Global and Phylogenetic Distribution of Quorum Sensing Signals, Acyl Homoserine Lactones, in the Family of Vibrionaceae

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Method Performance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Limit of Detection (nmol L−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| C4 | C6 | C8 | C10 | O-C6 | O-C8 | O-C10 | OH-C6 | |
| UHPLC-HRMS a | 40 | 25 | 30 | 70 | 25 | 150 | 125 | 14 |
| C. violaceum b | 10,200 | 250 | 220 | 15,700 | 235 | 200 | 1850 | 230 |
| A. tumefaciens b | ND | 12,600 | 22 | 200 | 25 | 10 | 2 | 230 |
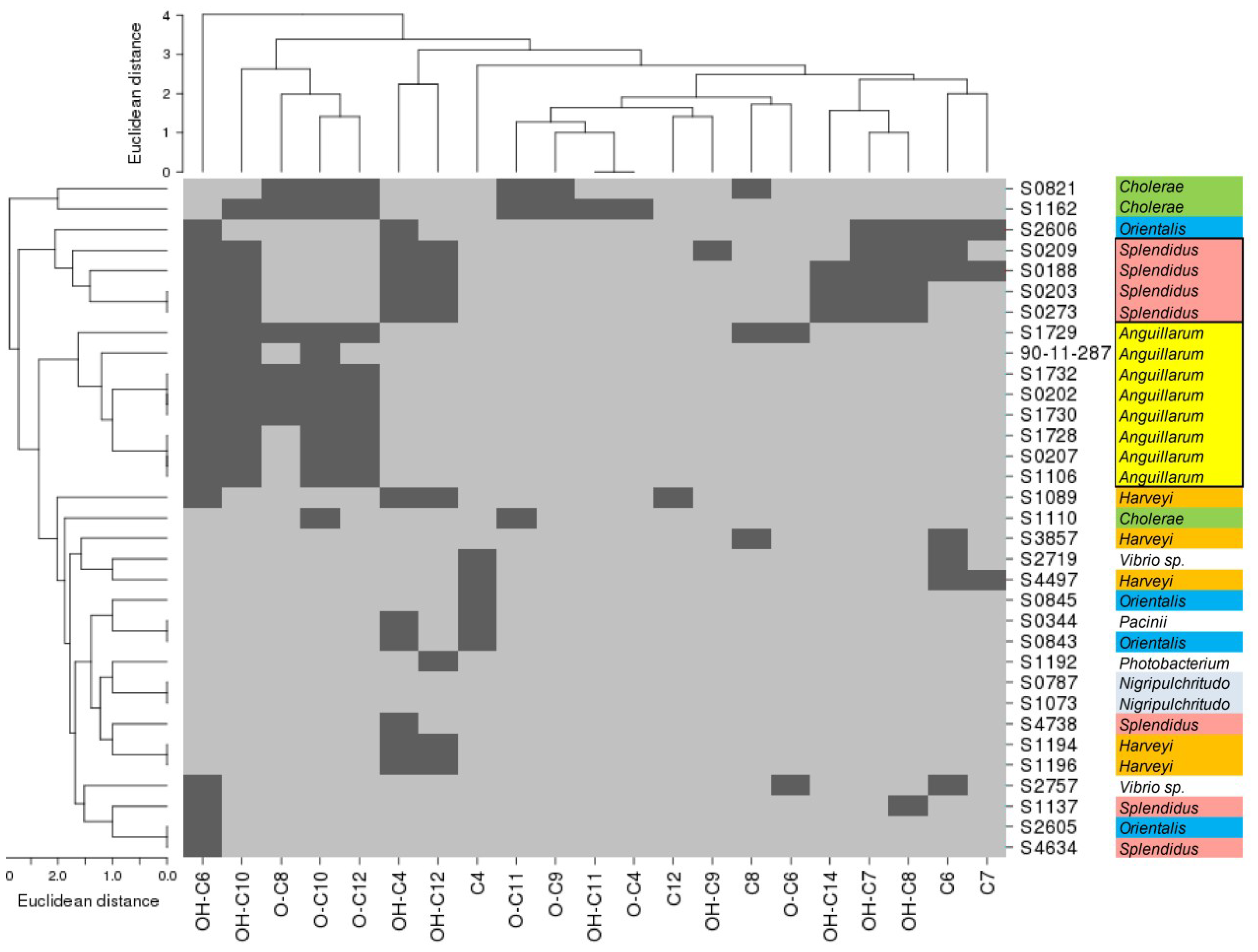
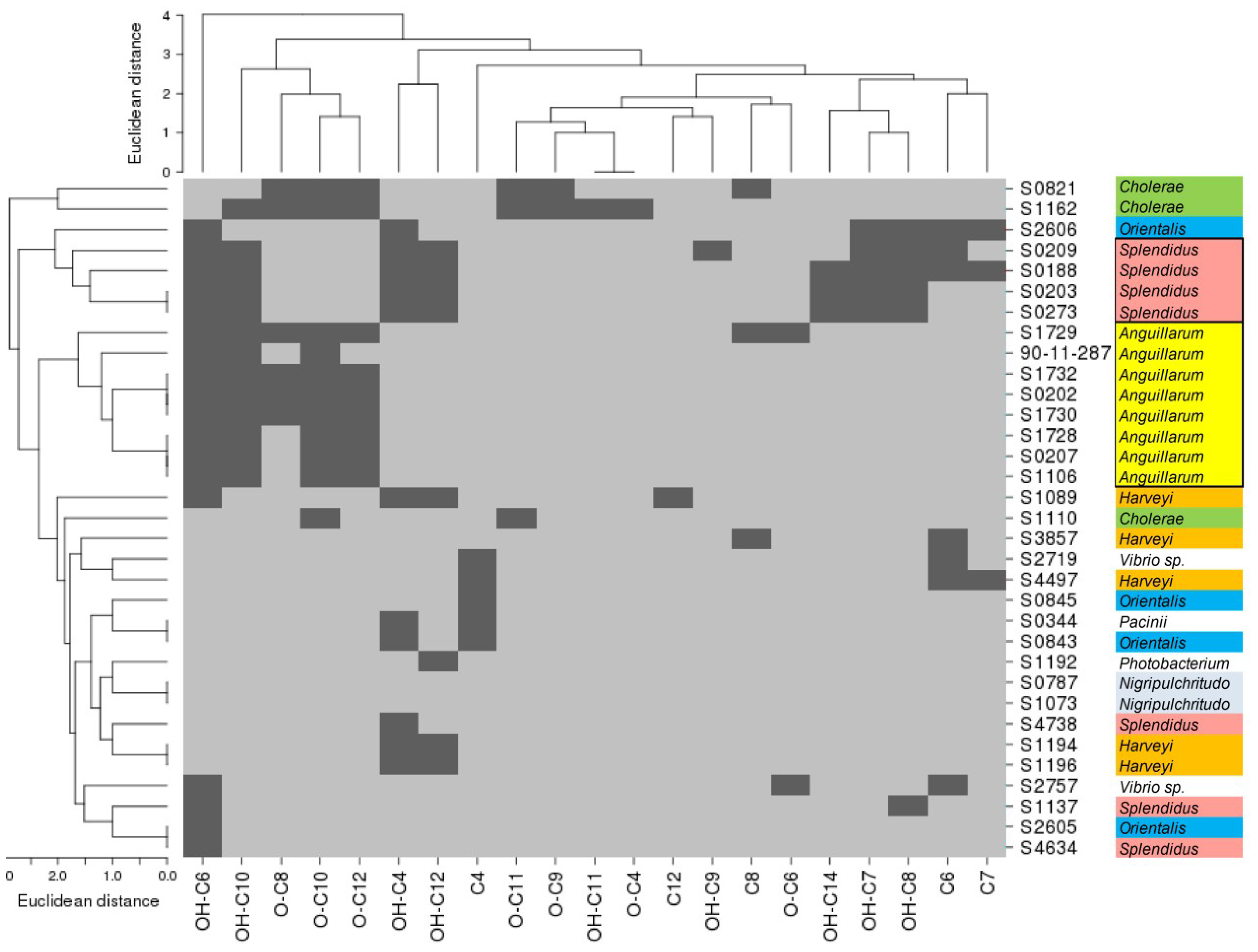
2.2. AHL-Producing Vibrionaceae Strains
| Strain | 16S rRNA GenBank Acc. No. | Closest Relative by MLSA | fur Gene Analysis | Isolation Site | ||
|---|---|---|---|---|---|---|
| Species | E Value | Latitude | Longitude | |||
| S0188 | FJ457302 | V. lentus a | V. splendidus | 0 | 62.03815 | −9.99592 |
| S0202 | FJ457304 | V. anguillarum a | V. anguillarum b | 0 | 62.03815 | −9.99592 |
| S0203 | FJ457305 | V. lentus a | V. splendidus | 0 | 62.03815 | −9.99592 |
| S0207 | KM273118 | V. anguillarum | V. anguillarum b | 0 | 62.16527 | −16.5731 |
| S0209 | FJ457309 | Vibrio sp. | V. splendidus | 0 | 62.16527 | −16.5731 |
| S0273 | FJ457310 | V. lentus a | V. splendidus | 0 | 62.2569 | −20.8819 |
| S0344 | FJ457313 | V. pacinii a | V. pacinii | 0 | 66.7494 | −53.8954 |
| S0787 | FJ457334 | V. mediterranei a | V. shilonii | 0 | 33.7571 | −25.4239 |
| S0821 | FJ457339 | V. furnissii | V. fluvialis | 0 | 23.0918 | −24.0417 |
| S0843 | FJ457343 | V. brasiliensis | V. brasiliensis | 0 | 12.20592 | −21.0234 |
| S0845 | FJ457345 | V. brasiliensis | V. brasiliensis | 0 | 12.20592 | −21.0234 |
| S1073 | FJ457350 | V. nigripulchritudo | V. nigripulchritudo | 0 | 1.6207 | −10.5021 |
| S1089 | FJ457355 | V. campbellii | V. campbelli | 0 | 4.570467 | −1.72975 |
| S1106 | FJ457363 | V. anguillarum | V. anguillarum b | 0 | 4.570467 | −1.72975 |
| S1110 | FJ457366 | V. fluvialis | V. fluvialis | 0 | 4.570467 | −1.72975 |
| S1137 | FJ457371 | V. chagasii a | V. splendidus | 9.32E−158 | 4.9119 | −0.3376 |
| S1162 | FJ457375 | V. fluvialis | V. fluvialis | 0 | 4.570467 | −1.72975 |
| S1192 | KM273119 | Photobacterium angustum a | ND | 4.570467 | −1.72975 | |
| S1194 | KM273120 | V. campbellii | V. campbelli | 0 | 4.570467 | −1.72975 |
| S1196 | KM273121 | V. campbellii | V. campbelli | 0 | 4.570467 | −1.72975 |
| S1728 | FJ457408 | V. anguillarum | V. anguillarum b | 2.52E−170 | −19.7461 | 114.8573 |
| S1729 | FJ457409 | V. anguillarum | V. anguillarum b | 0 | −19.7461 | 114.8573 |
| S1730 | FJ457410 | V. anguillarum | V. anguillarum b | 0 | −19.7461 | 114.8573 |
