
Cadmium-Containing Carbonic Anhydrase CDCA1 in Marine Diatom Thalassiosira weissflogii





Abstract
:
{kind=link}
{kind=link}
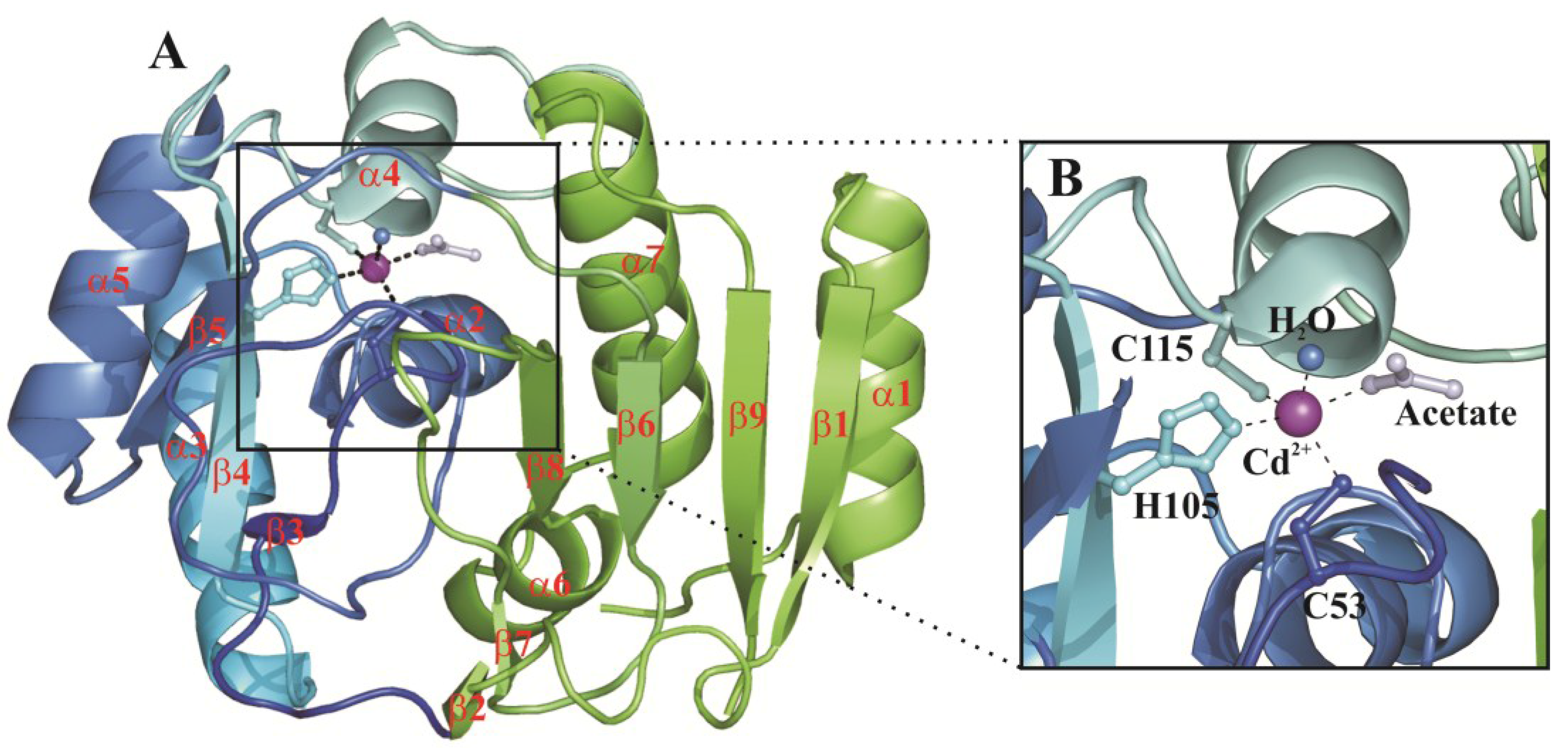
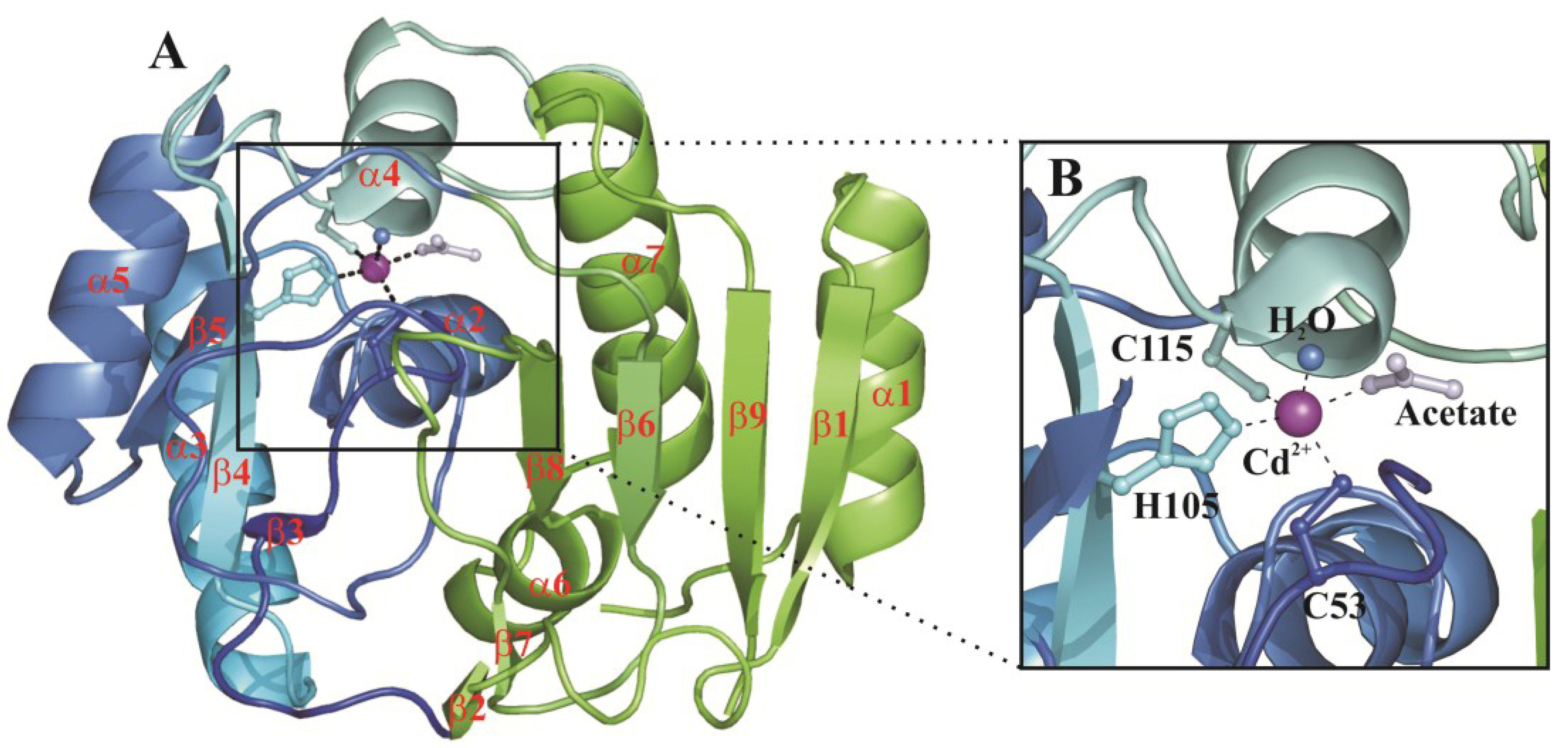{kind=link}
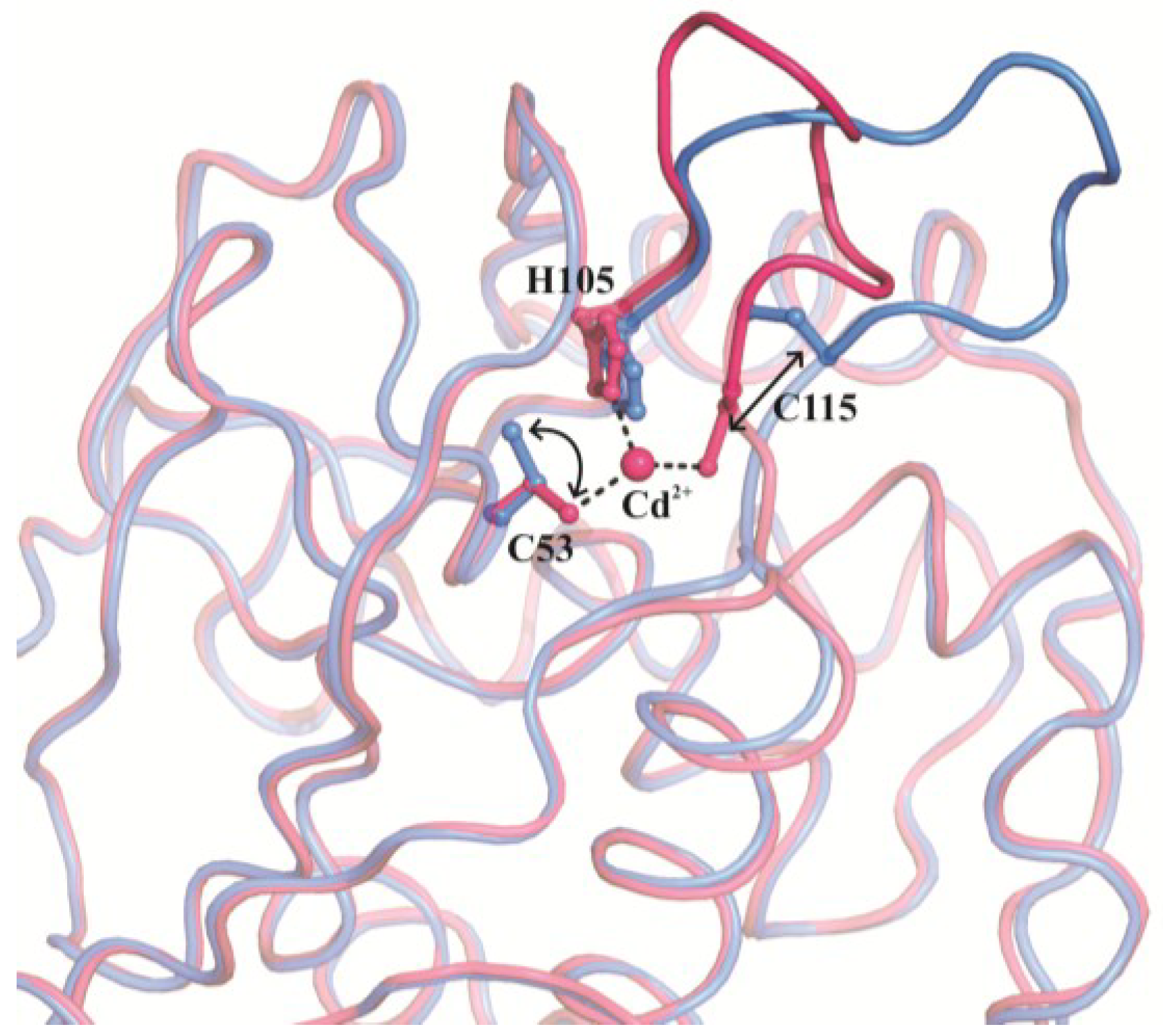{kind=link}
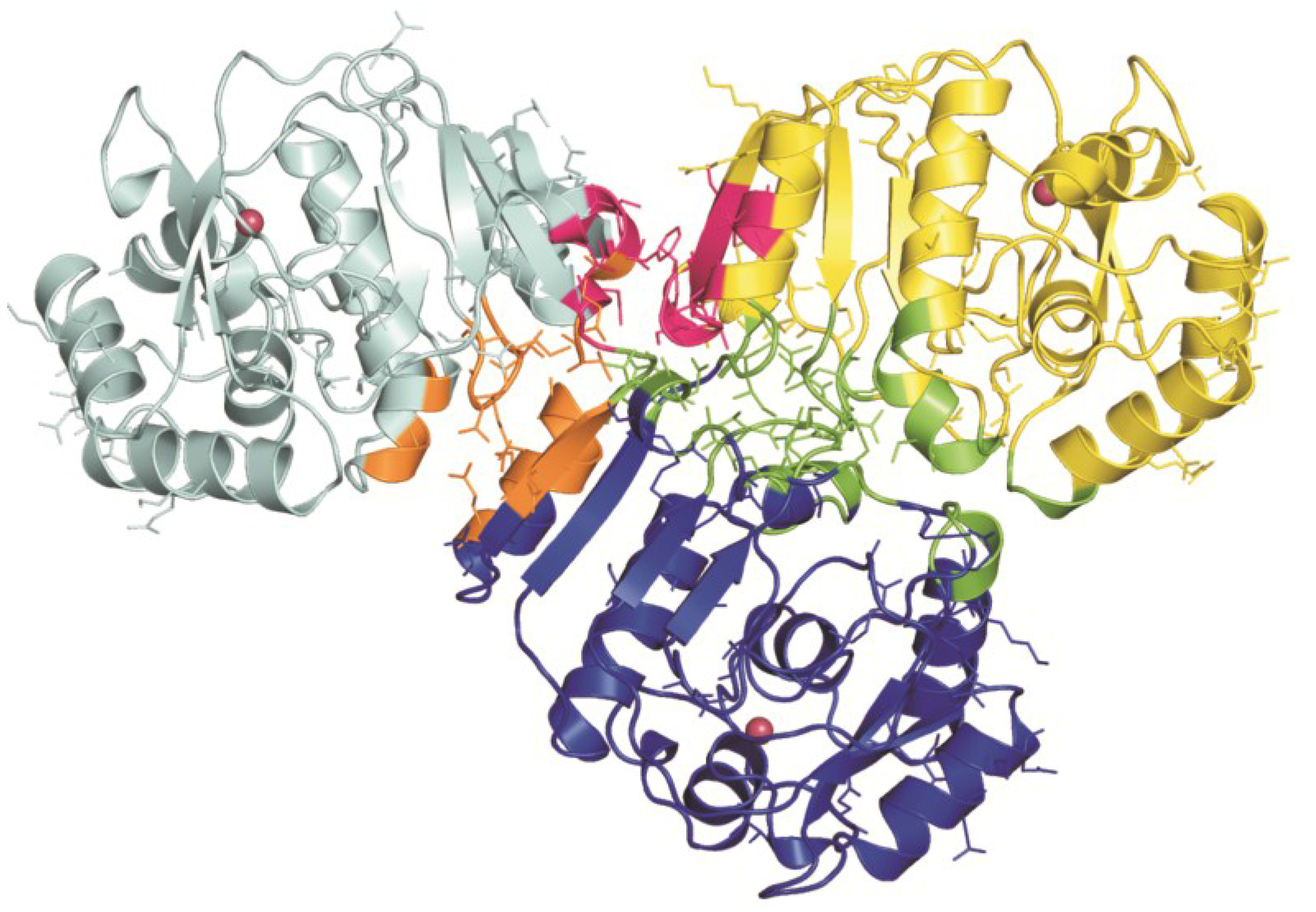{kind=link}
1. Introduction
2. Kinetic and Inhibition Assays

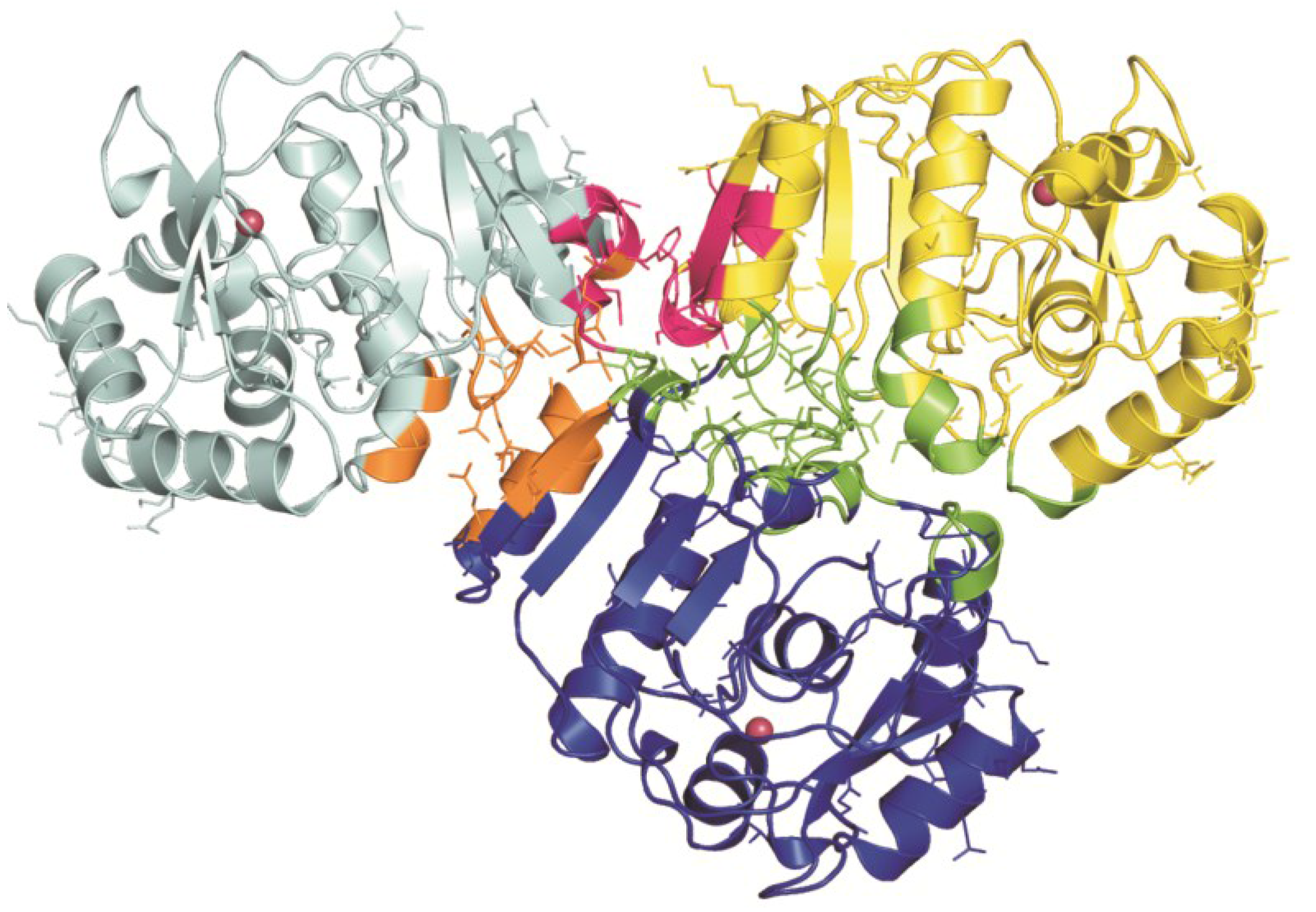
3. Structural Characterization of CDCA1

4. Potential Biotechnological Applications of CDCA1 Enzyme
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Lane, T.W.; Morel, F.M. A biological function for cadmium in marine diatoms. Proc. Natl. Acad. Sci. USA 2000, 97, 4627–4631. [Google Scholar] [CrossRef] [PubMed]
- Boyle, E.A.; Sclater, F.; Edmond, J.M. Marine geochemistry of cadmium. Nat. Chem. Biol. 1976, 263, 3. [Google Scholar]
- Xu, Y.; Feng, L.; Jeffrey, P.D.; Shi, Y.; Morel, F.M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 2008, 452, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, V.M.; Kaufman, G.K.; Urbach, A.R.; Gitlin, I.; Gudiksen, K.L.; Weibel, D.B.; Whitesides, G.M. Carbonic anhydrase as a model for biophysical and physical-organic studies of proteins and protein-ligand binding. Chem. Rev. 2008, 108, 946–1051. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Winum, J.-Y. Drug Design of zinc-Enzyme Inhibitors: Functional, Structural, and Disease Applications; Wiley: Hoboken, NJ, USA, 2009; Volume X, Part II. [Google Scholar]
