
Heterotrophic Production of Omega-3 Long-Chain Polyunsaturated Fatty Acids by Trophically Converted Marine Diatom Phaeodactylum tricornutum

Abstract
:
1. Introduction
2. Results and Discussion
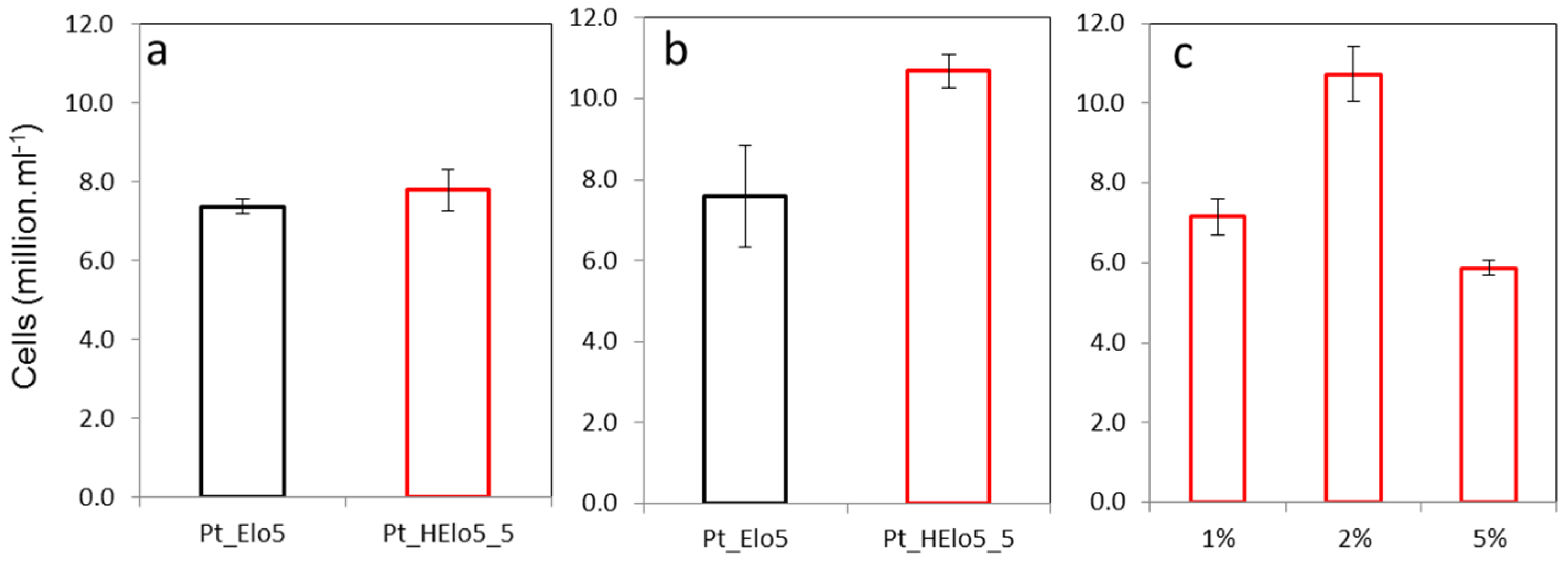
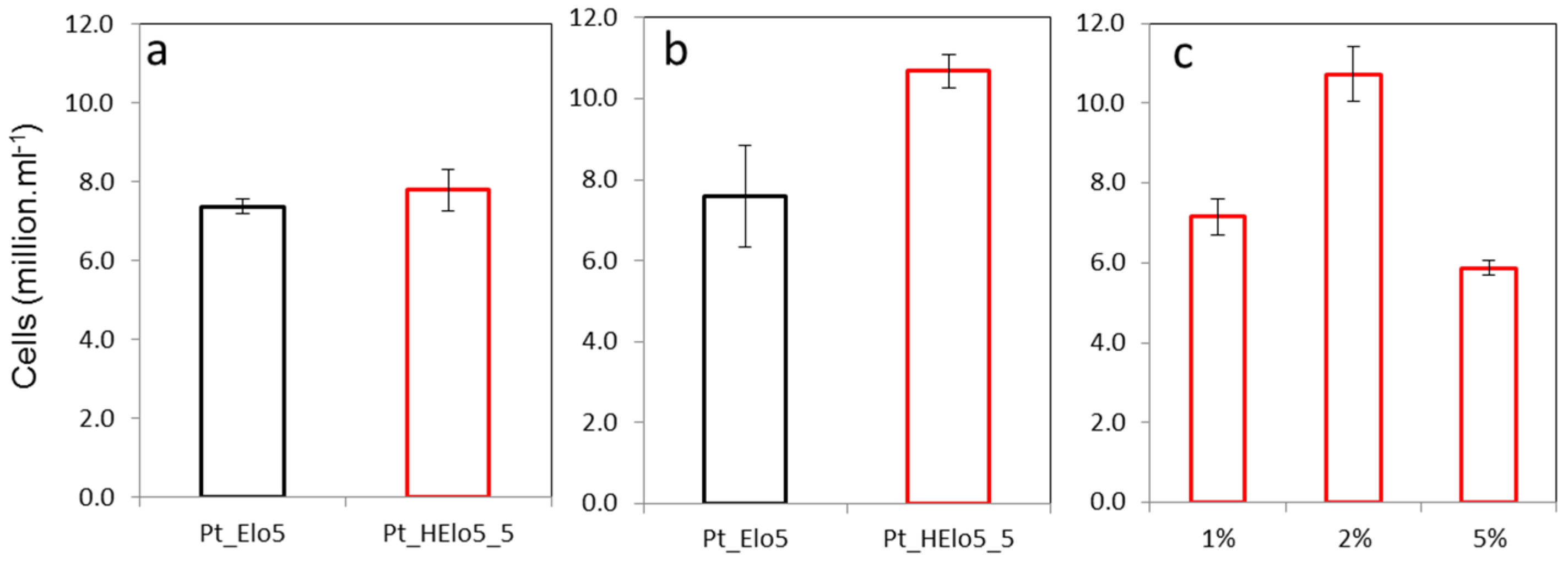
2.1. Trophic Conversion of Transgenic Pt_Elo5 Strain Accumulating Enhanced Levels of DHA
2.2. Impact of a Glucose on Cell Growth
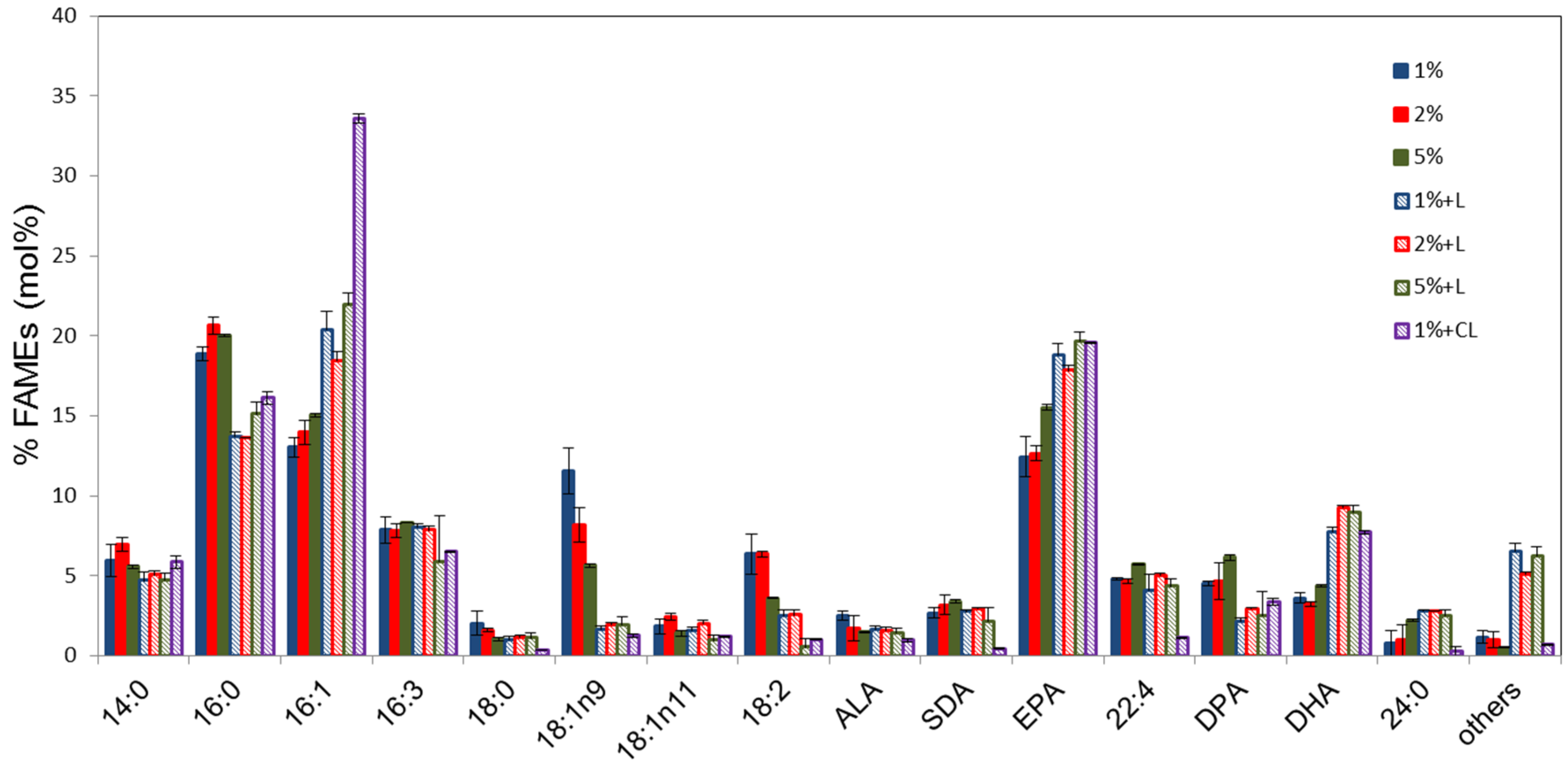
2.3. Analyses of Fatty Acid Composition of Pt_HElo5_5 Cells Grown at Different Glucose Concentrations
2.4. Quantification of LC-PUFAs Accumulation in Cells under Photo-, Mixo- and Heterotrophic Conditions
2.5. Comparison of Lipid-Bound FAs in Cells Grown under Photo, Mixo and Heterotrophic Conditions
3. Experimental Section
3.1. Strains and Growth Conditions
3.2. Plasmid Design and Transformation
3.3. Biomass Determination
3.4. Fatty Acid Analysis
3.5. Lipid Analysis
3.6. Quantitative Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Borowitzka, M.A. High-value products from microalgae-their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Benemann, J. Microalgae for biofuels and animal feeds. Energies 2013, 6, 5869–5886. [Google Scholar] [CrossRef]
- Calder, P.C. The role of marine omega-3 (n-3) fatty acids in inflammatory processes, atherosclerosis and plaque stability. Mol. Nutr. Food Res. 2012, 56, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Gharami, K.; Das, M.; Das, S. Essential role of docosahexaenoic acid towards development of a smarter brain. Neurochem. Int. 2015, 89, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Influences of culture temperature on the growth, lipid content and fatty acid composition of Aurantiochytrium sp. strain mh0186. Mar. Biotechnol. 2009, 11, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Barclay, W.R. Method of Aquaculture Comprising Feeding Microflora Having a Small Cell Aggregate Size. U.S. Patent 5,688,500, 18 November 1997. [Google Scholar]
