Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids


 , ,
, ,
Abstract
:
1. Introduction
2. Plastid Phylogeny

{kind=link}
{kind=link}
{kind=link}
| Pigment type | Cyanobacteria | Glaucophytes | Red algae | Brown algae | Diatoms | Green algae | Land plants |
|---|---|---|---|---|---|---|---|
| PBS | + | + | + | - | - | - | - |
| Chl a | +, except in Acaryochloris marina and related taxa | + | + | + | + | + | + |
| Chl b | except in Prochlorophytes | - | - | - | - | + | + |
| Chl c | - | - | - | + | + | - | - |
| Chl d | Only in Acaryochloris marina and related taxa | - | - | - | - | - | - |
| Chl f | Only in filamentous cyanobacteria from stromatolites | - | - | - | - | - | - |
| β-Carotene | + | + | Unicellular | + | + | + | + |
| Fucoxanthin | - | - | - | + | + | - | - |
| Diadinoxanthin | - | - | - | Traces | + | - | - |
| Diatoxanthin | - | - | - | Traces | + | - | - |
| Violaxanthin | - | - | + | + | + | + | + |
| Lutein | Depends on species | Macrophytes | - | - | + | + | |
| Zeaxanthin | Depends on species | + | + | + | Traces | + | + |
| Xanthophyll cycle | - | - | - | + | + | + | + |
| Echinenone | Depends on species | - | Traces | Traces | - | Adonis sp. | |
| Myxoxanthophyll | Depends on species | - | - | - | - | - | - |
| Canthaxanthin | Anabaena | - | - | - | - | Depen on species | Adonis sp. |
| Aphanizophyll | Depends on species | - | - | - | - | - | - |
| β-Cryptoxanthin | Aphanizomenon | - | + | - | - | + | + |
| Lycopene | Nostoc | - | - | - | - | Depend on species | + |
| Synechoxanthin | Synechococcus | - | - | - | - | - | - |
3. Bioactivity of Pigments and Lipids from Plastids of Marine Phytoplankton
3.1. Pigments
3.1.1. Tetrapyrroles
3.1.1.1. Anti-Oxidant, Anti-Inflammatory and Anti-Mutagenic Activities
3.1.1.2. Anti-Cancer
3.1.1.3. Miscellaneous
3.1.2. Carotenoids
| Pigments | Bioactivities | References |
|---|---|---|
| Aphanizophyll | Photoprotection | [97,98,99] |
| Astaxanthin | Anti-allergic, anti-cancer, anti cardiovascular diseases | [100,101,102] |
| Anti-oxidant | [100,103,104] | |
| Photoprotection | [38,105] | |
| Canthaxanthin | Anti-cancer | [106] |
| Anti-oxidant | [107] | |
| β-Carotene | Anti-allergic | [108,109] |
| Anti-cancer | [110,111] | |
| Anti-oxidant | [112] | |
| β-Cryptoxanthin | Anti-inflammatory | [113] |
| Improvement of skin health | [114] | |
| Diadinoxanthin/Diatoxanthin | Photoprotection | [115,116] |
| Fucoxanthin | Anti-allergic | [109,117,118] |
| Anti-cancer | [119,120] | |
| Anti-inflammatory | [121,122,123] | |
| Anti-obesity | [124,125,126,127] | |
| Anti-oxidant | [128,129] | |
| Photoprotection | [130] | |
| Fucoxanthinol | Anti-cancer | [131] |
| Anti-obesity | [124,125,126] | |
| Lutein | Anti-inflammatory | [113] |
| Protection of eyes | [131,132,133] | |
| Myxoxanthophyll | Anti-oxidant | [134,135] |
| Photoprotection | [97,98,99] | |
| Neoxanthin | Anti-cancer | [134] |
| Orange Carotenoid Protein | Photoprotection | [136,137,138,139] |
| Peridin | Anti-cancer | [140] |
| Sporopollenin | Photoprotection | [141] |
| Synechoxanthin | Anti-oxidant | [142] |
| Violaxanthin | Anti-cancer | [111,112,134] |
| Violeaxanthin | Anti-cancer | [111,112,134] |