- Maren, T.H. Carbonic anhydrase: Chemistry, physiology, and inhibition. Physiol. Rev. 1967, 47, 595–781. [Google Scholar] [PubMed]
- Beauchemin, M.; Morse, D. Delta carbonic anhydrases: Structure, distribution and potential roles. In Carbonic Anhydrases as Biocatalysts from Theory to Medical and Industrial Applications; Supuran, C.T., De Simone, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 337–349. [Google Scholar]
- Xu, Y.; Morel, F.M. Cadmium in marine phytoplankton. Met. Ions Life Sci. 2013, 11, 509–528. [Google Scholar] [PubMed]
- Chmielowska-Bak, J.; Izbianska, K.; Deckert, J. The toxic doppelganger: On the ionic and molecular mimicry of cadmium. Acta Biochim. Pol. 2013, 60, 369–374. [Google Scholar] [PubMed]
- Belyaeva, E.A.; Dymkowska, D.; Wieckowski, M.R.; Wojtczak, L. Mitochondria as an important target in heavy metal toxicity in rat hepatoma as-30d cells. Toxicol. Appl. Pharmacol. 2008, 231, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, J.F.; Antes, F.G.; Maldaner, J.; Pereira, L.B.; Tabaldi, L.A.; Rauber, R.; Rossato, L.V.; Bisognin, D.A.; Dressler, V.L.; Flores, E.M.; et al. Cadmium and mineral nutrient accumulation in potato plantlets grown under cadmium stress in two different experimental culture conditions. Plant Physiol. Biochem. 2009, 47, 814–821. [Google Scholar]
- Kippler, M.; Hoque, A.M.; Raqib, R.; Ohrvik, H.; Ekstrom, E.C.; Vahter, M. Accumulation of cadmium in human placenta interacts with the transport of micronutrients to the fetus. Toxicol. Lett. 2010, 192, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Matovic, V.; Buha, A.; Bulat, Z.; Dukic-Cosic, D. Cadmium toxicity revisited: Focus on oxidative stress induction and interactions with zinc and magnesium. Arh. Hig. Rada Toksikol. 