- Burja, A.M.; Radianingtyas, H.; Windust, A.; Barrow, C.J. Isolation and characterization of polyunsaturated fatty acid producing thraustochytrium species: Screening of strains and optimization of omega-3 production. Appl. Microbiol. Biotechnol. 2006, 72, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Kyle, D.J. The large scale production of a single cell oil which is highly enriched in docosahexaenoic acid. Abstr. Pap. Am. Chem. Soc. 1999, 217, AGFD 008. [Google Scholar]
- Martins, D.A.; Custodio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Liu, Y.; Cheng, J.J.; Mos, M.; Daroch, M. Biological potential of microalgae in china for biorefinery-based production of biofuels and high value compounds. New Biotechnol. 2015, 32, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Martínez, F.; Orús, M.I. Interactions between glucose and inorganic carbon metabolism in Chlorella vulgaris strain UAM 101. Plant Physiol. 1991, 95, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Lowrey, J.; Brooks, M.S.; McGinn, P.J. Heterotrophic and mixotrophic cultivation of microalgae for biodiesel production in agricultural wastewaters and associated challenges—A critical review. J. Appl. Phycol. 2015, 27, 1485–1498. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Rohit, M.V.; Chiranjeevi, P.; Chandra, R.; Navaneeth, B. Heterotrophic microalgae cultivation to synergize biodiesel production with waste remediation: Progress and perspectives. Bioresour. Technol. 