| Zeaxanthin | Anti-allergic | [109] |
| Antioxidant | [135,143] | |
| Photoprotection | [139] |
3.1.2.1. Anti-Oxidant, Anti-Inflammatory and Anti-Mutagenic Activities
3.1.2.2. Anti-Cancer
3.1.2.3. Anti-Allergic
3.1.2.4. Anti-Obesity
3.1.2.5. Protection against Light
3.2. Lipids
3.2.1. Omega-3 Polyunsaturated Fatty Acids
3.2.1.1. Cardioprotective Effects
3.2.1.2. Anti-Obesity
3.2.1.3. Anti-Cholesterol
3.2.1.4. Normalizing Platelet Hyper-Aggregability
3.2.2. Glycerides
3.2.3. Phospholipids
4. Changing the Cultivation Conditions for Optimizing the Production of Bioactive Compounds
4.1. Pigments
4.1.1. Tetrapyrroles
4.1.2. Carotenoids
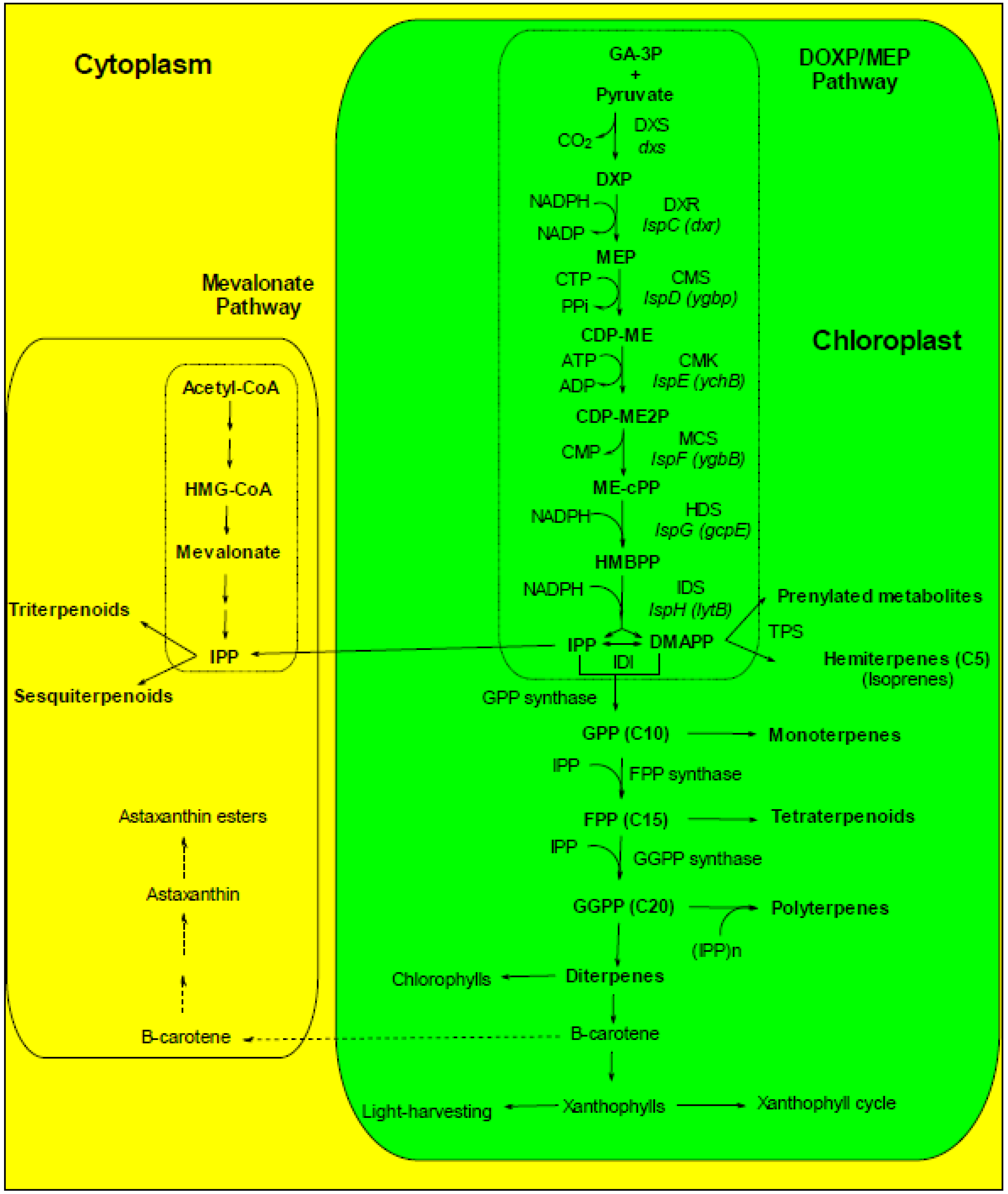
4.1.2.1. Temperature
4.1.2.2. Light Intensity
4.1.2.3. Nutrients
4.1.2.4. Other Stress
4.2. Lipids
4.2.1. Temperature
4.2.2. Light Intensity
4.2.3. Nutrients
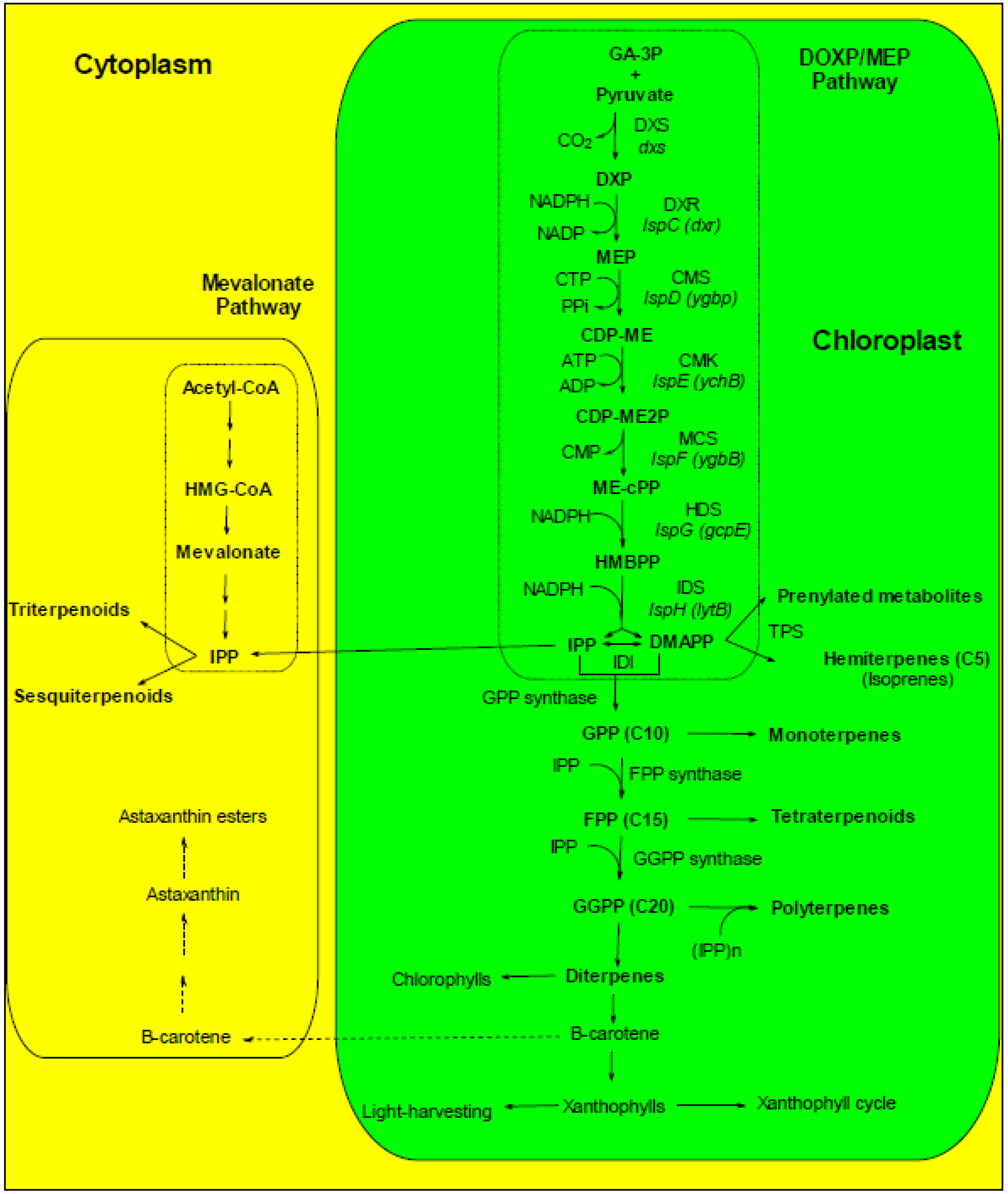
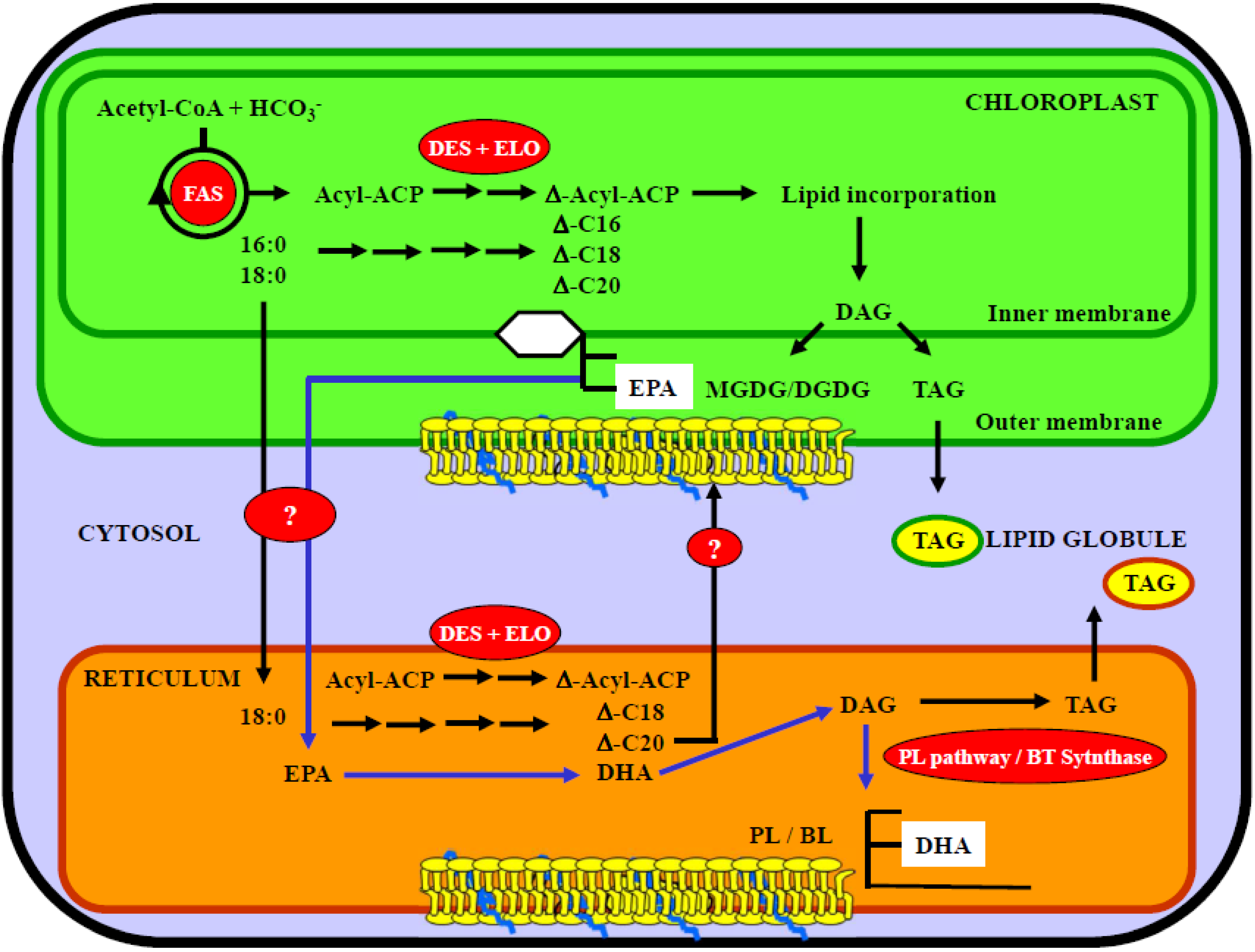
5. Role of Chloroplast and Reticulum Compartments in Lipid and Fatty Acid Synthesis in Marine Microalgae
5.1. Lipid and Fatty Acid Plastid Synthesis in Microalgae
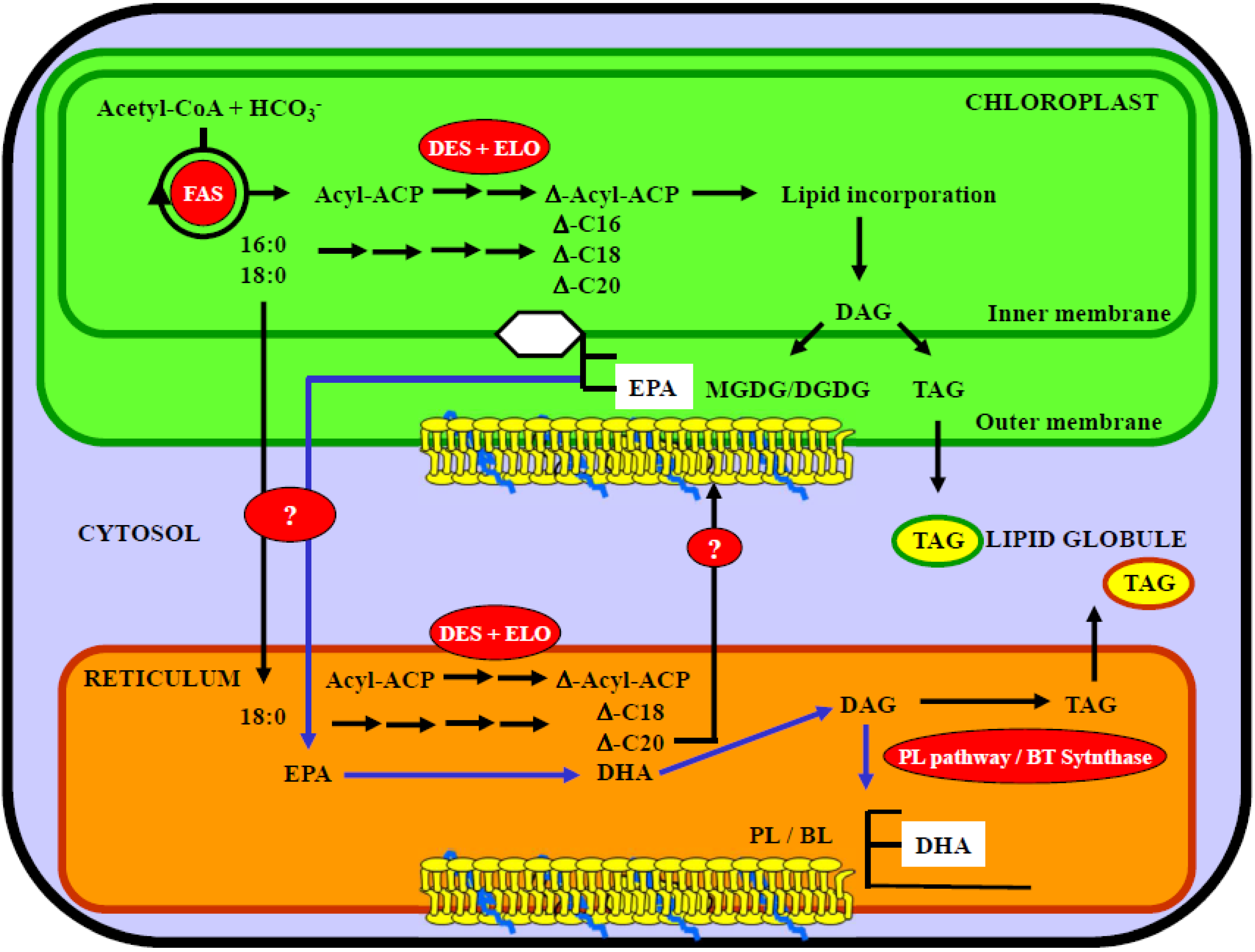
5.2. Compartmentalization of EPA and DHA Synthesis
6. Conclusion and Perspectives
Conflicts of Interest
References
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Blackwell Scientific: Oxford, England, 2007; p. 375. [Google Scholar]
- Tartar, A.; Boucias, D.G.; Becnel, J.J.; Adams, B.J. Comparison of plastid 16S rRNA (rrn16) genes from Helicosporidium spp.: Evidence supporting the reclassification of Helicosporidia as green algae (Chlorophyta). Int. J. Syst. Evol. Microbiol. 2003, 53, 1719–1723. [Google Scholar] [CrossRef]
- Ueno, R.; Urano, N.; Suzuki, M. Phylogeny of the non-photosynthetic green micro-algal genus Prototheca (Trebouxiophyceae, Chlorophyta) and related taxa inferred from SSU and LSU ribosomal DNA partial sequence data. FEMS Microbiol. Lett. 2003, 223, 275–280. [Google Scholar] [CrossRef]
- Gastineau, R.; Davidovich, N.A.; Bardeau, J.F.; Caruso, A.; Leignel, V.; Davidovich, O.I.; Rincé, Y.; Gaudin, P.; Cox, E.J.; Mouget, J.L. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the black sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar] [CrossRef]
- Caron, D.A.; Countway, P.D.; Jones, A.C.; Kim, D.Y.; Schnetzer, A. Marine protistan diversity. Annu. Rev. Mar. Sci. 2012, 4, 467–493. [Google Scholar] [CrossRef]
- Morales, E.A.; Rivera, S.F.; Wetzel, C.E.; Novais, M.H.; Hamilton, P.B.; Hoffmann, L.; Ector, L. New epiphytic araphid diatoms (Bacillariophyta) from Lake Titicaca, Bolivia. Diatom Res 2013. submitted for publication. [Google Scholar]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.P.; Morant-Manceau, A.; Schoefs, B. The potential of microalgae for the production of bioactive molecules of pharmaceutical interest. Curr. Pharmaceut. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef]
- Irigoien, X.; Huisman, J.; Harris, R.P. Global biodiversity patterns of marine phytoplankton and zooplankton. Nature 2004, 429, 863–867. [Google Scholar] [CrossRef]
- Kopp, R.E.; Kirschvink, J.L.; Hilburn, I.A.; Nash, C.Z. The paleoproterozoic snowball earth: A climate disaster triggered by the evolution of oxygenic photosynthesis. Proc. Natl. Acad. Sci. USA 2005, 102, 11131–11136. [Google Scholar]