2011, 62, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Pytharopoulou, S.; Grintzalis, K.; Sazakli, E.; Leotsinidis, M.; Georgiou, C.D.; Kalpaxis, D.L. Translational responses and oxidative stress of mussels experimentally exposed to hg, cu and cd: One pattern does not fit at all. Aquat. Toxicol. 2011, 105, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Li, J.; Chen, D.; Liu, Z. Simultaneous effects of lead and cadmium on primary cultures of rat proximal tubular cells: Interaction of apoptosis and oxidative stress. Arch. Environ. Contam. Toxicol. 2011, 61, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Deckert, J.; Rucinska-Sobkowiak, R.; Gzyl, J.; Pawlak-Sprada, S.; Abramowski, D.; Jelonek, T.; Gwozdz, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Filipic, M. Mechanisms of cadmium induced genomic instability. Mutat. Res. 2012, 733, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.W.; Saito, M.A.; George, G.N.; Pickering, I.J.; Prince, R.C.; Morel, F.M. Biochemistry: A cadmium enzyme from a marine diatom. Nature 2005, 435, 42. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Song, B.; Morel, F.M. Diversity of the cadmium-containing carbonic anhydrase in marine diatoms and natural waters. Environ. Microbiol. 2007, 9, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Viparelli, F.; Monti, S.M.; De Simone, G.; Innocenti, A.; Scozzafava, A.; Xu, Y.; Morel, F.M.; Supuran, C.T. Inhibition of the r1 fragment of the cadmium-containing zeta-class carbonic anhydrase from the diatom thalassiosira weissflogii with anions. Bioorganic Med. Chem. Lett. 2010, 20, 4745–4748. [Google Scholar] [CrossRef]
- Xu, Y.; Supuran, C.T.; Morel, F.M.M. Cadmium-Carbonic Anhydrase. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Scott, R.A., Ed.; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Alterio, V.; Langella, E.; Viparelli, F.; Vullo, D.; Ascione, G.; Dathan, N.A.; Morel, F.M.; Supuran, C.T.; De Simone, G.; Monti, S.M. Structural and inhibition insights into carbonic anhydrase cdca1 from the marine diatom thalassiosira weissflogii. Biochimie 2012, 94, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A.M.; Traversa, U. Mechanisms of inhibitory effects of zinc and cadmium ions on agonist binding to adenosine a1 receptors in rat brain. Biochem. Pharmacol. 1999, 58, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Gautam, N.; Mishra, A.; Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 2011, 43, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, L.O.; Harbak, H.; Bennekou, P. Cobalt metabolism and toxicology—A brief update. Sci. Total Environ. 2012, 432, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, M.; Nunez, M.T. Iron and copper metabolism. Mol. Asp. Med. 2005, 26, 313–327. [Google Scholar] [CrossRef]
