
Determination of Lipid Hydroperoxides in Marine Diatoms by the FOX2 Assay


 ,
, 

Abstract
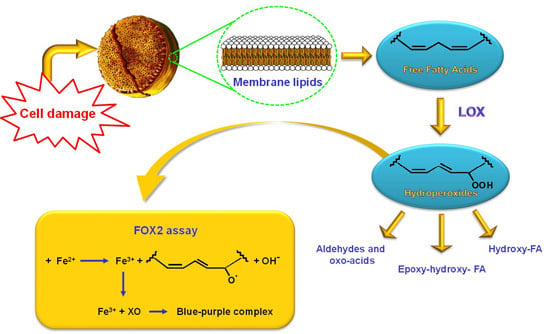
:
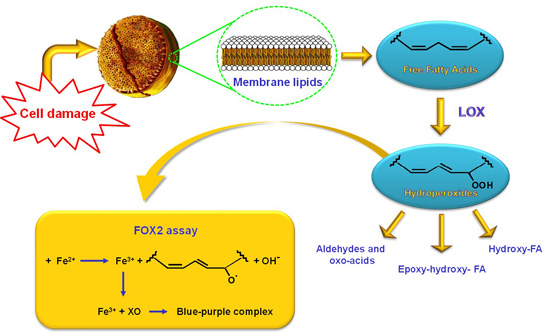
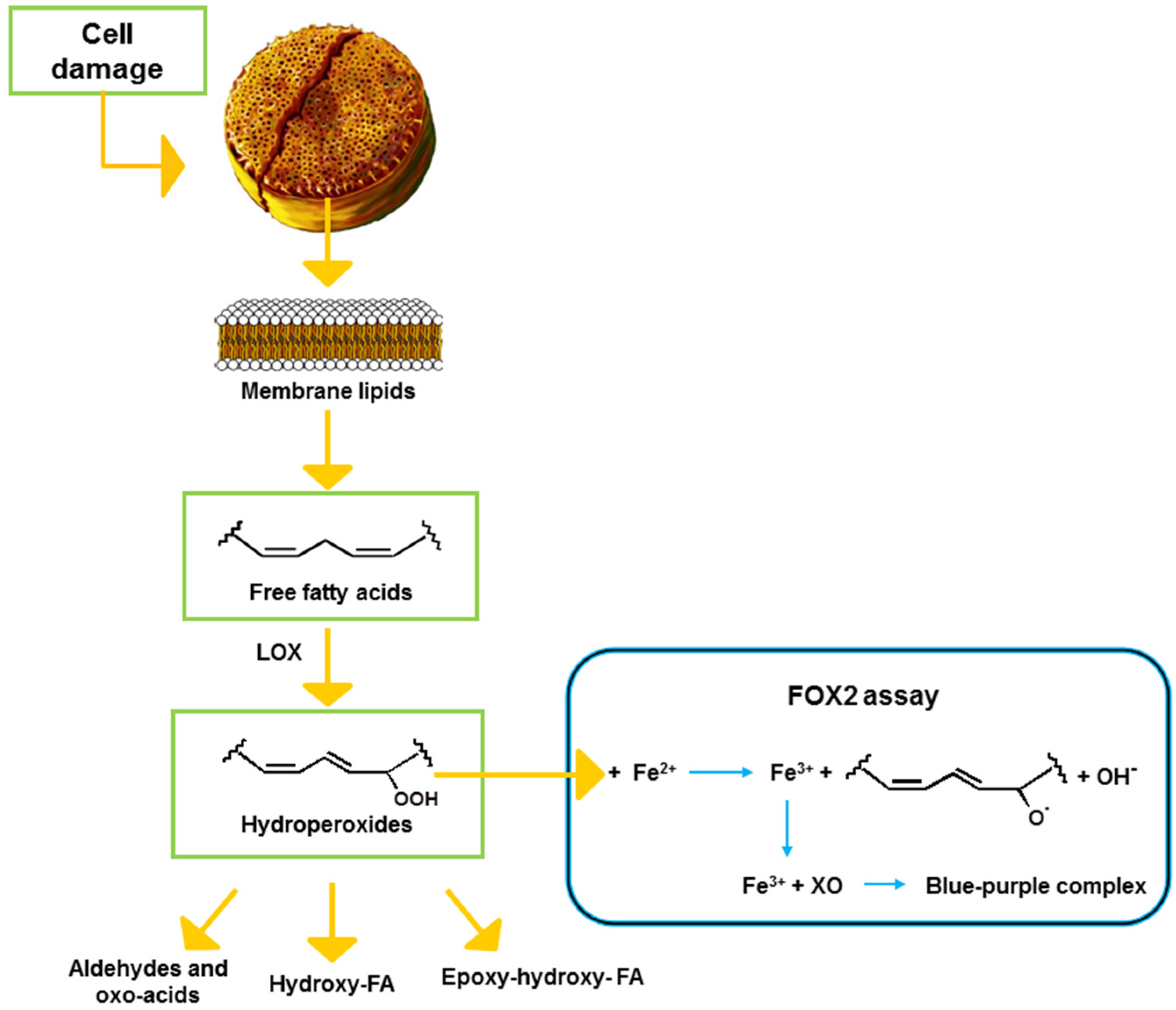
1. Introduction
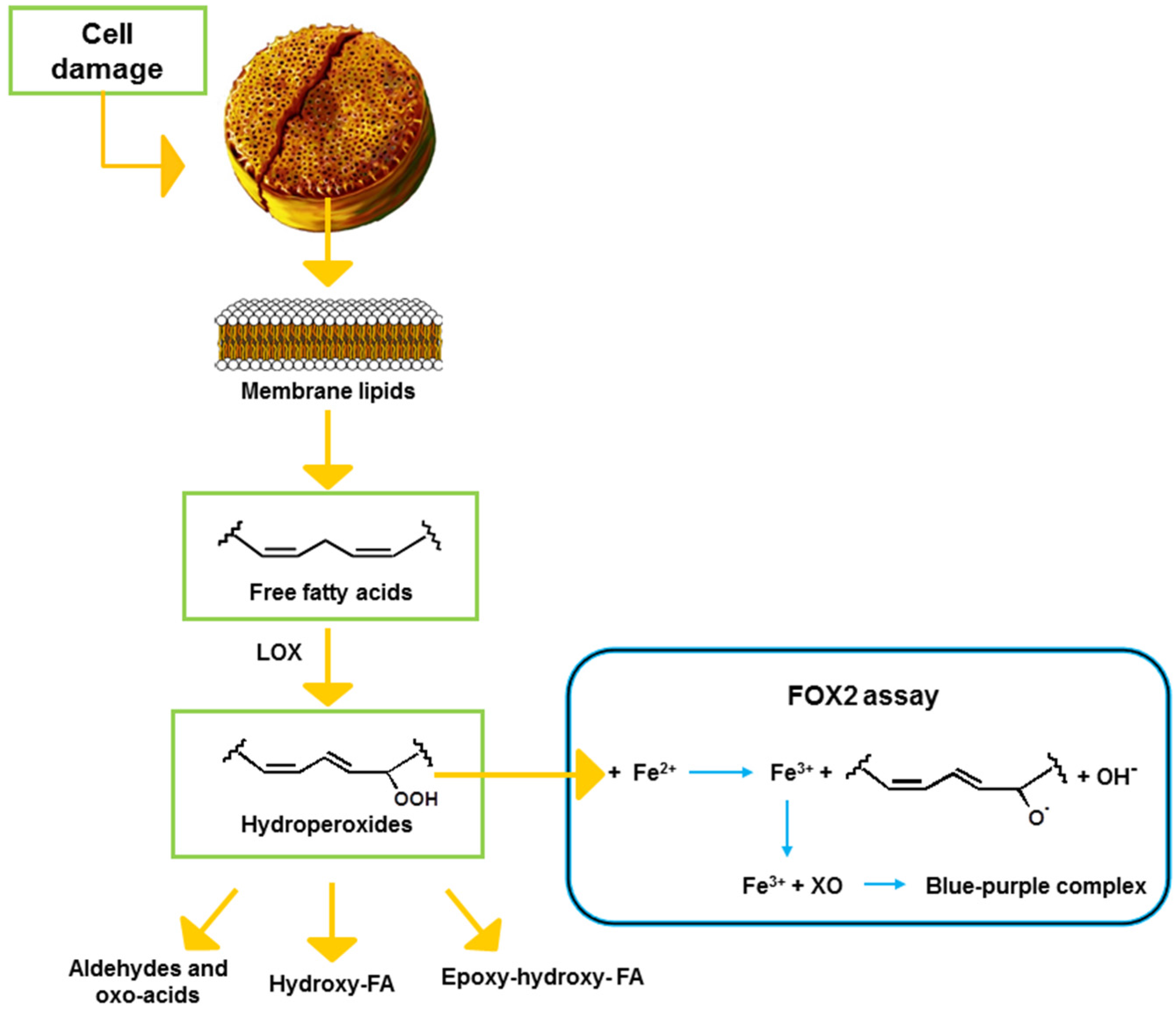
2. Results
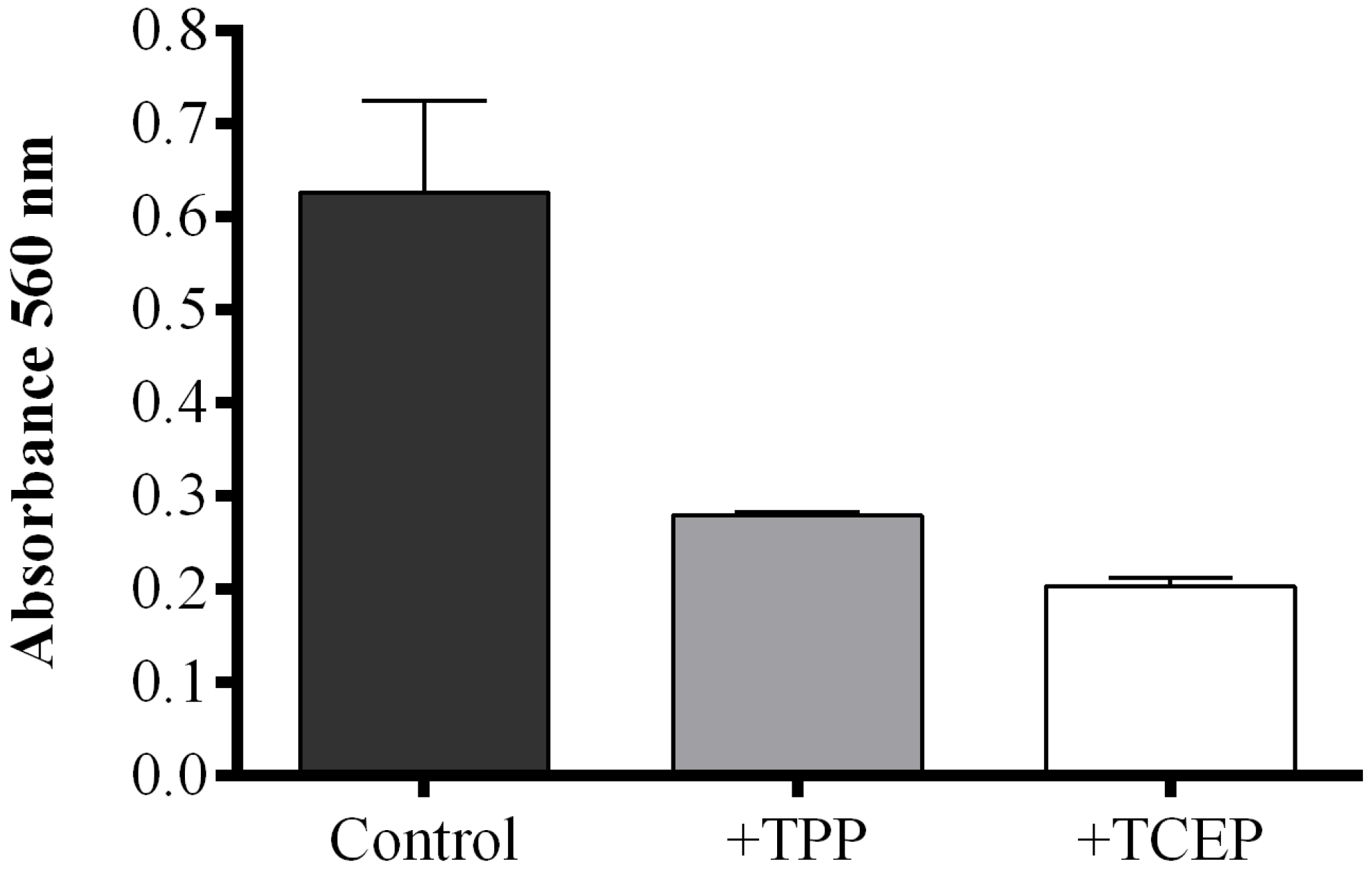
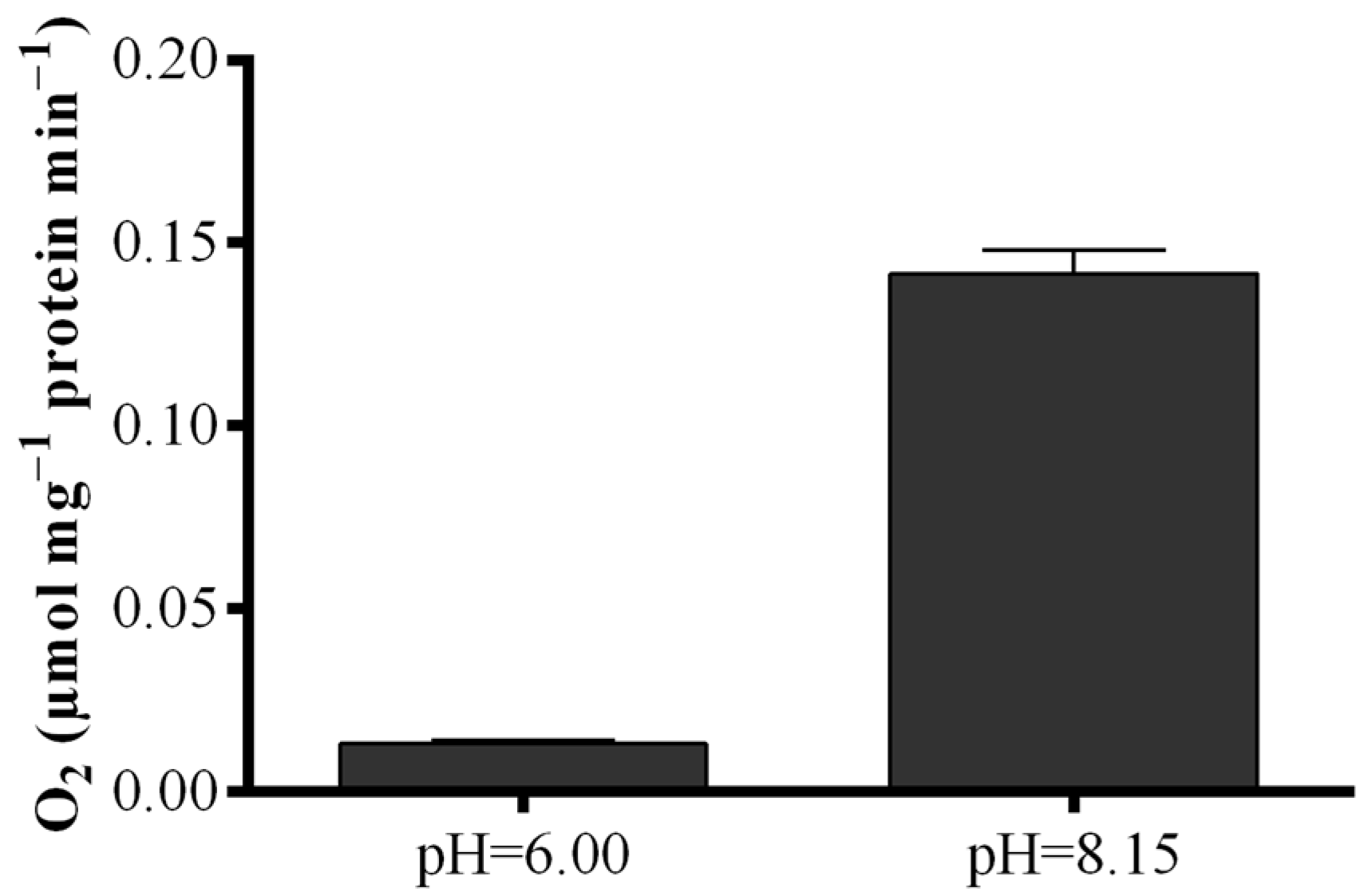
2.1. Optimization of the FOX2 Assay for Diatom Samples
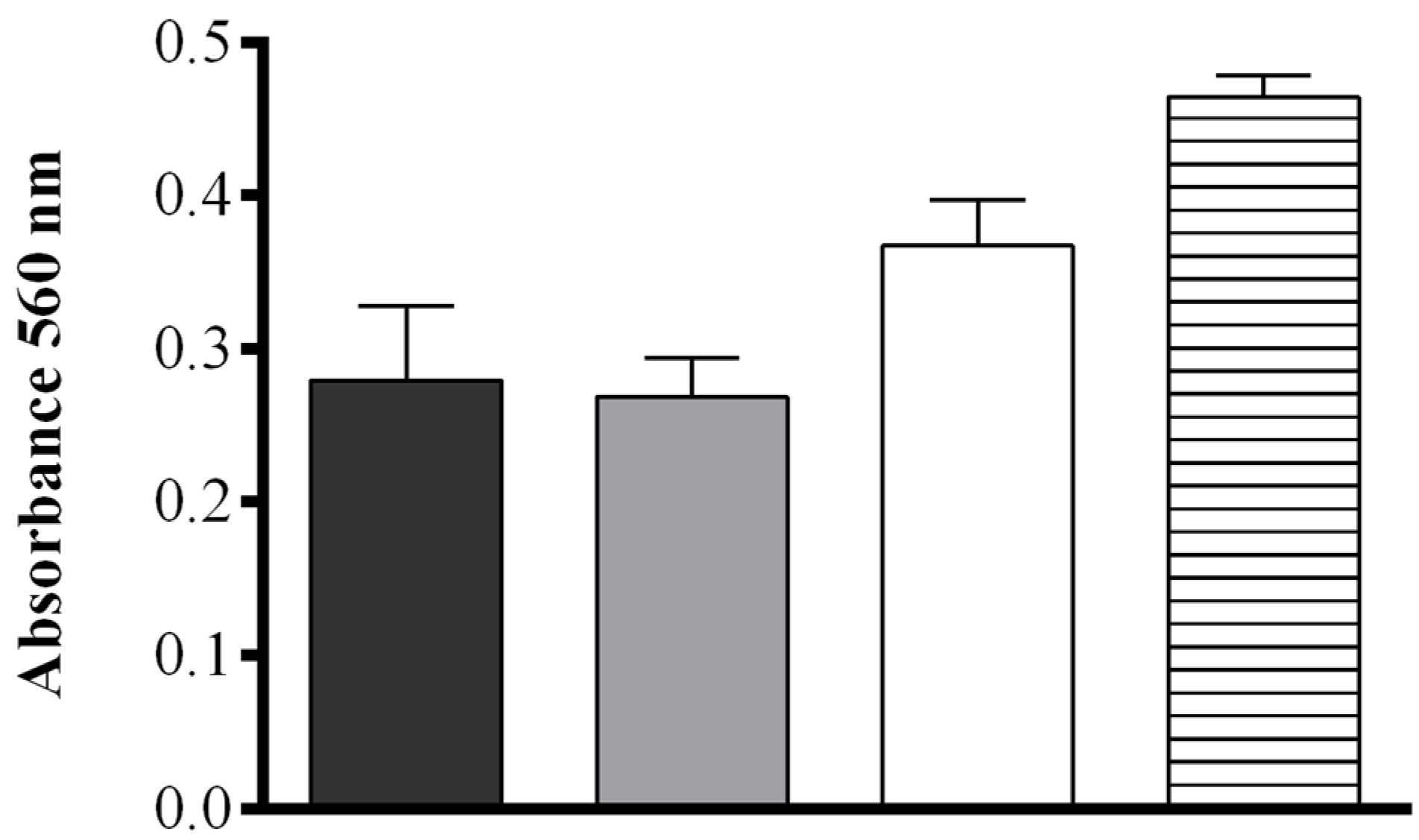
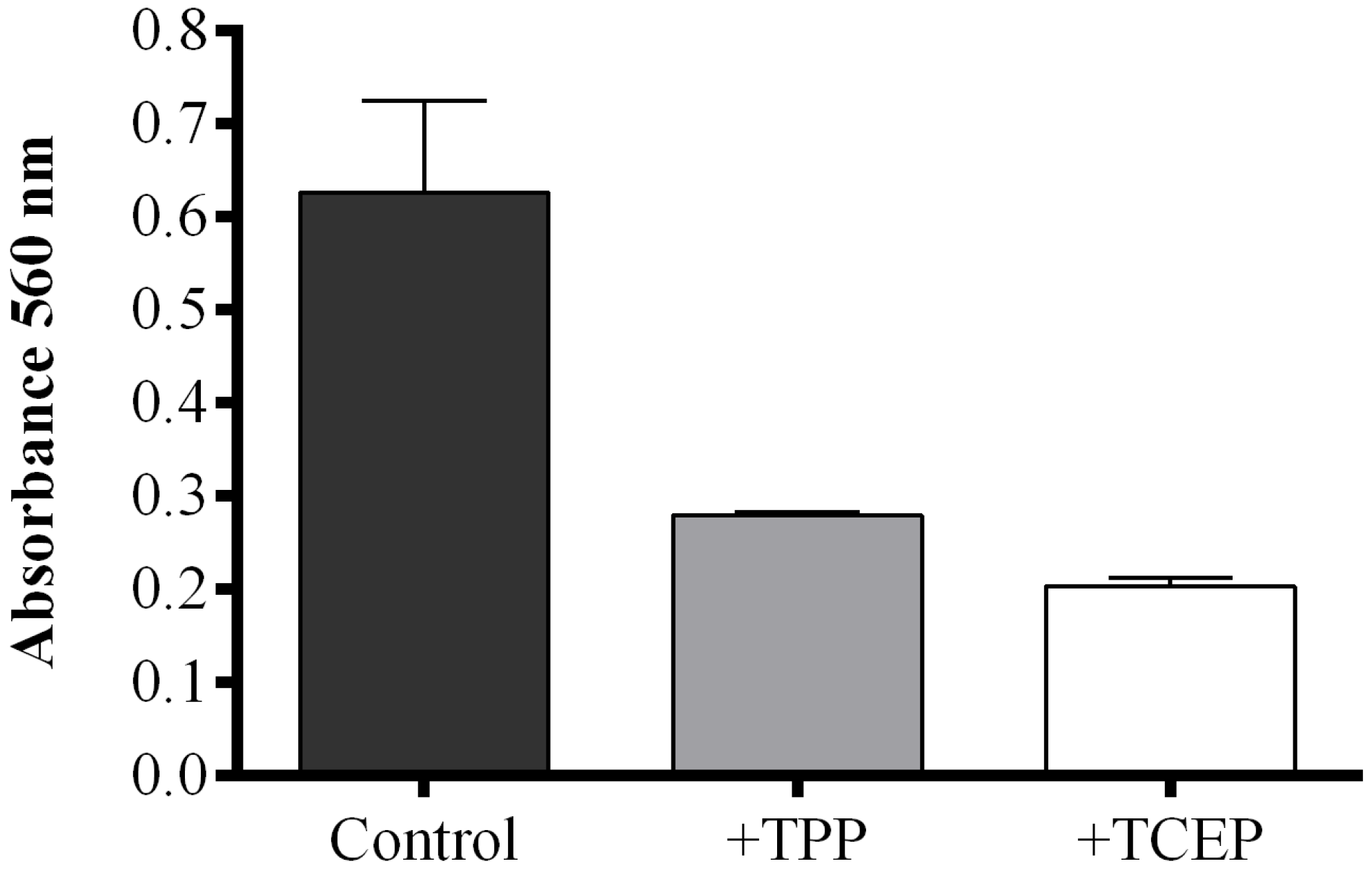
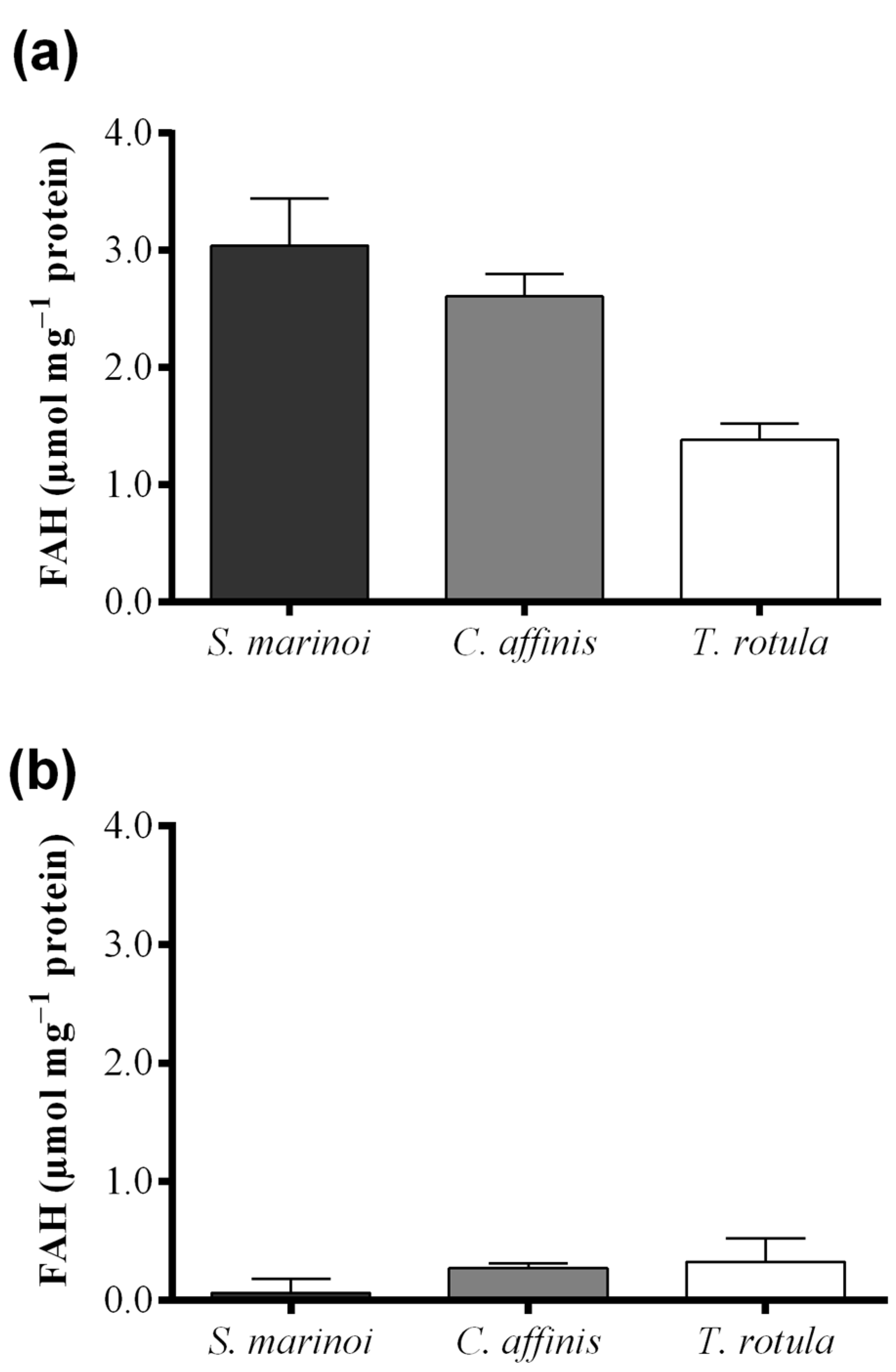
2.2. Fatty Acid Hydroperoxide Production in Selected Diatom Species
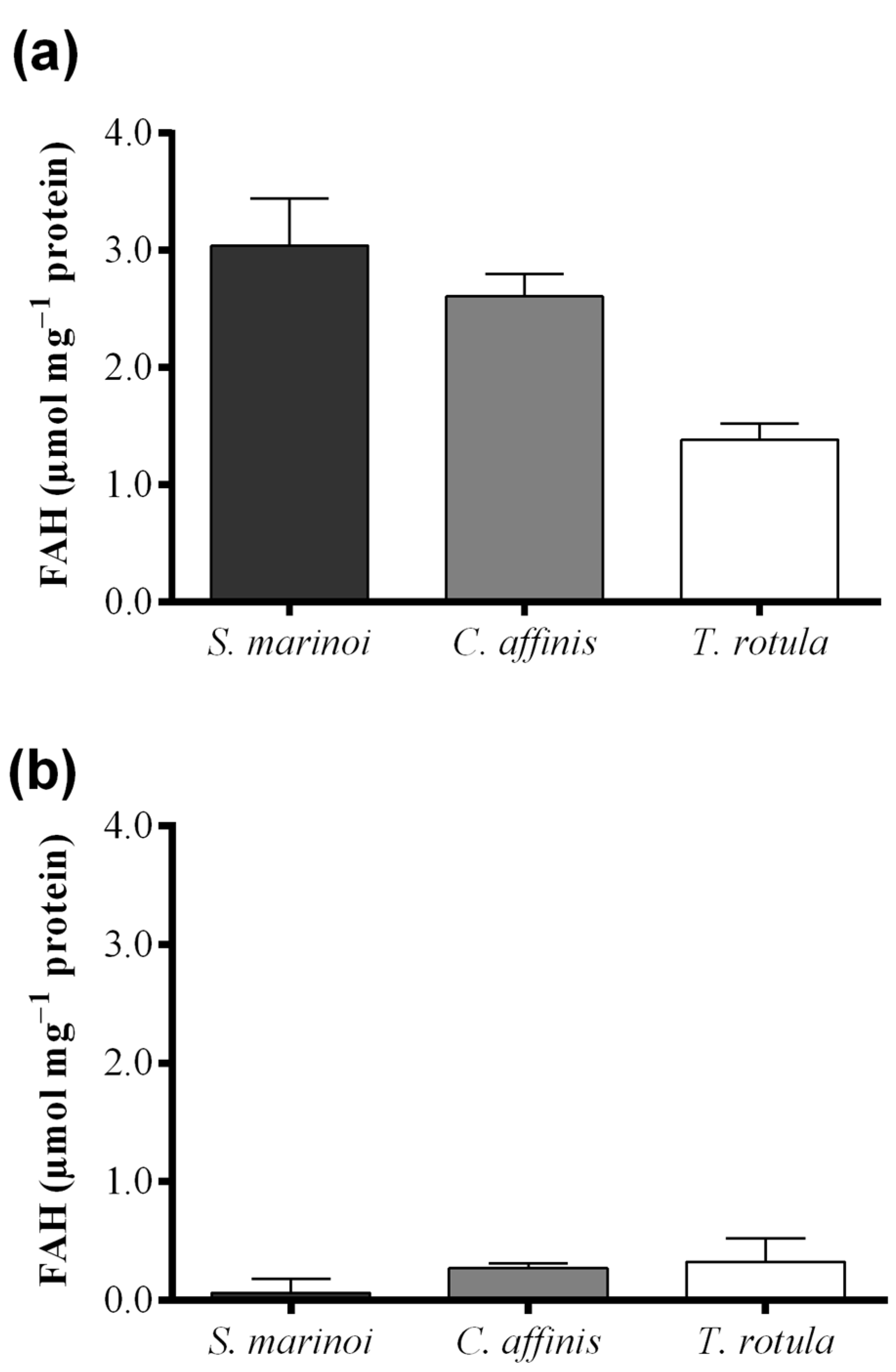
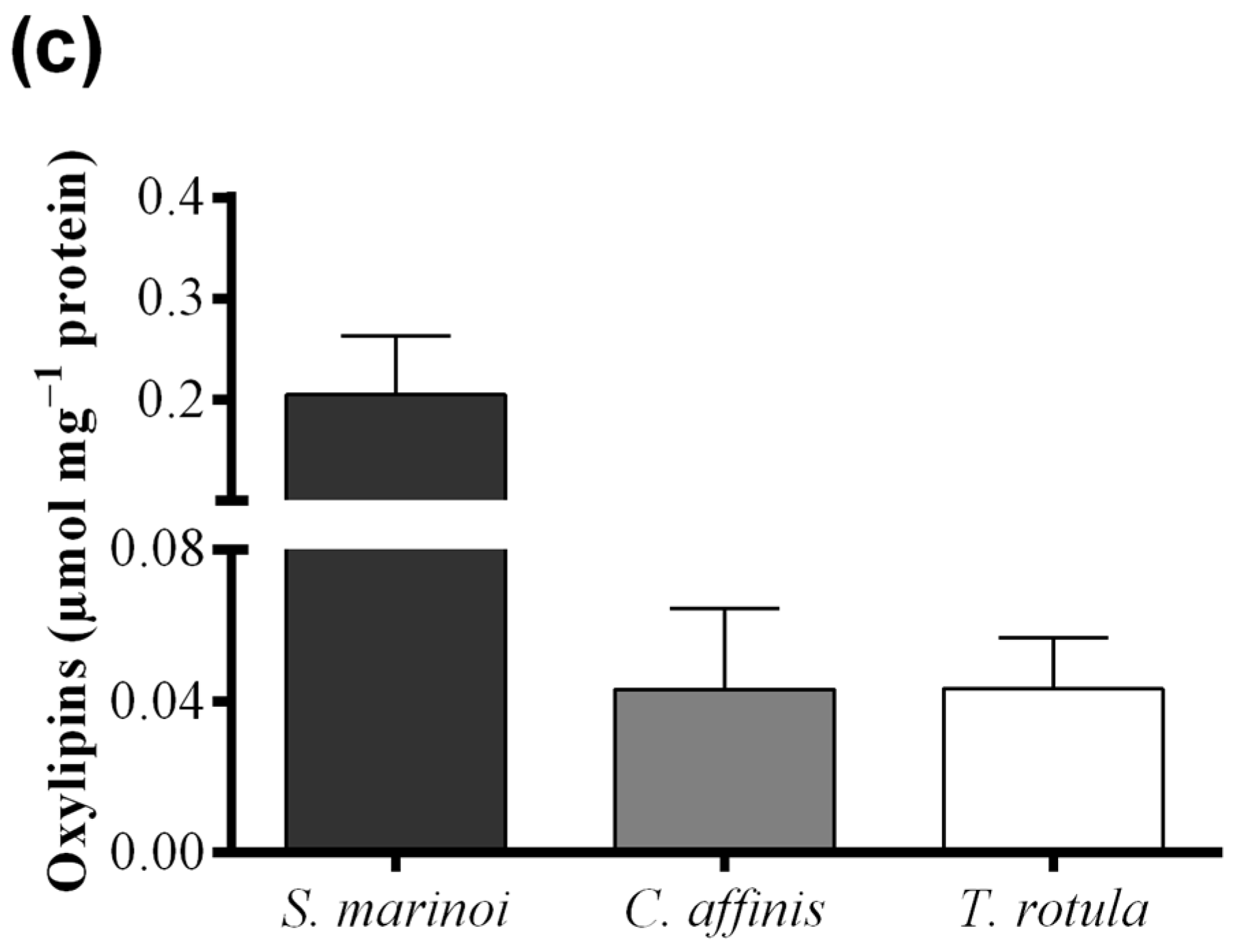

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Pairs | Mean Diff. | q | p Value |
|---|---|---|---|
| FOX2 assay | |||
| S. marinoi–C. affinis | 0.43 | 1.61 | p > 0.05 |
| S. marinoi–T. rotula | 1.65 | 6.16 | p < 0.05 |
| C. affinis–T. rotula | 1.22 | 4.55 | p < 0.05 |
| “Anthon and Barrett” assay | |||
| S. marinoi–C. affinis | −0.21 | 3.44 | p > 0.05 |
| S. marinoi–T. rotula | −0.26 | 4.27 | p < 0.05 |
| C. affinis–T. rotula | −0.05 | 0.75 | p > 0.05 |
| LC and GC–MS | |||
| S. marinoi–C. affinis | 0.16 | 7.10 | p < 0.001 |
| S. marinoi–T. rotula | 0.16 | 7.09 | p < 0.001 |
| C. affinis–T. rotula | −0.00023 | 0.0087 | p > 0.05 |
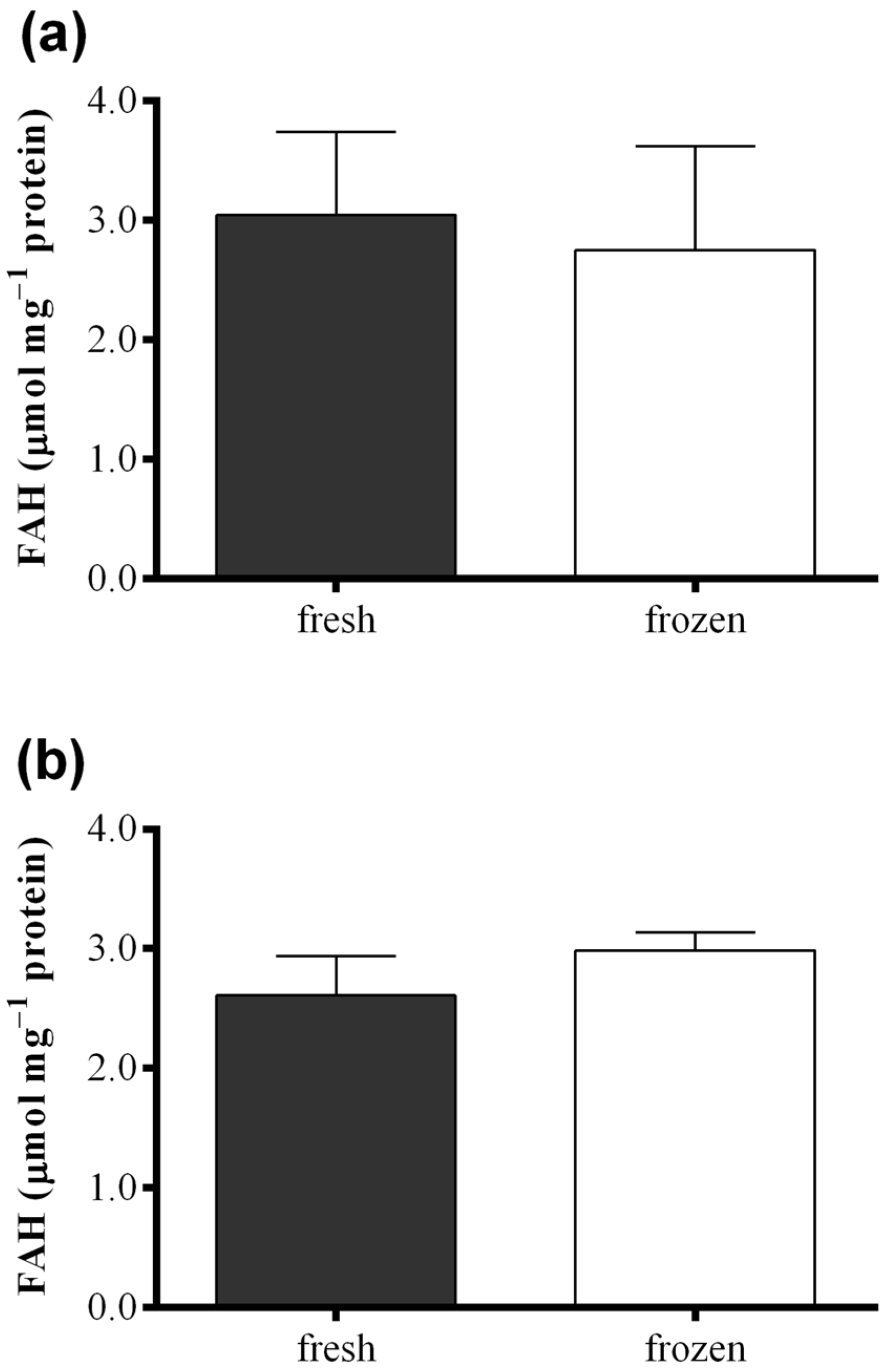
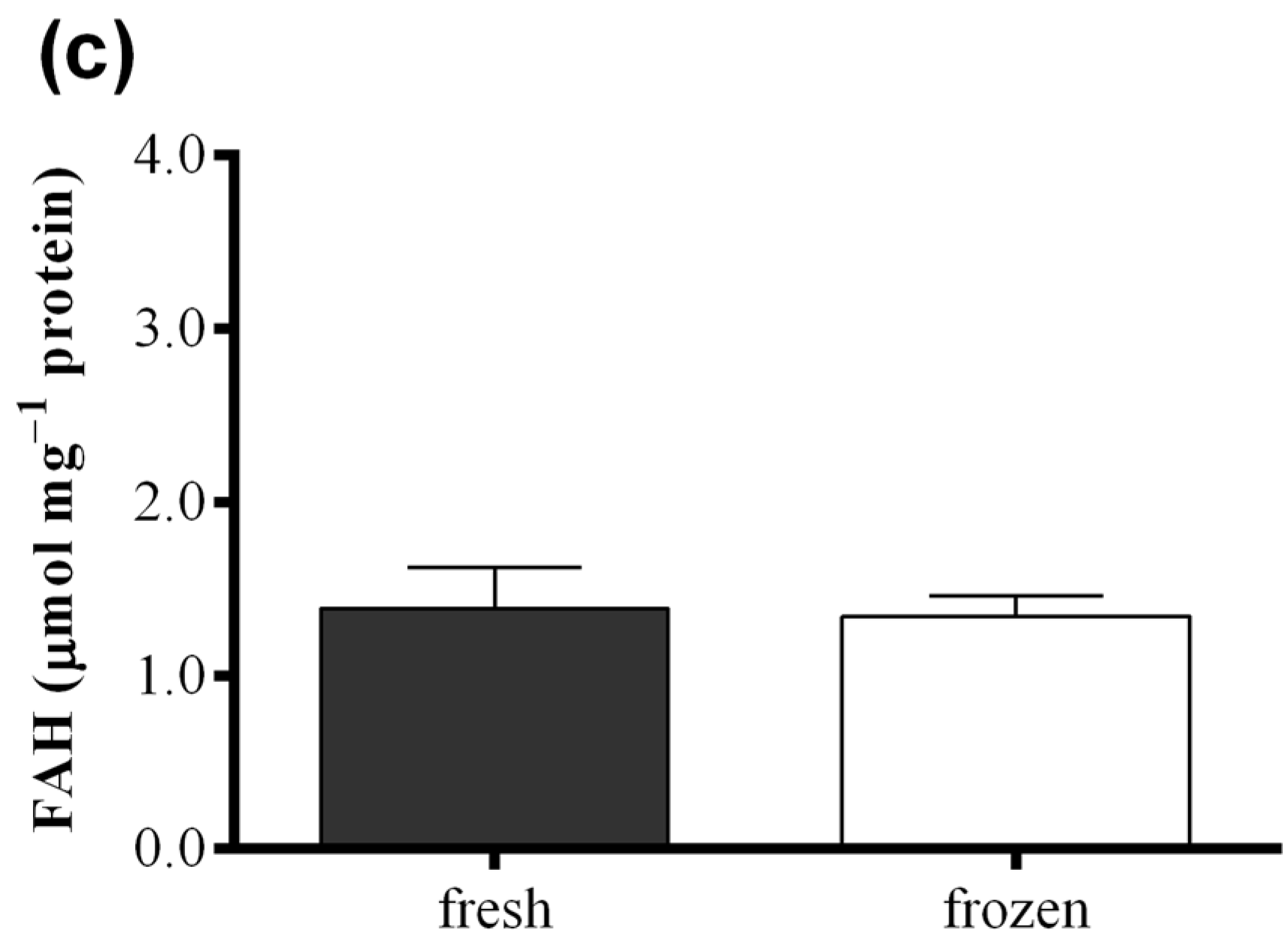
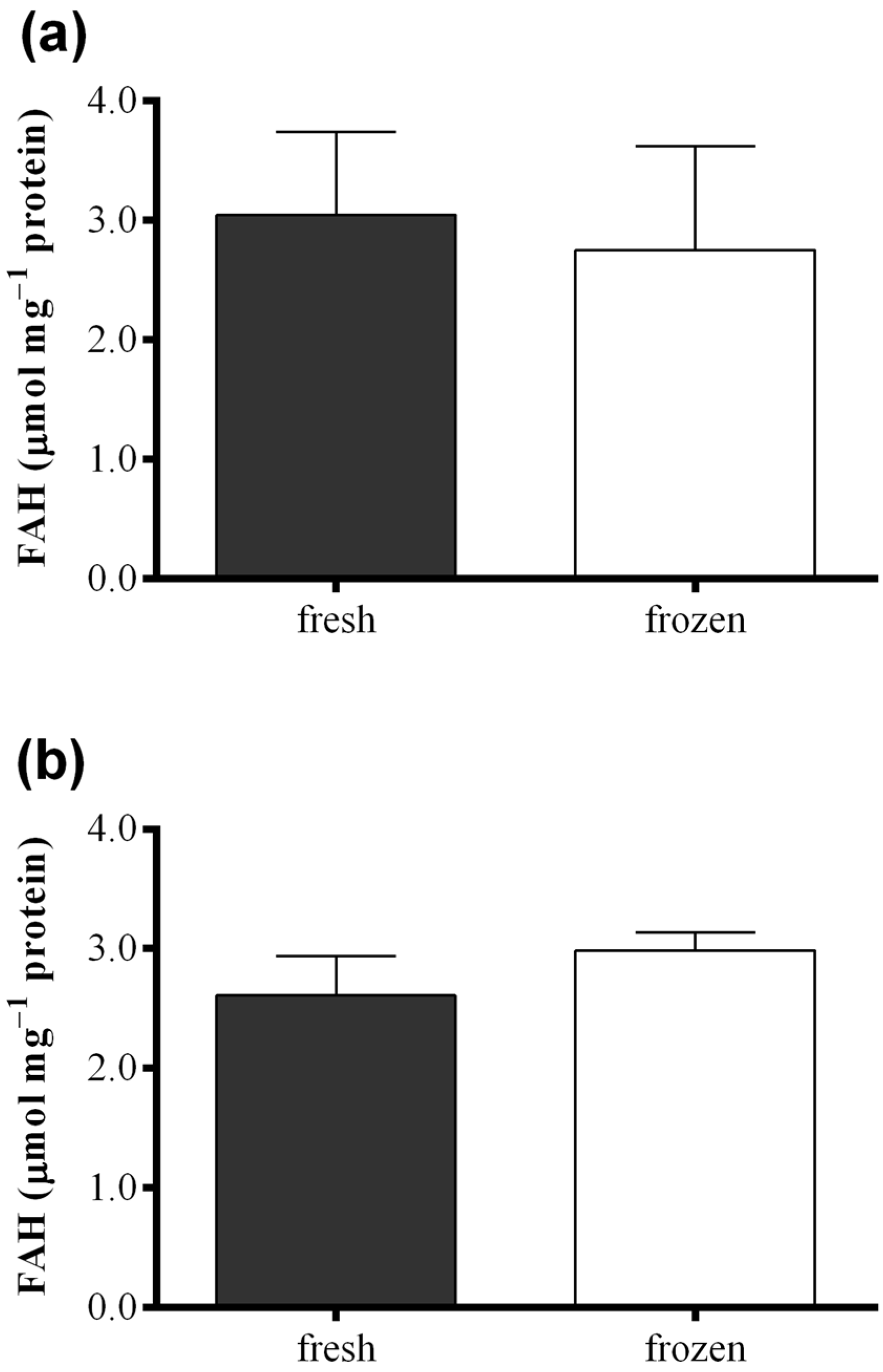
2.3. Effect of Freezing on Diatom Samples