- Allen, J.F.; Martin, W. Evolutionary biology: Out of thin air. Nature 2007, 445, 610–612. [Google Scholar] [CrossRef]
- Frei, R.; Gaucher, C.; Poulton, S.W.; Canfield, D.E. Fluctuations in precambrian atmospheric oxygenation recorded by chromium isotopes. Nature 2009, 461, 250–253. [Google Scholar] [CrossRef]
- Nelson, D.M.; Tréguer, P.; Brzezinski, M.A.; Leynaet, A.; Quéguiner, B. Production and dissolution of biogenic silica in the ocean. Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar]
- Bowler, C.; Vardi, A.; Allen, A.E. Oceanogarphic and biogeochemical insights from diatom genomes. Annu. Rev. Mar. Sci. 2010, 2, 333–365. [Google Scholar] [CrossRef]
- Moreau, D. Étude de Nouvelles Cibles Moléculaires de Cancer Bronchopulmonaire non à Petites Cellules Pharmacomodulées par des Substances Originales Naturelles et Synthétiques. Ph.D. Thesis, University of Nantes, Nantes, France, 23 November 2006; p. 280. [Google Scholar]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High lipid induction in microalgae for biodiesel production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-mining the microbial treasures of the ocean: New natural products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Beneficial effects of marine algal compounds in cosmeceuticals. Mar. Drugs 2013, 11, 146–164. [Google Scholar] [CrossRef]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from Spirulina platensis protect against diabetic nephropathy by inhibiting ooxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R110–R112. [Google Scholar] [CrossRef]
- Solymosi, K. Plastid structure, diversification and interconversions I. Algae. Curr. Chem. Biol. 2012, 6, 167–186. [Google Scholar] [CrossRef]
- Ramachandra, T.V.; Mahapatra, D.M.; Kartrick, B.; Gordon, R. Milking diatoms for sustainable energy: Biochemical engineering versus gasoline-secreting diatom solar panels. Ind. Eng. Chem. Res. 2009, 48, 8769–8788. [Google Scholar] [CrossRef]
- Amard, B.; Bertrand-Sarfati, J. Microfossils in 2000 Ma old Cherty Stromatolites of the Franceville Group, Gabon. Precambrian Res. 1997, 81, 197–221. [Google Scholar] [CrossRef]
- Jeltsch, A. Oxygen, epigenetic engineering, and the evolution of early life. Trends Biochem. Sci. 2013, 38, 172–176. [Google Scholar] [CrossRef]
- Nelson, N.; Ben-Shem, A. The complex architecture of oxygenic photosynthesis. Nat. Rev. Mol. Cell. Biol. 2004, 5, 971–982. [Google Scholar] [CrossRef]
- Schoefs, B. Determination of pigments in vegetables. J. Chromatogr. 2004, 1054, 217–226. [Google Scholar]
- Chen, M.; Blankenship, R.E. Expanding the solar spectrum used by photosynthesis. Trends Plant Sci. 2011, 16, 427–431. [Google Scholar] [CrossRef]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids. Volume 1A: Isolation and Analysis; Birkhäuser Verlag: Basel, Switzerland, 1995; p. 328. [Google Scholar]
- De Chlerck, O.; Bogaret, K.; Leliaert, F. Diversity and evolution of algae: Primary endosymbiosis. Adv. Bot. Res. 2012, 64, 55–86. [Google Scholar] [CrossRef]