- Luevano, J.; Damodaran, C. A review of molecular events of cadmium-induced carcinogenesis. J. Environ. Pathol. Toxicol. Oncol. 2014, 33, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Jarup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.R.; Schoenung, J.M. Human health and ecological toxicity potentials due to heavy metal content in waste electronic devices with flat panel displays. J. Hazard. Mater. 2010, 177, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.B.; Maliwal, B.P.; Feliccia, V.L.; Fierke, C.A.; McCall, K. Determination of picomolar concentrations of metal ions using fluorescence anisotropy: Biosensing with a “reagentless” enzyme transducer. Anal. Chem. 1998, 70, 4717–4723. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.H.; Thompson, R.B.; Maliwal, B.P.; Fones, G.R.; Moffett, J.W.; Fierke, C.A. Real-time determination of picomolar free cu(ii) in seawater using a fluorescence-based fiber optic biosensor. Anal. Chem. 2003, 75, 6807–6812. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.A.; Lesburg, C.A.; Christianson, D.W.; Thompson, R.B.; Fierke, C.A. Active-site engineering of carbonic anhydrase and its application to biosensors. In The Carbonic Anhydrases: New Horizons; Chegwidden, W.R., Carter, N.D., Edwards, Y.H., Eds.; Springer, Birkhäuser Verlag: Basel, Switzerland, 2000; Volume 90, pp. 221–240. [Google Scholar]
- Boone, C.D.; Habibzadegan, A.; Gill, S.; McKenna, R. Carbonic anhydrases and their biotechnological applications. Biomolecules 2013, 3, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.K.; Wang, D.; Thompson, R.B.; Fierke, C.A. Carbonic anhydrase ii-based metal ion sensing: Advances and new perspectives. Biochim. Biophys. Acta 2010, 1804, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.S.; Ragavan, K.V. Biosensors in food processing. J. Food Sci. Technol. 