
Cyanobacterial Toxic and Bioactive Peptides in Freshwater Bodies of Greece: Concentrations, Occurrence Patterns, and Implications for Human Health



Abstract
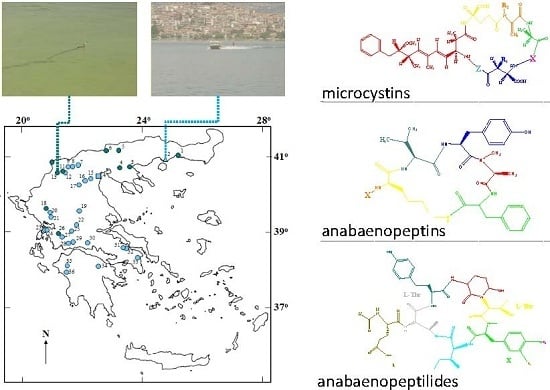
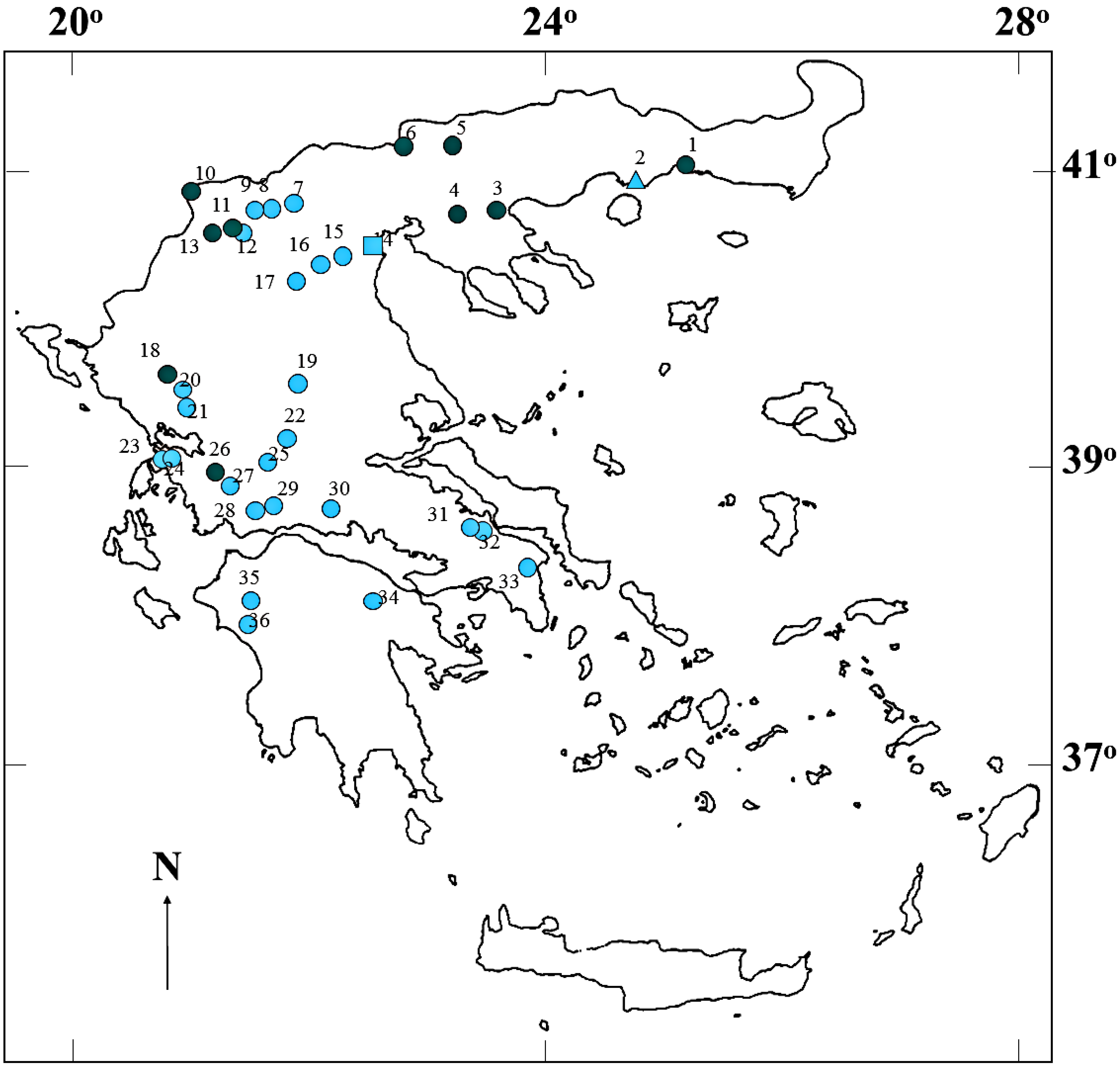
:
1. Introduction
2. Results
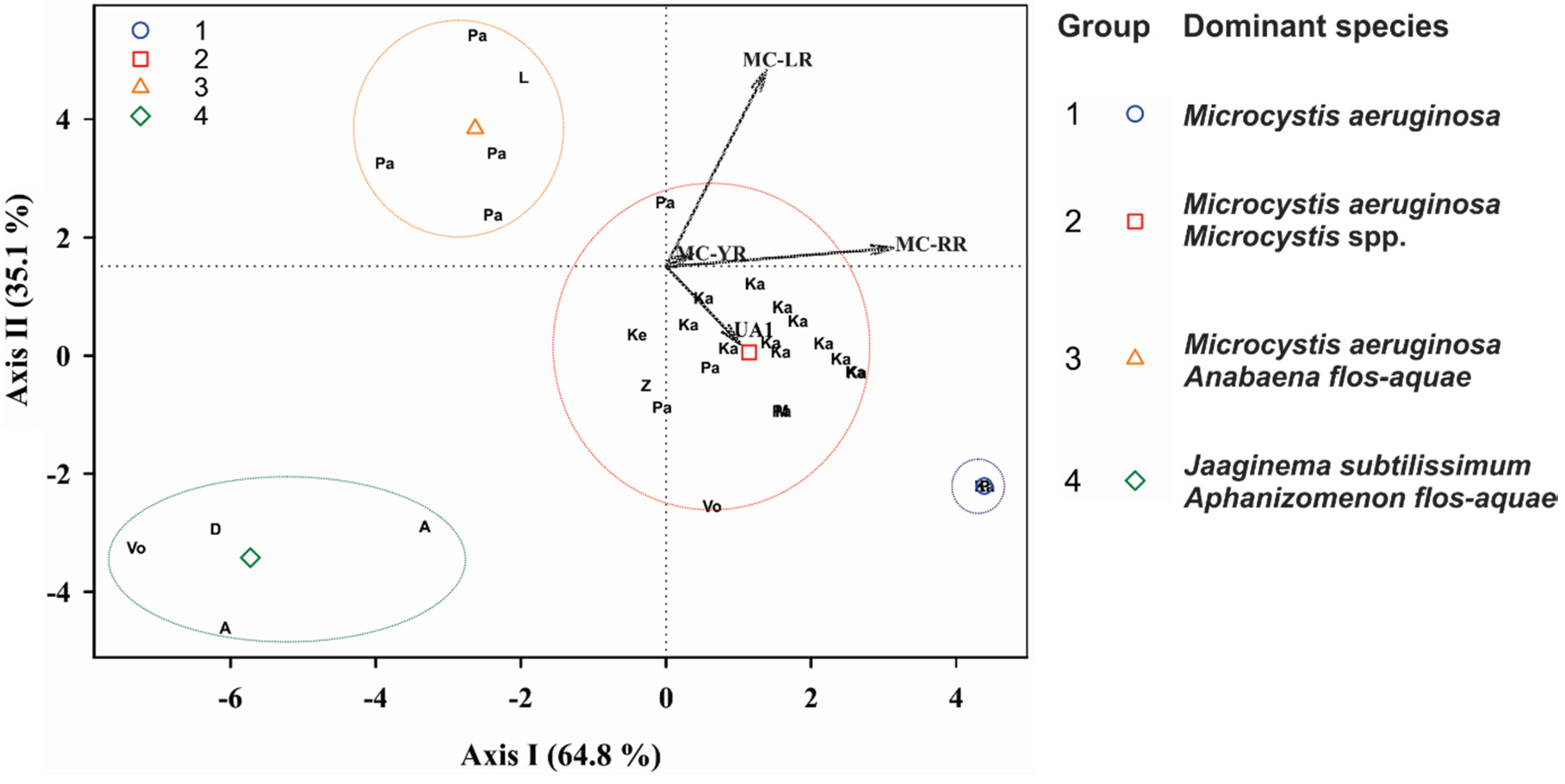
2.1. Peptide Diversity
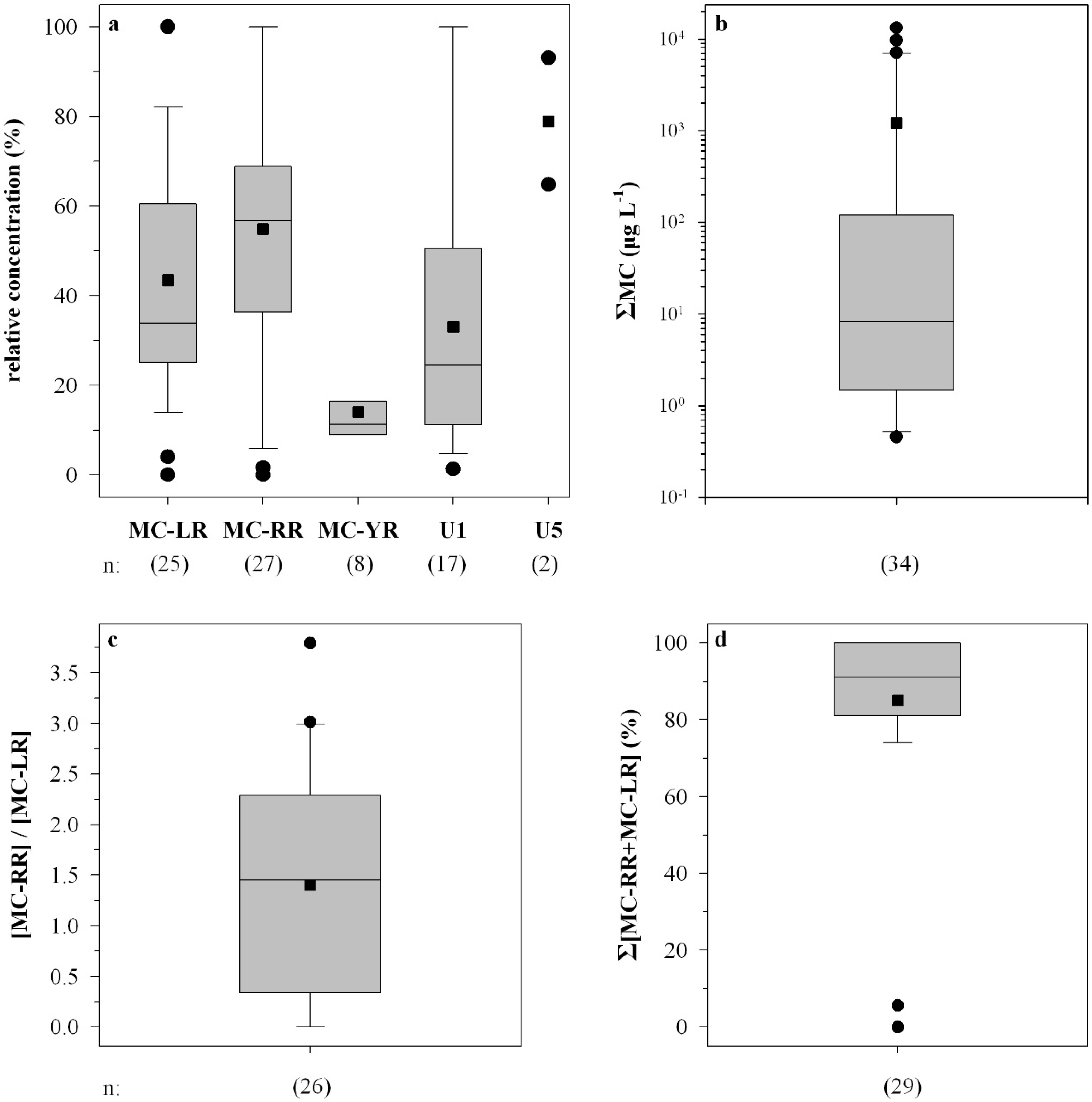
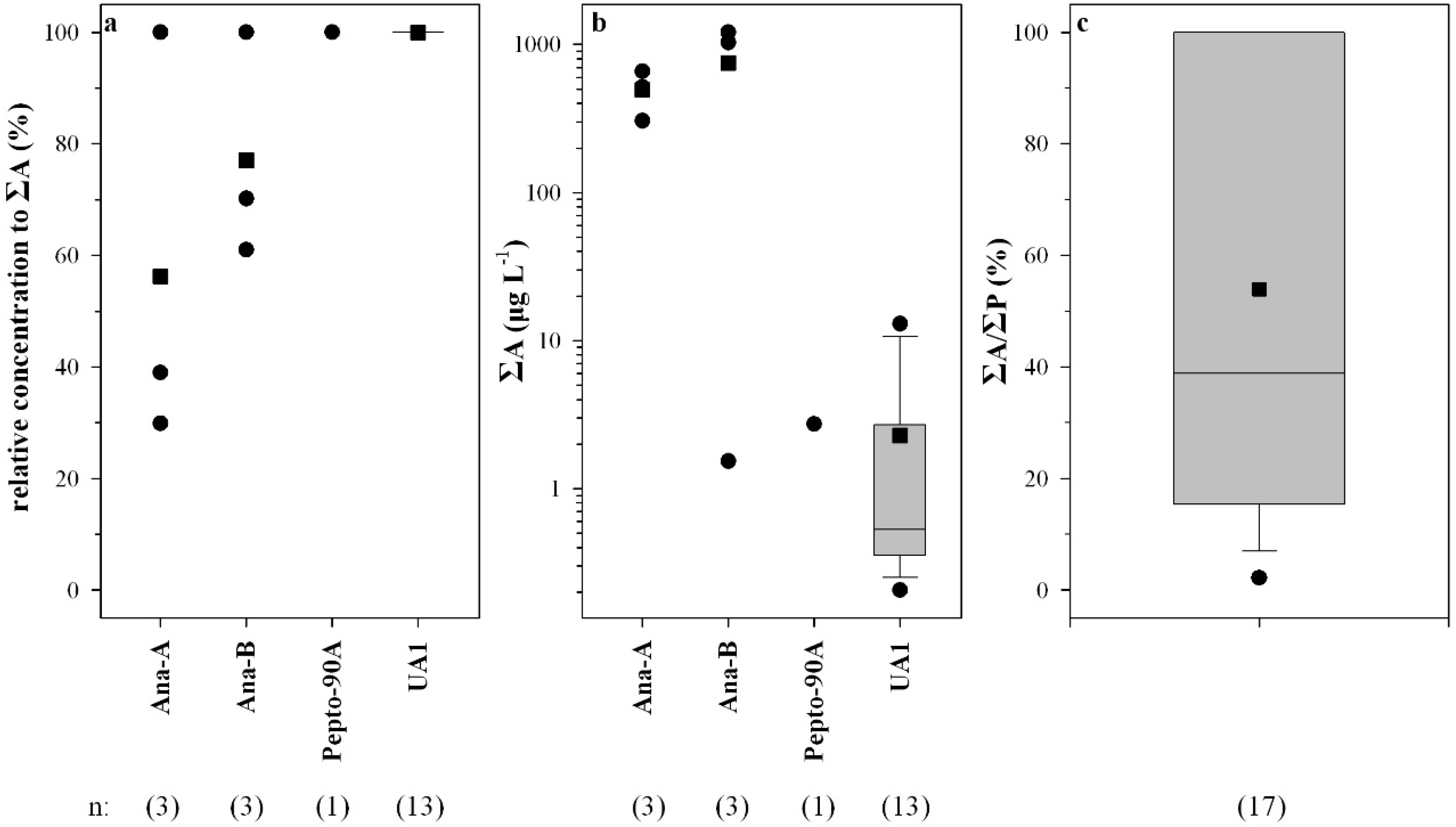
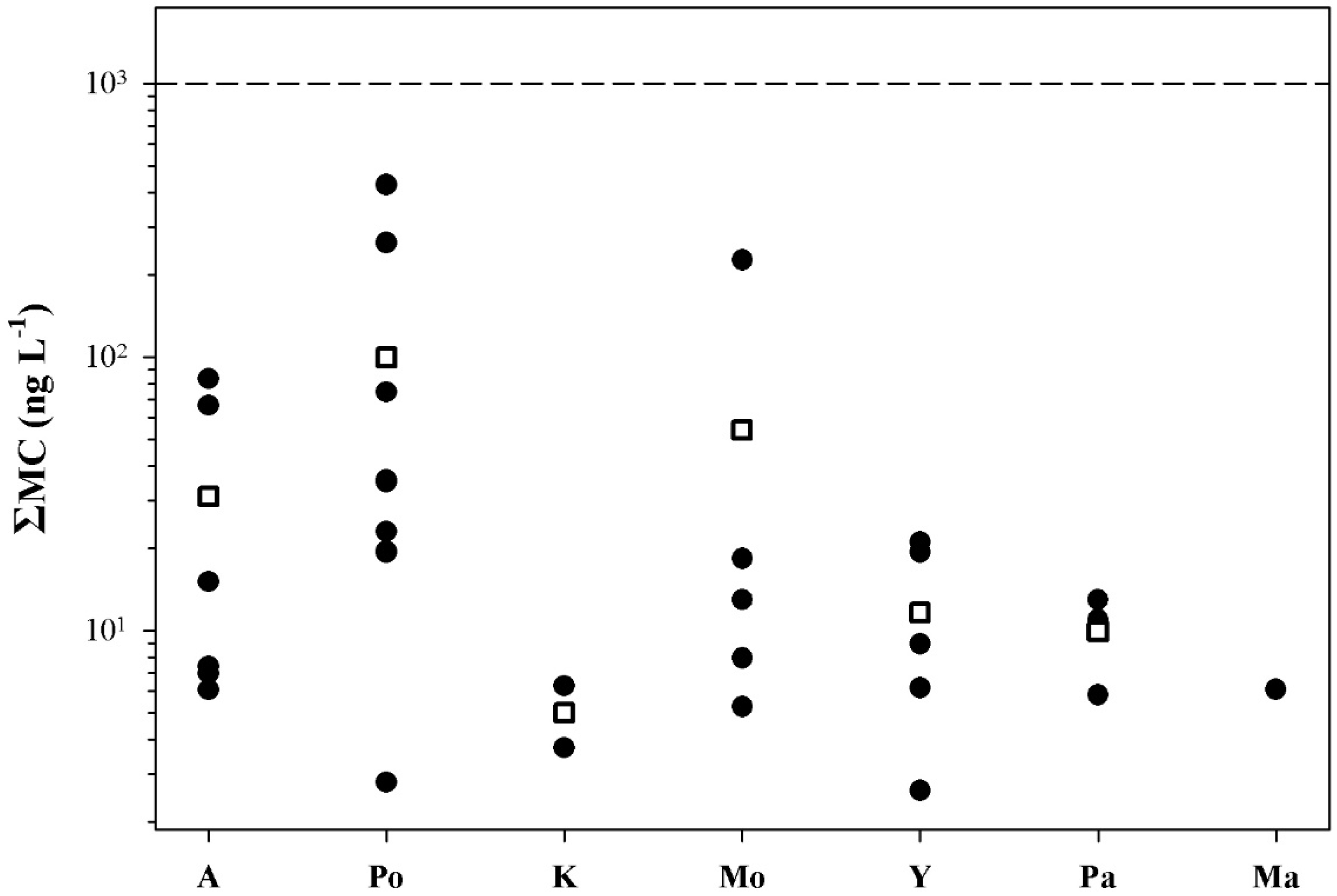
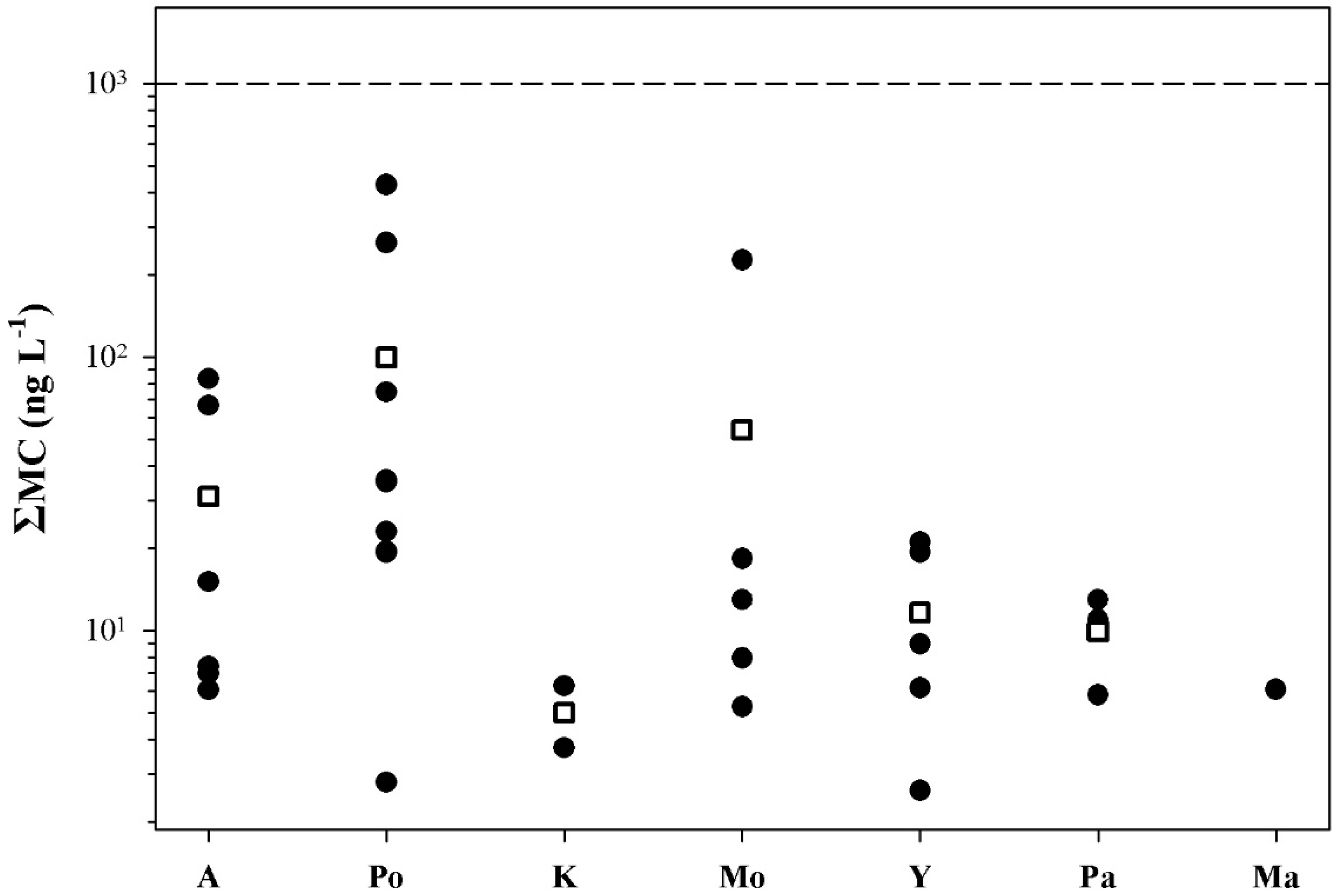
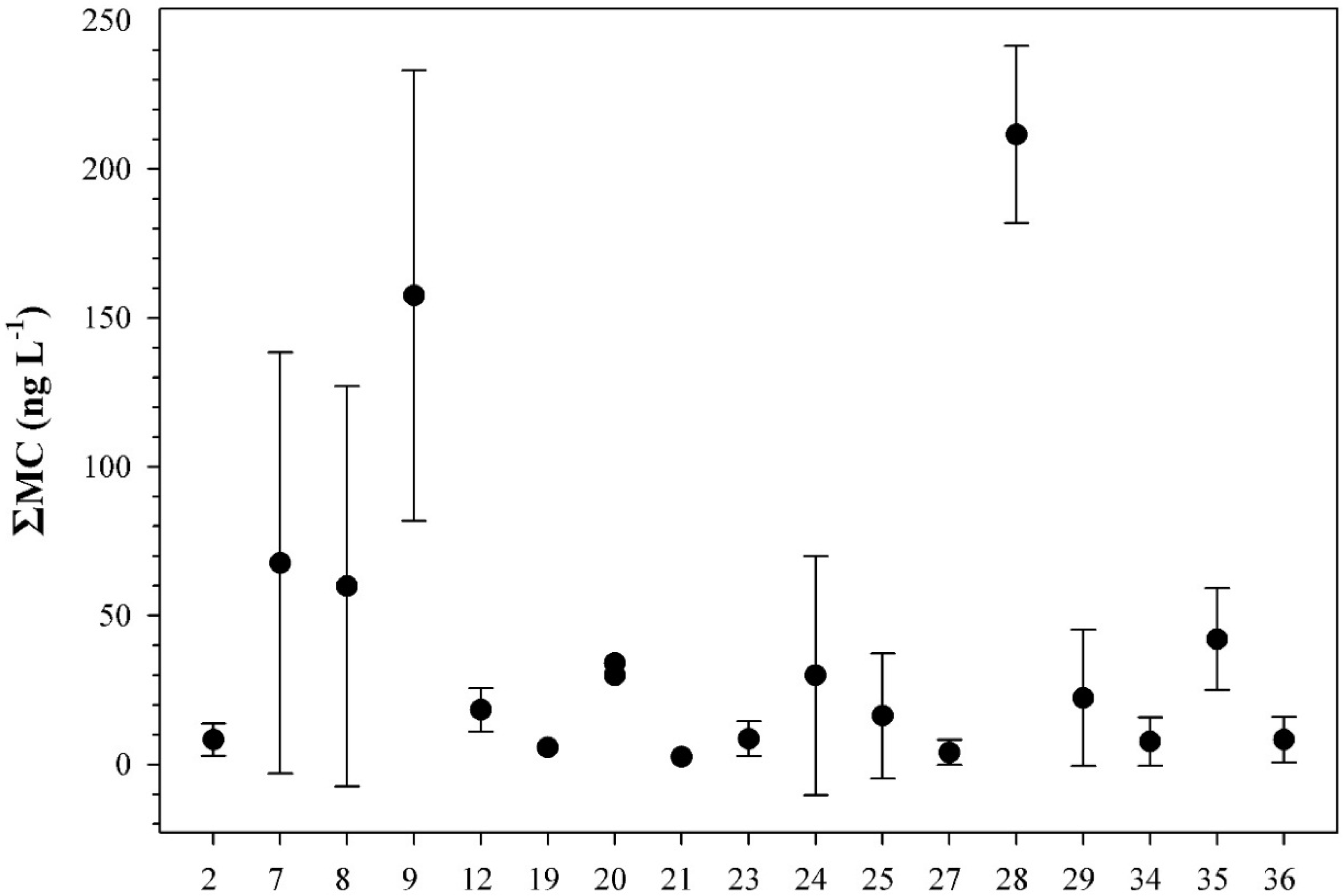
2.2. Peptide Concentrations and Occurrence Patterns

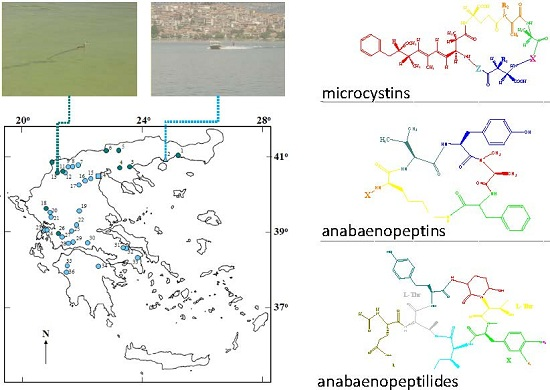{kind=link}
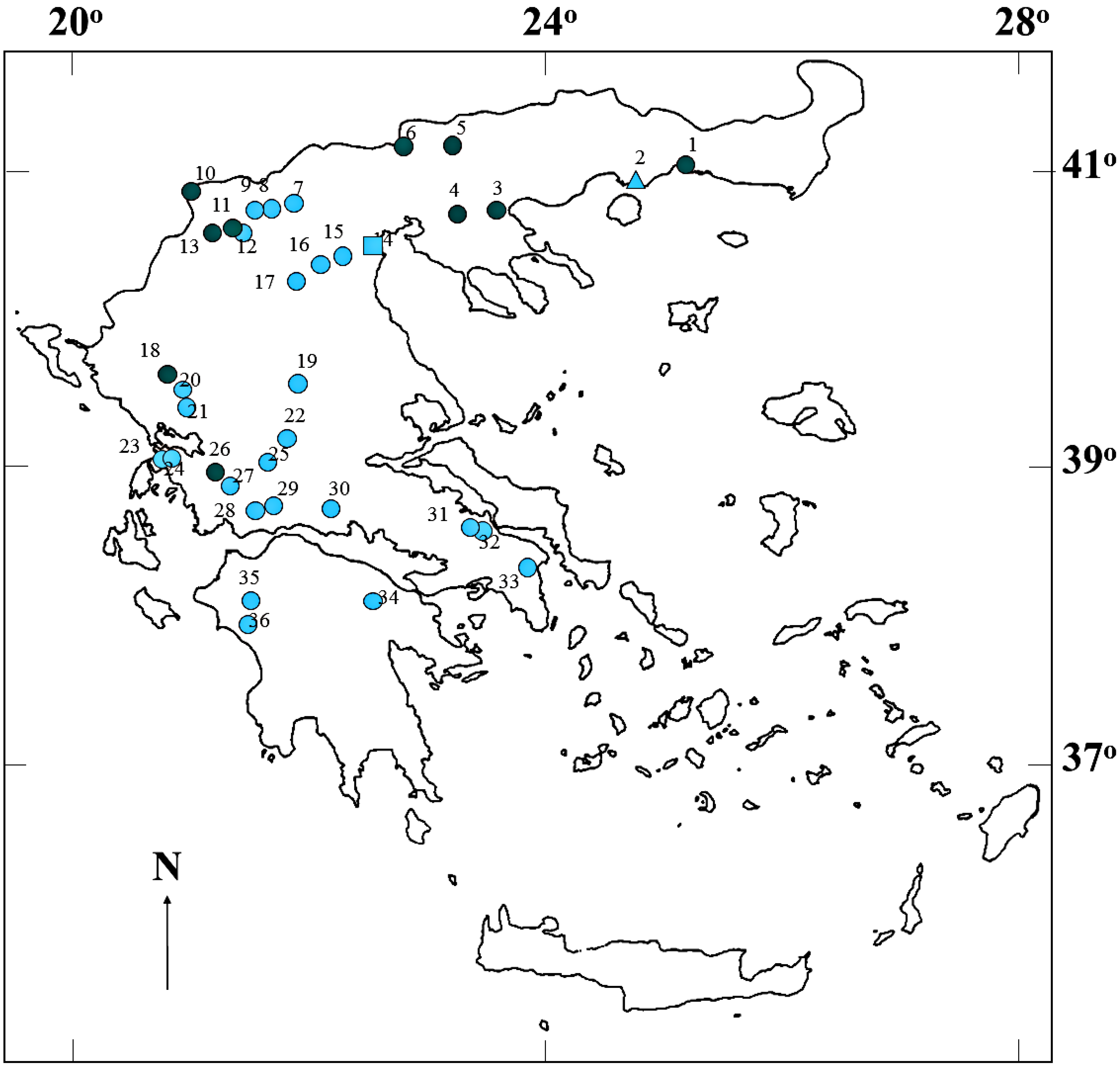{kind=link}
{kind=link}
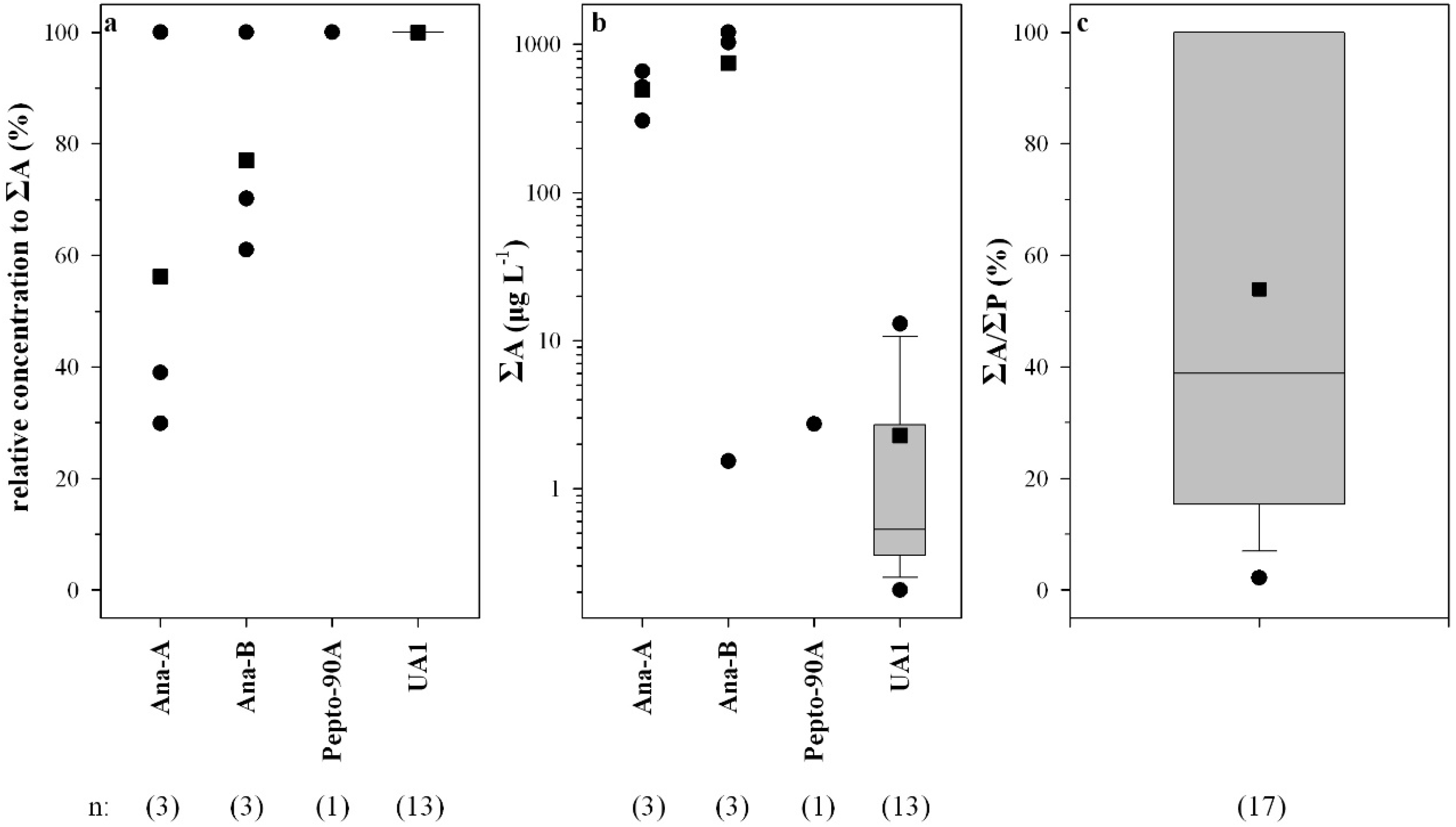{kind=link}
{kind=link}
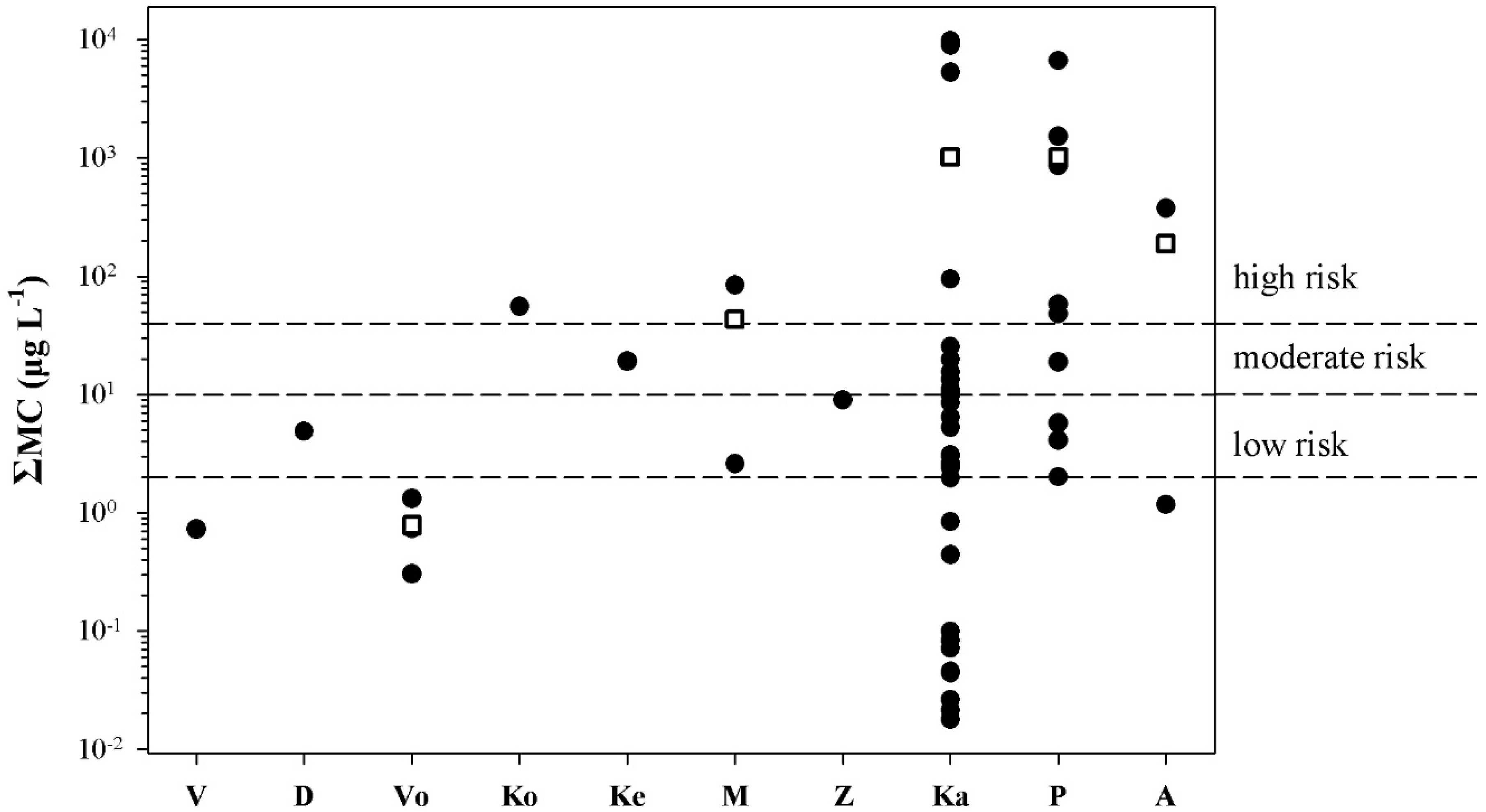{kind=link}
{kind=link}
{kind=link}
| Number of Samples | Sampling Frequency | ||
|---|---|---|---|
| 1 | 2–10 | >10 | |
| total (n) | 20 | 41 | 54 |
| with MC (n) | 15 | 39 | 54 |
| frequency (%) | 75 | 95 | 100 |
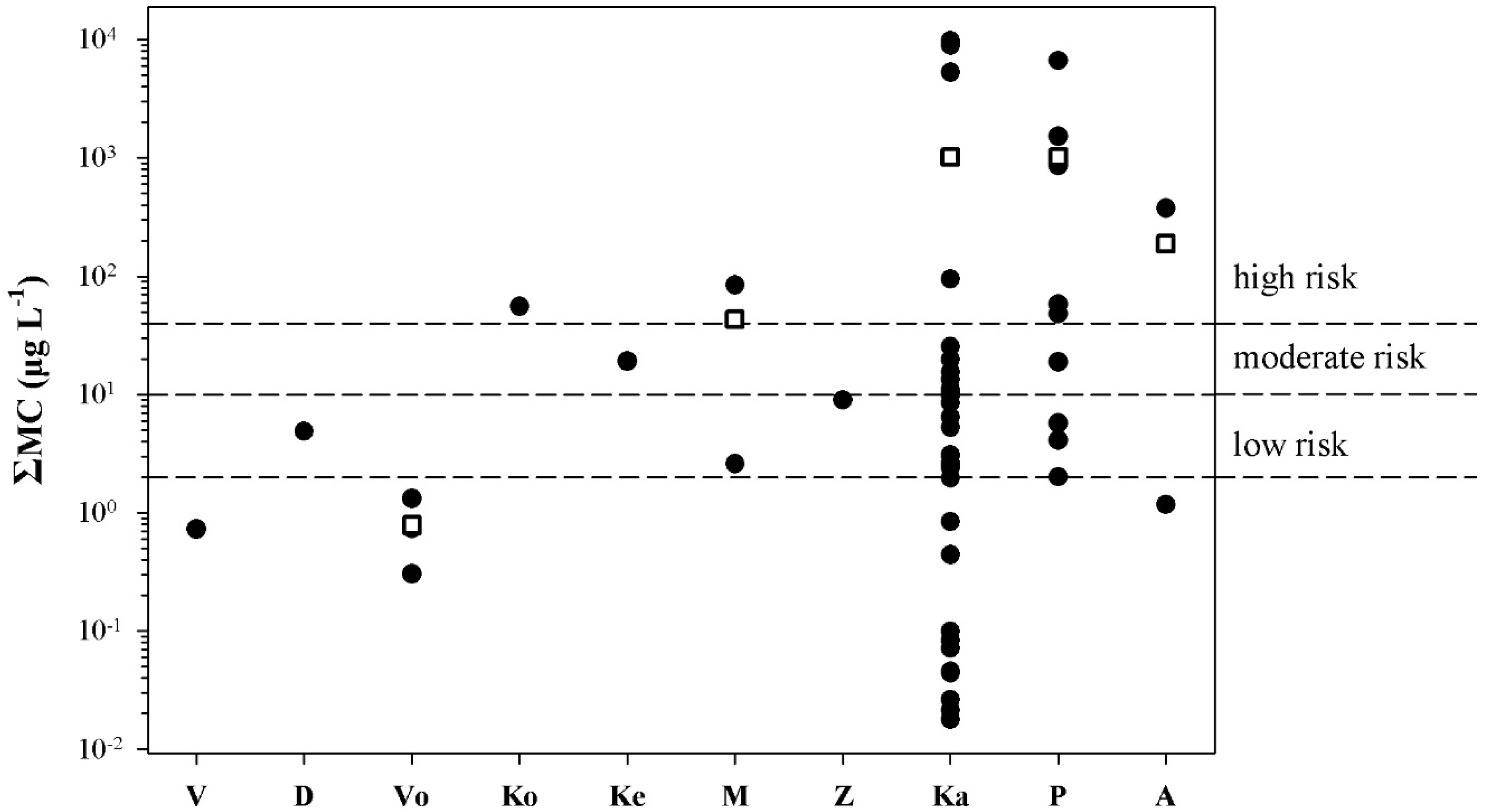
2.3. Risk Assessment
| Water Volume (L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Vistonis | Doirani | Volvi | Koronia | Kerkini | Mikri Prespa | Zazari | Kastoria | Pamvotis | Amvrakia | ||
| TDI a | child | 0.4 | 0.027 | 0.3 | 0.007 | 0.01 | 0.004 | 0.03 | 3.4 × 10−5 | 3.0 × 10−5 | 3.0 × 10−4 |
| adult | 2.2 | 0.16 | 1.8 | 0.04 | 0.09 | 0.02 | 0.18 | 2.1 × 10−3 | 1.8 × 10−3 | 3.0 × 10−3 | |
| NOAEL b | child | 376.6 | 27.7 | 302.8 | 7.2 | 15.8 | 3.7 | 30.1 | 0.034 | 0.031 | 0.5 |
