Inhibition of Bacterial Quorum Sensing by Extracts from Aquatic Fungi: First Report from Marine Endophytes


 ,
, 

Abstract
:1. Introduction
2. Results
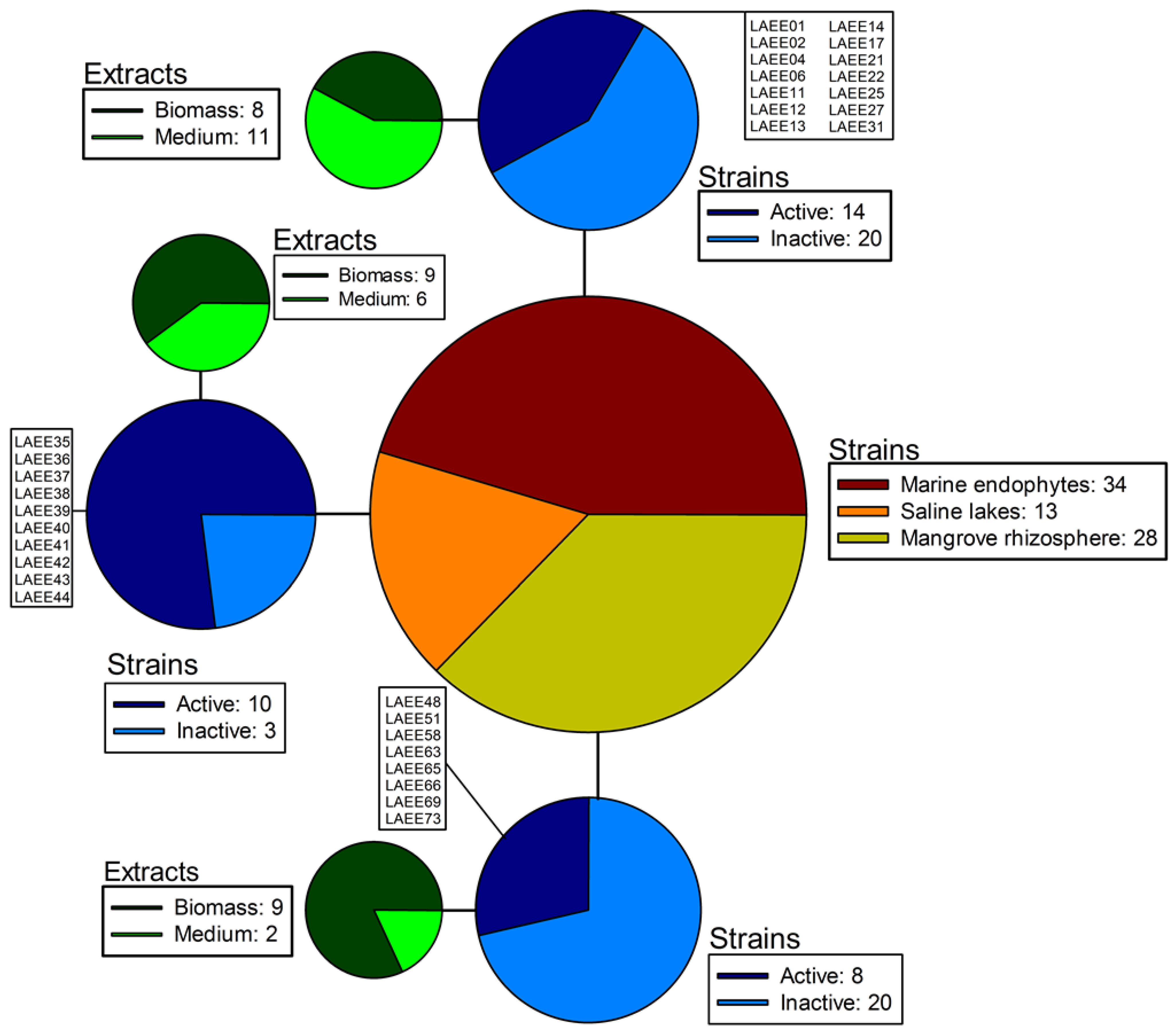
2.1. Sampling and Isolation

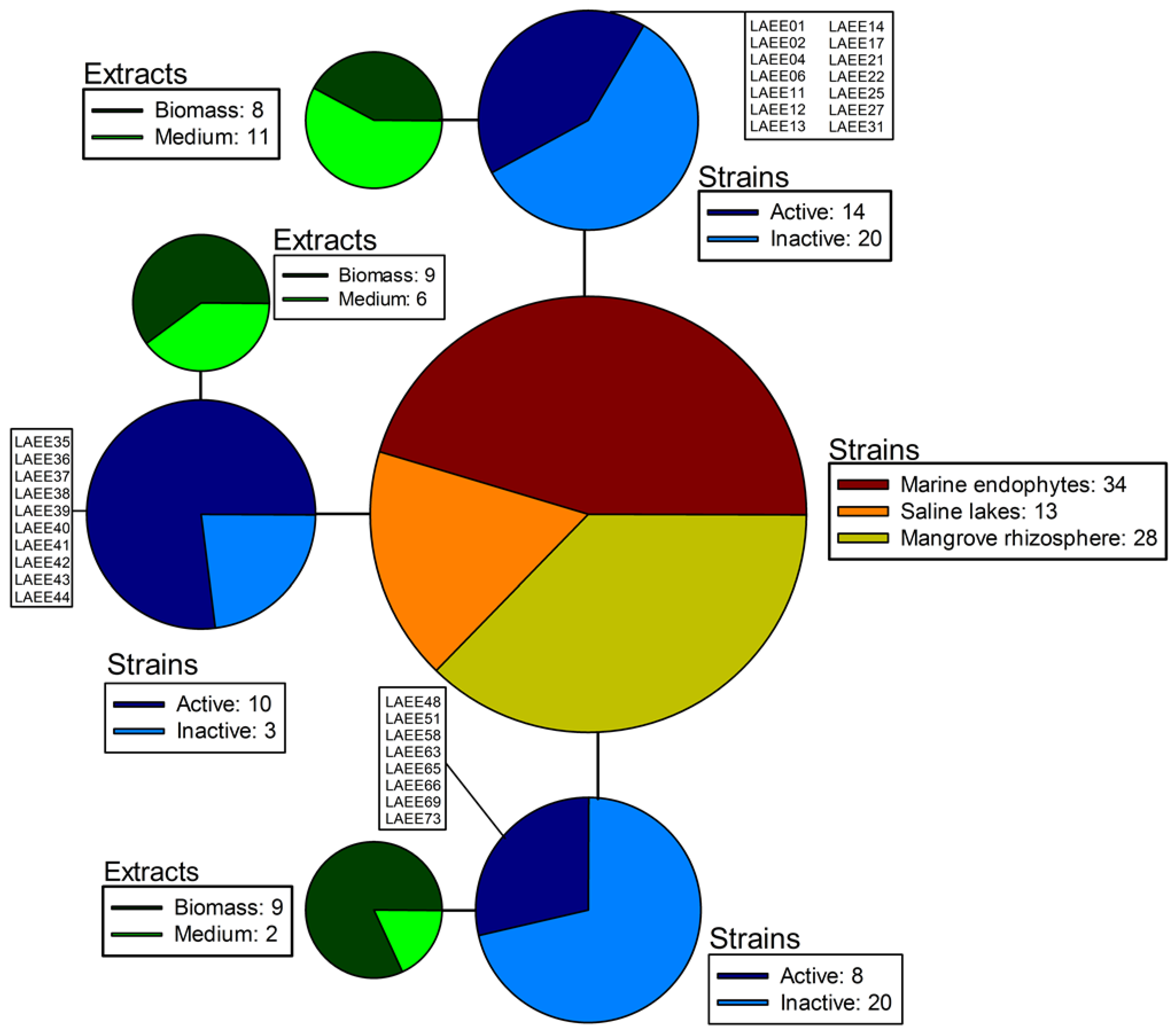{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID. | Fungus Genus | Biological Source | Location |
|---|---|---|---|
| LAEE 01 | Cladosporium sp. | Amphimedon compressa | Arrecifes Blancas |
| LAEE 02 | Aspergillus sp. | Amphimedon compressa | Arrecifes Blancas |
| LAEE 03 | Curvularia sp. | Amphimedon compressa | Arrecifes Blancas |
| LAEE 04 | Fusarium sp. | Agelas sp. | Arrecifes Blancas |
| LAEE 05 | Aspergillus sp. | Agelas sp. | Arrecifes Blancas |
| LAEE 06 | Sarocladium sp. | Agelas sp. | Arrecifes Blancas |
| LAEE 07 | Fusarium sp. | Aplysina sp. | Arrecifes Blancas |
| LAEE 08 | Acremonium sp. | Chondrilla sp. | Arrecifes Blancas |
| LAEE 09 | Fusarium sp. | Acropora palmata | Arrecifes Blancas |
| LAEE 10 | Acremonium sp. | Acropora palmata | Arrecifes Blancas |
| LAEE 11 | Acremonium sp. | Acropora palmata | Arrecifes Blancas |
| LAEE 12 | Fusarium sp. | Diploria clivosa | Arrecifes Blancas |
| LAEE 13 | Fusarium sp. | Diploria strigosa | Arrecifes Blancas |
| LAEE 14 | Epicoccum sp. | Diploria strigosa | Arrecifes Blancas |
| LAEE 15 | Trichoderma sp. | Diploria strigosa | Arrecifes Blancas |
| LAEE 16 | Aspergillus sp. | Diploria strigosa | Arrecifes Blancas |
| LAEE 17 | Fusarium sp. | Diploria strigosa | Arrecifes Blancas |
| LAEE 18 | Aspergillus sp. | Montastrea cavernosa | Arrecifes Blancas |
| LAEE 19 | Aspergillus sp. | Montastrea cavernosa | Arrecifes Blancas |
| LAEE 20 | Aspergillus sp. | Montastrea cavernosa | Arrecifes Blancas |
| LAEE 21 | Khuskia sp. | Plexaura flexuosa | Arrecifes Blancas |
| LAEE 22 | Cladosporium sp. | Plexaura flexuosa | Arrecifes Blancas |
| LAEE 23 | Trichoderma sp. | Plexaura flexuosa | Arrecifes Blancas |
| LAEE 24 | Monilia sp. | Plexaura flexuosa | Arrecifes Blancas |
| LAEE 25 | Acremonium sp. | Pseudoplexaura porosa | Arrecifes Blancas |
| LAEE 26 | Acremonium sp. | Pseudoterogorgia americana | Isla de Sacrificios |
| LAEE 27 | Aspergillus sp. | Pseudoterogorgia americana | Isla de Sacrificios |
| LAEE 28 | Alternaria sp. | Siderastrea siderea | Arrecifes Blancas |
| LAEE 29 | Aspergillus sp. | Siderastrea siderea | Arrecifes Blancas |
| LAEE 30 | Aspergillus sp. | Siderastrea siderea | Arrecifes Blancas |
| LAEE 31 | Fusarium sp. | Siderastrea siderea | Arrecifes Blancas |
| LAEE 32 | Aspergillus sp. | Siderastrea siderea | Arrecifes Blancas |
| LAEE 33 | Fusarium sp. | Zoanthus sp. | Isla de Sacrificios |
| LAEE 34 | Aspergillus sp. | Hypnea cervicornis | Isla de Sacrificios |
| LAEE 35 | Monilia sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 36 | Ulocladium sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 37 | Stachybotrys sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 38 | Fusarium sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 39 | Fusarium sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 40 | Absidia sp. | Potamogeton sp. | Laguna de Atexcac |