2013, 50, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Rani, R. Development of biosensors for the detection of biological warfare agents: Its issues and challenges. Sci. Prog. 2013, 96, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Bozym, R.; Hurst, T.K.; Westerberg, N.; Stoddard, A.; Fierke, C.A.; Frederickson, C.J.; Thompson, R.B. Determination of zinc using carbonic anhydrase-based fluorescence biosensors. Methods Enzymol. 2008, 450, 287–309. [Google Scholar] [PubMed]
- Fierke, C.A.; Thompson, R.B. Fluorescence-based biosensing of zinc using carbonic anhydrase. Biometals 2001, 14, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Lindskog, S.; Nyman, P.O. Metal-binding properties of human erythrocyte carbonic anhydrases. Biochim. Biophys. Acta 1964, 85, 462–474. [Google Scholar] [PubMed]
- Chen, R.F.; Kernohan, J.C. Combination of bovine carbonic anhydrase with a fluorescent sulfonamide. J. Biol. Chem. 1967, 242, 5813–5823. [Google Scholar] [PubMed]
- Nair, S.K.; Elbaum, D.; Christianson, D.W. Unexpected binding mode of the sulfonamide fluorophore 5-dimethylamino-1-naphthalene sulfonamide to human carbonic anhydrase ii. Implications for the development of a zinc biosensor. J. Biol. Chem. 1996, 271, 1003–1007. [Google Scholar]
