31] |
| Mite | Rhipicephalus microplus (formerly, Boophilus microplus) | Acaricide | [32] |
| Algae | Ceramium codii | Anti-fouling | [33] |
| Coral | Algae | Allelopathy | [34] |
| Plant | Amaranthus caudatus | Anti-germinating | [35] |
| Natural Product | Chemical Class | Biological Activity | Species | Reference | |
|---|---|---|---|---|---|
| agelastatin A | unusual C11 alkaloid | antiproliferative kinase inhibitor | IC50 (µg/mL) KB 0.075, L1210 0.033 GSK-3β selective inhibitor (IC50 12 µM). | Agelas dendromorpha | [41,42,43,44] |
| ageliferin | dimeric C11 oroidin-related alkaloid | neurotransmitter inhibitor | Active on SRIF 2.21 µM (Ki 2.47 µM); VIP 19.8 µM (Ki 63.8 µM). | Agelas novaecaledoniae | [45] |
| sceptrin | Active on SRIF 0.27 µM (Ki 0.30 µM); VIP 19.2 µM (Ki 61.8 µM). | ||||
| callipeltin A | cyclic depsidecapeptide | antiproliferative | IC50 (µg/mL) P388 < 3.3, NSCLC-N6 < 1.1, NSCLC-N6 C15 > 30, NSCLC-N6 C92 < 3.3, NSCLC-N6 C98 < 3.3, E39 < 1.1, M96 < 3.3 | Callipelta sp. | [46,47] |
| antiviral | Day 6 post-infection: CD50 0.29 µg/mL and ED50 HIV-1 0.01 µg/mL (SI 29), AZT reference CD50 50 µM and ED50 30 nM. | ||||
| antifungal | Ca 30 mm/100 µg/disk. | ||||
| callipeltin B | antiproliferative antiviral | IC50 (µg/mL) P388 < 3.3, NSCLC-N6 1.3, NSCLC-N6 C15 22.5, NSCLC-N6 C92 > 30, NSCLC-N6 C98 < 3.3, E39 > 10, M96 < 3.3 Inactive against HIV-1. | [47] | ||
| callipeltin C | acyclic depsidecapeptide | antiproliferative antifungal antiviral | IC50 (µg/mL) NSCLC-N6 53.5, E39 36.1 Ca 9 mm/100 µg/disk. Inactive against HIV-1. | [47] | |
| callipeltoside A | glycoside macrolide | antiproliferative mechanism of action | IC50 (µg/mL) P388 15.26, NSCLC-N6 11.26 G1 cell cycle arrest in NSCLC-N6 cells. | [48] | |
| callipeltoside B | antiproliferative | IC50 NSCLC-N6 15.1 µg/mL. | [49] | ||
| callipeltoside C | IC50 NSCLC-N6 30.0 µg/mL. | ||||
| corallistin A | porphyrin | antiproliferative | IC50 KB 10 µg/mL. Inactive in vivo and in tubuline assay. | Corallistes sp. | [50,51] |
| coscinosulfate | sulfated sesquiterpene | antiproliferative antimitotic antibacterial | Against Jurkat and HBL100 cells (data not shown). CDC25A inhibitor: IC50 3 µM. Sa 12 mm/50 µg/disk. | Coscinoderma mathewsi | [52] |
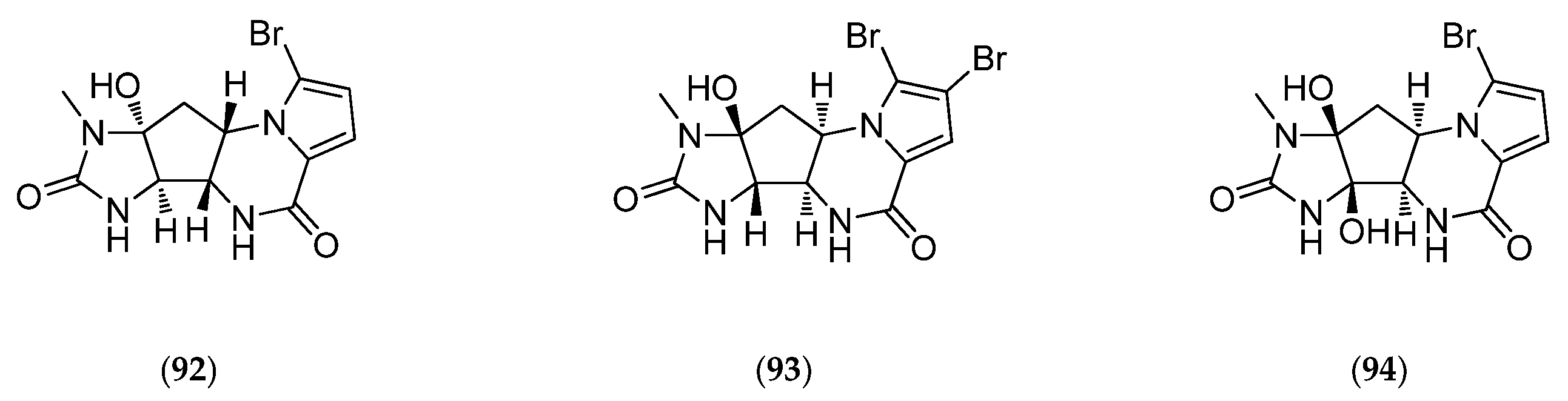
| dibromocantharelline | C11 alkaloid | kinase inhibitor | IC50 GSK3-β 3 µM. | Cymbastela cantharella (formerly Pseudaxinyssa cantharella) | [44,53] |
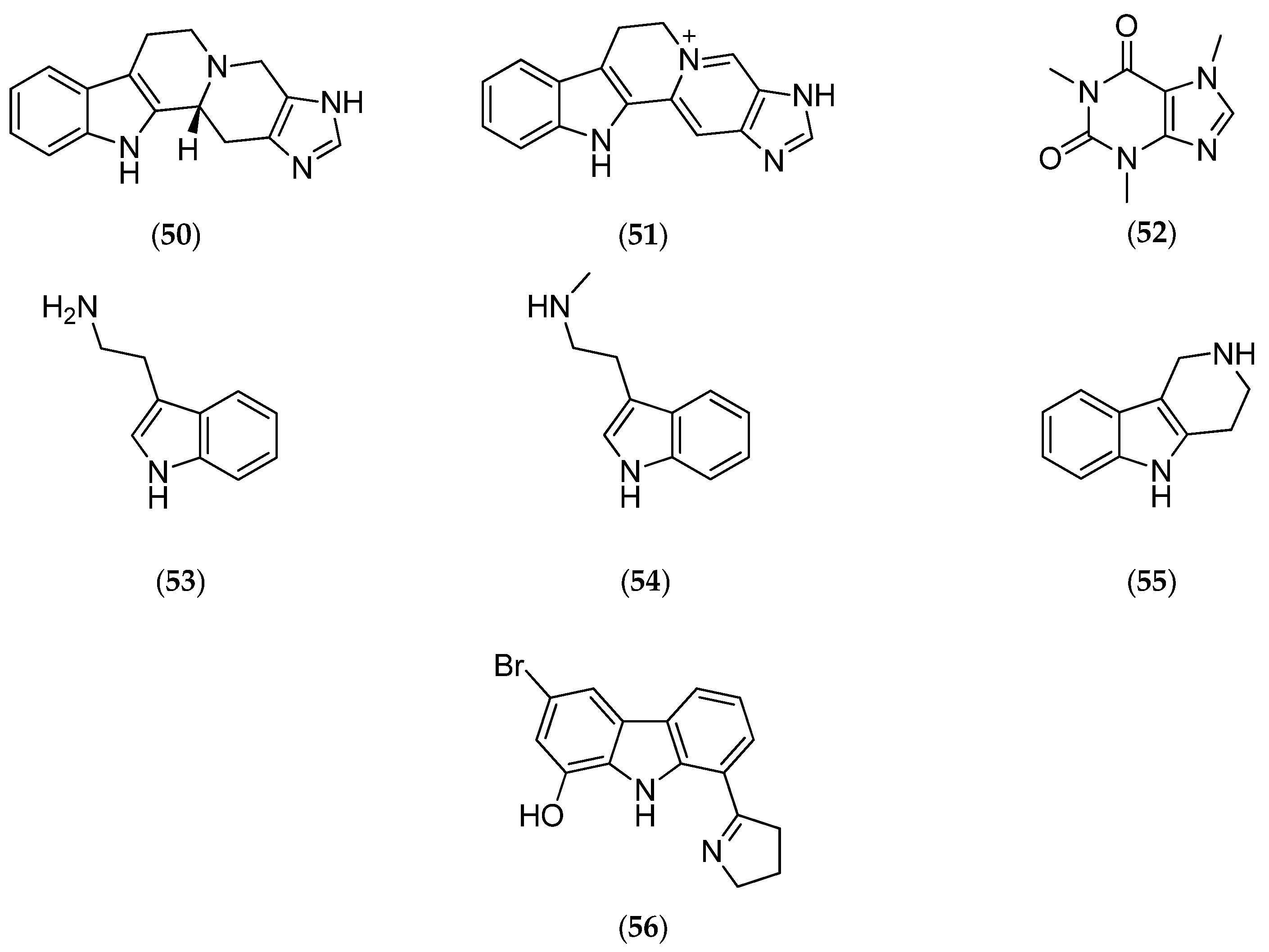
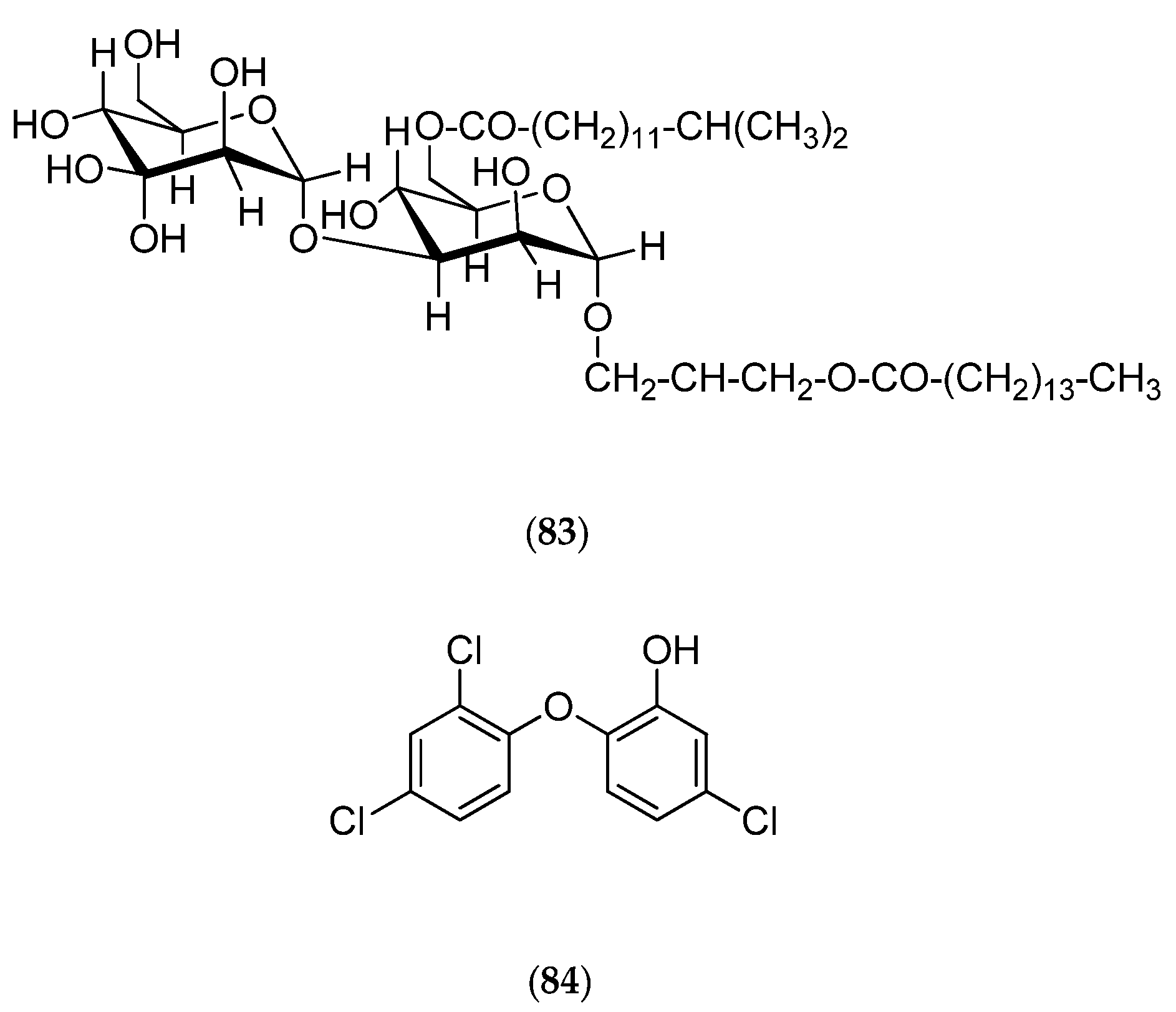
| girolline | degraded C11 alkaloid | antiproliferative antimalarial | IC50 (µM) P388 0.06, P388/Dox 0.08, KB 0.21, T24 0.19 IC50 Pf 77-215 nM. | [54,55,56,57], [58,59,60,61,62] | |
| pyraxinine | 3-pyridylguanidine | anti-inflammatory | Inhibition of macrophagic NO synthase at 100 µM. | [63] | |
| hymenialdisine | tricyclic pyrrole alkaloid | kinase inhibitor | IC50 (nM) CDK1/cyclin B 22, CDK2/cyclin A 70, CDK2/cyclin E 40, CDK5/p25 28, GSK-3β 10, CK1 35 and PLK-1 10,000. | [44,64] | |
| methyl diacarnoate A | epi-dioxy norditerpene | antiproliferative | Inactive against KB cells. | Diacarnus levii | [65] |
| methyl 3-epinuapapuanoate | antimalarial antiproliferative | IC50 7.4 and 7.2 µM on CQ-sensitive and CQ-resistant Pf, resp. IC50 KB >> 20 µg/mL. | [65,66] | ||
| 2-epimukubilin benzyl ester | epi-dioxy norsesterterpene | antiproliferative | IC50 KB 1.0 µg/mL. | [65] | |
| methyl prenyldiacarnoate A | IC50 KB 3.3 µg/mL. | ||||
| methyl 2-epiprenyldiacarnoate A | IC50 KB 0.9 µg/mL. | ||||