| adult | 2259.8 | 166.0 | 1816.8 | 43.3 | 94.9 | 22.3 | 180.8 | 0.206 | 0.181 | 3.0 | |
| LOAEL c | child | 941.6 | 69.2 | 757 | 18 | 39.5 | 9.3 | 75.3 | 0.08 | 0.075 | 1.2 |
| adult | 5649.7 | 415.2 | 4542.1 | 108.3 | 237.3 | 55.8 | 452.1 | 0.516 | 0.45 | 7.5 | |
3. Discussion
4. Experimental Section
4.1. Sample Collection
4.2. Peptide Extraction, Assays, and High-Performance Liquid Chromatography
4.3. Risk Assessment
4.4. Statistical Analysis
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Codd, G.A.; Lindsay, J.; Young, F.M.; Morrison, L.F.; Metcalf, J.S. Harmful cyanobacteria: From mass mortalities to management measures. In Harmful Cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 3, pp. 1–23. [Google Scholar]
- Paerl, H.W.; Otten, G.T. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water, 1st ed.; Chorus, I., Bartram, J., Eds.; E. & F.N. Spon: London, UK, 1999; pp. 41–110. [Google Scholar]
- Chorus, I.; Fastner, J. Recreational exposure to cyanotoxins. In Cyanotoxins: Occurrence, Causes, Consequences; Chorus, I., Ed.; Springer: Berlin, Germany, 2001; pp. 190–199. [Google Scholar]
- World Health Organization (WHO). Guidelines for Safe Recreational Water Environments. Volume 1. Coastal and Fresh Waters; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- World Health Organization (WHO). Guidelines for Drinking-Water Quality, 4th ed.; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Welker, M.; von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Erhard, M.; Döhren, H. Determination of oligopeptide diversity within a natural population of Microcystis spp. (Cyanobacteria) by typing single colonies by matrix-assisted laser desorption ionization–time of flight mass spectrometry. Appl. Environ. Microbiol. 