| LAEE 41 | Paecilomyces sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 42 | Chaetomium sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 43 | Stemphylium sp. | Juncus sp. | Laguna de Atexcac |
| LAEE 44 | Penicillium sp. | Cladophora sp. | Laguna de Atexcac |
| LAEE 45 | Acremonium sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 46 | Aspergillus sp. | Sample from littoral zone | Laguna de Atexcac |
| LAEE 47 | Aspergillus sp. | Cladophora sp. | Laguna de Atexcac |
| LAEE 48 | Fusarium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 49 | Alternaria sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 50 | Fusarium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 51 | Trichocladium sp. | Rhizosphereof Rhizophora mangle | Manglar de Tuxpan |
| LAEE 52 | Aspergillus sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 53 | Penicillium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 54 | Aspergillus sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 55 | Fusarium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 56 | Aspergillus sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 57 | Mucor sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 58 | Cladosporium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 59 | Fusarium sp. | Rhizosphere of Rhizophora mangle | Manglar de Tuxpan |
| LAEE 60 | Aspergillus sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 61 | Blastomyces sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 62 | Aspergillus sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 63 | Fusarium sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 64 | Fusarium sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 65 | Penicillium sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 66 | Acremonium sp. | Rhizosphere of Avicennia germinans | Manglar de Tuxpan |
| LAEE 67 | Aspergillus sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 68 | Aspergillus sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 69 | Penicillium sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 70 | Acremonium sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 71 | Aspergillus sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 72 | Aspergillus sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 73 | Fusarium sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 74 | Paecilomyces sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
| LAEE 75 | Paecilomyces sp. | Rhizosphere of Laguncularia racemosa | Manglar de Tuxpan |
2.2. Quorum Sensing Assays
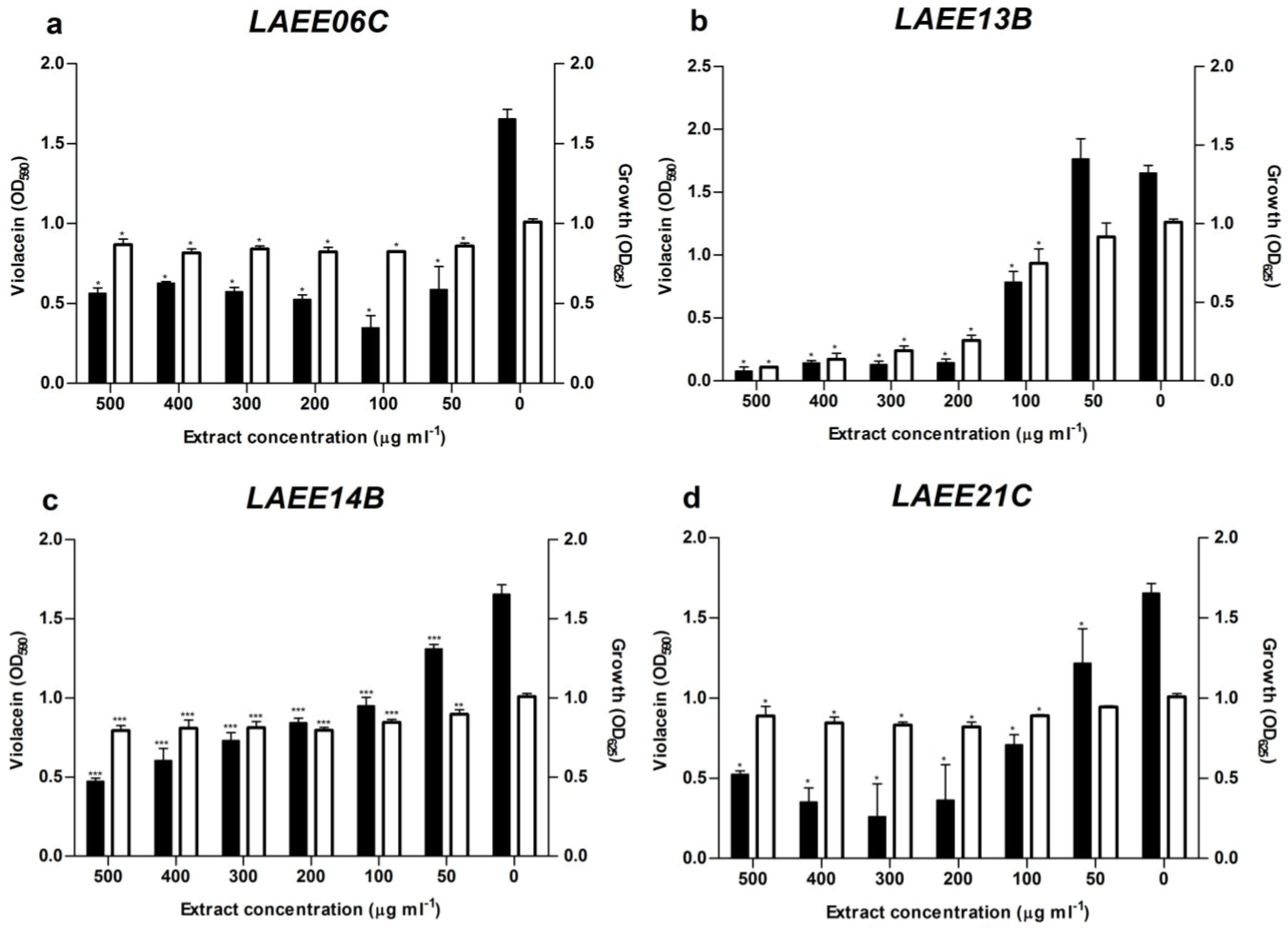
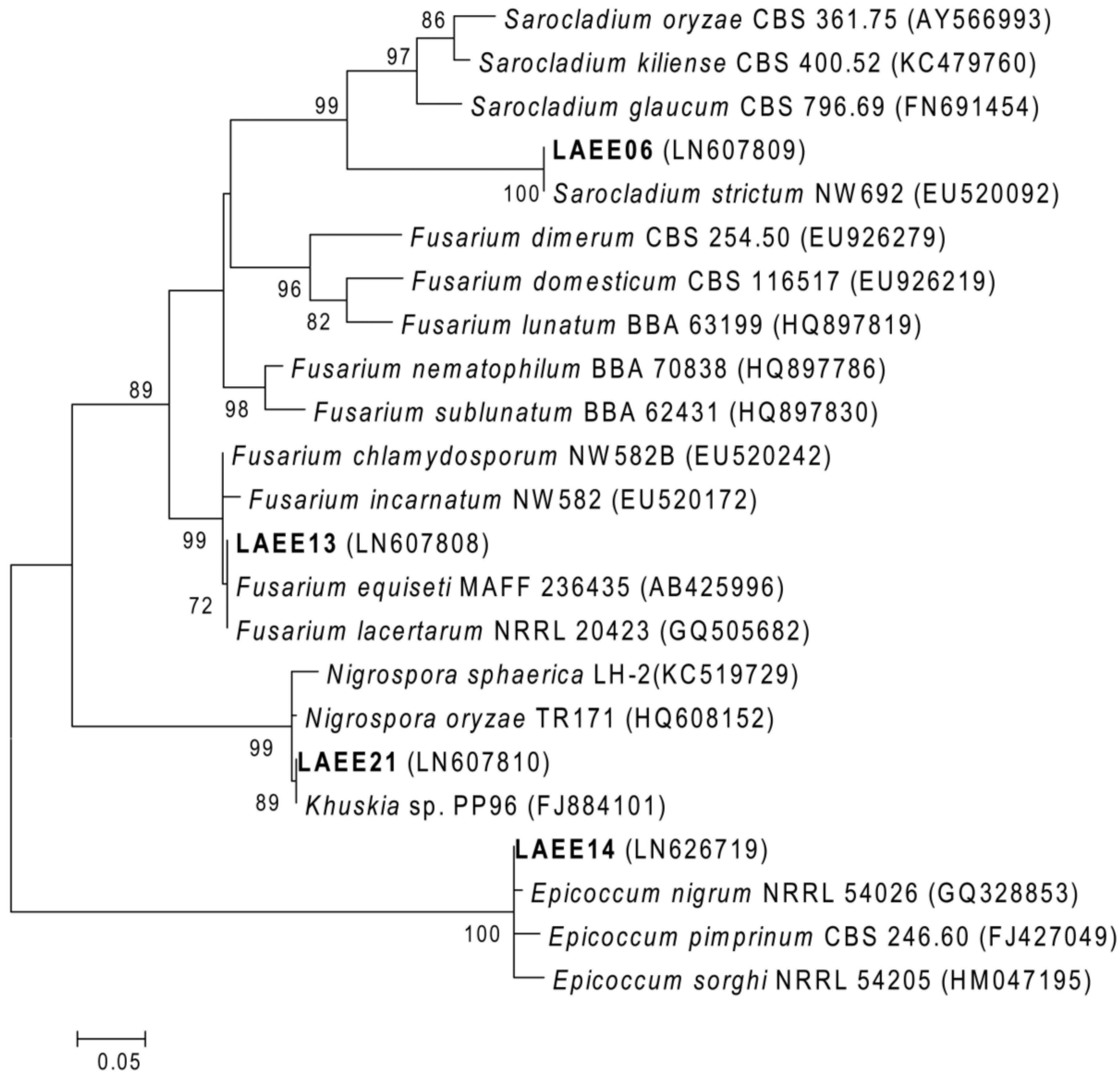
2.3. ITS Phylogeny and Taxonomic Identification of the Isolates
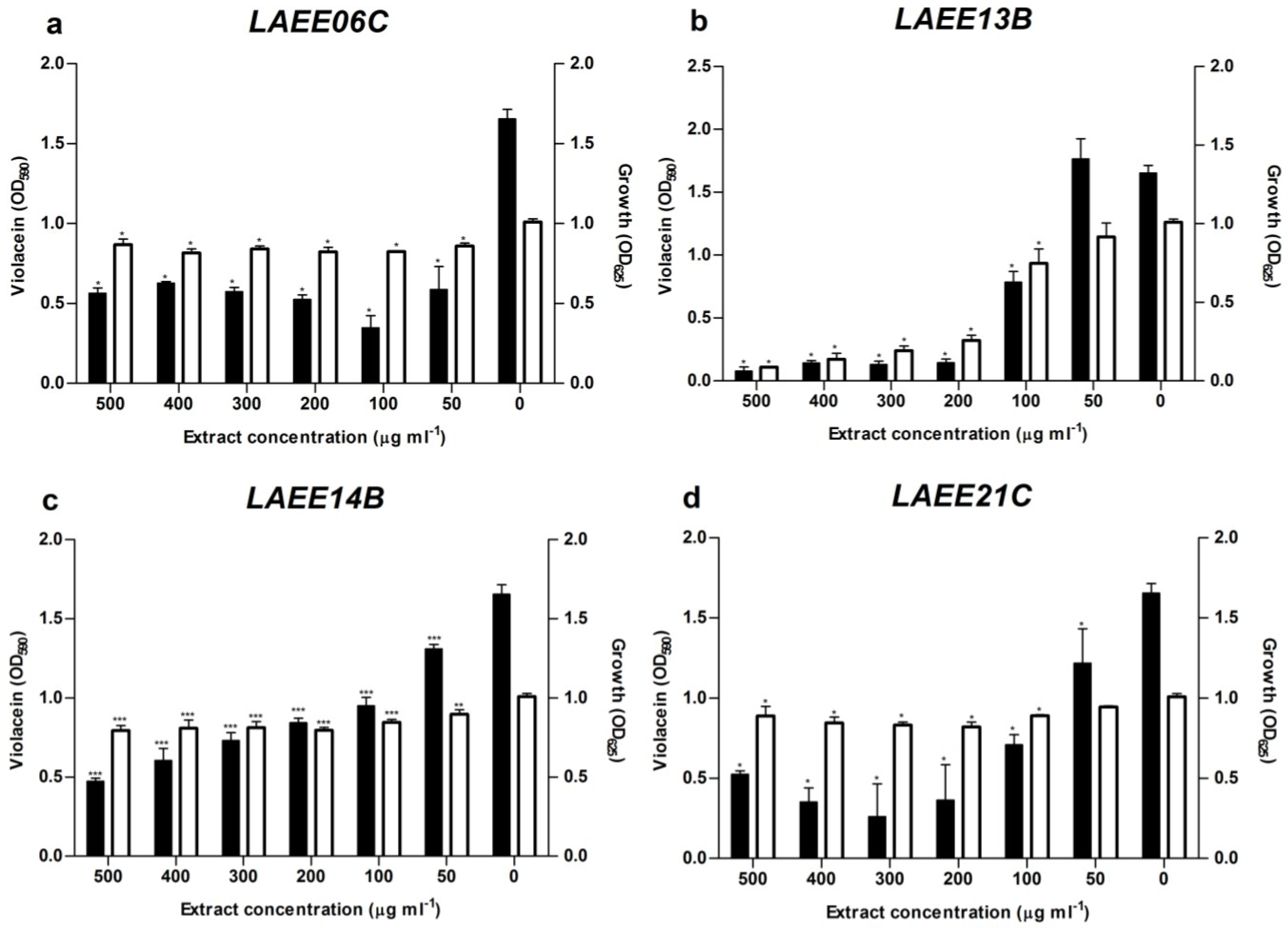
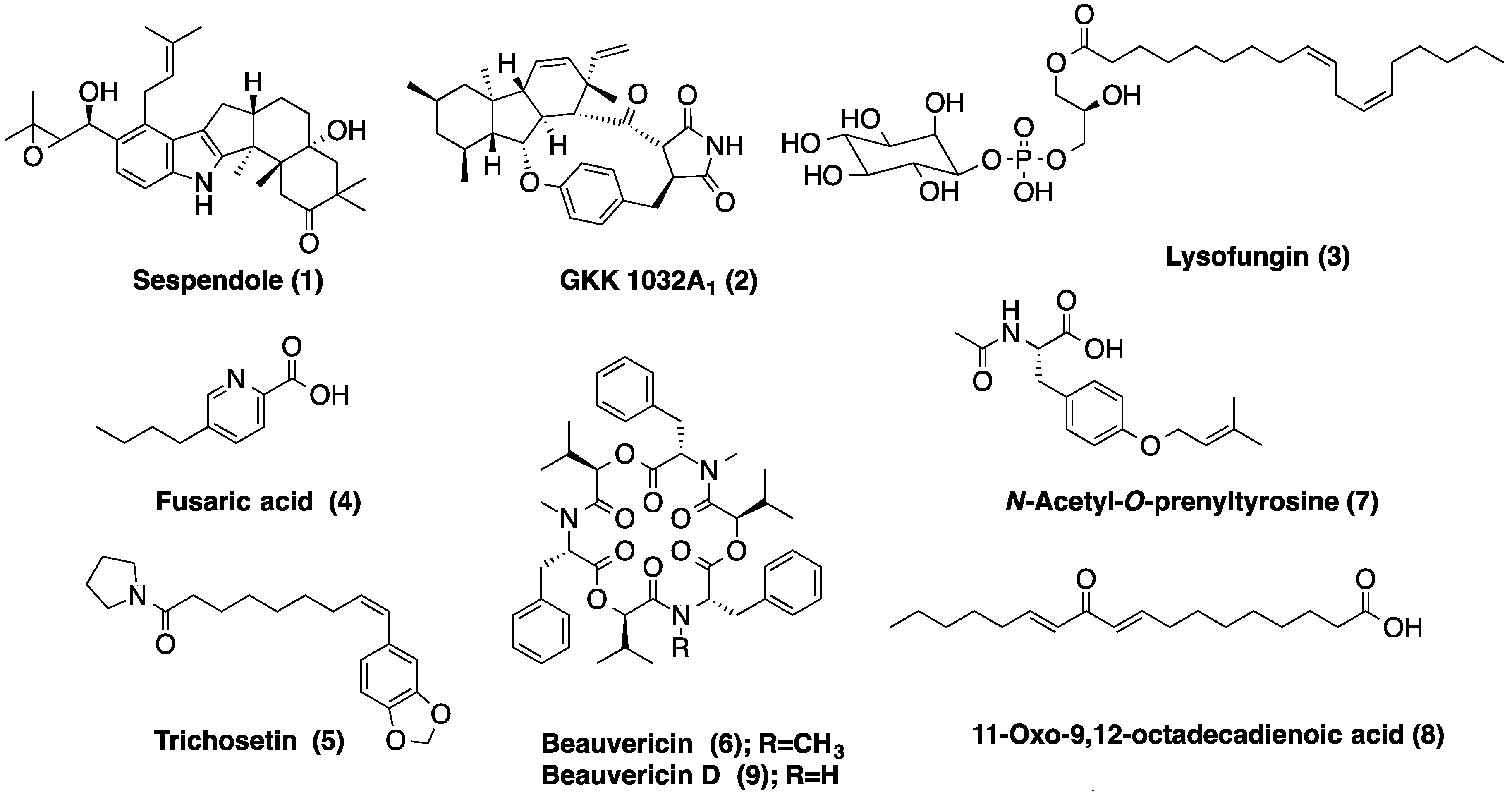
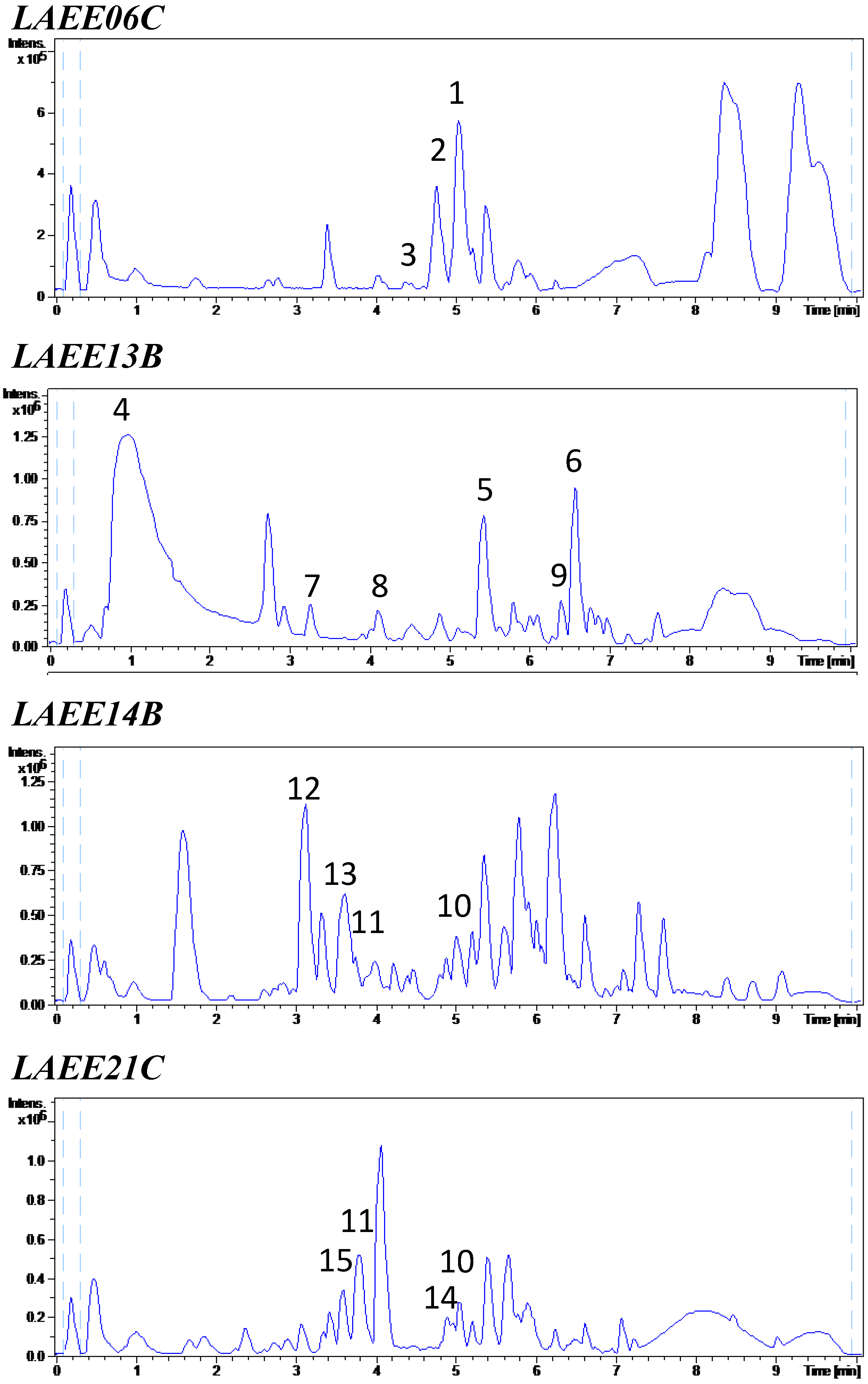
2.4. Metabolic Profiles
| Sample | RT (min) | Suggested MF | FM Database ID | Fungal Metabolites with This MF Included in the DNP |
|---|---|---|---|---|
| LAEE06C | 4.36 | C27H49O12P | Lysofungin (3) | |
| 4.76 | C33H43NO4 | Antibiotic GKK 1032A1 (2) | ||
| 5.04 | C33H45NO4 | Sespendole (1) | ||
| 5.39 | C18H30O2 | Linolenic acid | ||
| 6.64 | C18H35NO2 | Not found in the DNP | ||
| LAEE13B | 0.90 | C10H13NO2 | Fusaric acid (4) | |