| nortopsentin D | bis-indole alkaloid | antiproliferative | EC50 KB 0.014 µg/mL (permethylated derivative). | Dragmacidon sp. | [67] |
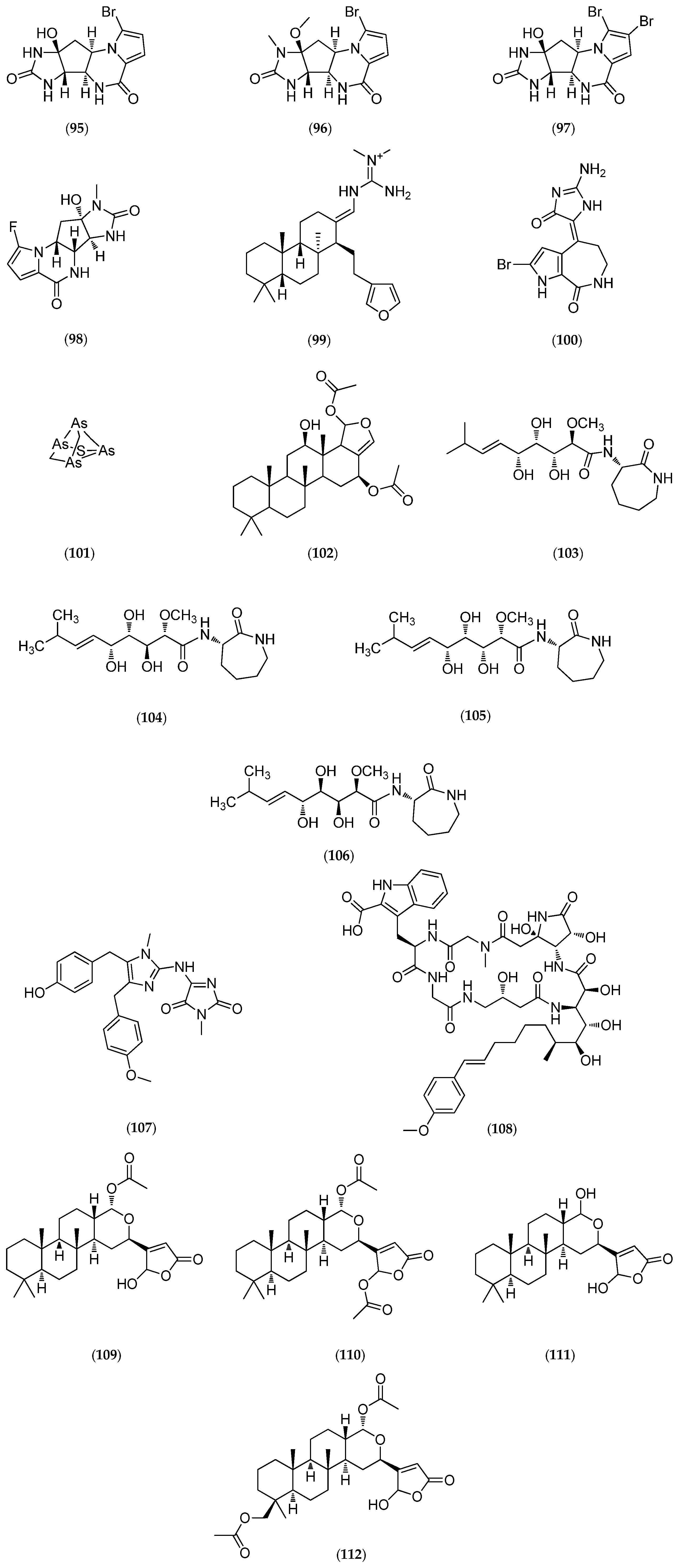
| arsenicin A | polyarsenic | antibacterial antifungal | 10 µg/disk, Sa/Ec/Ca: 24/28/26 (mm) for arsenicin A, and 22/30/22 (mm) for gentamicin. | Echinochalina bargibanti | [68] |
| euryspongiol A1 | polyhydroxylated 9,11-secosterol | antihistaminic | Reduction of histamine release by 26% (control 35%). | Euryspongia sp. | [69] |
| euryspongiol A2 | Reduction of histamine release by 15% (control 35%). | ||||
| homophymine A | cyclodepsipeptide | antiviral antiproliferative | IC50 HIV-1 75 nM. IC50 (nM) KB 7.3, MCF-7 23.6, MCF-7R 22.9, HCT116 6.0, HCT15 22.5, HT29 70.0, OVCAR8 5.4, OV3 7.5, PC3 4.2, Vero 5.0, MRC5 11.0, HL60 24.1, HL60R 22.4, K562 24.0, PaCa 31.4, SF268 9.9, A549 8.3, MDA231 10.9, MDA435 39.0, HepG2 68.6, EPC 5.0 | Homophymia sp. | [70,71] |
| homophymine B | antiproliferative mechanism of action | IC50 (nM) KB 18.0, MCF-7 16.8, MCF-7R 26.3, HCT116 13.8, HCT15 22.9, HT29 101.9, OVCAR8 8.0, OV3 9.9, PC3 6.2, Vero 8.6, MRC5 17.1, HL60 43.1, HL60R 36.7, K562 26.7, PaCa 62.0, SF268 17.2, A549 19.8, MDA231 17.0, MDA435 40.1, HepG2 99.0, EPC 8.0. Caspase-independent cell death pathway (HL60). | [71] | ||
| homophymine C | antiproliferative | IC50 (nM) KB 8.5, MCF-7 8.8, MCF-7R 10.8, HCT116 4.9, HCT15 19.2, HT29 62.8, OVCAR8 4.3, OV3 3.7, PC3 3.0, Vero 4.2, MRC5 16.8, HL60 23.0, HL60R 23.5, K562 22.5, PaCa 25.9, SF268 13.6, A549 8.3, MDA231 16.2, MDA435 35.0, HepG2 72.1, EPC 9.3 | |||
| homophymine D | IC50 (nM) KB 12.7, MCF-7 19.6, MCF-7R 37.7, HCT116 19.8, HCT15 43.2, HT29 81.3, OVCAR8 8.1, OV3 10.6, PC3 6.3, Vero 10.9, MRC5 16.9, HL60 29.6, HL60R 24.9, K562 35.3, PaCa 37.4, SF268 17.9, A549 13.8, MDA231 18.9, MDA435 49.9, HepG2 78.7, EPC 11.1 | ||||
| homophymine E | IC50 (nM) KB 6.0, MCF-7 14.2, MCF-7R 15.6, HCT116 5.5, HCT15 27.2, HT29 35.1, OVCAR8 4.6, OV3 4.2, PC3 3.9, Vero 7.0, MRC5 9.5, HL60 23.3, HL60R 21.4, K562 22.2, PaCa 18.1, SF268 8.1, A549 9.6, MDA231 13.3, MDA435 38.3, HepG2 60.5, EPC 9.5 | ||||
| homophymine A1 | IC50 (nM) KB 7.1, MCF-7 12.4, MCF-7R 13.5, HCT116 6.1, HCT15 13.5, HT29 30.9, OVCAR8 5.1, OV3 5.5, PC3 3.7, Vero 6.1, MRC5 7.8, HL60 17.3, HL60R 11.1, K562 12.8, PaCa 19.2, SF268 6.3, A549 6.0, MDA231 8.4, MDA435 27.0, HepG2 91.4, EPC 7.8 | ||||
| homophymine B1 | cyclodepsipeptide | antiproliferative | IC50 (nM) KB 16.4, MCF-7 14.2, MCF-7R 12.3, HCT116 11.4, HCT15 14.1, HT29 93.8, OVCAR8 6.5, OV3 8.0, PC3 4.7, Vero 6.1, MRC5 10.2, HL60 18.7, HL60R 25.8, K562 16.6, PaCa 22.2, SF268 11.7, A549 8.6, MDA231 18.2, MDA435 29.5, HepG2 100.3, EPC 6.6 | Homophymia sp. | [71] |
| homophymine C1 | antiproliferative mechanism of action | IC50 (nM) KB 6.8, MCF-7 6.3, MCF-7R 5.4, HCT116 2.7, HCT15 17.2, HT29 38.2, OVCAR8 2.6, OV3 2.4, PC3 2.6, Vero 3.1, MRC5 8.0, HL60 14.6, HL60R 17.1, K562 11.9, PaCa 14.4, SF268 7.1, A549 6.2, MDA231 15.8, MDA435 20.3, HepG2 58.6, EPC 12.2 Caspase-independent cell death pathway (HL60). | |||
| homophymine D1 | antiproliferative mechanism of action | IC50 (nM) KB 10.6, MCF-7 3.5, MCF-7R 3.5, HCT116 1.8, HCT15 11.4, HT29 32.2, OVCAR8 1.6, OV3 1.4, PC3 1.4, Vero 1.8, MRC5 10.5, HL60 13.1, HL60R 21.9, K562 12.9, PaCa 17.6, SF268 7.9, A549 5.0, MDA231 11.1, MDA435 23.4, HepG2 80.4, EPC 7.7 Caspase-independent cell death pathway (HL60). | |||