2015, 184, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Bumbak, F.; Cook, S.; Zachleder, V.; Hauser, S.; Kovar, K. Best practices in heterotrophic high-cell-density microalgal processes: Achievements, potential and possible limitations. Appl. Microbiol. Biotechnol. 2011, 91, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Doucha, J.; Livansky, K. Productivity, CO2/O2 exchange and hydraulics in outdoor open high density microalgal (Chlorella sp.) photobioreactors operated in a middle and southern european climate. J. Appl. Phycol. 2006, 18, 811–826. [Google Scholar] [CrossRef]
- Zaslavskaia, L.A.; Lippmeier, J.C.; Shih, C.; Ehrhardt, D.; Grossman, A.R.; Apt, K.E. Trophic obligate conversion of an photoautotrophic organism through metabolic engineering. Science 2001, 292, 2073–2075. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.L.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab. Eng. 2014, 22, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Kirsch, H.; Domergue, F.; Abbadi, A.; Sperling, P.; Bauer, J.; Cirpus, P.; Zank, T.K.; Moreau, H.; Roscoe, T.J.; et al. Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis. J. Lipid Res. 2004, 45, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Guiheneuf, F.; Mimouni, V.; Tremblin, G.; Ulmann, L. Light intensity regulates LC-PUFA incorporation into lipids of Pavlova lutheri and the final desaturase and elongase activities involved in their biosynthesis. J. Agric. Food Chem. 2015, 63, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Chauton, M.S.; Winge, P.; Brembu, T.; Vadstein, O.; Bones, A.M. Gene regulation of carbon fixation, storage, and utilization in the diatom Phaeodactylum tricornutum acclimated to light/dark cycles. Plant Physiol. 2013, 161, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, O.; Escalante, F.M.; de-Bashan, LE.