- Nozaki, H.; Yang, Y.; Maruyama, S.; Suzaki, T. A case study for effects of operational taxonomic units from intracellular endoparasites and ciliates on the eukaryotic phylogeny: Phylogenetic position of the haptophyta in analyses of multiple slowly evolving genes. PLoS One 2012, 7, e50827. [Google Scholar]
- Bisanz, C.; Botté, C.; Saidani, N.; Bastien, O.; Cesbron-Delauw, M.-F.; Maréchal, E. Structure, Function and Biogenesis of the Secondary Plastid of Apicomplexan Parasites. In Plant Cell Compartments—Selected Topics; Schoefs, B., Ed.; Research Signpost: Kerala, India, 2008; p. 293. [Google Scholar]
- Moustafa, A.; Beszteri, B.; Maier, U.G.; Bowler, C.; Valentin, K.; Bhattacharya, D. Genomic footprints of a cryptic plastid endosymbiosis in diatoms. Science 2009, 324, 1724–1726. [Google Scholar] [CrossRef]
- Deschamps, P.; Moreira, D. Re-evaluating the green contribution to diatom genomes. Genome Biol. Evol. 2012, 4, 795–800. [Google Scholar]
- Archibald, J.M. The evolution of algae by secondary and tertiary endosymbiosis. Adv. Bot. Res. 2012, 64, 87–118. [Google Scholar] [CrossRef]
- Tirichine, L.; Bowler, C. Decoding algal genomes: Tracking back the history of photosynthetic life on earth. Plant J. 2011, 66, 45–57. [Google Scholar] [CrossRef]
- Roy, S.; Llewellyn, C.A.; Skarstadt Egeland, E.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; University Press: Cambridge, NY, USA, 2011. [Google Scholar]
- Schoefs, B. Chlorophyll and carotenoid analysis in food products. Properties of the pigments and methods of analysis. Trends Food Sci. Technol. 2002, 13, 361–371. [Google Scholar] [CrossRef]
- Beale, S.I.; von Weinstein, D. Biochemistry and Regulation of Photosynthetic Pigment Formation in Plants and Algae. In Biosynthesis of Tetrapyrroles; Jordan, P.M., Ed.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1991; pp. 155–309. [Google Scholar]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; de Girolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Micallef, M.A.; Garg, M.L. Beyond blood lipids: Phytosterols, statins and omega-3 polyunsaturated fatty acid therapy for hyperlipidemia. J. Nutr. Biochem. 2009, 20, 927–939. [Google Scholar] [CrossRef]
- Sakdarat, S.; Shuyprom, A.; Pientong, C.; Ekalaksananan, T.; Thongchai, S. Bioactive constituents from the leaves of Clinacanthus nutans lindau. Bioorg. Med. Chem. 2009, 17, 1857–1860. [Google Scholar] [CrossRef]
- Wang, S.Y.; Tseng, C.P.; Tsai, K.C.; Lin, C.F.; Wen, C.Y.; Tsay, H.S.; Sakamoto, N.; Tseng, C.H.; Cheng, J.C. Bioactivity-guided screening identifies pheophytin a as a potent anti-hepatitis C virus compound from Lonicera hypoglauca Miq. Biochem. Biophys. Res. Commun. 2009, 385, 230–235. [Google Scholar] [CrossRef]
- Baxter, H. Absorption of chlorophyll phytol in normal man and patients with refsum’s disease. J. Lipid Res. 1968, 9, 636–641. [Google Scholar]
- Ferruzzi, M.G.; Failla, M.; Schwartz, S.J. Assessment of degradation and intestinal cell uptake of chlorophyll derivatives from spinach puree using an in vitro digestion and Caco-2 human cell model. J. Agric. Food Chem. 2001, 49, 2082–2089. [Google Scholar] [CrossRef]