| homophymine E1 | antiproliferative | IC50 (nM) KB 12.5, MCF-7 3.9, MCF-7R 7.1, HCT116 2.3, HCT15 10.1, HT29 31.8, OVCAR8 4.0, OV3 2.7, PC3 3.5, Vero 4.4, MRC5 12.3, HL60 20.5, HL60R 23.2, K562 17.8, PaCa 10.6, SF268 10.1, A549 11.4, MDA231 20.0, MDA435 37.0, HepG2 62.8, EPC 29.0 | |||
| 12-epi-heteronemin | scalarane sesterterpene | farnesyltransferase inhibitor | Inactive on farnesyl transferase. | Hyrtios erecta | [72,73] |
| heteronemin | IC50 3 µM. | Hyrtios reticulata | [73] | ||
| thorectolide | terpene | antiproliferative antiviral | IC50 KB 5.3 µg/mL. HIV-1 nucleocapside and integrase inhibitor 10 and 20 µg/mL, resp. | Hyrtios sp. | [74] |
| thorectolide monoacetate | antiproliferative anti-inflammatory | IC50 KB 0.3 µg/mL. Cobra venom PLA2 inhibitor 2 µM, bee venom PLA2 inactive. | |||
| puupehenone | merosesquiterpene | antiproliferative antifungal antibacterial antimalarial | IC50 KB 0.8 µg/mL. Ct 12 mm/50 µg/disk. Sa 12 mm/50 µg/disk. IC50 (µg/mL) Pf F32 0.6, FcB1 2.1 and PFB 1.5 | [75,76] | |
| dipuupehedione | antiproliferative antifungal antibacterial | IC50 KB 3 µg/mL. Inactive against Ct. Inactive against Sa. | [75] | ||
| 15α-methoxypuupehenol | antiproliferative antibacterial antifungal antimalarial | IC50 KB 6 µg/mL. Sa 7 mm/1 µg/disk. Ct 9 mm/50 µg/disk. IC50 (µg/mL) Pf F32 0.4, FcB1 1.4 and PFB 1.2 | [76] | ||
| pentaprenylhydro-quinone 4-sulfate | terpene | NPY inhibitor kinase inhibitor antiviral | IC50 50.8 µg/mL. IC50 TPK 8 µg/mL. HIV-1 integrase: 65% inhibition at 1 µg/mL. | Ircinia sp. | [77] |
| hexaprenylhydro-quinone 4-sulfate | kinase inhibitor | IC50 TPK 4.0 µg/mL. | |||
| heptaprenylhydro-quinone 4-sulfate | IC50 TPK 8.0 µg/mL. | ||||
| leucascandrolide A | macrolide | antiproliferative antifungal | IC50 (µg/mL) KB 0.05, P388 0.25 Ca 26 mm/40 µg/disk. | Leucascandra caveolata | [78] |
| leucascandrolide B | IC50 (µg/mL) KB 5, P388 >10 Inactive against Ca. | [79] | |||
| microsclerodermin A | cyclic hexapeptide | antifungal | Ca 2.5 µg/disk. | Microscleroderma sp. | [80] |
| microsclerodermin B | |||||
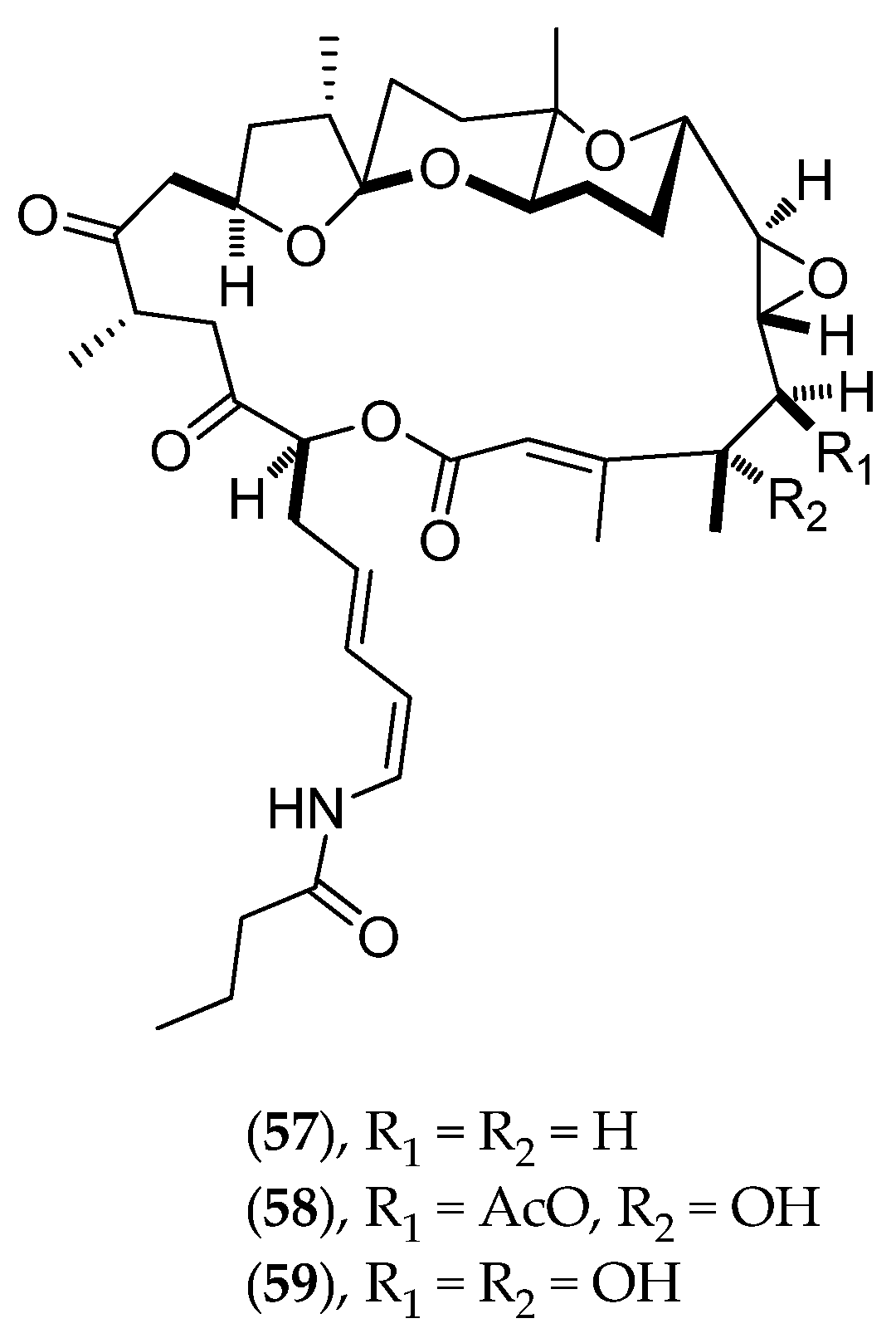
| sphinxolide | macrolide | antiproliferative antifungal | IC50 (ng/mL) NSCLC-N6 27, P388/Dox 0.33, P388 4.1, KB 7.0, HT29 115 IC90 values < 2 ppm against Pc, Pci, Pr, Pv, Bc, Po, Fr, Aa, Rs, Ph, Sn, Hg and Un. | Neosiphonia superstes | [81,82] |
| sphinxolide B | IC50 (ng/mL) KB 0.03, P388 3.1, P388/Dox 0.02, NSCLC-N6 16, HT29 2.4 IC90 values < 2 ppm against Pc, Pci, Pr, Pv, Bc, Po, Fr, Aa, Rs, Ph, Sn, Hg and Un. | ||||
| sphinxolide C | IC50 (ng/mL) KB 40, P388 40, P388/Dox 30, NSCLC-N6 30, HT29 30 IC90 values < 2 ppm against Pc, Pci, Pr, Pv, Bc, Po, Fr, Aa, Rs, Ph, Sn, Hg and Un. | ||||
| sphinxolide D | antiproliferative | IC50 (ng/mL) NSCLC-N6 60, P388/Dox 8, P388 3, KB 3.0, HT29 22 | [81] | ||
| sphinxolide E | NCI screening: 60 human cell lines (9 cancer types: leukemia, lung, colon, brain, melanoma, ovarian, renal, prostate and breast). Sphinxolides F and G: less potent by 10-100 times compared to E. Same degree of cell line selectivity. | [83] | |||
| sphinxolide F | |||||
| sphinxolide G | |||||
| superstolide A | IC50 (µg/mL) KB 0.02, P388 0.003, P388/Dox 0.02, NSCLC-N6-L16 0.04, HT29 0.04 | [84] | |||
| superstolide B | IC50 (µg/mL) KB 0.005, P388 0.003, NSCLC-N6-L16 0.039 | [85] | |||
| neosiphoniamolide A | cyclic depsipeptide | antifungal | IC90 Po and Hg 5 ppm, > 5 ppm against Pc, Pci, Pr, Pv, Bc, Fr, Aa, Rs, Ph, Sn and Un. | [82] | |
| nepheliosyne A | C47 polyoxygenated acetylenic acid | antiproliferative | IC50 (µM) K562 200, U266 170, SKM1 250, Kasumi 200 | Niphates sp. | [86] |
| nepheliosyne B | IC50 (µM) K562 150, U266 200, SKM1 > 250, Kasumi 150 | ||||