2001, 67, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Döhren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Elkobi-Peer, S.; Carmeli, S. New prenylated aeruginosin, microphycin, anabaenopeptin and micropeptin analogues from a Microcystis bloom material collected in Kibbutz Kfar Blum, Israel. Mar. Drugs 2015, 13, 2347–2375. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Harada, K.-I.; Suzuki, M.; Kondo, F.; Ikai, Y.; Oka, H.; Carmichael, W.W.; Sivonen, K. Occurrence of novel cyclic peptides together with microcystins from toxic cyanobacteria, Anabaena species. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 559–562. [Google Scholar]
- Fujii, K.; Sivonen, K.; Nakano, T.; Harada, K.-I. Structural elucidation of cyanobacterial peptides encoded by peptide synthetase gene in Anabaena species. Tetrahedron 2002, 58, 6863–6871. [Google Scholar] [CrossRef]
- Tonk, L.; Welker, M.; Huisman, J.; Visser, P.M. Production of cyanopeptolins, anabaenopeptins, and microcystins by the harmful cyanobacteria Anabaena 90 and Microcystis PCC 7806. Harmful Algae 2009, 8, 219–224. [Google Scholar] [CrossRef]
- Martins, J.; Saker, M.L.; Moreira, C.; Welker, M.; Fastner, J.; Vasconcelos, V.M. Peptide diversity in strains of the cyanobacterium Microcystis aeruginosa isolated from Portuguese water supplies. Appl. Microbiol. Biotechnol. 