- Negishi, T.; Rai, H.; Hayatsu, H. Antigenotoxicity activity of natural chlorophylls. Mut. Res. 1997, 376, 97–100. [Google Scholar] [CrossRef]
- Dashwood, R.H. Chlorophylls as anticarcinogens. Int. J. Oncol. 1997, 10, 721–727. [Google Scholar]
- Ferruzzi, M.G.; Blakeslee, J. Digestion, absorption, and cancer preventative activity of dietary chlorophyll derivatives. Nutr. Res. 2007, 27, 1–12. [Google Scholar] [CrossRef]
- De Vogel, J.; Jonker-Termont, D.S.; Katan, M.B.; van der Meer, R. Natural chlorophyll but not chlorophyllin prevents heme-induced cytotoxic and hyperproliferative effects in rat colon. J. Nutr. 2000, 135, 1995–2000. [Google Scholar]
- Tang, P.M.; Zhang, D.M.; Bui Xuan, N.H.; Tsui, S.K.; Waye, M.M.; Kong, S.K.; Fong, W.P.; Fung, K.P. Photodynamic therapy inhibits P-glycoprotein mediated multidrug resistance via JNK activation in human hepatocellular carcinoma using the photosensitizer pheophorbide a. Mol. Cancer 2009, 8, 56–68. [Google Scholar] [CrossRef]
- Lohrey, E.; Tapper, B.; Hove, E.L. Photosensitization of albino-rats fed on luccerne-protein concentrate. Br. J. Nutr. 1974, 31, 159–166. [Google Scholar] [CrossRef]
- Holden, M. Chlorophyll degradation products in leaf protein preparations. J. Sci. Food Agric. 1974, 25, 1427–1432. [Google Scholar] [CrossRef]
- Hwang, D.F.; Tsai, J.S.; Chou, S.S.; Liu, S.; Wu, J.T.; Lin, S.; Tu, W.C. HPLC determination of pheophorbide a and pyropheophorbide a in dried laver products implicated in food poisoning. J. Food Hyg. Soc. Jpn. 2005, 46, 145–148. [Google Scholar]
- Gandul-Rojas, B.; Gallardo-Guerrero, L.; Minguez-Mosquera, M.I. Influence of chlorophyll structure on its transfer from an oily food matrix to intestinal epithelium cells. J. Agric. Food Chem. 2009, 57, 5306–5314. [Google Scholar]
- Gallardo-Guerrero, L.; Gandul-Rojas, B.; Minguez-Mosquera, M.I. Digestive stability, micellanization, and uptake of caco-2 human intestinal cell of chlorophyll derivatives from different preparations of pea (Pisum sativum L.). J. Food Chem. 2008, 56, 8379–8386. [Google Scholar] [CrossRef]
- Schoefs, B. Chlorophylls, Chlorophyll-Related Molecules, and Open-Chain Tetrapyrroles. In Handbook of Analysis of Active Compounds in Functional Foods; Nollet, L., Toldrà, F., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 665–685. [Google Scholar]
- Wrolstad, R.E. Handbook of Food Analytical Chemistry, 1st ed.; Wiley and Sons: Hookoken, NJ, USA, 2005. [Google Scholar]
- Cubas, C.; Lobos, M.G.; Gonzalez, M. Optimization of extraction of chlorophylls in green beans (Phaseolus vulgaris L.) by N,N-dimethylformamide using response surface methodology. J. Food Comp. Anal. 2008, 21, 125–133. [Google Scholar] [CrossRef]
- Harrison, J.W.; Levin, S.E.; Trabin, B. The safety of potassium copper chlorophyllin. J. Am. Pharm. Assoc. 1954, 43, 722–737. [Google Scholar]
- Reber, E.F.; Willigan, D.A. The effects of a chlorophyll derivative when included in a ration fed rats. II. Reproduction, blood, and tissue studies. Am. J. Vet. Res. 1954, 15, 643–646. [Google Scholar]
- Anonymous 1. 2002. Food Additives in Europe 2000. Thema Nord. Available online: http://www.norden.org/en/publications/publications/2002–560set_language=en (accessed on 1 April 2013).