| gelliusine A | racemic tris-indole alkaloid | antiproliferative | 10 < IC50 (µg/mL) < 20 against KB, P388, P388/Dox, HT29 and NSCLC-N6 cells. | Orina sp. (formerly Gellius sp.) | [87,88] |
| serotoninergic activity | Serotonin agonist (10-100 µM). SRIF (100% displacement) and NPY (90%) active at 5 µg/mL. Bradykinin receptor 100%. Inactive on NK3, AMPA, CGRP, galanin, glycine, NT and VIP-binding assays. | ||||
| gelliusine B | racemic tris-indole alkaloid | serotoninergic activity | Bradykinin receptor 93% and NPY 62%. Inactive on NK3, AMPA, CGRP, galanin, glycine, NT and VIP-binding assays. | Orina sp. (formerly Gellius sp.) | [87,88] |
| gelliusine E | racemic bis-indole alkaloid | Inactive on serotonin receptor. SRIF (87% displacement) active at 5 µg/mL. NPY 63% and bradykinin receptor 63%. Inactive on NK3, AMPA, CGRP, galanin, glycine, NT and VIP-binding assays. | [88] | ||
| gelliusine F | Inactive on serotonin receptor. SRIF (91% displacement) active at 5 µg/mL. Bradykinin receptor 89% and NPY 67%. Inactive on NK3, AMPA, CGRP, galanin, glycine, NT and VIP-binding assays. | ||||
| petrosaspongiolide A | cheilantane-type sesterterpene | antiproliferative | IC50 NSCLC-N6 13.0 µg/mL. | Petrosaspongia nigra | [89] |
| petrosaspongiolide B | IC50 NSCLC-N6 14.8 µg/mL. | ||||
| petrosaspongiolide C | IC50 NSCLC-N6 0.5 µg/mL. | ||||
| petrosaspongiolide D | IC50 NSCLC-N6 5.2 µg/mL. | ||||
| petrosaspongiolide E | IC50 NSCLC-N6 4.5 µg/mL. | ||||
| petrosaspongiolide F | IC50 NSCLC-N6 8.7 µg/mL. | ||||
| petrosaspongiolide G | Inactive against NSCLC-N6. | ||||
| petrosaspongiolide H | IC50 NSCLC-N6 8.1 µg/mL. | ||||
| petrosaspongiolide I | IC50 NSCLC-N6 6.8 µg/mL. | ||||
| petrosaspongiolide J | IC50 NSCLC-N6 6.3 µg/mL. | ||||
| petrosaspongiolide K | IC50 NSCLC-N6 1.3 µg/mL. | ||||
| petrosaspongiolide L | IC50 NSCLC-N6 5.7 µg/mL. | ||||
| petrosaspongiolide M | anti-inflammatory | PLA2 inhibitors (10 µM) 71% bee venom, 11.5% N. naja venom, 12.3% porcine pancreas and 68.6% human synovial. | [90] | ||
| petrosaspongiolide N | PLA2 inhibitors (10 µM) 43.9% bee venom, 6.8% N. naja venom, 11.6% porcine pancreas and 44.0% human synovial. | ||||
| petrosaspongiolide P | PLA2 inhibitors (10 µM) 37.9% bee venom, 3.0% N. naja venom, 0.0% porcine pancreas and 60.9% human synovial. | ||||
| petrosaspongiolide Q | PLA2 inhibitors (10 µM) 12.5% bee venom, 4.2% N. naja venom, 0.0% porcine pancreas and 30.1% human synovial. | ||||
| petrosaspongiolide R | PLA2 inhibitors (10 µM) 18.8% bee venom, 1.0% N. naja venom, 0.8% porcine pancreas and 7.1% human synovial. | ||||
| phloeodictine A | guanidine alkaloid | antiproliferative antibacterial | IC50 KB 1.5 µg/mL. MIC (µg/mL) Sf 5, Sa 1, Ec 1, Pa 10 | Phloeodictyon sp. | [91] |
| phloeodictine B | IC50 KB 11.2 µg/mL. MIC (µg/mL) Sf > 15, Sa 3, Ec 30, Pa > 30 | ||||
| phloeodictine A1 | IC50 KB 2.2 µg/mL. 2.6:1 mixture of phloeodictine A1 and A2. MIC (µg/mL) Sa 3, Ec 3, Pa 30, Cp 30, Bf 10 and Pas 10 | [92] | |||
| phloeodictine A2 | |||||
| phloeodictine A3 | IC50 KB 3.5 µg/mL. 2.6:0.7:0.3 mixture of phloeodictine A3, A4 and A5. MIC (µg/mL) Sa 30, Ec 30, Pa > 30, Cp > 30, Bf > 30 and Pas > 30 | ||||
| phloeodictine A4 | |||||
| phloeodictine A5 | |||||
| phloeodictine A6 | IC50 KB 0.6 µg/mL. 1:1.4 mixture of phloeodictine A6 and A7. MIC (µg/mL) Sa 1, Ec 3, Pa 30, Cp 1, Bf 3 and Pas 3 | ||||
| phloeodictine A7 | |||||
| phloeodictine C1 | IC50 KB 1.8 µg/mL. 1:1 mixture of phloeodictine C1 and C2. MIC (µg/mL) Sa 3, Ec > 30, Pa > 30, Cp > 100, Bf > 100 and Pas > 100 | ||||