| S1732 | FJ457411 | V. anguillarum | V. anguillarum b | 0 | −19.7461 | 114.8573 |
| S2605 | FJ457458 | V. brasiliensis a | V. tubiashii | 3.02E−144 | −10.3454 | 157.7956 |
| S2606 | FJ457459 | V. brasiliensis a | V. tubiashii | 3.02E−144 | −10.3454 | 157.7956 |
| S2719 | FJ457471 | Vibrio sp. | V. brasiliensis | 2.35E−154 | −8.1005 | 156.8451 |
| S2757 | FJ457478 | Vibrio sp. | V. tubiashii | 2.88E−170 | −15.2329 | 156.665 |
| S3857 | FJ457565 | Vibrio sp. | V. rotiferianus | 0 | −14.159 | −77.4004 |
| S4497 | FJ457596 | Vibrio sp. | V. rotiferianus | 0 | 24.9963 | −67.0246 |
| S4634 | FJ457597 | V. splendidus | V. splendidus | 3.25E−116 | 43.0309 | −66.2774 |
| S4738 | FJ457608 | Vibrio sp. | V. splendidus b | 0 | 58.8041 | −3.0564 |
| Strain | AHL Response in | No of AHL/Strain | Peak Area (1000 Counts) of AHL in Chromatogram | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biomass Screen | Extract Well Assay | |||||||||||||||||||||||||
| Cv | At | Cv | At | C4 | C6 | C7 | C8 | C12 | OH-C4 | OH-C6 | OH-C4 | OH-C6 | OH-C9 | OH-C10 | OH-C11 | OH-C12 | OH-C14 | O-C4 | O-C6 | O-C8 | O-C9 | O-C10 | O-C11 | O-C12 | ||
| S0188 a | − | + | + | + | 9 | ND | (114) ᶜ | 178 ᵇ | ND | ND | 32 b (161) b | 880 b (4822) b | 32 b (161) b | 880 b (4822) b | ND | 46 c (561) b | ND | 12 c (163) b | (21313) b | ND | ND | ND | ND | ND | ND | ND |
| S0202 | − | + | − | + | 5 | ND | ND | ND | ND | ND | ND | 118 c (1659) b | ND | 118 c (1659) b | ND | (112) b | ND | ND | ND | ND | ND | (182) b | ND | 90 c (3385) b | ND | (30) b |
| S0203 a | − | + | + | + | 7 | ND | ND | ND | ND | ND | 40 b (3709) b | 964 b (6479) b | 40 b (3709) b | 964 b (6479) b | ND | 58 c (412) b | ND | 16 c (256) b | (94) b | ND | ND | ND | ND | ND | ND | ND |
| S0207 | + | + | − | + | 4 | ND | ND | ND | ND | ND | ND | 167 c (1807) b | ND | 167 c (1807) b | ND | (70) b | ND | ND | ND | ND | ND | ND | ND | 96 c (3637) b | ND | (43) b |
| S0209 a | + | + | + | + | 8 | ND | (116) ᵇ | ND | ND | ND | 17 c (342) b | 627 b (4297) b | 17ᶜ (342) b | 627 b (4297) ᵇ | (30) b | 28 c (452) b | ND | (64) b | ND | ND | ND | ND | ND | ND | ND | ND |
| S0273 a | + | + | + | + | 7 | ND | ND | ND | ND | ND | 28 b (273) b | 896 b (5918) b | 28 b (273) b | 896 b (5918) b | ND | 75 c (485) b | ND | 17 c (278) b | (115) b | ND | ND | ND | ND | ND | ND | ND |
| S0344 | + | − | − | − | 2 | 92 b (5352) b | ND | ND | ND | ND | (50)ᵇ | ND | (50)ᵇ | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S0787 | − | + | − | + | 0 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S0821 | + | + | + | + | 6 | ND | ND | ND | 57 b (6284) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | (294) b | 11 c (423) b | (2991) b 82 c | 10 c | (72) b |