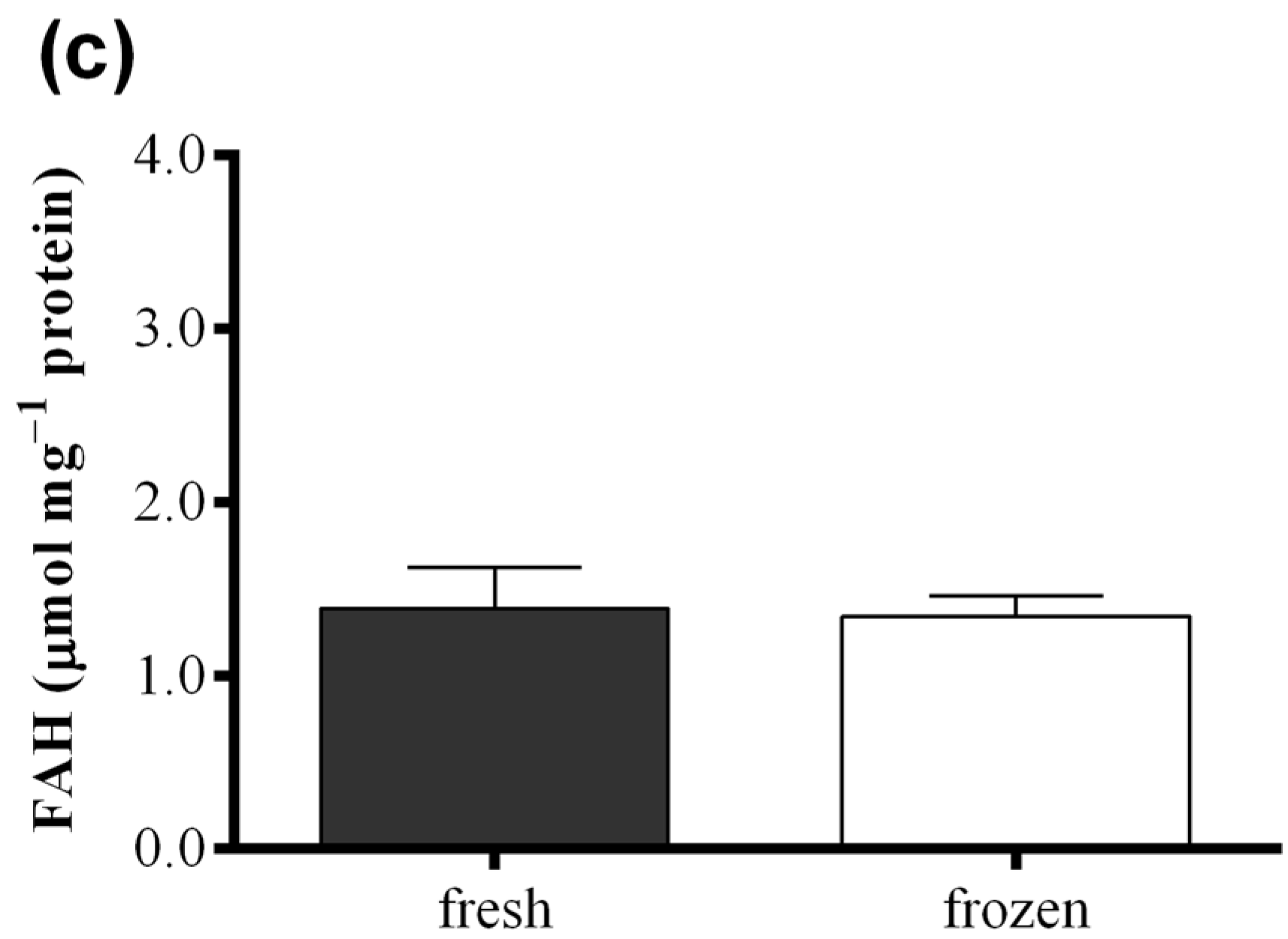
3. Discussion
4. Experimental Section
4.1. Cell Culturing and Harvesting
4.2. Colorimetric Assay According to the FOX2 Method
4.3. Colorimetric Assay According to Anthon and Barrett
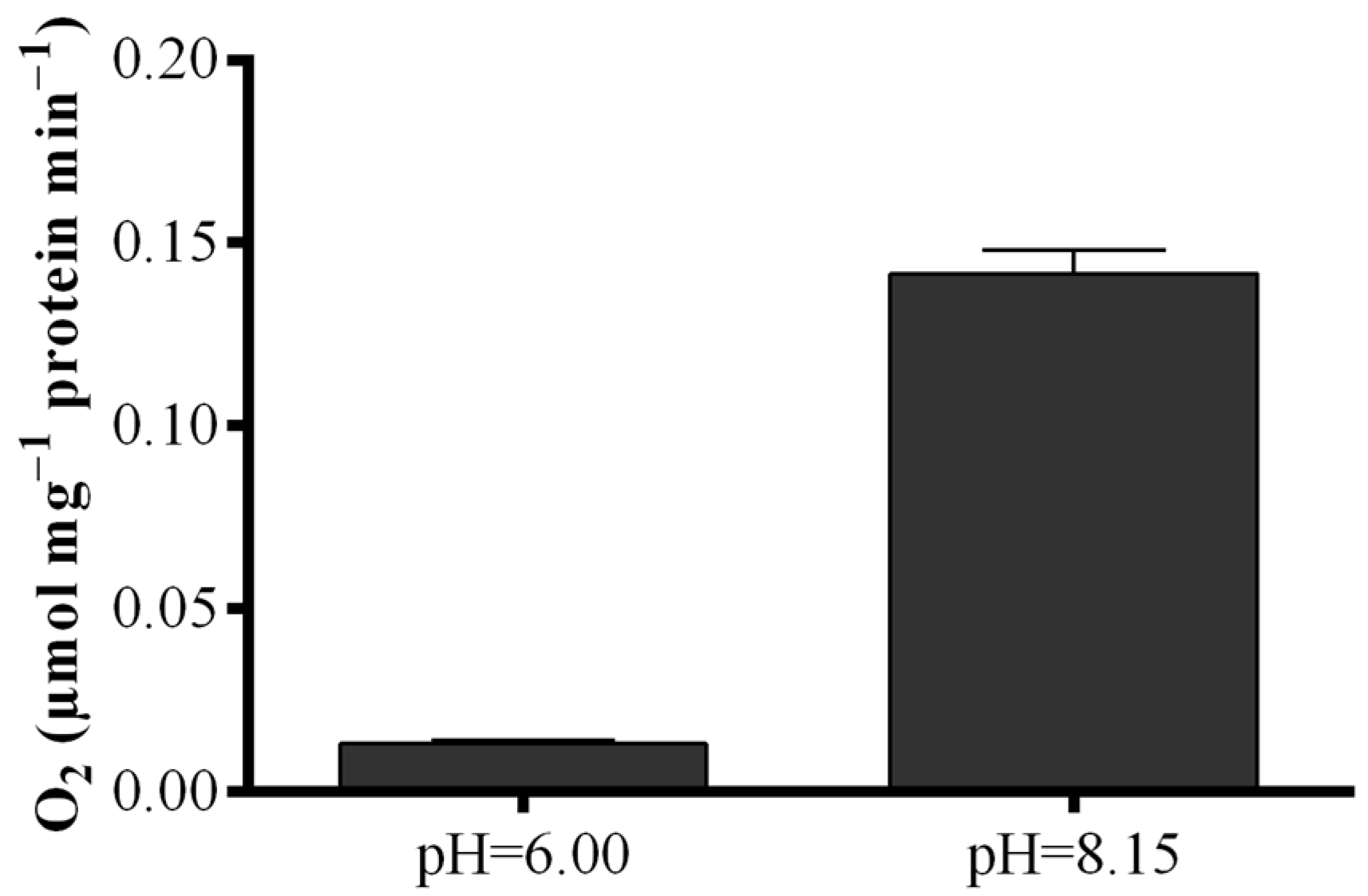
4.4. Polarographic Assay
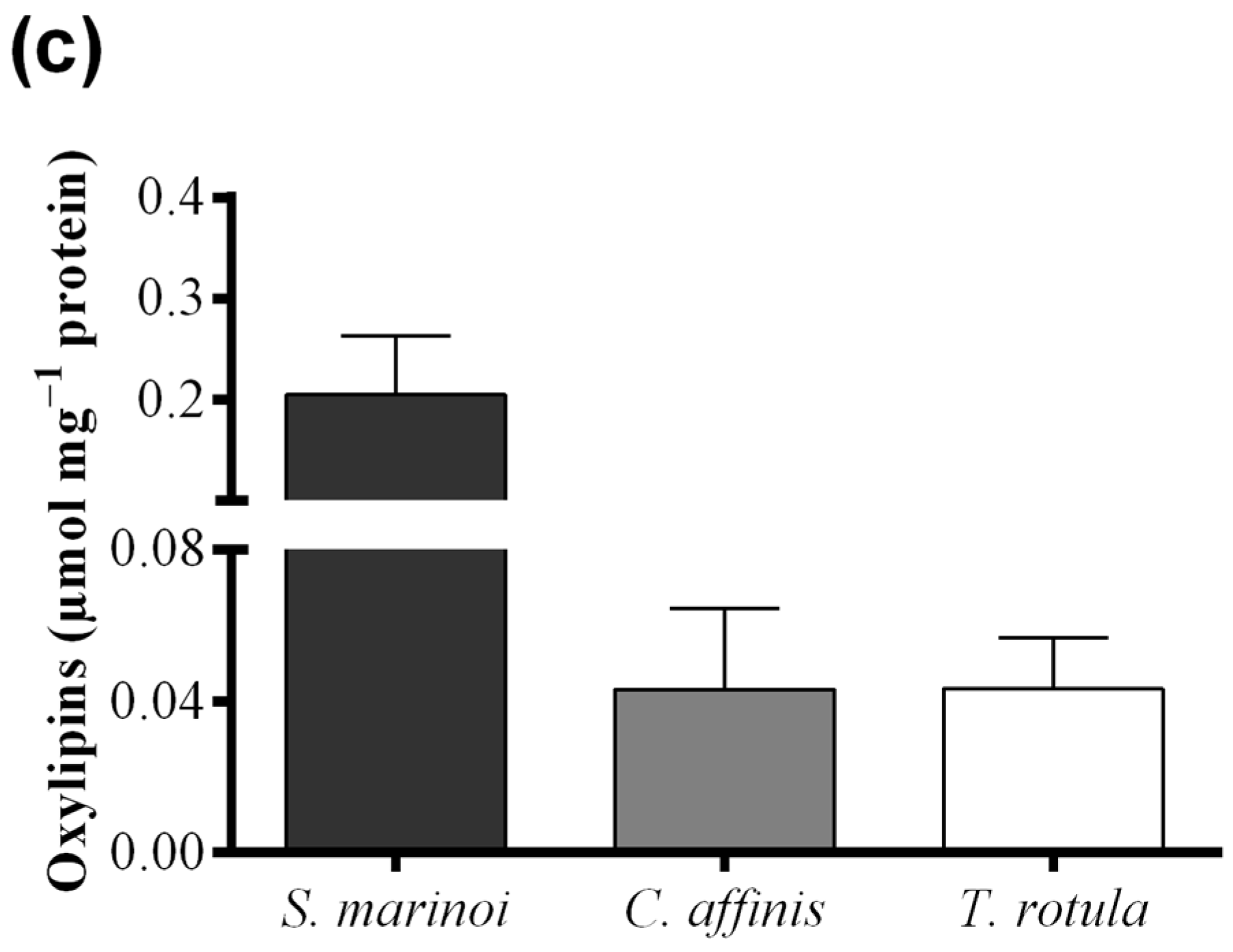
4.5. Oxylipin Quantification
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Riley, G.A. A theoretical analysis of the zooplankton population on Georges Bank. J. Mar. Res. 1947, 6, 104–113. [Google Scholar]
- Caldwell, G.S. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar. Drugs 2009, 7, 367–400. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, S.; Ianora, A. Different diatoms induce contrasting effects on the reproductive success of the copepod Temora stylifera. J. Exp. Mar. Biol. Ecol. 2003, 294, 189–202. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A.; Carotenuto, I.; Buttino, I.; Romano, G.; Casotti, R.; Pohnert, G.; Wichard, T.; Colucci-D’Amato, L.; Terrazzano, G.; et al. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Miralto, A.; Guglielmo, L.; Zagami, G.; Buttino, I.; Granata, A.; Ianora, A. Inhibition of population growth in the copepods Acartia clausi and Calanus helgolandicus during diatom blooms. Mar. Ecol. Prog. Ser. 2003, 254, 253–268. [Google Scholar] [CrossRef]