| phloeodictine C2 | |||||
| chondropsin A | macrolide lactam | antiproliferative | IC50 (nM) KB 1.5, HCT116 1.2, T47D 0.45, HBL100 1.7 and Chang liver 2.4 G2/M cell cycle arrest in HL60 and KB cell lines (→ apoptosis). | Psammoclemma sp. | [93] |
| 73-deoxychondropsin A | IC50 (nM) KB 0.28, HCT116 0.22, T47D 0.18, HBL100 0.60, Chang liver 0.24 G2/M cell cycle arrest in HL60 and KB cell lines (→ apoptosis). | ||||
| echinosulfonic acid D | alkaloid | IC50 KB 2 µg/mL. | [94] | ||
| psammaplysene C | bromotyrosine alkaloid | IC50 THP-1 7 µM. | [95] | ||
| psammaplysene D | |||||
| reidispongiolide A | sphinxolide-type macrolide | antiproliferative | IC50 (µg/mL) KB 0.10, P388 0.16, P388/Dox 0.01, NSCLC-N6 0.07, HT29 0.04 | Reidispongia coerulea | [83,96] |
| reidispongiolide B | IC50 (µg/mL) KB 0.06, P388 0.06, P388/Dox 0.02, NSCLC-N6 0.05, HT29 0.04 | [96] | |||
| reidispongiolide C | NCI screening: 60 human cell lines (9 cancer types: leukemia, lung, colon, brain, melanoma, ovarian, renal, prostate and breast). Same degree of cell line selectivity. | [83] | |||
| auroral 1 | unusual (C(3)-α-OH) truncated isomalabaricane triterpene | antiproliferative | IC50 KB 0.2 µg/mL. | Rhabdastrella globostellata (formerly Aurora sp.) | [97] |
| auroral 2 | |||||
| auroral 3 | IC50 KB 8.0 µg/mL. | ||||
| auroral 4 | |||||
| (+)-aeroplysinin-1 | bromotyrosine derivative | antibacterial | Active against Pecten maximus larvae. | Suberea creba | [98] |
| dibromoverongiaquinol | |||||
| demethylxestospongin B | bis-1-oxaquinolizidine alkaloid | antiproliferative somatostatin inhibitor | IC50 (µg/mL) KB 2.5, L1210 0.8 No in vivo activity against P388 leukemia cells. IP3 active: IC50 12 µM (Ki 13.4 µM). | Xestospongia exigua | [99] |
| xestospongin B | IC50 (µg/mL) KB 2.5, L1210 2.0. No in vivo activity against P388 leukemia cells. IP3 active: IC50 12 µM. | [45,99] | |||
| xestospongin D | antiproliferative | IC50 (µg/mL) KB 2.0, L1210 0.2 No in vivo activity against P388 leukemia cells. | [99] | ||
| xestoamine | β-carboline alkaloid | Inactive against KB and L1210 cell lines. | |||
| Natural Product | Chemical Class | Biological Activity | Species | Reference | |
|---|---|---|---|---|---|
| eudistalbin A | β-carboline alkaloid | antiproliferative | ED50 KB 3.2 µg/mL. | Eudistoma album | [113] |
| eudistalbin B | Inactive on KB cell line. | ||||
| eudistomin E | ED50 KB < 5 ng/mL. | ||||
| (−)-woodinine | alkaloid | antibacterial | Sa 16 mm/100 µg/disk and 18 mm/200 µg/disk. Ec 8/100 µg/disk and 11 mm/200 µg/ disk. | Eudistoma fragum | [114] |
| 5-bromo-N,N-dimethylamino-ethyltryptamine | Sa 12 mm/100 µg/disk and 17 mm/200 µg/disk. Ec 17 mm/100µg/disk and 22 mm/200 µg/disk. | ||||
| bistramide A | tetrahydropyran derivative | antiproliferative | IC50 (nM) KB 45, P388 20 and normal human endothelial cells 22 IC50 NSCLCN6-L16 0.49 µM at 67 h. IC50 (µg/mL) KB 0.53, P388 0.20, P388/Dox 0.05, B16 0.10, HT29 0.32, NSCLC-N6 0.03 | Lissoclinum bistratum | [115,116,117,118,119,120] |
| antibacterial | Inactive agaisnt Ec, Kp, Mm, Pm, Pv, Pa, Sm, Sa and Streptococcus group D (500 µg/mL). | ||||
| mechanism of action | G1 cell cycle arrest in NSCLCN6-L16 cells; polyploidy-inaptitude for cytokinesis. | ||||
| Na+ channels inhibitor | At rest and in the inactivated state, occupied a site which was not located on the inactivation gate. | ||||
| Ca2+ sensitivity immunomodulator | Binding to contractile proteins for which it competes with Ca2+. Inhibition of T cell proliferation and activation of B cell proliferation. | ||||