| S0843 a | + | − | + | + | 2 | 317 b (11974) b | ND | ND | ND | ND | (94)ᵇ | ND | (94)ᵇ | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S0845 | + | − | − | − | 1 | 83 b (5009) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S1073 a | − | + | − | + | 0 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S1089 | − | + | − | + | 4 | ND | ND | ND | ND | (125322) b | 310 b (2279) b | (119) b | 310 b (2279) b | (119) b | ND | ND | ND | 42 c (1313) b | ND | ND | ND | ND | ND | ND | ND | ND |
| S1106 | + | + | − | + | 4 | ND | ND | ND | ND | ND | ND | 140 c (2050) b | ND | 140 c (2050) b | ND | (93) c | ND | ND | ND | ND | ND | ND | ND | 86 c (3661) b | ND | (41) b |
| S1110 | + | + | − | + | 2 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | 58 c (2770) b | (305) b | ND |
| S1137 | + | + | − | + | 2 | ND | ND | ND | ND | ND | ND | 210 c (3154) b | ND | 210 c (3154) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S1162 | − | + | − | + | 8 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | 239 c (134) b | 59 b (503) b | ND | ND | 30 b | ND | 12 c (273) b | 31 c (620) b | 241 c (5862) b | 623 b (10784) b | 12 c |
| S1192 | − | + | − | + | 1 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | (104) b | ND | ND | ND | ND | ND | ND | ND | ND |
| S1194 | − | + | − | − | 2 | ND | ND | ND | ND | ND | 61 b (491) b | ND | 61 b (491) b | ND | ND | ND | ND | (215) b | ND | ND | ND | ND | ND | ND | ND | ND |
| S1196 | − | + | − | + | 2 | ND | ND | ND | ND | ND | 163 b (1425) b | ND | 163 b (1425) b | ND | ND | ND | ND | ND (80) b | ND | ND | ND | ND | ND | ND | ND | ND |
| S1728 | − | + | − | + | 4 | ND | ND | ND | ND | ND | ND | 61 c (617) b | ND | 61 c (617) b | ND | (78) b | ND | ND | ND | ND | ND | ND | ND | 89 c (3069) b | ND | (106) b |
| S1729 | − | + | + | + | 7 | ND | ND | ND | 53 b (5170) b | ND | ND | 268 c (2943) b | ND | 268 c (2943) b | ND | 17 b (124) b | ND | ND | ND | ND | (303) b | 11 c (293) b | ND | 171 c (6066) b | ND | 16 c (77) b |
| S1730 | − | + | − | + | 5 | ND | ND | ND | ND | ND | ND | 142 c (2643) b | ND | 142 c (2643) b | ND | (147) b | ND | ND | ND | ND | ND | (274) b | ND | 115 c (5836) b | ND | (52) b |
| S1732 | − | + | − | + | 5 | ND | ND | ND | ND | ND | ND | 201ᶜ (2201) ᵇ | ND | 201 c (2201) b | ND | 12 c (100) b | ND | ND | ND | ND | ND | 10 c (233) b | ND | 149 c (5687) b | ND | (68) b |
| S2605 | + | − | − | + | 1 | ND | ND | ND | ND | ND | ND | 33c (369) b | ND | 33 c (369) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S2606 a | + | − | + | + | 6 | ND | (773) c | 806 b | ND | ND | (90)b | 961b (6248) b | (90)b | 961 b (6248) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S2719 | + | − | − | + | 2 | 77 b (4578) c | (54) c | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S2757 | + | + | + | + | 3 | ND | (69) c | ND | ND | ND | ND | 364 c (5464) b | ND | 364c (5464) b | ND | ND | ND | ND | ND | ND | (220) b | ND | ND | ND | ND | ND |