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Ceron-Garcia, M.C.; Fernandez-Sevilla, J.M.; Sanchez-Miron, A.; Garcia-Camacho, F.; Contreras-Gomez, A.; Molina-Grima, E. Mixotrophic growth of Phaeodactylum tricornutum on fructose and glycerol in fed-batch and semi-continuous modes. Bioresour. Technol. 2013, 147, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Gouveia, L.; Veloso, V.; Fernandes, H.L.; Empis, J.A.; Novais, J.M. Eicosapentaenoic acid-rich biomass production by the microalga Phaeodactylum tricornutum in a continuous-flow reactor. Bioresour. Technol. 1996, 55, 83–88. [Google Scholar] [CrossRef]
- Garces, M.; Mancha, R. One-step lipid extraction and fatty acid methyl esters preparation from fresh plant tissues. Anal. Biochem. 1993, 211, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, N.; Haslam, R.P.; Usher, S.L.; Napier, J.A.; Sayanova, O. Reconstitution of EPA and DHA biosynthesis in Arabidopsis: Iterative metabolic engineering for the synthesis of n-3 LC-PUFAs in transgenic plants. Metab. Eng. 2013, 17, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Chen, F. Growing phototrophic cells without light. Biotechnol. Lett. 2006, 28, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Grima, E.M.; Fernández, F.A.; Camacho, F.G.; Chisti, Y. Photobioreactors: Light regime, mass transfer, and scaleup. J. Biotechnol. 1999, 70, 231–247. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| 14:0 | 16:0 | 16:1 | 16:3 | 18:0 | 18:1 | 18:2 | EPA | DPA | DHA | Others | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| WT + L | 5.2 ± 0.1 | 14.6 ± 0.2 | 38.5 ± 1.4 | 2.7 ± 0.1 | 0.4 ± 0.1 | 7.4 ± 1.1 | 0.3 ± 0.2 | 24.7 ± 0.6 | nd | 1.8 ± 0.1 | 2.7 ± 0.1 |
| Pt_Elo5 + L | 5.1 ± 0.2 | 14.1 ± 0.5 | 35.1 ± 1.2 | 7.7 ± 0.3 | 1.5 ± 0.1 | 2.5 ± 0.1 | 0.7 ± 0.1 | 15.9 ± 0.5 | 1.9 ± 0.3 | 6.3 ± 0.1 | 11.6 ± 1.7 |