| bistramide B | antiproliferative mechanism of action | IC50 (µg/mL) KB 2.10, P388 0.20, P388/Dox 1.16, B16 1.20, HT29 0.71, NSCLC-N6 0.32. Significant decreases of S phase in NSCLC-N6 cells. | [117] | ||
| bistramide C | IC50 (µg/mL) KB 0.65, P388 0.02, P388/Dox 0.05, B16 0.06, HT29 0.50, NSCLC-N6 0.05. Significant decreases of S phase in NSCLC-N6 cells. | ||||
| bistramide D | IC50 (µg/mL) KB 10.00, P388 0.36, P388/Dox 5.82, B16 0.10, HT29 2.76, NSCLC-N6 3.43; In vivo (IV and IP) antitumor activity in nude mice engrafted SC with NSCLC-N6, T/C 53% at day 30. Significant decreases of S phase; partial G1 cell cycle arrest in NSCLC-N6 cells. | ||||
| bistramide K | tetrahydropyran derivative | antiproliferative mechanism of action | IC50 (µg/mL) KB > 10.00, P388 0.57, P388/Dox > 10.00, B16 1.90, HT29 5.60, NSCLC-N6 3.23; In vivo (IV and IP) antitumor activity in nude mice engrafted SC with NSCLC-N6, T/C 49% at day 30. G1 cell cycle arrest in NSCLC-N6 cells. | Lissoclinum bistratum | [117] |
| dichlorolissoclimide | nitrogenous labdane diterpene | antiproliferative mechanism of action | IC50 (ng/mL) KB 14, P388 1, P388/Dox 300 and NSCLC-N6 9; G1 cell cycle arrest in NSCLC-N6 cells (irreversible, total, dose-and time-dependent). | Lissoclinum voeltzkowi | [121,122] |
| chlorolissoclimide | antiproliferative mechanism of action | IC50 (ng/mL) KB 52, P388 1.7, P388/Dox 200 and NSCLC-N6 10; G1 cell cycle arrest in NSCLC-N6 cells. | [121,122,123] | ||
| arborescidine D | indole alkaloid | antiproliferative | IC50 KB 3 µg/mL. | Pseudodistoma arborescens | [124] |
| Natural Product | Chemical Class | Biological Activity | Species | Reference | |
|---|---|---|---|---|---|
| 7-epi-11,19-desoxyhavannahine | xenicane diterpene | anti-fouling | Ceramium codii RGR after 2 days: 4% of control at 50 ppm, 21% at 25 ppm, 42% at 12.5 ppm. | Xenia garciae | [33] |
| iela melst | protein | elastase inhibitor | Inhibitor of amidolysis of Suc(Ala)3p-NA by porcine pancreatic elastase (Ki 1.5 nM). | Melithea cf. stormii | [37] |
| villogorgin A | caffeine-xanthine type alkaloid | anti-inflammatory | Acetylcholine antagonist. Anti-aggregatory (thrombin, A23187). | Villogorgia rubra | [127] |
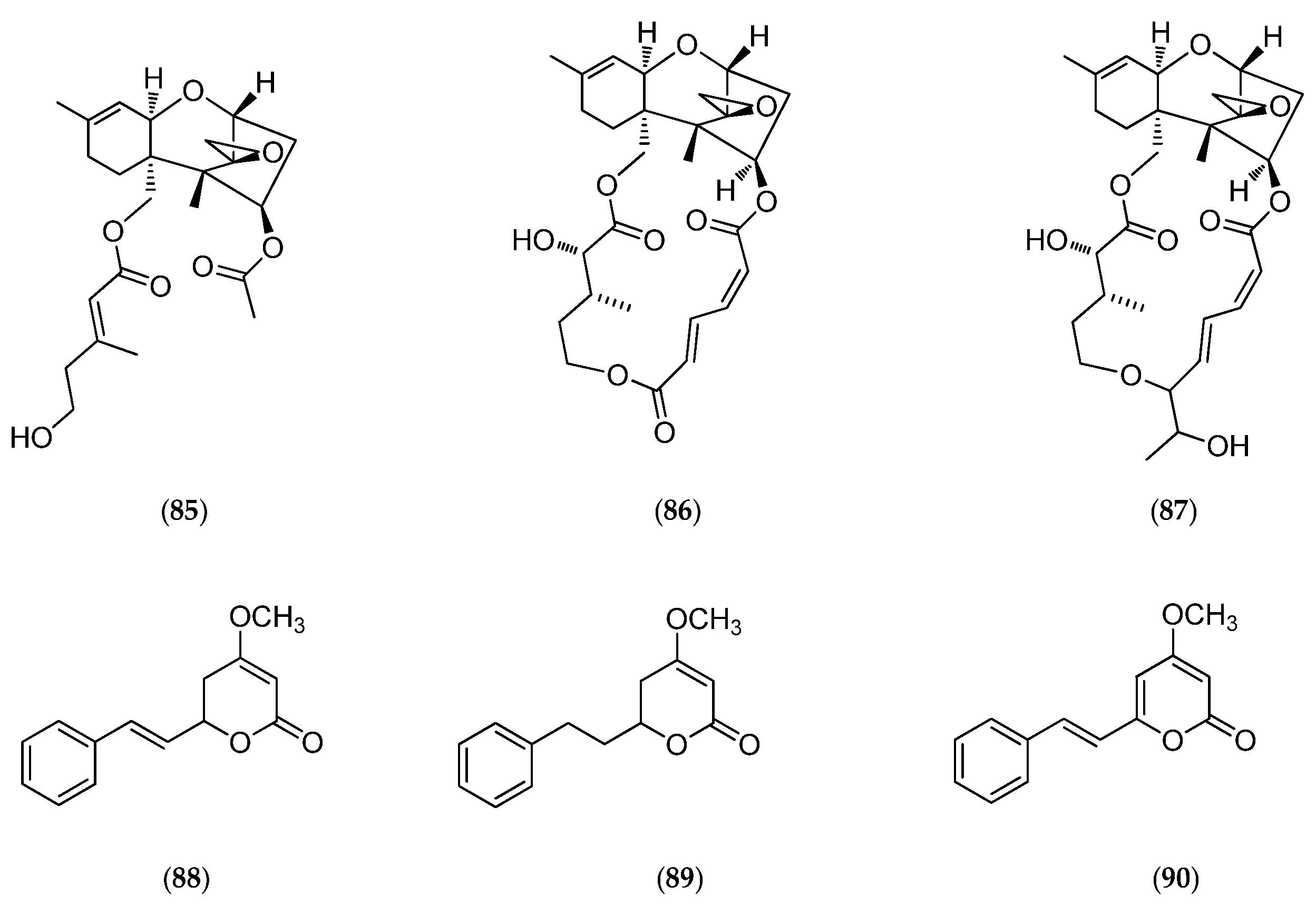
| lituarine A | polyethermacrolide | antiproliferative | IC50 KB 3.7–5 ng/mL. | Lituaria australasiae | [128] |
| lituarine B | IC50 KB 1–2 ng/mL. | ||||
| lituarine C | IC50 KB 5–6 ng/mL. | ||||
| pteroidin | briarane diterpene | ichtyotoxic | LD100 50 µg/mL, t = 90 min (fish of the genus Mugil). | Pteroides laboutei | [129] |
| O-deacetyl-12-O-benzoyl-12-pteroidin | LD100 50 µg/mL, t = 150 min (fish of the genus Mugil). | ||||
| Natural Product | Chemical Class | Biological activity | Species | Reference | |
|---|---|---|---|---|---|
| gymnochrome B | phenanthroperylene-quinone pigment | antiviral | DENV RF50% 1 µg/mL. | Gymnochrinus richeri | [141,142] |
| gymnochrome D | DENV RF50% < 1 µg/mL. | ||||
| isogymnochrome D | |||||
| ptilomycalin A | guanidine alkaloid | antiviral | IC50 HIV-1 0.11 µg/mL. DENV inactive. | Celerina heffernani | [142,143] |
| celeromycalin | IC50 HIV-1 0.32 µg/mL. DENV inactive. | ||||
| fromiamycalin | IC50 HIV-1 0.11 µg/mL. | Fromia monolis | |||
| crambescidin 800 | IC50 HIV-1 0.11 µg/mL. DENV inactive. | ||||
| (25S)-5α-cholestane-3β,4β,6β, 7α,8,15α,16β,26-octol | sterol | antifungal | Active at 5 µg against Clodosporium cucumerinum. | Rosaster sp. | [144] |