| S3857 | + | + | + | + | 2 | ND | 52 c (5321) b | ND | (768) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S4497 a | + | − | + | + | 3 | 19 b (917) b | 286 b (12817) b | 631 b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S4634 | + | − | − | + | 1 | ND | ND | ND | ND | ND | ND | (148)b | ND | (148) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| S4738 a | − | + | − | + | 1 | ND | ND | ND | ND | ND | 219 b (2638) b | ND | 219 b (2638) b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| 90-11-287 | − | + | + | + | 3 | ND | ND | ND | ND | ND | ND | 192 c (2162) b | ND | 192 c (2162) b | ND | (46) c | ND | ND | ND | ND | ND | ND | ND | 111 c (3228) b | ND | ND |
| No of occurrence/AHL | 5 | 7 | 3 | 3 | 1 | 11 | 17 | 11 | 17 | 1 | 12 | 1 | 8 | 3 | 1 | 2 | 6 | 2 | 11 | 3 | 9 | |||||
2.3. Structural Abundance of AHLs in Vibrio Strains
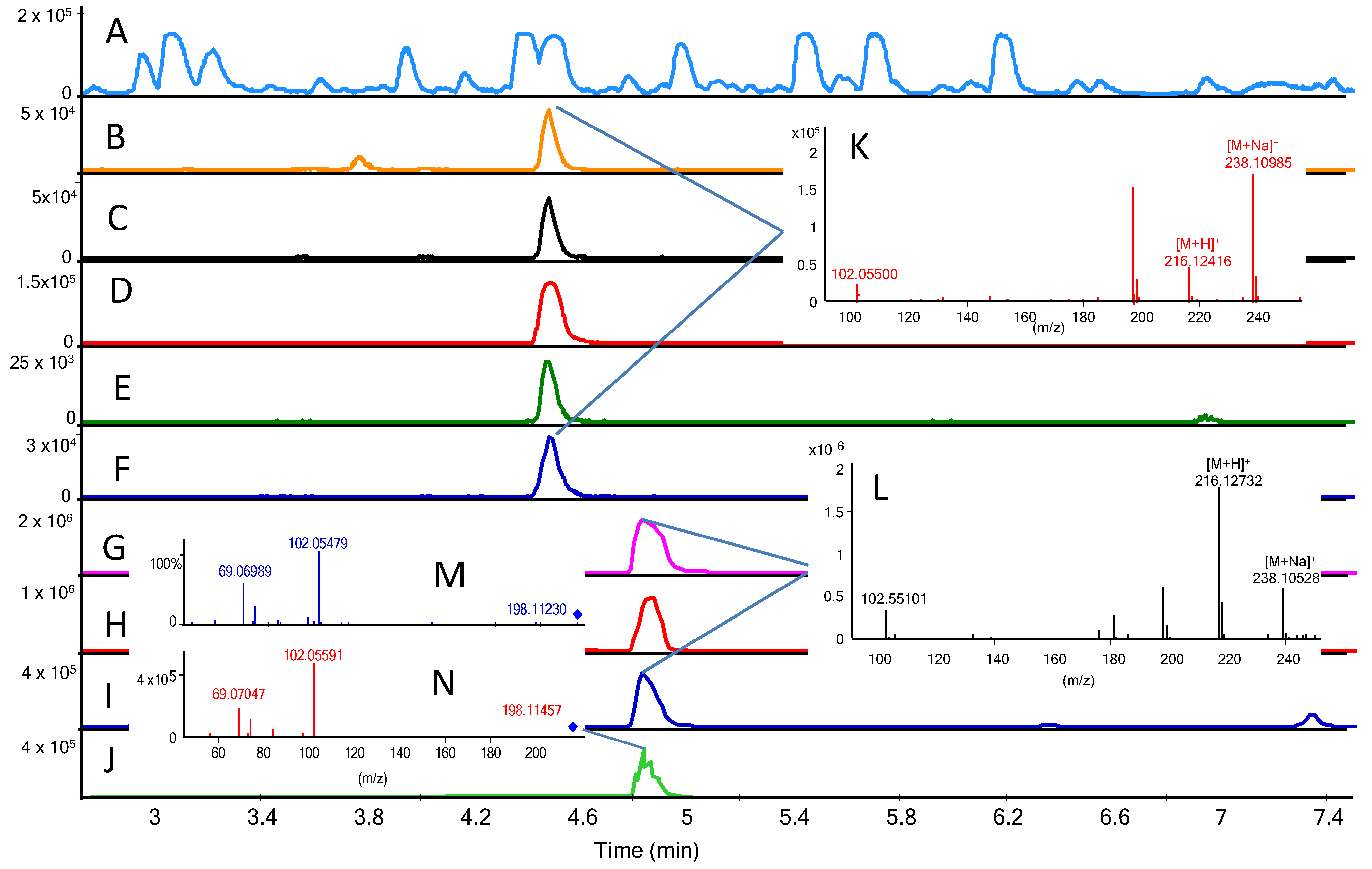
2.4. MS/HRMS Screening of AHLs
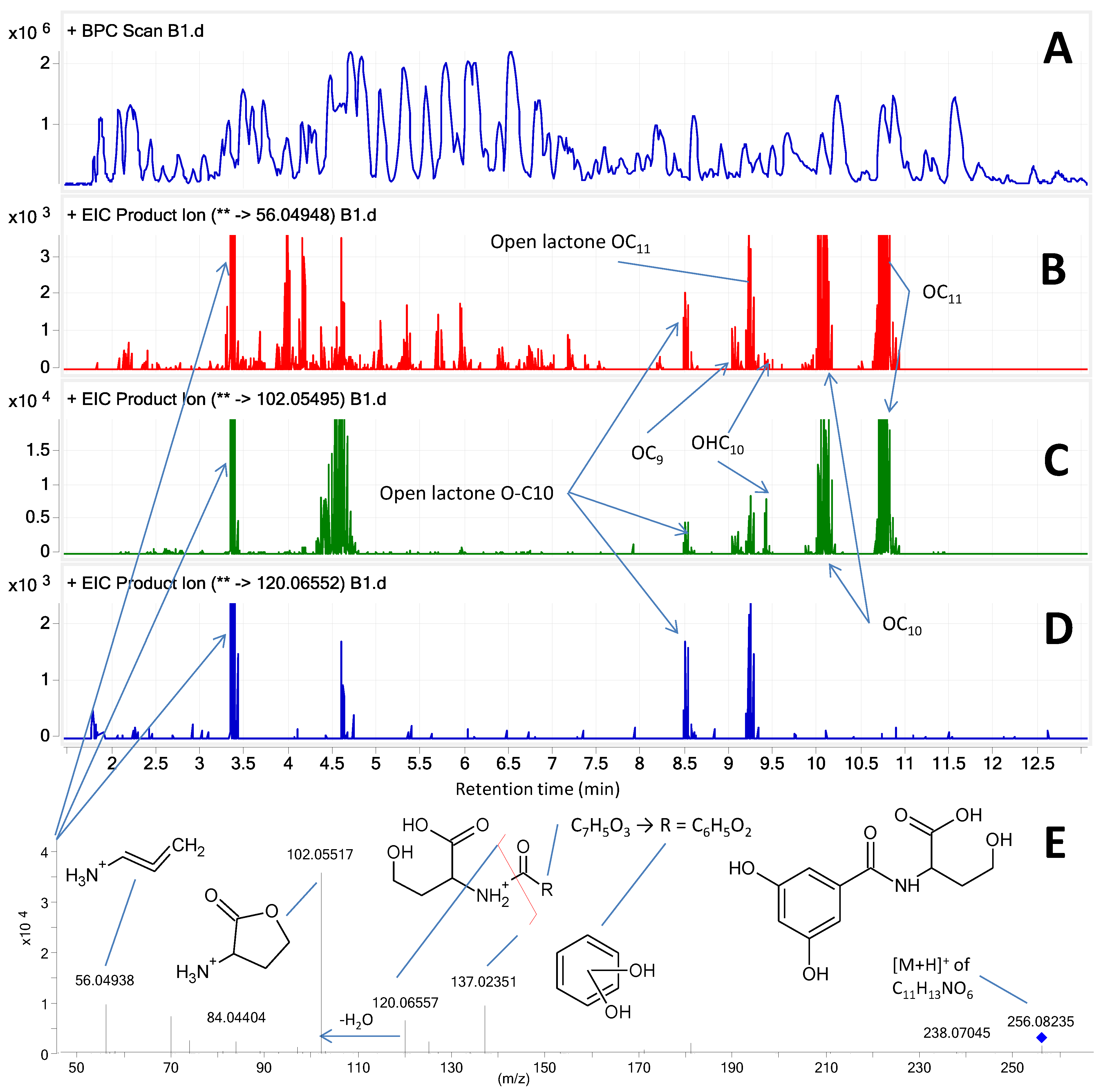
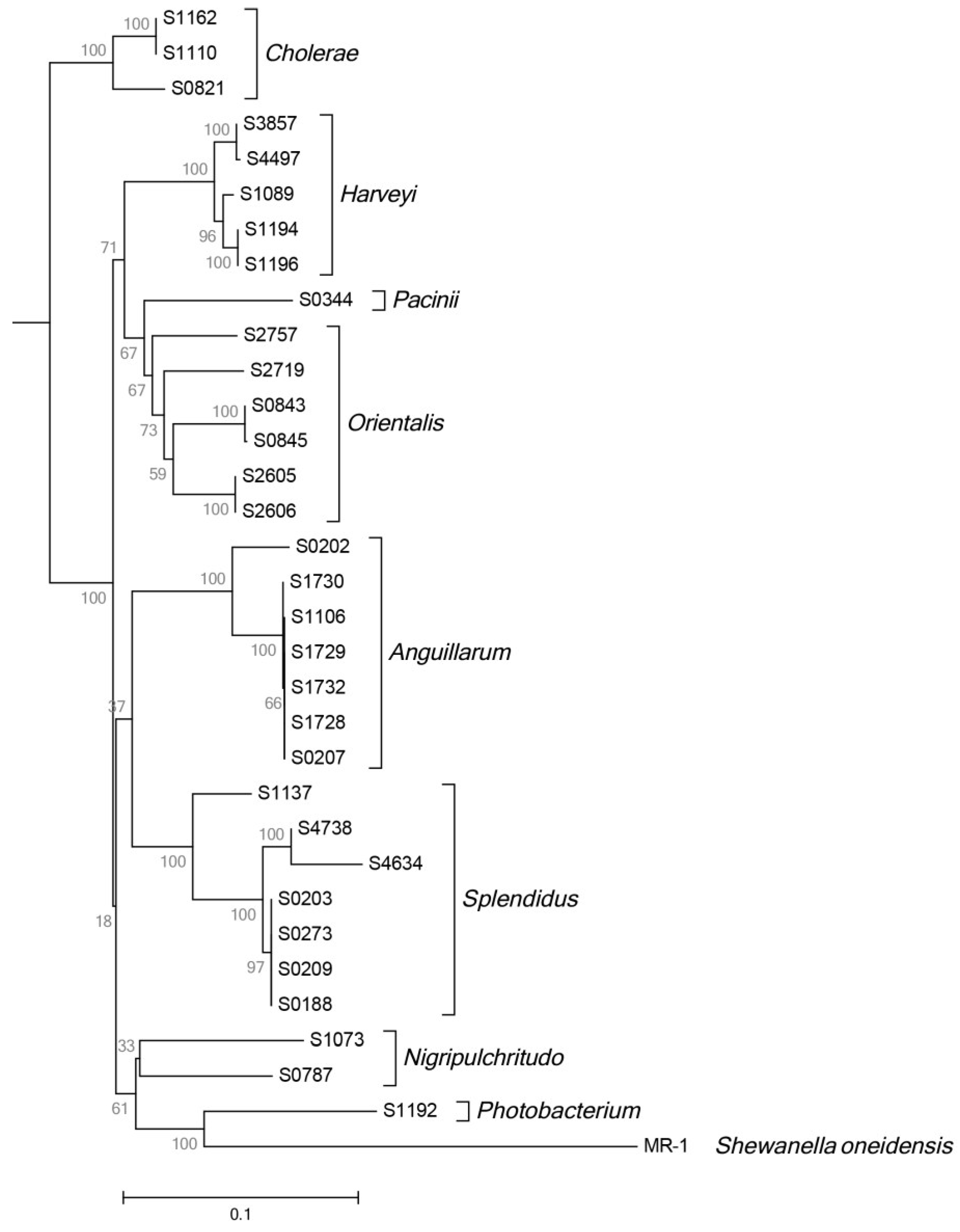
2.5. Phylogeny and Geographical Distribution of AHL-Producing Strains
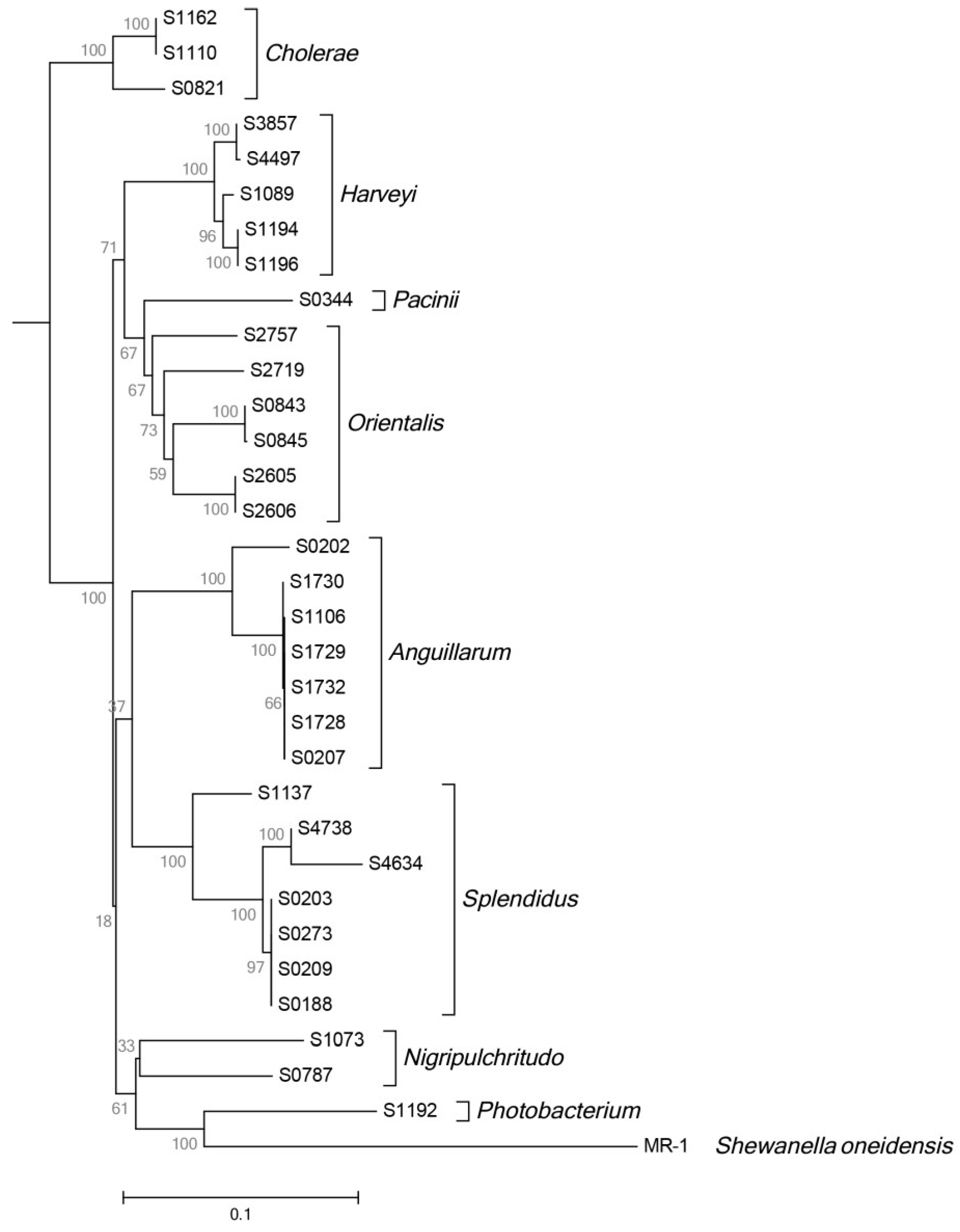
3. Experimental Section
3.1. Bacterial Strains and Growth Conditions
3.2. Phylogenetic Analysis of Strains
3.3. Detection of AHL Compounds
3.4. Extracts for Bioassays and UHPLC-DAD-QTOFMS
3.5. AHL Detection via UHPLC-DAD-QTOFMS
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bandara, H.M.H.N.; Lam, O.L.T.; Jin, L.J.; Samaranayake, L. Microbial chemical signaling: A current perspective. Crit. Rev. Microbiol. 2012, 38, 217–249. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.; Surette, M.G. Communication in bacteria: An ecological and evolutionary perspective. Nat. Rev. Microbiol. 2006, 4, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Bassler, B. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Teplitski, M.; Paul, V. Mini-review: Quorum sensing in the marine environment and its relationship to biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Henke, J.M.; Bassler, B.L. Three parallel quorum-seonsing systems regulate gene expression in Vibrio harveyi. J. Bacteriol. 2004, 186, 6902–6914. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.L. Quorum sensing in vibrios: Complexity for diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Callow, M.E.; Joint, I.; Callow, J.A. Specificity in the settlement—Modifying response of bacterial biofilms towards zoospores of the marine alga Enteromorpha. Environ. Microbiol. 2003, 5, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Takemura, A.F.; Chien, D.M.; Polz, M.F. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 2014, 5, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Han, Y.; Zhang, X.-H. Detection of quorum sensing signal molecules in the family Vibrionaceae. J. Appl. Microbiol. 2011, 110, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H. Autoinduction of bacterial luciferase. Occurrence, mechanism and significance. Arch. Microbiol. 1977, 112, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [PubMed]