- Conrad, M.E.; Benjamin, B.I.; Williams, H.L.; Foy, A.L. Human absorption of haemoglobin iron. Gastroenterology 1967, 53, 5–10. [Google Scholar]
- Shayeghi, M.; Latunde-Dada, G.O.; Oakhill, J.S.; Laftah, A.H.; Takeuchi, K.; Halliday, N.; Khan, Y.; Warley, A.; McCann, F.E.; Hider, R.C.; et al. Identification of an intestinal haem transporter. Cell 2005, 122, 789–801. [Google Scholar]
- Raffin, S.B.; Woo, C.H.; Roost, K.T.; Price, D.C.; Schmid, R. Intestinal absorption of hemoglobin iron-heme cleavage by mucosal heme oxygenase. J. Clin. Invest. 1974, 54, 1344–1352. [Google Scholar] [CrossRef]
- Geiselmann, J.; Houmard, J.; Schoefs, B. Regulation of Phycobilisome Biosynthesis. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; Marcel. Dekker: New York, NY, USA, 2005; pp. 425–438. [Google Scholar]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Jelic, D.; Iva Tatic, I.; Trzun, M.; Hrvacic, B.; Brajsa, K.; Verbanac, D.; Tomaskovic, M.; Culic, O.; Antolovic, R.; Glojnaric, I.; et al. Porphyrins as new endogenous anti-inflammatory agents. Eur. J. Pharmacol. 2012, 691, 251–260. [Google Scholar] [CrossRef]
- Von Wettstein, D.; Gough, S.; Kannangara, C.G. Chlorophyll biosynthesis. Plant Cell 1995, 7, 1039–1057. [Google Scholar]
- Franck, F.; Schoefs, B.; Barthélemy, X.; Mysliwa-Kurdziel, B.; Strzalka, K.; Popovic, R. Protection of native chlorophyll(ide) forms and of photosystem II against photodamage during early stages of chloroplast differentiation. Acta Physiol. Plant 1995, 17, 123–132. [Google Scholar]
- Schoefs, B.; Franck, F. Protochlorophyllide reduction: Mechanisms and evolution. Photochem. Photobiol. 2003, 78, 543–557. [Google Scholar] [CrossRef]
- Priyadarsini, R.V.; Kumar, N.; Khan, I.; Thiyagarajan, P.; Kondaiah, P.; Nagini, S. Gene expression signature of DMBA-induced hamster buccal pouch carcinomas: Modulation by chlorophyllin and ellagic acid. PloS One 2012, 7, e34628. [Google Scholar]
- Ge, B.; Qin, S.; Han, L.; Lin, F.; Ren, R. Antioxidant properties of recombinant allophycocyanin expressed in Escherichia coli. J. Photochem. Photobiol. B Biol. 2006, 84, 175–180. [Google Scholar] [CrossRef]
- Patel, A.; Mishra, S.; Ghosh, P.K. Antioxidant potential of C-phycocyanin isolated from cyanobacterial species Lyngbya, Phormidium and Spirulina spp. Ind. J. Biochem. Biophys. 2006, 43, 25–31. [Google Scholar]
- Soni, B.; Visavadiya, N.P.; Madamwar, D. Attenuation of diabetic complications by c-phycoerythrin in rats: Antioxidant activity of C-phycoerythrin including copper-induced lipoprotein and serum oxidation. Br. J. Nutr. 2009, 102, 102–109. [Google Scholar] [CrossRef]
- Romay, C.; Armesto, J.; Remirez, D.; Gonzalez, R.; Ledon, N.; Garcia, I. Antioxidant and antiinflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar]
- Benedetti, S.; Benvenuti, F.; Scoglio, S.; Canestrari, F. Oxygen Radical absorbance capacity of phycocyanin and phycocyanobilin from the food supplement Aphanizomenon flos-aquae. J. Med. Food 2010, 13, 223–227. [Google Scholar] [CrossRef]
- Nollet, L.; Toldrà, F. Handbook of Analysis of Active Compounds in Functional Foods; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Gross-Steinmeyer, K.; Eaton, D.L. Dietary modulation of the biotransformation and genotoxicity of aflatoxin B(1). Toxicology 2012, 299, 69–79. [Google Scholar] [CrossRef]
- Bedard, L.L.; Massey, T.E. Aflatoxin B1-induced damage and its repair. Cancer Lett. 2006, 241, 174–183. [Google Scholar] [CrossRef]
- Hayatsu, H.; Negishi, T.; Arimoto-Kobayashi, S. Chlorophyll. In Chemoprevention of Cancer and DNA Damage by Dietary Factors; Knasmueller, S., Demarini, D.M., Johnson, I., Gerhauser, C., Eds.; Wiley-VCH: Weinheim, Germany, 2009; pp. 699–708. [Google Scholar]
- Sinha, R.; Perters, U.; Cross, A.J.; Kulldorff, M.; Weissfeld, J.L.; Pinsky, P.F.; Rothman, N.; Hayes, R.B. Meat, meat cooking methods and preservation, and risk for colorectal adenoma. Cancer Res. 2005, 65, 8034–8041. [Google Scholar] [CrossRef]
- Breinholt, V.; Schimerlik, M.; Dashwood, R.; Bailley, G. Mechanisms of chlorophyllin anti-carcinogenesis against aflatoxin B1. Complex formation with the carcinogen. Chem. Res. Toxicol. 1999, 8, 506–514. [Google Scholar]
- Yun, C.Y.; Hye, G.J.; Jhoun, J.W.; Guengerich, F.P. Non-specific inhibition of cytochrome P450 activites by chlorophyllin in human and rat liver microsomes. Carcinogenesis 1995, 16, 1437–1440. [Google Scholar] [CrossRef]
- Egner, P.A.; Wang, J.B.; Zhu, Y.R.; Zhang, B.C.; Wu, Y.; Zhang, Q.N.; Qian, G.S.; Kuang, S.Y.; Gange, S.J.; Jacobson, L.P.; et al. Chlorophyllin intervention reduces aflatoxin-DNA adducts in individuals at high risk for liver cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 14601–14606. [Google Scholar]
- Shaughnessy, D.T.; Gangarosa, L.M.; Schliebe, B.; Umbach, D.M.; Xu, Z.; MacIntosh, B.; Knize, M.G.; Mattews, P.P.; Swank, A.E.; Sandler, R.S.; et al. Inhibition of fried meat-induced colorectal DNA damage and altered systemic genotoxicity in humans by crucifera, chlorophyllin, and yogurt. PloS One 2011, 6, e18707. [Google Scholar] [CrossRef]
- Chernomorsky, S.; Segalman, A.; Poretz, R.D. Effects of dietary chlorophyll derivatives on mutagenesis and tumor cell growth. Teratogen. Carcinogen. Mutagen. 1999, 19, 313–322. [Google Scholar] [CrossRef]
- Chiu, L.C.; Kong, C.K.; Ooi, V.E. The Chlorophyllin-induced cell cycle arrest and apoptosis in human breast MCF-7 cells is associated with ERK deactivation and cyclin D1 depletion. Int. J. Mol. Med. 2005, 16, 735–740. [Google Scholar]
- Carter, O.; Bailey, G.S.; Dashwood, R.H. The dietary phytochemical chlorophyllin alters E-cadherin and beta-catenin expression in human colon cancer cells. J. Nutr. 2004, 34, 341–344. [Google Scholar]
- Chiu, L.C.; Kong, C.K.; Ooi, V.E. Antiproliferative effect of chlorophyllin derived from a traditional Chinese medicine Bombyx mori excreta on human breast cancer. Int. J. Oncol. 2003, 23, 729–735. [Google Scholar]
- Nelson, R.L. Chlorophyllin, an antimutagen acts as a tumor promoter in the rat-dimethylhydrazide colon carcinogenesis model. Anticancer Res. 1992, 12, 737–739. [Google Scholar]