- Ianora, A.; Casotti, R.; Bastianini, M.; Brunet, C.; d’Ippolito, G.; Acri, F.; Fontana, A.; Cutignano, A.; Turner, J.; Miralto, A. Low reproductive success for copepods during a bloom of the non-aldehyde-producing diatom Cerataulina pelagica in the North Adriatic Sea. Mar. Ecol. 2008, 29, 399–410. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Cutignano, A.; Briante, R.; Febbraio, F.; Cimino, G.; Fontana, A. New C16 fatty-acid-based oxylipin pathway in the marine diatom Thalassiosira rotula. Org. Biomol. Chem. 2005, 3, 4065–4070. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Romano, G.; Lamari, N.; Massa-Gallucci, A.; Cimino, G.; Miralto, A.; Ianora, A. LOX-induced lipid peroxidation mechanism responsible for the detrimental effect of marine diatoms on zooplankton grazers. ChemBioChem 2007, 8, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Pohnert, G. Phospholipase A2 activity triggers the wound-activated chemical defence in the diatom Thalassiosira rotula. Plant Physiol. 2002, 129, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Miralto, A.; Ianora, A.; Romano, G.; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl. Chem. 2007, 79, 481–490. [Google Scholar] [CrossRef]
- Gerecht, A.; Romano, G.; Ianora, A.; d’Ippolito, G.; Cutignano, A.; Fontana, A. Plasticity of oxylipin metabolism among clones of the marine diatoms Skeletonema marinoi (bacillariophyceae). J. Phycol. 2011, 47, 1050–1056. [Google Scholar] [CrossRef]