2009, 82, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Fastner, J.; Dittmann, E.; Christiansen, G.; Vasconcelos, V.M. Variation between strains of the cyanobacterium Microcystis aeruginosa isolated from a Portuguese river. J. Appl. Microbiol. 2005, 99, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Harjunpää, V.; Lanaras, T.; Sivonen, K. Diversity of hepatotoxic microcystins and bioactive anabaenopeptins in cyanobacterial blooms from Greek freshwaters. Environ. Toxicol. 2005, 20, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, T.; Kagalou, I.; Bacopoulos, V.; Leonardos, I.D. Accumulation of microcystins in water and fish tissues: An estimation of risks associated with microcystins in most of the Greek Lakes. Environ. Toxicol. 2010, 25, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Papadimitriou, T.; Zaoutsos, N.; Leonardos, I. Anthropogenic and climate-induced change favors toxic cyanobacteria blooms: Evidence from monitoring a highly eutrophic, urban Mediterranean lake. Harmful Algae 2014, 39, 322–333. [Google Scholar] [CrossRef]
- Gkelis, S.; Zaoutsos, N. Cyanotoxin occurrence and potentially toxin producing cyanobacteria in freshwaters of Greece: A multi-disciplinary approach. Toxicon 2014, 78, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Moustaka-Gouni, M.; Sivonen, K.; Lanaras, T. First report of the cyanobacterium Aphanizomenon ovalisporum Forti in two Greek lakes and cyanotoxin occurrence. J. Plankton Res. 2005, 27, 1295–1300. [Google Scholar] [CrossRef]