| 2.74 | C9H13NO | Not found in the DNP | ||
| 3.26 | C16H21NO4 | N-Acetyl-O-prenyltyrosine (7) | ||
| 4.02 | C13H16O3 | 5 coincidences in the DNP | ||
| 4.11 | C18H30O3 | 11-Oxo-9,12-octadecadienoic acid (8) | ||
| 5.41 | C21H29NO4 | Trichosetin (5) | ||
| 6.39 | C44H55N3O9 | Beauvericin D (9) | ||
| 6.56 | C45H57N3O9 | Beauvericin (6) | ||
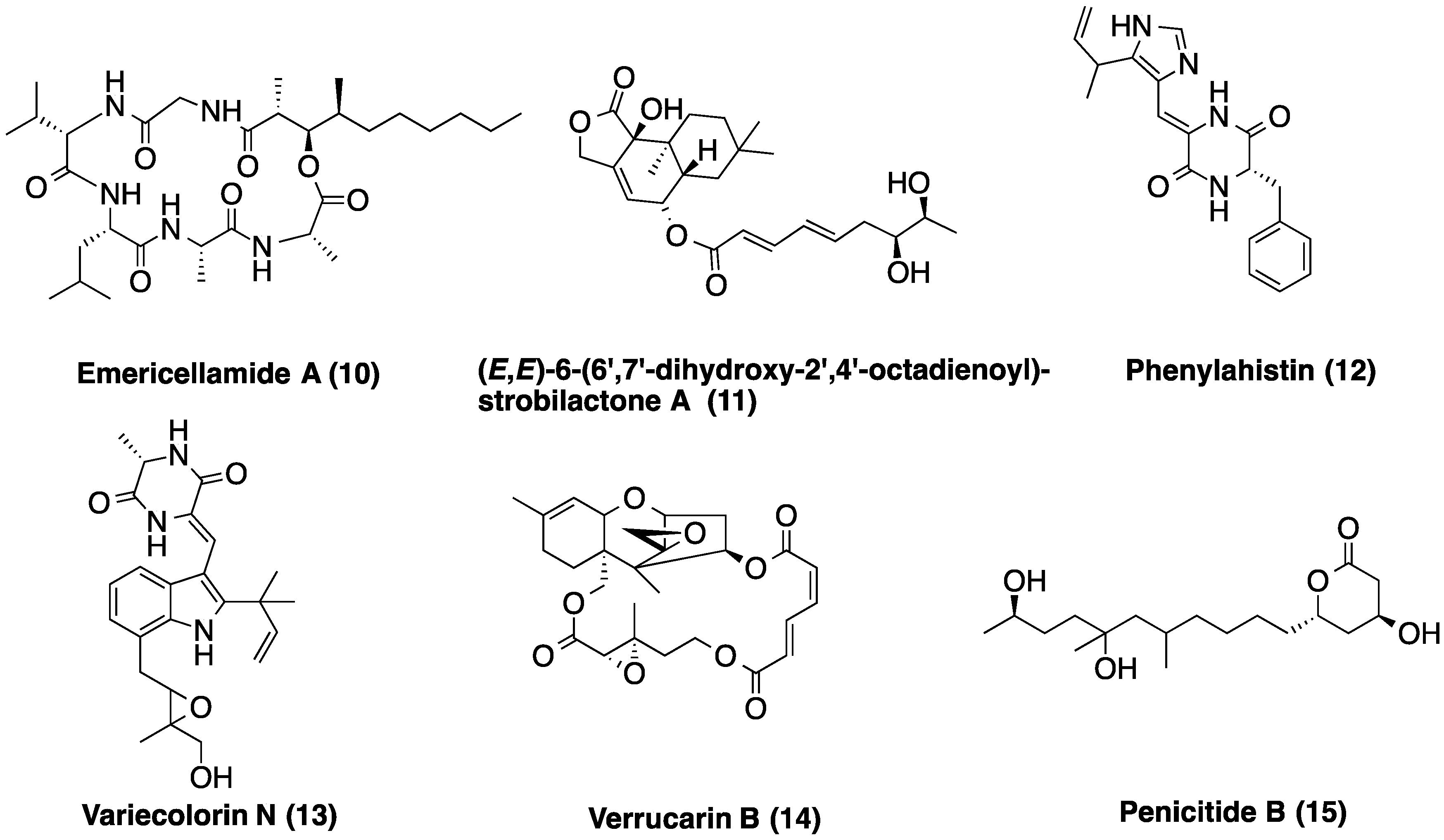
| LAEE14B | 3.29 | C20H22N4O2 | Phenylahistin (12) | |
| 3.48 | C24H29N3O4 | Variecolorin N (13) | ||
| 3.76 | C23H32O7 | (E,E)-6-(6′,7′-dihydroxy-2′,4′-octadienoyl)-strobilactone A | ||
| 5.00 | C31H55N5O7 | Emericellamide A (10) | ||
| 5.74 | C15H20O3 | >50 coincidences in the DNP | ||
| 6.15 | C25H34O | Not found in the DNP | ||
| 6.53 | C18H32O2 | Linoleic acid | ||
| LAEE21C | 2.38 | C10H16O4 | 13 coincidences in the DNP | |
| 3.08 | C15H22O4 | >50 coincidences in the DNP | ||
| 3.59 | C18H34O5 | Penicitide B (15) | ||
| 3.79 | C23H32O7 | (E,E)-6-(6′,7′-dihydroxy-2′,4′-octadienoyl)-strobilactone A | ||
| 4.06 | C17H14N2S | Not found in the DNP | ||
| 4.58 | C27H32O9 | Verrucarin B (14) | ||
| 5.04 | C31H55N5O7 | Emericellamide A (10) | ||
| 6.61 | C18H32O2 | Linoleic acid |
3. Discussion
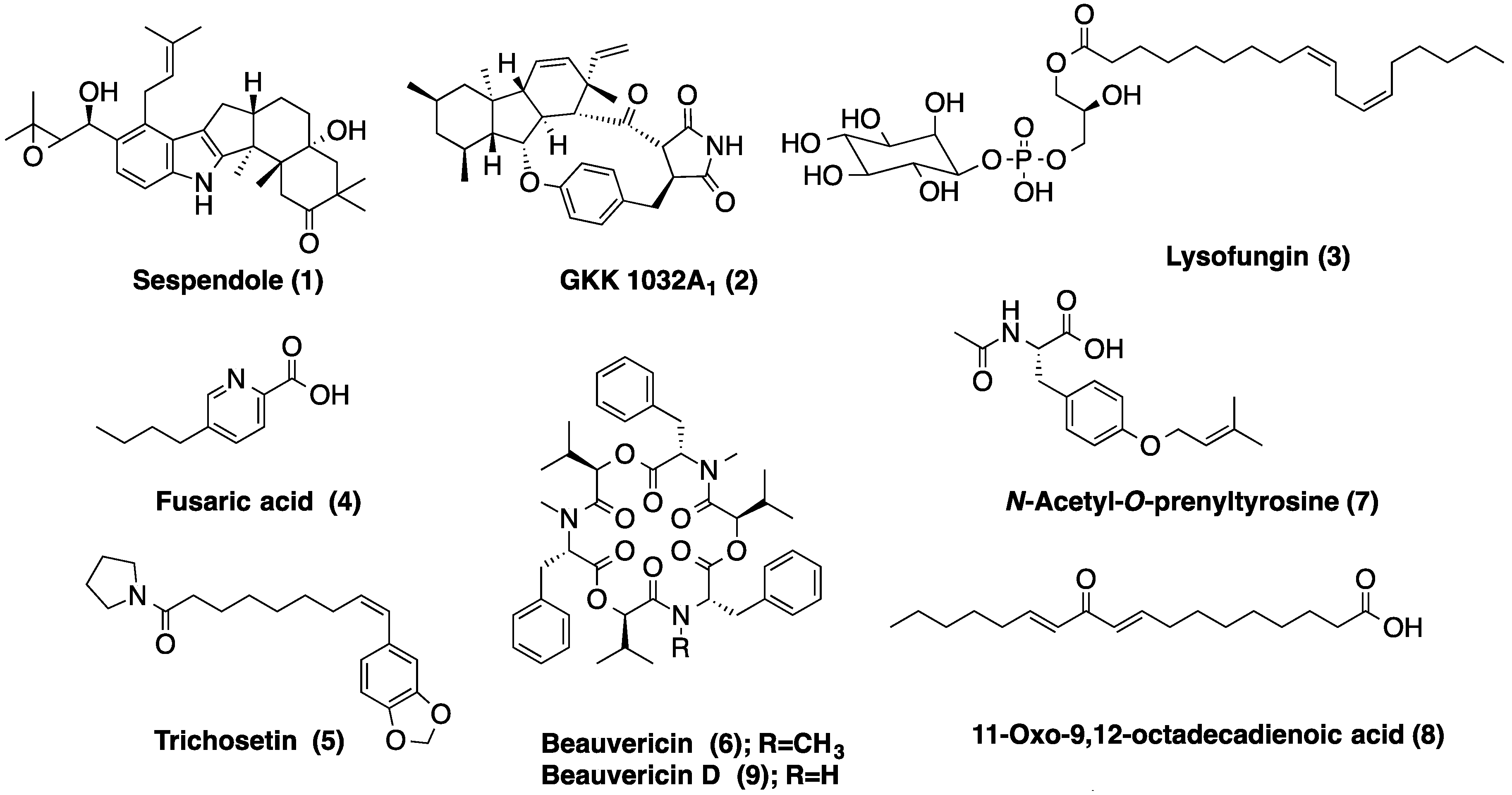
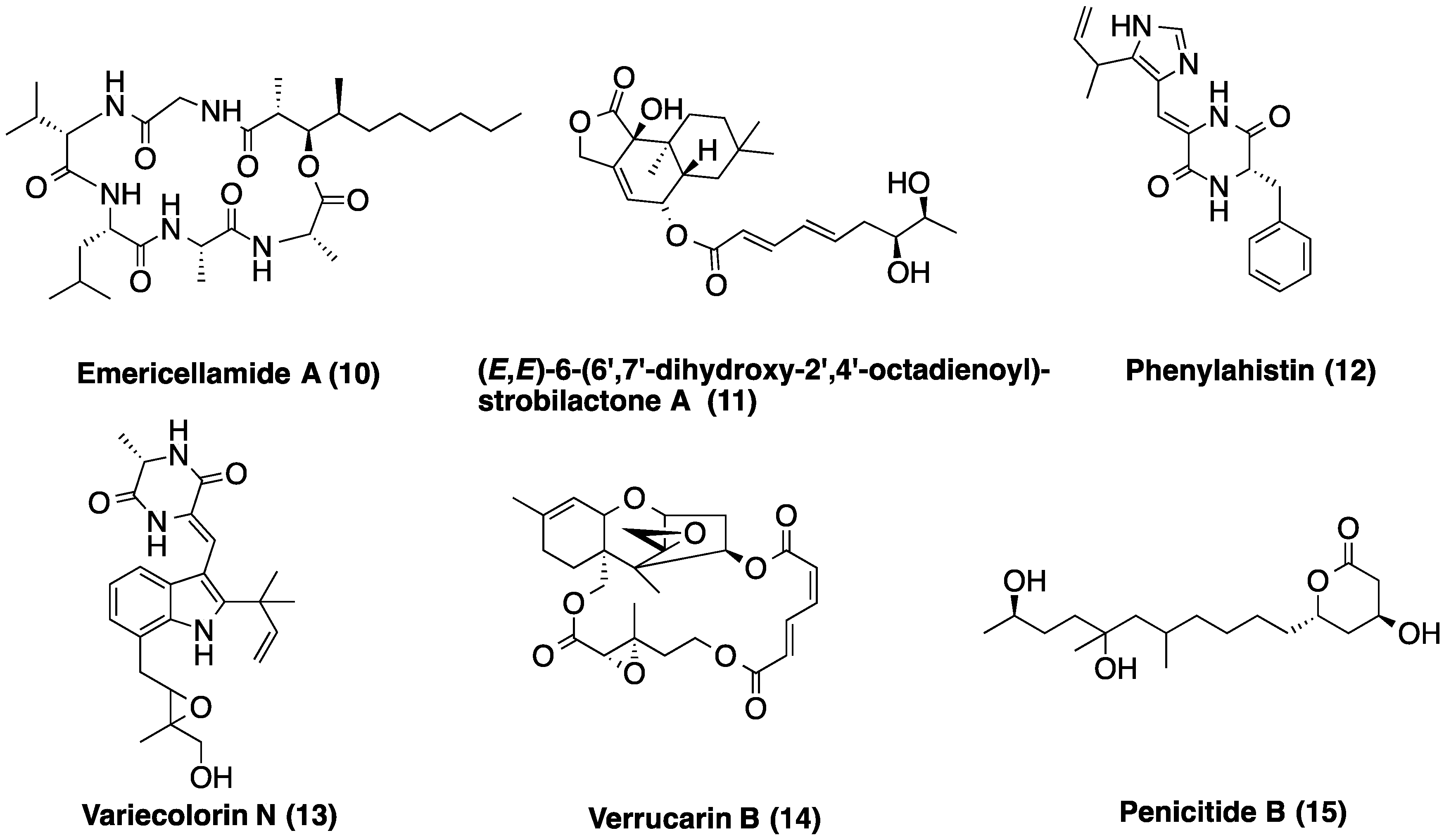
4. Experimental Section
4.1. Sampling and Isolation of Filamentous Fungi
4.1.1. Marine Endophytic Fungi
4.1.2. Root-Associated Fungi from Mangroves
4.1.3. Fungi from Saline Lakes
4.2. Small-Scale Fermentation and Preparation of Fungal Extracts
4.3. Quorum Sensing Inhibition
4.4. Statistics
4.5. DNA Extraction, PCR Amplification and Sequencing
4.6. Phylogenetic Analysis
4.7. LC-HRMS Metabolic Profiling
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Marques, J.C.; Lamosa, P.; Russell, C.; Ventura, R.; Maycock, C.; Semmelhack, M.F.; Miller, S.T.; Xavier, K.B. Processing the interspecies quorum-sensing signal autoinducer-2 (AI-2): Characterization of phospho-(S)-4,5-dihydroxy-2,3-pentanedione isomerization by LsrG protein. J. Biol. Chem. 2011, 286, 18331–18343. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Dow, J.M. Communication with a growing family: Diffusible signal factor (DSF) signaling in bacteria. Trends Microbiol. 2011, 19, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed]
- Brachmann, A.O.; Brameyer, S.; Kresovic, D.; Hitkova, I.; Kopp, Y.; Manske, C.; Schubert, K.; Bode, H.B.; Heermann, R. Pyrones as bacterial signaling molecules. Nat. Chem. Biol. 2013, 9, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S. A new bacterial chemical signal: Mapping the chemical space used for communication. Chembiochem 2014, 15, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Galloway, W.R.J.D.; Hodgkinson, J.T.; Bowden, S.D.; Welch, M.; Spring, D.R. Quorum sensing in Gram-negative bacteria: Small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev. 2011, 111, 28–67. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-L.; Dobretsov, S.; Xiong, H.; Qian, P.-Y. Effect of biofilm formation by Pseudoalteromonas spongiae on induction of larval settlement of the polychaete Hydroides elegans. Appl. Environ. Microbiol. 2007, 73, 6284–6288. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-L.; Shen, P.-J.; Liang, X.; Li, Y.