- Lamari, N.; Ruggiero, M.V.; d’Ippolito, G.; Kooistra, W.H.; Fontana, A.; Montresor, M. Specificity of lipoxygenase pathways supports species delineation in the marine diatom genus Pseudo-nitzschia. PLoS ONE 2013, 8, e73281. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar] [PubMed]
- Buttino, I.; de Rosa, G.; Carotenuto, Y.; Mazzella, M.; Ianora, A.; Esposito, F.; Vitiello, V.; Quaglia, F.; la Rotonda, M.I.; Miralto, A. Aldehyde-encapsulating liposomes impair marine grazer survivorship. J. Exp. Biol. 2008, 211, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, S.; Nejstgaard, J.C.; Pohnert, G. Accumulation of polyunsaturated aldehydes in the gonads of the copepod Acartia tonsa revealed by tailored fluorescent probes. PLoS ONE 2014, 9, e112522. [Google Scholar] [CrossRef] [PubMed]
- Lilius, E.M.; Laaskso, S. A sensitive lipoxygnase assay based on chemiluminescence. Anal. Biochem. 1982, 119, 135–141. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S. Lipoxygenase from soybeans. Methods Enzymol. 1981, 71, 441–451. [Google Scholar]
- Cutignano, A.; Lamari, N.; d’ippolito, G.; Manzo, E.; Cimino, G.; Fontana, A. Lipoxygenase products in marine diatoms: A concise analytical method to explore the functional potential of oxylipins. J. Phycol. 2011, 47, 233–243. [Google Scholar] [CrossRef]
- Anthon, G.E.; Barrett, D.M. Colorimetric method for the determination of lipoxygenase activity. J. Agric. Food Chem. 2001, 49, 32–37. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Cutignano, A.; Tucci, S.; Romano, G.; Cimino, G.; Fontana, A. Biosynthetic intermediates and stereochemical aspects of aldehyde biosynthesis in the marine diatom Thalassiosira rotula. Phytochemistry 2006, 67, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.P. Ferrous ion oxidation in presence of ferric ion indicator xylenol orange for measurement of hydroperoxides. Methods Enzymol. 1994, 233, 182–189. [Google Scholar]
- DeLong, J.M.; Prange, R.K.; Hodges, D.M.; Forney, C.F.; Bishop, M.C.; Quilliam, M. Using a modified ferrous oxidation-xylenol orange (FOX) assay for detection of lipid hydroperoxides in plant tissue. J. Agric. Food Chem. 2002, 50, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bou, R.; Codony, R.; Tres, A.; Decker, E.A.; Guardiola, F. Determination of hydroperoxides in foods and biological samples by the ferrous oxidation–xylenol orange method: A review of the factors that influence the method’s performance. Anal. Biochem. 2008, 377, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sodergren, E.; Nourooz-Zadeh, J.; Berglund, L.; Vessby, B. Re-evaluation of the ferrous oxidation in xylenol orange assay for the measurement of plasma lipid hydroperoxides. J. Biochem. Biophys. Methods 1998, 37, 137–146. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Shantha, N.C.; Decker, E.A. Rapid, sensitive, iron-based spectrophotometric methods for determination of peroxide values of food lipids. J. AOAC Int. 1994, 77, 421–424. [Google Scholar] [PubMed]