- Engebrecht, J.; Silverman, M. Identification of genes and gene products necessary for bacterial bioluminescence. Proc. Natl. Acad. Sci. USA 1984, 81, 4154–4158. [Google Scholar] [CrossRef] [PubMed]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: Isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Mansson, M.; Gram, L.; Larsen, T.O. Production of bioactive secondary metabolites by marine Vibrionaceae. Mar. Drugs 2011, 9, 1440–1468. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Kawabata, K.; Adachi, K.; Katsuta, A.; Shizuri, Y. Unnarmicins A and C, new antibacterial depsipeptides produced by marine bacterium Photobacterium sp. MBIC06485. J. Antibiot. Tokyo 2008, 61, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Sandy, M.; Han, A.; Blunt, J.; Munro, M.; Haygood, M.; Butler, A. Vanchrobactin and anguibactin siderophores produced by Vibrio sp. DS40M4. J. Nat. Prod. 2010, 73, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- De Nys, R.; Kumar, N.; Sharara, K.A.; Srinivasan, S.; Ball, G.; Kjelleberg, S. A new metabolite from the marine bacterium Vibrio angustum S14. J. Nat. Prod. 2001, 64, 531–532. [Google Scholar] [CrossRef] [PubMed]
- Wietz, M.; Mansson, M.; Gotfredsen, C.; Larsen, T.; Gram, L. Antibacterial compounds from marine Vibrionaceae isolated on a global expedition. Mar. Drugs 2010, 8, 2946–2960. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.; Carmeli, S.; Sar, N. Vibrindole A, a metabolite of the marine bacterium, Vibrio parahaemolyticus, isolated from the toxic mucus of the boxfish Ostracion cubicus. J. Nat. Prod. 1994, 57, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Imamura, N.; Nishijima, M.; Takadera, T.; Adachi, K.; Sakai, M.; Sano, H. New anticancer antibiotics pelagiomicins, produced by a new marine bacterium Pelagiobacter variabilis. J. Antibiot. Tokyo 1997, 50, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Kjaerulff, L.; Nielsen, A.; Mansson, M.; Gram, L.; Larsen, T.O.; Ingmer, H.; Gotfredsen, C.H. Identification of four new agr quorum sensing-interfering cyclodepsipeptides from a marine Photobacterium. Mar. Drugs 2013, 11, 5051–5062. [Google Scholar] [CrossRef] [PubMed]