- Cook, C.M.; Vardaka, E.; Lanaras, T. Toxic cyanobacteria in Greek freshwaters, 1987–2000: Occurrence, toxicity, and impacts in the Mediterranean region. Acta Hydrochim. Hydrobiol. 2004, 32, 107–124. [Google Scholar] [CrossRef]
- Fastner, J.; Wirsing, B.; Wiedner, C.; Heinze, R.; Neumann, U.; Chorus, I. Microcystins and hepatocyte toxicity. In Cyanotoxins: Occurrence, Causes, Consequences; Chorus, I., Ed.; Springer: Berlin, Germany, 2001; pp. 22–37. [Google Scholar]
- Gkelis, S. Characterisation of Planktic Cyanobackteria and Their Bioactive Peptides. Ph.D. Thesis, Scientific Annals of the School of Biology, Aristotle University of Thessaloniki, Thessaloniki, Greece, March 2006. Available online: http://thesis.ekt.gr/thesisBookReader/id/15304#page/1/mode/2up (accessed on 3 August 2015). [Google Scholar]
- Vardaka, E.; Moustaka-Gouni, M.; Cook, C.M.; Lanaras, T. Cyanobacterial blooms and water quality in Greek waterbodies. J. Appl. Phycol. 2005, 17, 391–401. [Google Scholar] [CrossRef]
- Znachor, P.; Jurczak, T.; Komárková, J.; Jezberová, J.; Mankiewicz, J.; Kaštovská, K.; Zapomělova, E. Summer changes in cyanobacterial bloom composition and microcystin concentration in eutrophic Czech reservoirs. Environ. Toxicol. 2006, 21, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Dyble, J.; Fahnenstiel, G.L.; Litaker, W.L.; Millie, D.F.; Tester, P.A. Microcystin concentrations and genetic diversity of Microcystis in the Lower Great Lakes. Environ. Toxicol. 