-F.; Bao, W.-Y.; Li, J.-L. Larval settlement and metamorphosis of the mussel Mytilus coruscus in response to monospecific bacterial biofilms. Biofouling 2013, 29, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Twigg, M.S.; Tait, K.; Williams, P.; Atkinson, S.; Cámara, M. Interference with the germination and growth of Ulva zoospores by quorum-sensing molecules from Ulva-associated epiphytic bacteria. Environ. Microbiol. 2013, 16, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Tait, K.; Havenhand, J. Investigating a possible role for the bacterial signal molecules N-acylhomoserine lactones in Balanus improvisus cyprid settlement. Mol. Ecol. 2013, 22, 2588–2602. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, F.; Beltran, J.; Correa, J.A.; Lion, U.; Pohnert, G.; Kumar, N.; Steinberg, P.; Kloareg, B.; Potin, P. Spore release in Acrochaetium sp. (Rhodophyta) is bacterially controlled. J. Phycol. 2007, 43, 235–241. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; Steinberg, P.; Kjelleberg, S.; Givskov, M. How Delisea pulchra furanones affect quorum sensing and swarming motility in Serratia liquefaciens MG1. Microbiology 2000, 146, 3237–3244. [Google Scholar] [PubMed]
- Janssens, J.C.A.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated furanones inhibit biofilm formation by Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, G.; Milne, L.; Carter, G. Isolation and identification of antifungal polyesters from the marine fungus Hypoxylon oceanicum LL-15G256. Tetrahedron 2002, 58, 6825–6835. [Google Scholar] [CrossRef]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Bhadury, P.; Mohammad, B.T.; Wright, P.C. The current status of natural products from marine fungi and their potential as anti-infective agents. J. Ind. Microbiol. Biotechnol. 2006, 33, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.-Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. Biofouling 2010, 26, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, R.; Vitello, M.P.; Leitermann, F.; Syldatk, C. Advances in the production of sponge biomass Aplysina aerophoba-a model sponge for ex situ sponge biomass production. J. Biotechnol. 2006, 124, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Putz, A.; Ortlepp, S.; Kjer, J.; Bayer, M. Bioactive natural products from marine sponges and fungal endophytes. Phytochem. Rev. 2010, 9, 475–489. [Google Scholar] [CrossRef]
- Kobayashi, D.Y.; Crouch, J.A. Bacterial/Fungal interactions: From pathogens to mutualistic endosymbionts. Ann. Rev. Phytopathol. 2009, 47, 63–82. [Google Scholar] [CrossRef]
- Tarkka, M.T.; Sarniguet, A.; Frey-Klett, P. Inter-kingdom encounters: Recent advances in molecular bacterium-fungus interactions. Curr. Genet. 2009, 55, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef] [PubMed]