- Mansson, M.; Nielsen, A.; Kjærulff, L.; Gotfredsen, C.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T. Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a marine Photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Melchiorsen, J.; Bruhn, J. Antibacterial activity of marine culturable bacteria collected from a global sampling of ocean surface waters and surface swabs of marine organisms. Mar. Biotechnol. N. Y. 2010, 12, 439–451. [Google Scholar] [CrossRef]
- Bassler, B.L.; Greenberg, E.P.; Stevens, A.M. Cross-species induction of luminescence in the quorum-sensing bacterium Vibrio harveyi. J. Bacteriol. 1997, 179, 4043–4045. [Google Scholar] [PubMed]
- Zhu, J.; Miller, M.B.; Vance, R.E.; Dziejman, M.; Bassler, B.L.; Mekalanos, J.J. Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2002, 99, 3129–3134. [Google Scholar] [CrossRef] [PubMed]
- Buchholtz, C.; Nielsen, K.F.; Milton, D.L.; Larsen, J.L.; Gram, L. Profiling of acylated homoserine lactones of Vibrio anguillarum in vitro and in vivo: Influence of growth conditions and serotype. Syst. Appl. Microbiol. 2006, 29, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.L.; Hardman, A.; Camara, M.; Chhabra, S.R.; Bycroft, B.W.; Stewart, G.S.; Williams, P. Quorum sensing in Vibrio anguillarum: Characterization of the vanI/vanR locus and identification of the autoinducer N-(3-oxodecanoyl)-l-homoserine lactone. J. Bacteriol. 1997, 179, 3004–3012. [Google Scholar] [PubMed]
- Bruhn, J.B.; Dalsgaard, I.; Nielsen, K.F.; Buchholtz, C.; Larsen, J.L.; Gram, L. Quorum sensing signal molecules (acylated homoserine lactones) in gram-negative fish pathogenic bacteria. Dis. Aquat. Organ. 2005, 65, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Valiente, E.; Bruhn, J.B.; Nielsen, K.F.; Larsen, J.L.; Roig, F.J.; Gram, L.; Amaro, C. Vibrio vulnificus produces quorum sensing signals of the AHL-class. FEMS Microbiol. Ecol. 2009, 69, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Flodgaard, L.R.; Dalgaard, P.; Andersen, J.B.; Nielsen, K.F.; Givskov, M.; Gram, L. Nonbioluminescent strains of Photobacterium phosphoreum produce the cell-to-cell communication signal N-(3-Hydroxyoctanoyl) homoserine lactone. Appl. Environ. Microbiol. 2005, 71, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.M.; Schaefer, A.L.; Ferreira, R.B.R.; Qin, N.; Stevens, A.M.; Ruby, E.G.; Greenberg, E.P. Transcriptome analysis of the Vibrio fischeri LuxR-LuxI regulon. J. Bacteriol. 2007, 189, 8387–8391. [Google Scholar] [CrossRef] [PubMed]
- García-Aljaro, C.; Vargas-Cespedes, G.J.; Blanch, A.R. Detection of acylated homoserine lactones produced by Vibrio spp. and related species isolated from water and aquatic organisms. J. Appl. Microbiol. 2012, 112, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Purohit, A.A.; Johansen, J.A.; Hansen, H.; Leiros, H.-K.S.; Kashulin, A.; Karlsen, C.; Smalås, A.; Haugen, P.; Willassen, N.P. Presence of acyl-homoserine lactones in 57 members of the Vibrionaceae family. J. Appl. Microbiol. 