| Natural Product and/or Analogs | Chemical Synthesis and Biological Activity | Reference |
|---|---|---|
| (−)-agelastatin A–F | -Enantioselective total synthesis involving late-stage C-ring formation. -(−)-agelastatin A: antiproliferative activity highly potent on blood cancer cell lines (CEM EC50 20 nM; Jurkat EC50 74 nM; Daudi EC50 20 nM; HL-60 EC50 138 nM; CA46 EC50 187 nM) vs. normal red blood cells (EC50 > 333 μM); dose-dependent induction of apoptosis (G2/M phase arrest), no effect on tubulin dynamics. | [181] |
| agelastatin A/13-debromo-13-trifluoromethyl agelastatin A | -In vitro and in vivo antiproliferative activity: CLL patient (CLL1 and CLL2) and JVM-2 cell line (EC50 0.064 µM and 0.16 µM respectively). -Chemical modifications outside the pyrrole ring result in significant loss in activity. -Importance of an electronegative functional group at position C-13 for CLL activity. | [182] |
| suvanine | -Anti-inflammatory activity, Hsp 60 inhibitor. | [183] |
| -Antagonist of farnesoid-X-receptor (FXR). -Identification of conformational changes responsible for agonist/antagonist form on FXR using suvanine as template. | [184] | |
| -Inhibitor of hepatitis C virus NS3 helicase (IC50 of 3 µM). -Inhibition of ATPase, RNA binding, and serine protease activities of NS3 helicase with IC50 7, 3, and 34 µM, respectively. -Interaction with an allosteric site in NS3 rather than binding to the catalytic core. | [185] | |
| isohymenialdisine and hymenialdisine | -Inhibition of CLK kinases (hymenialdisine only). | [186] |
| -Stimulation of translation: isohymenialdisine and hymenialdisine act on PKR (RNA-dependent protein kinase) by inhibiting its autophosphorylation and pertub the PKR-eIF2α phosphorylation axis; models indicate that it fits in the PKR ATP binding site. | [187] | |
| -Short (6 steps), concise, and high yielding (44%) total synthesis of hymenialdisine will enable the synthesis of novel libraries for subsequent SAR studies. | [188] | |
| arsenicin A | -Synthesis of sulfur-derivatives. -Antiproliferative activity of monosulfide (±)-arsenicin A (more potent on APL cells than (±)-arsenicin A and arsenic (III) oxide); induction of apoptosis. | [189] |
| heteronemin | -Anti-intravasative properties. | [190] |
| -Targets TDP-43, binds to specific sequences on DNA and RNA. | [191] | |
| -Antiproliferative (A498 EC50 1.57 μM). -Induction of both apoptosis and autophagy (heteronemin inhibits the phosphorylation of ERK and AKT signaling pathways and increases the phosphorylation of p38 and JNK). | [192] | |
| bengamides and analogs | -Synthesis of novel caprolactam-ring-opened bengamide analogs with antitumor activity on MDA-MB-435 (compounds 3a (EC50 4 nM) and 2i (EC50 9 nM) more potent activity than LAF389, analog of bengamide (EC50 40 nM) and the original caprolactam analog 10’ (EC50 17 nM). -Improved water solubility. | [193] |
| Methionine aminopeptidases inhibitors (HsMetAP1 and HsMetAP2). | [194] | |
| -Synthesis of a series of inhibitors of methionine aminopeptidases of Mycobacterium tuberculosis. -New X-ray structures of MtMetAP1c in complex with inhibitors in Mn(II) and Ni(II) forms; all amide moieties bind to the unique shallow cavity and interact with flat surface created by His-212 of MtMetAP1c in the Mn(II) form. Influence of active site metal on binding mode (amide takes on a different conformation in the Ni(II) form). | [195] | |
| -Analogs of bengamide E with antiproliferative properties (modified at the terminal olefinic position). -More potent activity: compound 56 (a cyclopentyl group replaced the isopropyl group at the terminal olefinic position). | [196] | |