2008, 23, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Fewer, D.; Rohrlack, T.; Jokela, J.; Rouhiainen, L.; Sivonen, K.; Kristensen, T.; Jakobsen, K.S. Evidence for positive selection acting on microcystin synthetase adenylation domains in three cyanobacterial genera. BMC Evol. Biol. 2008, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Suzuki, S.; Ishida, K.; Murakami, M. Anabaenopeptins G and H, potent carboxypeptidase A inhibitors from the cyanobacterium Oscillatoria agardhii (NIES-595). Bioor. Med. Chem. Lett. 1999, 9, 1243–1246. [Google Scholar] [CrossRef]
- Gkelis, S.; Lanaras, T.; Sivonen, K. The presence of microcystins and other cyanobacterial bioactive peptides in aquatic fauna collected from Greek freshwaters. Aquat. Toxicol. 2006, 78, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Grach-Pogrebinsky, O.; Sedmak, B.; Carmeli, S. Protease inhibitors from a Slovenian lake Bled toxic waterbloom of the cyanobacterium Planktothrix rubescens. Tetrahedron 2003, 59, 8329–8336. [Google Scholar] [CrossRef]
- Kurmayer, R.; Schober, E.; Tonk, L.; Visser, P.M.; Christiansen, G. Spatial divergence in the proportions of genes encoding toxic peptide synthesis among populations of the cyanobacterium Planktothrix in European lakes. FEMS Microbiol. Lett. 2011, 317, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Repka, S.; Koivula, M.; Harjunpää, V.; Rouhiainen, L.; Sivonen, K. Effects of phosphate and light on growth of and bioactive peptide production by the cyanobacterium Anabaena strain 90 and its anabaenopeptilide mutant. Appl. Environ. Microbiol. 2004, 70, 4551–4560. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, P.; Fabbrocino, S.; Cerulo, M.G.; Bruno, M.; Serpe, L.; Gallo, P. Characterisation of biotoxins produced by a cyanobacteria bloom in Lake Averno using two LC-MS based techniques. Food Addit. Contam. 