- Avvaru, B.S.; Busby, S.A.; Chalmers, M.J.; Griffin, P.R.; Venkatakrishnan, B.; Agbandje-McKenna, M.; Silverman, D.N.; McKenna, R. Apo-human carbonic anhydrase ii revisited: Implications of the loss of a metal in protein structure, stability, and solvent network. Biochemistry 2009, 48, 7365–7372. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.A.; Ahmed, M.; Fierke, C.A. Metal binding specificity in carbonic anhydrase is influenced by conserved hydrophobic core residues. Biochemistry 1999, 38, 9054–9062. [Google Scholar] [CrossRef] [PubMed]
- DiTusa, C.A.; McCall, K.A.; Christensen, T.; Mahapatro, M.; Fierke, C.A.; Toone, E.J. Thermodynamics of metal ion binding. 2. Metal ion binding by carbonic anhydrase variants. Biochemistry 2001, 40, 5345–5351. [Google Scholar]
- McCall, K.A.; Fierke, C.A. Probing determinants of the metal ion selectivity in carbonic anhydrase using mutagenesis. Biochemistry 2004, 43, 3979–3986. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.E.; Hunt, J.A.; Fierke, C.A.; Oas, T.G. Novel disulfide engineering in human carbonic anhydrase ii using the pairwise side-chain geometry database. Protein Sci. 2000, 9, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Frederickson, C.J.; Giblin, L.J.; Weiss, J.H.; Medvedeva, Y.; Bentley, P.A. Sensitive and selective detection of zinc ions in neuronal vesicles using pydpy1, a simple turn-on dipyrrin. Chem. Commun. (Camb.) 2011, 47, 7107–7109. [Google Scholar] [CrossRef]