- Tang, P.M.K.; Chan, J.Y.W.; Zhang, D.M.; Au, S.W.N.; Fong, W.P.; Kong, S.K.; Tsui, S.K.W.; Waye, M.M.Y.; Mak, T.C.W.; Fung, K.P. Pheophorbide a, an active component in Scutellaria barbata, reverses P-glycoprotein-mediated multidrug resistance on a human hepatoma cell line R-HepG2. Cancer Biol. Ther. 2007, 6, 504–509. [Google Scholar] [CrossRef]
- Yoshioka, H.; Kamata, A.; Konishi, T.; Takahashi, J.; Oda, H.; Tamai, T.; Toyohara, H.; Sugahara, T. Inhibitory effect of chlorophyll c2 from brown algae, Sargassum horneri, on degranulation of RBL-2H3 cells. J. Func. Foods 2012, 5, 204–210. [Google Scholar]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 103–136. [Google Scholar]
- Eonseon, J.; Polle, J.E.W.; Lee, H.K.; Hyun, S.M.; Chang, M. Xanthophylls in microalgae: From biosynthesis to biotechnological mass production and application. J. Microbiol. Biotechnol. 2003, 13, 165–174. [Google Scholar]
- Berera, R.; van Stokkum, I.H.M.; Gwizdala, M.; Wilson, A.; Kirilovsky, D.; van Grondelle, R. The photophysics of the orange carotenoid protein, a light-powered molecular switch. J. Phys. Chem. B 2012, 116, 2568–2574. [Google Scholar] [CrossRef]
- Kirilovsky, D.; Kerfeld, C.R. The orange carotenoid protein: A blue-green light photoactive protein. Photochem. Photobiol. Sci. 2013, 12, 1135–1143. [Google Scholar] [CrossRef]
- Hashtroudi, M.S.; Shariatmadari, Z.; Riahi, H.; Ghassempour, A. Analysis of Anabaena vaginicola and Nostoc calcicola from Northern Iran, as rich sources of major carotenoids. Food Chem. 2013, 136, 1148–1153. [Google Scholar] [CrossRef]
- Mehnert, G.; Rücker, J.; Nicklish, A.; Leunert, F.; Wiedner, C. Effects of thermal acclimation and photoacclimation on lipophilic pigments in an invasive and a native cyanobacterium of temperate regions. Eur. J. Phycol. 2012, 47, 182–192. [Google Scholar] [CrossRef]
- Bonilla, S.; Aubriot, L.; Soares, M.C.S.; González-Piana, M.; Fabre, A.; Huszar, V.L.M.; Lürling, M.; Antoniades, D.; Padisák, J.; Kruk, C. What drives the distribution of the bloom-forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? FEMS Microbiol. Ecol. 2012, 79, 594–607. [Google Scholar]
- Montero, M.; Sanchez-Gujo, A.; Lubian, L.M.; Martinez-Rodriguez, G. Changes in membrane lipids and carotenoids during light acclimation in a marine cyanobacterium Synechococcus sp. J. Biosci. 2012, 37, 635–645. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potentialin human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Miki, W. Biological functions and activities of carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Djordjevic, B.; Baralic, I.; Kotur-Stevuljevic, J.; Stefanovic, A.; Ivanisevic, J.; Radivojevic, N.; Andjelkovic, M.; Dikic, N. Effect of astaxanthin supplementation on muscle damage and oxidative stress markers in elite young soccer players. J. Sports Med. Phys. Fitness. 2012, 52, 382–392. [Google Scholar]
- Caramujo, M.J.; de Carvalho, C.C.C.R.; Silva, S.J.; Carman, K.R. Dietary carotenoids regulate astaxanthin content of copepods and modulate their susceptibility to UV light and copper toxicity. Mar. Drugs 2012, 10, 998–1018. [Google Scholar] [CrossRef]
- Dlamini, N.; Moroka, T. Indigenous Edible Plants as Sources of Nutrients and Health Benefitting Components (Nutraceuticals). Available online: http://researchspace.csir.co.za/dspace/bitstream/10204/4224/3/Dlamini_2010_Presentation.pdf (accessed on 15 April 2013).
- Malik, K.; Tokkas, J.; Goyal, S. Microbial pigments: A review. Int. J. Microbial. Resour. Technol. 2012, 1, 361–365. [Google Scholar]
- Koizumi, T.; Bando, N.; Terao, J.; Yamanishi, R. Feeding with both β-Carotene and supplemental α-tocopherol enhances type 1 Helper T cell activity among splenocytes isolated from DO11.10 mice. Biosci. Biotechnol. Biochem. 2006, 70, 3042–3045. [Google Scholar] [CrossRef]
- Vo, T.S.; Ngo, D.H.; Kim, S.K. Marine algae as a potential pharmaceutical source for anti-allergic therapeutics. Process Biochem. 2012, 47, 386–394. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; de Brabanter, J.; de Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Molnar, J.; Gyemant, N.; Mucsi, I.; Molnar, A.; Szabo, M.; Kortvelyesi, T.; Varga, A.; Molnar, P.; Toth, G. Modulation of multidrug resistance and apoptosis of cancer cells by selected carotenoids. In Vivo 2004, 18, 237–244. [Google Scholar]
- Molnar, J.; Engi, H.; Hohmann, J.; Molnar, P.; Deli, J.; Wesolowska, O.; Michalak, K.; Wang, Q. Reversal of multidrug resistance by natural substances from plants. Curr. Top. Med. Chem. 2010, 10, 1757–1768. [Google Scholar] [CrossRef]
- Di Filippo, M.M.; Mathison, B.D.; Park, J.S.; Chew, B.P. Lutein and β-cryptoxanthin inhibit inflammatory mediators in human chondrosarcoma cells induced with IL-1β. Open Nutr. J. 2012, 6, 41–47. [Google Scholar] [CrossRef]
- Sayo, T.; Sugiyama, Y.; Inoue, S. Lutein, a nonprovitamin a, activates the retinoic acid receptor to induce HAS3-dependent hyaluronan synthesis in keratinocytes. Biosci. Biotechnol. Biochem. 2013, in press. [Google Scholar]
- Moulin, P.; Lemoine, Y.; Schoefs, B. Carotenoids and Stress in Higher Plants and Algae. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, M., Ed.; Taylor and Francis: New York, NY, USA, 2011; pp. 407–433. [Google Scholar]
- Cadoret, J.P.; Garnier, M.; Saint-Jean, B. Microalgae, functional genomics and biotechnoloy. Adv. Bot. Res. 2012, 64, 285–341. [Google Scholar] [CrossRef]
- Sakai, S.; Sugawara, T.; Matsubara, K.; Hirata, T. Inhibitory effect of carotenoids on the degranulation of mast cells via suppression of antigen-induced aggregation of high affinity IgE receptors. J. Biol. Chem. 2009, 284, 28172–28179. [Google Scholar] [CrossRef]
- Vo, T.S.; Ngo, D.H.; Kim, S.K. Potential targets for anti-inflammatory and anti-allergic activities of marine algae: An overview. Inflamm. Allergy Drug Targets 2012, 11, 90–101. [Google Scholar] [CrossRef]
- Kim, K.N.; Heo, S.J.; Kang, S.M.; Ahn, G.; Jeon, Y.J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. In Vitro 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Moreau, D.; Tomasoni, C.; Cathrine, J.; Kaas, R.; le Guedes, R.; Cadoret, J.P.; Muller-Feuga, A.; Konitza, I.; Vagias, C.; Roussis, V.; et al. Cultivated microalgae and the carotenoid fucoxanthin from Odontella aurita as potent anti-proliferative agents in bronchopulmonary and epithelial cell lines. Envir. Toxicol. Pharmacol. 2006, 22, 97–103. [Google Scholar] [CrossRef]