- Nourooz-Zadeh, J.; Tajaddini-Sarmadi, J.; Birlouez-Aragon, I.; Wolff, S.P. Measurement of hydroperoxides in edible oils using the ferrous oxidationin xylenol orange assay. J. Agric. Food Chem. 1995, 43, 17–21. [Google Scholar] [CrossRef]
- Ugalde-Benítez, V.; Rojo-Domínguez, A.; Jaramillo-Flores, M.E.; Guerrero-Legarreta, I. Effect of sample solubilized aliquot volume and concentration on hydroperoxide detection in PUFA-rich fish oil. Int. J. Food Prop. 2014, 17, 650–660. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Willmore, W.G.; Storey, K.B. Quantification of lipid peroxidation in tissue extracts based on Fe(III)xylenol orange complex formation. Free Radic. Biol. Med. 1995, 19, 271–280. [Google Scholar] [CrossRef]
- Grau, A.; Codony, R.; Rafecas, M.; Barroeta, A.C.; Guardiola, F. Lipid hydroperoxide determination in dark chicken meat through a ferrous oxidation-xylenol orange method. J. Agric. Food Chem. 2000, 48, 4136–4143. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Singh, R.P.; Bijo, A.J.; Reddy, C.R.K.; Jha, B. Estimation of lipid hydroperoxide levels in tropical marine macroalgae. J. Phycol. 2012, 48, 1362–1373. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Santos, S.A.O.; Simões, M.M.Q.; Neto, C.P.; Domingues, M.R.M.; Silvestre, A.J.D. Analysis of linoleic acid hydroperoxides generated by biomimetic and enzymatic systems through an integrated methodology. Ind. Crop. Prod. 2011, 34, 1474–1481. [Google Scholar] [CrossRef]
- Koski, M.; Wichard, T.; Jonasdottir, S.H. “Good” and “bad” diatoms: Development, growth and juvenile mortality of the copepod Temora longicornis on diatom diets. Mar. Biol. 2008, 154, 719–734. [Google Scholar] [CrossRef]