- Thompson, R.B.; Maliwal, B.P.; Zeng, H.H. Zinc biosensing with multiphoton excitation using carbonic anhydrase and improved fluorophores. J. Biomed. Opt. 2000, 5, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.B.; Peterson, D.; Mahoney, W.; Cramer, M.; Maliwal, B.P.; Suh, S.W.; Frederickson, C.; Fierke, C.; Herman, P. Fluorescent zinc indicators for neurobiology. J. Neurosci. Methods 2002, 118, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.B.; Patchan, M.W. Lifetime-based fluorescence energy transfer biosensing of zinc. Anal. Biochem. 1995, 227, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Moulis, J.M. The bioinorganic chemistry of cadmium in the context of its toxicity. Met. Ions Life Sci. 2013, 11, 1–29. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alterio, V.; Langella, E.; De Simone, G.; Monti, S.M. Cadmium-Containing Carbonic Anhydrase CDCA1 in Marine Diatom Thalassiosira weissflogii. Mar. Drugs 2015, 13, 1688-1697. https://doi.org/10.3390/md13041688
Alterio V, Langella E, De Simone G, Monti SM. Cadmium-Containing Carbonic Anhydrase CDCA1 in Marine Diatom Thalassiosira weissflogii. Marine Drugs. 2015; 13(4):1688-1697. https://doi.org/10.3390/md13041688
Chicago/Turabian StyleAlterio, Vincenzo, Emma Langella, Giuseppina De Simone, and Simona Maria Monti. 2015. "Cadmium-Containing Carbonic Anhydrase CDCA1 in Marine Diatom Thalassiosira weissflogii" Marine Drugs 13, no. 4: 1688-1697. https://doi.org/10.3390/md13041688
APA StyleAlterio, V., Langella, E., De Simone, G., & Monti, S. M. (2015). Cadmium-Containing Carbonic Anhydrase CDCA1 in Marine Diatom Thalassiosira weissflogii. Marine Drugs, 13(4), 1688-1697. https://doi.org/10.3390/md13041688