- Cugini, C.; Calfee, M.W.; Farrow, J.M.; Morales, D.K.; Pesci, E.C.; Hogan, D.A. Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol. Microbiol. 2007, 65, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.A.; Vik, A.; Kolter, R. A Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol. Microbiol. 2004, 54, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Yonezawa, T.; Lee, K.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Molecular phylogeny of the higher and lower taxonomy of the Fusarium genus and differences in the evolutionary histories of multiple genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, J.-P.; Hellio, C. Challenges for the development of new non-toxic antifouling solutions. Int. J. Mol. Sci. 2009, 10, 4623–4637. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Dahms, H.-U.; Qian, P.-Y. Inhibition of biofouling by marine microorganisms and their metabolites. Biofouling 2006, 22, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, Y.; Shao, C.-L.; Yang, R.-Y.; Zheng, C.-J.; Chen, Y.-Y.; Fu, X.-M.; Qian, P.-Y.; She, Z.-G.; de Voogd, N.J.; et al. Antibacterial bisabolane-type sesquiterpenoids from the sponge-derived fungus Aspergillus sp. Mar. Drugs 2012, 10, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.-H.; Xu, Y.; Xiong, H.-R.; Qian, P.-Y.; Zhang, S. Antifouling and antibacterial compounds from a marine fungus Cladosporium sp. F14. World J. Microbiol. Biotechnol. 2008, 25, 399–406. [Google Scholar] [CrossRef]
- He, F.; Liu, Z.; Yang, J.; Fu, P.; Peng, J.; Zhu, W.-M.; Qi, S.-H. A novel antifouling alkaloid from halotolerant fungus Penicillium sp. OUCMDZ-776. Tetrahedron Lett. 2012, 53, 2280–2283. [Google Scholar] [CrossRef]
- Bao, J.; Sun, Y.-L.; Zhang, X.-Y.; Han, Z.; Gao, H.-C.; He, F.; Qian, P.-Y.; Qi, S.-H. Antifouling and antibacterial polyketides from marine gorgonian coral-associated fungus Penicillium sp. SCSGAF 0023. J. Antibiot. 2013, 66, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Kwong, T.F.N.; Miao, L.; Li, X.; Qian, P.-Y. Novel antifouling and antimicrobial compound from a marine-derived fungus Ampelomyces sp. Mar. Biotechnol. 2006, 8, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Qian, P-Y.; Li, X.; Kwong, N.; Yang, H.; Dobretsov, S. Use of marine fungus originated compounds as antifouling agents. U.S. Patent 2006/0147410A1, 2006. [Google Scholar]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2006, 16, 49–67. [Google Scholar] [CrossRef]
- Jones, E.B.G. Are there more marine fungi to be described? Bot. Mar. 2011, 54, 343–354. [Google Scholar] [CrossRef]
- Khot, M.; Kamat, S.; Zinjarde, S.; Pant, A.; Chopade, B.; Ravikumar, A. Single cell oil of oleaginous fungi from the tropical mangrove wetlands as a potential feedstock for biodiesel. Microb. Cell Fact. 2012, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Smedsgaard, J.; Nielsen, J. Metabolite profiling of fungi and yeast: From phenotype to metabolome by MS and informatics. J. Exp. Bot. 2005, 56, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS One 2011, 6, e25321. [Google Scholar] [CrossRef] [PubMed]
- Cueto, M.; Jensen, P.R.; Kauffman, C.; Fenical, W.; Lobkovsky, E.; Clardy, J. Pestalone, a new antibiotic produced by a marine fungus in response to bacterial challenge. J. Nat. Prod. 2001, 64, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Kwong, T.F.N.; Qian, P.-Y. Effect of culture conditions on mycelial growth, antibacterial activity, and metabolite profiles of the marine-derived fungus Arthrinium c.f. saccharicola. Appl. Microbiol. Biotechnol. 2006, 72, 1063–1073. [Google Scholar] [CrossRef]