| Pt_HElo5_5 + L | 5.7 ± 1.0 | 15.6 ± 0.4 | 32.6 ± 0.6 | 6.3 ± 0.8 | 0.3 ± 0.1 | 2.8 ± 0.5 | 1.0 ± 0.1 | 19.0 ± 1.2 | 3.3 ± 0.2 | 7.5 ± 0.3 | 5.1 ± 0.4 |
| Pt_HElo5_5 − L | 7.1 ± 0.9 | 23.4 ± 1.3 | 22.4 ± 3.8 | 8.2 ± 1.8 | 2.0 ± 0.7 | 11.7 ± 2.9 | 6.3 ± 1.8 | 19.9 ± 2.6 | 5.2 ± 0.6 | 7.3 ± 1.8 | 14.2 ± 2.3 |
| Pt_HElo5_7 + L | 6.1 ± 0.5 | 11.8 ± 0.7 | 34.5 ± 3.1 | 7.0 ± 0.5 | 1.4 ± 0.2 | 1.8 ± 0.3 | 0.7 ± 0.1 | 11.9 ± 0.7 | 1.8 ± 1.1 | 8.1 ± 0.7 | 7.9 ± 0.4 |
| Pt_HElo5_7 − L | 9.5 ± 0.6 | 22.1 ± 0.2 | 13.8 ± 0.1 | 7.1 ± 0.1 | 1.0 ± 0.1 | 7.7 ± 0.1 | 1.4 ± 0.1 | 13.9 ± 0.1 | 9.1 ± 2.3 | 5.2 ± 0.3 | 5.7 ± 0.2 |
| F-test (p-values) | 0.217 | 0.002 | 0.045 | <0.001 | <0.001 | <0.001 | 0.159 | <0.001 | <0.001 | <0.001 | <0.001 |
| SED | - * | 0.70 | 2.58 | 0.40 | 0.20 | 0.79 | - | 0.72 | 0.18 | 0.81 | 0.19 |
| df | - | 8 | 8 | 8 | 8 | 8 | - | 8 | 6 | 8 | 8 |
| LSD (5%) | - | 1.60 | 5.96 | 0.91 | 0.40 | 1.83 | - | 1.65 | 0.44 | 1.87 | 0.43 |
| Cells | LC-PUFA μg/mg Dry Weight | ||
|---|---|---|---|
| EPA | DPA | DHA | |
| Pt_Elo5/L | 6.0 ± 0.8 | 1.5 ± 0.1 | 3.6 ± 0.8 |
| Pt_HElo5_5/L | 6.0 ± 1.0 | 1.1 ± 0.3 | 2.5 ± 0.3 |
| Pt_Elo5/L + 1% G | 4.7 ± 0.2 | 1.1 ± 0.1 | 2.8 ± 0.2 |
| Pt_HElo5_5/L + 1% G | 8.0 ± 0.5 | 1.4 ± 0.1 | 3.1 ± 0.2 |
| Pt_HElo5_5/−L + 1% G | 3.4 ± 0.2 | 1.3 ± 0.3 | 1.0 ± 0.1 |
| Pt_HElo5_5/−L + 2% G | 3.6 ± 0.1 | 1.7 ± 0.1 | 1.0 ± 0.1 |
| Pt_HElo5_5/−L + 5% G | 3.3 ± 0.4 | 1.3 ± 0.2 | 1.0 ± 0.1 |
| F-test (p-values) | <0.001 | 0.302 | <0.001 |
| SED | 0.76 | - * | 0.47 |
| df | 14 | - | 14 |
| LSD (5%) | 1.63 | - | 1.00 |
| Strain | NL | GL | PL | ||||||
|---|---|---|---|---|---|---|---|---|---|
| EPA | DPA | DHA | EPA | DPA | DHA | EPA | DPA | DHA | |