- Berge, J.P.; Gouyou, J.P.; Dubacq, J.P.; Durand, P. Reassessment of lipid composition of the diatom Skeletonema costatum. Phytochemistry 1995, 39, 1017–1021. [Google Scholar] [CrossRef]
- Nourooz-Zadeh, J.; Tajaddini-Sarmadi, J.; Wolff, S.P. Measurement of plasma hydroperoxide concentrations by the ferrous oxidation-xylenol orange assay in conjunction with triphenylphosphine. Anal. Biochem. 1994, 220, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Grintzalis, K.; Zisimopoulos, D.; Grune, T.; Weber, D.; Georgiou, C.D. Method for the simultaneous determination of free/protein malondialdehyde and lipid/protein hydroperoxides. Free Radic. Biol. Med. 2013, 59, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Ribalet, F.; Vidoudez, C.; Cassin, D.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. High plasticity in the production of diatom-derived polyunsaturated aldehydes under nutrient limitation: Physiological and ecological implications. Protist 2009, 160, 444–451. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Lamari, N.; Montresor, M.; Romano, G.; Cutignano, A.; Gerecht, A.; Cimino, G.; Fontana, A. 15S-Lipoxygenase metabolism in the marine diatom Pseudo-nitzschia delicatissima. New Phytol. 2009, 183, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Romano, G.; Iadicicco, O.; Miralto, A.; Ianora, A.; Cimino, G.; Fontana, A. New birth-control aldehydes from the marine diatom Skeletonema costatum: Characterization and biogenesis. Tetrahedron Lett. 2002, 43, 6133–6136. [Google Scholar] [CrossRef]