- Lema, K.A.; Willis, B.L.; Bourne, D.G. Corals form characteristic associations with symbiotic nitrogen-fixing bacteria. Appl. Environ. Microbiol. 2012, 78, 3136–3144. [Google Scholar] [CrossRef] [PubMed]
- Skindersoe, M.E.; Ettinger-Epstein, P.; Rasmussen, T.B.; Bjarnsholt, T.; de Nys, R.; Givskov, M. Quorum sensing antagonism from marine organisms. Mar. Biotechnol. 2008, 10, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.R.; Smith, S.M.; Downum, K.R.; Mydlarz, L.D. Microbial regulation in gorgonian corals. Mar. Drugs 2012, 10, 1225–1243. [Google Scholar] [CrossRef] [PubMed]
- Thenmozhi, R.; Nithyanand, P.; Rathna, J.; Pandian, S.K. Antibiofilm activity of coral-associated bacteria against different clinical M serotypes of Streptococcus pyogenes. FEMS Immunol. Med. Microbiol. 2009, 57, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Bakkiyaraj, D.; Sivasankar, C.; Pandian, S.K. Inhibition of quorum sensing regulated biofilm formation in Serratia marcescens causing nosocomial infections. Bioorg. Med. Chem. Lett. 2012, 22, 3089–3094. [Google Scholar] [CrossRef] [PubMed]
- Golberg, K.; Pavlov, V.; Marks, R.S.; Kushmaro, A. Coral-associated bacteria, quorum sensing disrupters, and the regulation of biofouling. Biofouling 2013, 29, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Alagely, A.; Krediet, C.J.; Ritchie, K.B.; Teplitski, M. Signaling-mediated cross-talk modulates swarming and biofilm formation in a coral pathogen Serratia marcescens. ISME J. 2011, 5, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Kharbikar, L.L.; Sharma, S.; Gupta, S.; Yadav, A. Phylogenetic relationships of Chaetomium isolates based on the internal transcribed spacer region of the rRNA gene cluster. Afr. J. Biotechnol. 2013, 12, 914–920. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Summerbell, R.C.; Gueidan, C.; Schroers, H.-J.; de Hoog, G.S.; Starink, M.; Rosete, Y.A.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Gazis, R.; Chaverri, P. Diversity of fungal endophytes in leaves and stems of wild rubber trees (Hevea brasiliensis) in Peru. Fungal Ecol. 2010, 3, 240–254. [Google Scholar] [CrossRef]
- Geiser, D.M.; Klich, M.A.; Frisvad, J.C.; Peterson, S.W.; Varga, J.; Samson, R.A. The current status of species recognition and identification in Aspergillus. Stud. Mycol. 2007, 59, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Steifert, K.A. Integrating DNA barcoding into the mycological sciences. Inoculum 2008, 59, 2–6. [Google Scholar]
- De Carvalho, M.P.; Abraham, W.-R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar]
- Holden, M.T.; Ram Chhabra, S.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Widmer, K.W.; Soni, K.A.; Hume, M.E.; Beier, R.C.; Jesudhasan, P.; Pillai, S.D. Identification of poultry meat-derived fatty acids functioning as quorum sensing signal inhibitors to autoinducer-2 (AI-2). J. Food Sci. 2007, 72, M363–M368. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.A.; Jesudhasan, P.; Cepeda, M.; Widmer, K.; Jayaprakasha, G.K.; Patil, B.S.; Hume, M.E.; Pillai, S.D. Identification of ground beef–derived fatty acid inhibitors of autoinducer-2-based cell signaling. J. Food Protect. 2008, 71, 134–138. [Google Scholar]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure op beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [PubMed]