2013, 115, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Mol. Plant Microbe Interact. 1998, 11, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; John, H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acyl homoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Kildgaard, S.; Mansson, M.; Dosen, I.; Klitgaard, A.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F. Accurate dereplication of bioactive secondary metabolites from marine-derived fungi by UHPLC-DAD-QTOFMS and a MS/HRMS library. Mar. Drugs 2014, 12, 3681–3705. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.; Chhabra, S.; Nys, R.; Stead, P.; Bainton, N.; Hill, P.; Manefield, M.; Kumar, N.; Labatte, M.; Engl, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S. Quorum sensing and bacterial biofilms. Nat. Prod. Rep. 2010, 27, 343–369. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.A.; Herman, J.; Krank, J.; Robert, C.; Churchill, M.E.A.; Murphy, R.C. Specificity of acyl-homoserine lactone synthases examined by mass spectrometry specificity of acyl-homoserine lactone synthases examined by mass spectrometry. J. Bacteriol. 2006, 188, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Kastbjerg, V.G.; Nielsen, K.F.; Dalsgaard, I.; Rasch, M.; Bruhn, J.B.; Givskov, M.; Gram, L. Profiling acylated homoserine lactones in Yersinia ruckeri and influence of exogenous acyl homoserine lactones and known quorum-sensing inhibitors on protease production. J. Appl. Microbiol. 2007, 102, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Inaba, T.; Kato, N.; Kanai, K.; Ikeda, T. Identification of Quorum-Sensing Signal Molecules and the LuxRI Homologs in Fish Pathogen Edwardsiella tarda. J. Biosci. Bioeng. 2004, 98, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.H.; Arnold, F.H.; Leadbetter, J.R. Directed evolution of Vibrio fischeri LuxR for increased sensitivity to a broad spectrum of acyl-homoserine lactones. Mol. Microbiol. 2005, 55, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.N.; Myers, T.G.; O’Connor, P.M.; Friend, S.H.; Fornace, A.J.; Kohn, K.W.; Fojo, T.; Bates, S.E.; Rubinstein, L.V.; Anderson, N.L.; et al. An information-intensive approach to the molecular pharmacology of cancer. Science 1997, 275, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Machado, H.; Gram, L. The fur gene as a phylogenetic marker in Vibrio species identification. 2014; submitted. [Google Scholar]
- Sawabe, T.; Kita-Tsukamoto, K.; Thompson, F.L. Inferring the evolutionary history of vibrios by means of multilocus sequence analysis. J. Bacteriol. 2007, 189, 7932–7936. [Google Scholar] [CrossRef] [PubMed]