| -Synthesis of stereoisomers of bengamide E (2,3-bis-epi- and the 2-epianalogues), a collection of C2-modified analogues, and various epoxy bengamides. -Stereochemistry at C2 and C3 positions and methoxyl group at C2: essential for retaining the cytotoxic potency. | [197] | |
| naamidine A | -Antitumor agent; induction of apoptosis (caspase-dependent). | [198,199] |
| microsclerodermin A | -Inhibition of NFκB transcriptional activity, reduced levels of phosphorylated (active) NFκB in the AsPC-1 cell line. -Antiproliferative activities: AsPC-1, BxPC-3, MIA PaCa-2 and PANC-1 pancreatic cancer cell lines. -Induction of apoptosis in the AsPC-1 mediated by GSK-3β pathway. | [200] |
| petrosaspongiolide M and analogs | -Synthetic derivative of benzo[b]thiophen-2-yl-3-bromo-5-hydroxy-5H-furan-2-one. -In vitro and in vivo potent anti-inflammatory activity (inhibition of NF-κB signaling pathway and STAT3 phosphorylation). | [201] |
| -Proteasome inhibitor. -Inhibitory activity by binding the active sites in the inner core of the immunoproteasome and/or covalently linking a Lys residue at the proteasome core/11S activator particle interface. -Modulation of intracellular proteolysis through a dual inhibition of the immunoproteasome and autophagy. | [202] | |
| aeroplysinin-1 | -Induction of apoptosis in endothelial cells (caspase dependent). | [203] |
| -Antiproliferative effect on acute myeloid leukemia (AML) cells (dose-dependent EC50 10–20 μM); induction of apoptosis. -Agent-specific pleiotropic effects. | [204] | |
| fistularin-3 | -Antiproliferative activity dose- and time-dependent (EC50 7.39 and 8.10 μM for Jurkat E6.1 and U937 resp.); Pro-apoptotic. | [205] |
| Analogs of bistramide A | -Design and synthesis of analog of bistramide A targeting cytoskeletal organization of cancer cells in vivo (combination of reversible G-actin binding and effective F-actin severing). -Potent and reversible binding of monomeric actin (Kd 9.0 nM), in vitro depolymerization of actin; in vitro and in vivo inhibition of A549 cells. | [206] |
| -Computational analyses of non-covalent actin-inhibitor interactions (AutoDock and DrugScore scoring function). -Design of a novel, highly modular class of hybrid analogs with a rationale to address both the bistramide and rhizopodin binding sites. -Antiproliferative activity is conserved in analogs resembling the original side chain of rhizopodin. | [207] | |
| luzonicoside A | -Potent immunomodulatory agent: more effective in stimulating lysosomal activity, intracellular ROS level elevation, and NO synthesis up-regulation in RAW 264.7 murine macrophages cells (0.01–0.1 μM). | [208] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motuhi, S.-E.; Mehiri, M.; Payri, C.E.; La Barre, S.; Bach, S. Marine Natural Products from New Caledonia—A Review. Mar. Drugs 2016, 14, 58. https://doi.org/10.3390/md14030058
Motuhi S-E, Mehiri M, Payri CE, La Barre S, Bach S. Marine Natural Products from New Caledonia—A Review. Marine Drugs. 2016; 14(3):58. https://doi.org/10.3390/md14030058
Chicago/Turabian StyleMotuhi, Sofia-Eléna, Mohamed Mehiri, Claude Elisabeth Payri, Stéphane La Barre, and Stéphane Bach. 2016. "Marine Natural Products from New Caledonia—A Review" Marine Drugs 14, no. 3: 58. https://doi.org/10.3390/md14030058
APA StyleMotuhi, S.-E., Mehiri, M., Payri, C. E., La Barre, S., & Bach, S. (2016). Marine Natural Products from New Caledonia—A Review. Marine Drugs, 14(3), 58. https://doi.org/10.3390/md14030058