- Deng, C.-M.; Liu, S.-X.; Huang, C.-H.; Pang, J.-Y.; Lin, Y.-C. Secondary metabolites of a mangrove endophytic fungus Aspergillus terreus (No. GX7-3B) from the South China Sea. Mar. Drugs 2013, 11, 2616–2624. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.; Kauffman, C.A.; Jensen, P.R.; Fenical, W.; Diego, S.; Jolla, L. Induced production of emericellamides A and B from the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 2007, 70, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.; Szewczyk, E.; Nayak, T.; Davidson, A.D.; James, F.; Lo, H.; Wen-yueh, H.; Simityan, H.; Kuo, E.; Praseuth, A.; et al. Molecular genetic mining of the Aspergillus secondary metabolome: Discovery of the emericellamide biosynthetic pathway. Chem. Biol. 2008, 15, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-H.; Miao, F.-P.; Qiao, M.-F.; Cichewicz, R.H.; Ji, N.-Y. Terretonin, ophiobolin, and drimane terpenes with absolute configurations from an algicolous Aspergillus ustus. RSC Adv. 2013, 3, 58–95. [Google Scholar]
- Cohen, E.; Koch, L.; Thu, K.M.; Rahamim, Y.; Aluma, Y.; Ilan, M.; Yarden, O.; Carmeli, S. Novel terpenoids of the fungus Aspergillus insuetus isolated from the Mediterranean sponge Psammocinia sp. collected along the coast of Israel. Bioorg. Med. Chem. 2011, 19, 6587–6593. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Edrada-Ebel, R.; Ebel, R.; Wang, Y.; Schulz, B.; Draeger, S.; Mu, W.E.G.; Wray, V.; Lin, W.; Proksch, P. Drimane sesquiterpenoids from the fungus Aspergillus ustus isolated from the marine sponge Suberites domuncula. J. Nat. Prod. 2009, 72, 1585–1588. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-N.; Zhu, T.-J.; Cai, S.-X.; Gu, Q.-Q.; Li, D.-H. Three new indole-containing diketopiperazine alkaloids from a deep-ocean sediment derived fungus Penicillium griseofulvum. Helv. Chim. Acta 2010, 93, 1758–1763. [Google Scholar] [CrossRef]
- Gao, S.-S.; Li, X.-M.; Du, F.-Y.; Li, C.-S.; Proksch, P.; Wang, B.-G. Secondary metabolites from a marine-derived endophytic fungus Penicillium chrysogenum QEN-24S. Mar. Drugs 2011, 9, 59–70. [Google Scholar] [CrossRef]
- Bacon, C.; Porter, J.; Norred, W.; Leslie, J. Production of fusaric acid by Fusarium species. Appl. Envir. Microbiol. 1996, 62, 4039–4043. [Google Scholar]
- Inokoshi, J.; Shigeta, N.; Fukuda, T.; Uchida, R.; Nonaka, K.; Masuma, R.; Tomoda, H. Epi-trichosetin, a new undecaprenyl pyrophosphate synthase inhibitor, produced by Fusarium oxysporum FKI-4553. J. Antibiot. 2013, 66, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Tatong, M.D.K.; Talontsi, F.M.; Abdel Rahim, H.M.D.; Islam, M.T.; Oswald, R.B.; Laatsch, H. Banchromene and other secondary metabolites from the endophytic fungus Fusarium sp. obtained from Piper guineense inhibit the motility of phytopathogenic Plasmopara viticola zoospores. Tetrahedron Lett. 2014, 55, 4057–4061. [Google Scholar] [CrossRef]
- van Rij, E.T.; Wesselink, M.; Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Influence of environmental conditions on the production of phenazine-1-carboxamide by Pseudomonas chlororaphis PCL1391. Mol. Plant. Microbe. Interact. 2004, 17, 557–566. [Google Scholar] [CrossRef] [PubMed]