- Thompson, F.L.; Gevers, D.; Thompson, C.C.; Dawyndt, P.; Naser, S.; Hoste, B.; Munn, C.B.; Swings, J. Phylogeny and molecular identification of vibrios on the basis of multilocus sequence analysis. Appl. Environ. Microbiol. 2005, 71, 5107–5115. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Macián, M.C.; Arahal, D.R.; Garay, E.; Pujalte, M.J. Multilocus sequence analysis of the central clade of the genus Vibrio by using the 16S rRNA, recA, pyrH, rpoD, gyrB, rctB and toxR genes. Int. J. Syst. Evol. Microbiol. 2010, 60, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, M.W.; Matsui, G.Y.; Friedman, R.; Lovell, C.R. Optimization of Multilocus Sequence Analysis for Identification of Species in the Genus Vibrio. Appl. Environ. Microbiol. 2014, 80, 5359–5365. [Google Scholar] [CrossRef] [PubMed]
- Skov, M.N.; Pedersen, K.; Larsen, J.L. Comparison of Pulsed-Field Gel Electrophoresis, Ribotyping, and Plasmid Profiling for Typing of Vibrio anguillarum Serovar O1. Appl. Environ. Microbiol. 1995, 61, 1540–1545. [Google Scholar] [PubMed]
- Gibson, L.F.; Khoury, J.T. Storage and survival of bacteria by ultra-freeze. Lett. Appl. Microbiol. 1986, 3, 127–129. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Ravn, L.; Christensen, A.B.; Molin, S.; Givskov, M.; Gram, L. Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinetics. J. Microbiol. Methods 2001, 44, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Grossart, H.; Schlingloff, A.; Kiørboe, T. Possible Quorum Sensing in Marine Snow Bacteria: Production of Acylated Homoserine Lactones by Roseobacter Strains Isolated from Marine Snow. Appl. Environ. Microbiol. 2002, 68, 4111–4116. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasmussen, B.B.; Nielsen, K.F.; Machado, H.; Melchiorsen, J.; Gram, L.; Sonnenschein, E.C. Global and Phylogenetic Distribution of Quorum Sensing Signals, Acyl Homoserine Lactones, in the Family of Vibrionaceae. Mar. Drugs 2014, 12, 5527-5546. https://doi.org/10.3390/md12115527
Rasmussen BB, Nielsen KF, Machado H, Melchiorsen J, Gram L, Sonnenschein EC. Global and Phylogenetic Distribution of Quorum Sensing Signals, Acyl Homoserine Lactones, in the Family of Vibrionaceae. Marine Drugs. 2014; 12(11):5527-5546. https://doi.org/10.3390/md12115527
Chicago/Turabian StyleRasmussen, Bastian Barker, Kristian Fog Nielsen, Henrique Machado, Jette Melchiorsen, Lone Gram, and Eva C. Sonnenschein. 2014. "Global and Phylogenetic Distribution of Quorum Sensing Signals, Acyl Homoserine Lactones, in the Family of Vibrionaceae" Marine Drugs 12, no. 11: 5527-5546. https://doi.org/10.3390/md12115527
APA StyleRasmussen, B. B., Nielsen, K. F., Machado, H., Melchiorsen, J., Gram, L., & Sonnenschein, E. C. (2014). Global and Phylogenetic Distribution of Quorum Sensing Signals, Acyl Homoserine Lactones, in the Family of Vibrionaceae. Marine Drugs, 12(11), 5527-5546. https://doi.org/10.3390/md12115527