| Pt_Elo5/L | 23.8 ± 7.4 | 16.2 ± 7.7 | 12.9 ± 3.0 | 58.9 ± 6.4 | 7.7 ± 4.2 | 8.5 ± 2.5 | 17.3 ± 4.0 | 76.1 ± 12.3 | 78.6 ± 17.1 |
| Pt_HElo5_5/ L | 36.5 ± 4.8 | 32.9 ± 4.6 | 23.6 ± 1.7 | 48.1 ± 9.6 | 12.8 ± 2.3 | 13.9 ± 1.3 | 15.4 ± 4.5 | 54.3 ± 24.6 | 62.5 ± 6.9 |
| Pt_Elo5/ L + G | 20.2 ± 2.3 | 14.5 ± 2.3 | 11.9 ± 2.3 | 63.9 ± 12.0 | 5.9 ± 0.9 | 6.7 ± 1.4 | 15.9 ± 1.6 | 79.5 ± 8.3 | 81.4 ± 13.5 |
| Pt_HElo5_5/L + G | 18.7 ± 0.8 | 17.3 ± 0.7 | 12.5 ± 0.1 | 56.1 ± 2.1 | 6.5 ± 0.2 | 7.1 ± 0.2 | 25.2 ± 1.0 | 76.1 ± 4.0 | 80.4 ± 2.3 |
| Pt_HElo5_5/−L + G | 19.1 ± 2.5 | 16.1 ± 2.2 | 10.9 ± 0.8 | 56.8 ± 0.1 | 11.8 ± 1.6 | 14.9 ± 3.8 | 24.1 ± 5.9 | 72.2 ± 5.9 | 74.1 ± 16.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, M.L.; Powers, S.; Napier, J.A.; Sayanova, O. Heterotrophic Production of Omega-3 Long-Chain Polyunsaturated Fatty Acids by Trophically Converted Marine Diatom Phaeodactylum tricornutum. Mar. Drugs 2016, 14, 53. https://doi.org/10.3390/md14030053
Hamilton ML, Powers S, Napier JA, Sayanova O. Heterotrophic Production of Omega-3 Long-Chain Polyunsaturated Fatty Acids by Trophically Converted Marine Diatom Phaeodactylum tricornutum. Marine Drugs. 2016; 14(3):53. https://doi.org/10.3390/md14030053
Chicago/Turabian StyleHamilton, Mary L., Stephen Powers, Johnathan A. Napier, and Olga Sayanova. 2016. "Heterotrophic Production of Omega-3 Long-Chain Polyunsaturated Fatty Acids by Trophically Converted Marine Diatom Phaeodactylum tricornutum" Marine Drugs 14, no. 3: 53. https://doi.org/10.3390/md14030053
APA StyleHamilton, M. L., Powers, S., Napier, J. A., & Sayanova, O. (2016). Heterotrophic Production of Omega-3 Long-Chain Polyunsaturated Fatty Acids by Trophically Converted Marine Diatom Phaeodactylum tricornutum. Marine Drugs, 14(3), 53. https://doi.org/10.3390/md14030053