- van Rij, E.T.; Girard, G.; Lugtenberg, B.J.J.; Bloemberg, G.V. Influence of fusaric acid on phenazine-1-carboxamide synthesis and gene expression of Pseudomonas chlororaphis strain PCL1391. Microbiology 2005, 151, 2805–2814. [Google Scholar] [CrossRef] [PubMed]
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 5, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Lugo, A.; Alcocer, J.; Sanchez, M. del R.; Escobar, E. Littoral protozoan assemblages from two Mexican hyposaline lakes. Hydrobiologia 1998, 381, 9–13. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Ilustrated Genera of Imperfect Fungi, 4th ed.; Macmillan Publishing Company: New York, NY, USA, 1998; p. 240. [Google Scholar]
- Watanabe, T. Pictorial Atlas of Soil and Seed Fungi Morphologies of Cultured Fungi and Key to Specie, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2002; p. 504. [Google Scholar]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; El Jaziri, M.; Baucher, M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010, 76, 243–253. [Google Scholar] [CrossRef]
- Martinelli, D.; Grossmann, G.; Séquin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.M.; Werner, K.E.; Kasai, M.; Francesconi, A.; Chanock, S.J.; Walsh, T.J. Rapid extraction of genomic DNA from medically important yeasts and filamentous fungi by high-speed cell disruption. J. Clin. Microbiol. 1998, 36, 1625–1629. [Google Scholar] [PubMed]
- Chung, W.-H.; Ishii, H.; Nishimura, K.; Ohshima, M.; Iwama, T.; Yoshimatsu, H. Genetic analysis and PCR-based identification of major Fusarium species causing head blight on wheat in Japan. J. Gen. Plant Pathol. 2008, 74, 364–374. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Bioinformatics applications note modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Rodríguez, A.J.; Reyes, F.; Martín, J.; Pérez-Yépez, J.; León-Barrios, M.; Couttolenc, A.; Espinoza, C.; Trigos, Á.; Martín, V.S.; Norte, M.; et al. Inhibition of Bacterial Quorum Sensing by Extracts from Aquatic Fungi: First Report from Marine Endophytes. Mar. Drugs 2014, 12, 5503-5526. https://doi.org/10.3390/md12115503
Martín-Rodríguez AJ, Reyes F, Martín J, Pérez-Yépez J, León-Barrios M, Couttolenc A, Espinoza C, Trigos Á, Martín VS, Norte M, et al. Inhibition of Bacterial Quorum Sensing by Extracts from Aquatic Fungi: First Report from Marine Endophytes. Marine Drugs. 2014; 12(11):5503-5526. https://doi.org/10.3390/md12115503
Chicago/Turabian StyleMartín-Rodríguez, Alberto J., Fernando Reyes, Jesús Martín, Juan Pérez-Yépez, Milagros León-Barrios, Alan Couttolenc, César Espinoza, Ángel Trigos, Víctor S. Martín, Manuel Norte, and et al. 2014. "Inhibition of Bacterial Quorum Sensing by Extracts from Aquatic Fungi: First Report from Marine Endophytes" Marine Drugs 12, no. 11: 5503-5526. https://doi.org/10.3390/md12115503
APA StyleMartín-Rodríguez, A. J., Reyes, F., Martín, J., Pérez-Yépez, J., León-Barrios, M., Couttolenc, A., Espinoza, C., Trigos, Á., Martín, V. S., Norte, M., & Fernández, J. J. (2014). Inhibition of Bacterial Quorum Sensing by Extracts from Aquatic Fungi: First Report from Marine Endophytes. Marine Drugs, 12(11), 5503-5526. https://doi.org/10.3390/md12115503