2008, 25, 1530–1537. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Cordero, P.; Agha, R.; Cirésa, S.; Lezcanoa, M.Á.; Sánchez-Contrerasa, M.; Waara, K.-O.; Utkilen, H.; Quesada, A. Effects of harmful cyanobacteria on the freshwater pathogenic free-living amoeba Acanthamoeba castellanii. Aquat. Toxicol. 2013, 130–131, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Nasri, A.B.; Bouaicha, N.; Fastner, J. First report of a microcystin-containing bloom of the cyanobacteria Microcystis spp. in Lake Oubeira, eastern Algeria. Arch. Environ. Contam. Toxicol. 2004, 46, 197–202. [Google Scholar] [PubMed]
- Fastner, J.; Neumann, U.; Wirsing, B.; Weckesser, J.; Wiedner, C.; Nixdorf, B.; Chorus, I. Microcystins (hepatotoxic heptapeptides) in German fresh water bodies. Environ. Toxicol. 1999, 14, 13–22. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Luukkainen, R.; Färdig, M.; Rouhiainen, L.; Evans, W.R.; Carmichael, W.W.; Rinehart, K.L.; Niemelä, S.I. Variation of cyanobacterial hepatotoxins in Finland. In The Contaminants in the Nordic Ecosystem: Dynamics, Processes & Fate; Munawar, M., Luotola, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherland, 1995; pp. 163–169. [Google Scholar]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R.; Bartram, J.; Chorus, I.; Kuiper-Goodman, T.; Utkilen, H.; Burch, M.; Codd, G.A. Safe levels and safe practices. In Toxic Cyanobacteria in Water, 1st ed.; Chorus, I., Bartram, J., Eds.; E. & F.N. Spon: London, UK, 1999; pp. 155–178. [Google Scholar]