- Guillard, R.L. Culture of phytoplankton for feeding marine invertebrates. In Cultures of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Publishing Corporation: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Iadicicco, O.; Romano, G.; Fontana, A. Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron Lett. 2002, 43, 6137–6140. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orefice, I.; Gerecht, A.; D'Ippolito, G.; Fontana, A.; Ianora, A.; Romano, G. Determination of Lipid Hydroperoxides in Marine Diatoms by the FOX2 Assay. Mar. Drugs 2015, 13, 5767-5783. https://doi.org/10.3390/md13095767
Orefice I, Gerecht A, D'Ippolito G, Fontana A, Ianora A, Romano G. Determination of Lipid Hydroperoxides in Marine Diatoms by the FOX2 Assay. Marine Drugs. 2015; 13(9):5767-5783. https://doi.org/10.3390/md13095767
Chicago/Turabian StyleOrefice, Ida, Andrea Gerecht, Giuliana D'Ippolito, Angelo Fontana, Adrianna Ianora, and Giovanna Romano. 2015. "Determination of Lipid Hydroperoxides in Marine Diatoms by the FOX2 Assay" Marine Drugs 13, no. 9: 5767-5783. https://doi.org/10.3390/md13095767
APA StyleOrefice, I., Gerecht, A., D'Ippolito, G., Fontana, A., Ianora, A., & Romano, G. (2015). Determination of Lipid Hydroperoxides in Marine Diatoms by the FOX2 Assay. Marine Drugs, 13(9), 5767-5783. https://doi.org/10.3390/md13095767