- Kuiper-Goodman, T.; Falconer, I.; Fitzgerald, J. Human health aspects. In Toxic Cyanobacteria in Water; Chorus, I., Bartram, J., Eds.; E. & F.N. Spon: London, UK, 1999; pp. 113–153. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical ecology, Developments in Environmental Modeling, 2nd English ed.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1998; Volume 20. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gkelis, S.; Lanaras, T.; Sivonen, K. Cyanobacterial Toxic and Bioactive Peptides in Freshwater Bodies of Greece: Concentrations, Occurrence Patterns, and Implications for Human Health. Mar. Drugs 2015, 13, 6319-6335. https://doi.org/10.3390/md13106319
Gkelis S, Lanaras T, Sivonen K. Cyanobacterial Toxic and Bioactive Peptides in Freshwater Bodies of Greece: Concentrations, Occurrence Patterns, and Implications for Human Health. Marine Drugs. 2015; 13(10):6319-6335. https://doi.org/10.3390/md13106319
Chicago/Turabian StyleGkelis, Spyros, Thomas Lanaras, and Kaarina Sivonen. 2015. "Cyanobacterial Toxic and Bioactive Peptides in Freshwater Bodies of Greece: Concentrations, Occurrence Patterns, and Implications for Human Health" Marine Drugs 13, no. 10: 6319-6335. https://doi.org/10.3390/md13106319
APA StyleGkelis, S., Lanaras, T., & Sivonen, K. (2015). Cyanobacterial Toxic and Bioactive Peptides in Freshwater Bodies of Greece: Concentrations, Occurrence Patterns, and Implications for Human Health. Marine Drugs, 13(10), 6319-6335. https://doi.org/10.3390/md13106319