- Shiratori, K.; Okgami, K.; Ilieva, I.; Jin, X.H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Affan, A.; Oh, C.; Jung, W.K.; Jeon, Y.J. Evaluation of antiinflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, M.S.; Choi, H.J.; Choi, J.W.; Shin, T.; Woo, H.C.; Kim, J.; Kim, H.R. Hexane fraction from Laminaria japonica exerts anti-inflammatory effects on lipopolysaccharide-stimulated RAW 264.7 macrophages via inhibiting NF-kappaB pathway. Eur. J. Nutr. 2013, 52, 409–421. [Google Scholar] [CrossRef]
- Jeon, S.M.; Kim, H.J.; Woo, M.N.; Lee, M.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin-rich seaweed extract suppresses body weight gain and improves lipid metabolism in high-Fat-Fed C57BL/6J Mice. Biotechnol. J. 2010, 5, 961–969. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hosokawa, M.; Matsukawa, N.; Hagio, M.; Shinoki, A.; Nishimukai, M.; Miyashita, M.; Yajima, T.; Hara, H. Suppressive effects of the marine carotenoids, fucoxanthin and fucoxanthinol on triglyceride absorption in lymph duct-cannulated rats. Eur. J. Nutr. 2010, 49, 243–249. [Google Scholar] [CrossRef]
- Woo, M.N.; Jeon, S.M.; Shin, Y.C.; Lee, M.K.; Kang, M.A.; Choi, M.S. Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice. Mol. Nutr. Food Res. 2009, 53, 1603–1611. [Google Scholar] [CrossRef]
- Hu, T.; Liu, D.; Chen, Y.; Wu, J.; Wang, S. Antioxidant activity of sulfated polysaccharide fractions extracted from Undaria pinnitafidain vitro. Int. J. Biol. Macromol. 2010, 46, 193–198. [Google Scholar] [CrossRef]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Biol. Int. 1997, 42, 361–370. [Google Scholar]
- Shimoda, H.; Tanaka, J.; Shan, S.J.; Maoka, T. Anti-pigmentary activity of fucoxanthin and its influence on skin mRNA expression of melanogenic molecules. J. Pharm. Pharmacol. 2010, 62, 1137–1145. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ozaki, Y.; Mizuno, M.; Yoshida, M.; Nishitani, Y.; Azuma, T.; Komoto, A. Pharmacokinetics of fucoxanthinol in human plasma after the oral administration of kombu extract. Br. J. Nutr. 2011, 107, 1–4. [Google Scholar]
- Alves-Rodrigues, A.; Shao, A. The science behind lutein. Toxicol. Lett. 2004, 150, 57–83. [Google Scholar] [CrossRef]
- Granado, F.; Olmedilla, B.; Blanco, I. Nutritional and clinical relevance of lutein in human health. Br. J. Nutr. 2003, 90, 487–502. [Google Scholar] [CrossRef]
- Ugocsai, K.; Varga, A.; Molnar, P.; Antus, S.; Molnar, J. Effects of selected flavonoids and carotenoids on drug accumulation and apoptosis induction in multidrug-resistant colon cancer cells expressing MDR1/LRP. In Vivo 2005, 19, 433–438. [Google Scholar]
- Kotake-Nara, E.; Nagao, A. Absorption and metabolism of xanthophylls. Mar. Drugs 2011, 9, 1024–1037. [Google Scholar] [CrossRef]
- Gwizdala, M.; Wilson, A.; Kirilovsky, D. In vitro reconstitution of the canobacterial photoprotective mechanism mediated by the orange carotenoid protein in Synechocystis PCC 6803. Plant Cell 2011, 23, 2631–2643. [Google Scholar] [CrossRef]
- Tian, L.; van Stokkum, I.H.M.; Koehorst, R.B.M.; Jongerius, A.; Kirilovsky, D.; van Amerongen, H. Site, rate, and mechanism of photoprotective quenching in cyanobacteria. J. Am. Chem. Soc. 2011, 133, 18304–18311. [Google Scholar]
- Polívka, T.; Chábera, P.; Kerfeld, C.A. Carotenoid-protein interaction alters the S1 energy of hydroxyechinenone in the orange carotenoid protein. Biochim. Biophys. Acta 2013, 1827, 248–254. [Google Scholar]
- Kirilovsky, D.; Kerfeld, C.A. The orange carotenoid protein in photoprotection of photosystem II in cyanobacteria. Biochim. Biophys. Acta 2012, 1817, 158–166. [Google Scholar]
- Gagez, A.L.; Thiery, V.; Pasquet, V.; Cadoret, J.P.; Picot, L. Epoxycarotenoids and cancer. Review. Curr. Bioact. Comp. 2012, 8, 109–141. [Google Scholar] [CrossRef]
- Xiong, X.; Lederer, F.; Lukavsky, J.; Nedbal, L. Screening of freshwater algae (Chlorophyta, Chromophyta) for ultraviolet-B sensitivity of the photosynthetic apparatus. J. Plant Physiol. 1996, 148, 42–48. [Google Scholar] [CrossRef]
- Fujii, S.; Chang, S.Y.; Burke, M.D. Snthesis of synechoxanthin. Synfacts 2011, 11, 1161–1161. [Google Scholar]
- Chakdar, H.; Jadhav, S.D.; Dhar, D.W.; Pabbi, S. Potential applications of blue green algae. J. Sci. Ind. Res. 2012, 71, 13–20. [Google Scholar]
- Erlinger, T.P.; Guallar, E.; Miller, E.R.; Stolzenberg-Solomon, R.; Appel, L.J. Relationshipbetween systemic markers of inflammation and serum beta-carotene levels. Arch. Intern. Med. 2001, 161, 1903–1908. [Google Scholar] [CrossRef]
- Rowley, K.; Walker, K.Z.; Cohen, J. Inflammation and vascular endothelial activation in an aboriginal population: Relationships to coronary disease risk factors and nutritional markers. Med. J. Aust. 2003, 178, 495–500. [Google Scholar]
- Paredes, S.; Girona, J.; Hurt-Camejo, E. Antioxidant vitamins and lipid peroxidation in patients with rheumatoid arthritis: Association with inflammatory markers. J. Rheumatol. 2002, 29, 2271–2277. [Google Scholar]
- Allen, R.G.; Tresini, M. Oxidative stress and gene regulation. Free Radic. Biol. Med. 2000, 28, 463–499. [Google Scholar] [CrossRef]
- Dufosse, L.; Galaup, P.; Yaron, A.; Malis Arad, S.; Blanc, P.; Murthy, K.N.C. Microorganisms and microalgae as sources of pigments for food use: A scientific oddity or an industrial reality? Trends Food Sci. Technol. 2005, 16, 389–406. [Google Scholar]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sust. Energ. Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Mostafa, S.S.M. Microalgal Biotechnology: Prospects and Applications. Available online: http://www.intechopen.com/books/plant-science/microalgal-biotechnology-prospects-and-applications (accessed on 1 April 2013).
- Paiva, S.A.R.; Russell, R.M.; Dutta, S.K. β-Carotene and other carotenoids as antioxidants. J. Am. Coll. Nutr. 1999, 18, 426–433. [Google Scholar] [CrossRef]
- Firestone, G.L.; Kapadia, B.J. Minireview: Regulation of gap junction dynamics by nuclear hormone receptors and their ligands. Mol. Endocrinol. 2012, 26, 1798–1807. [Google Scholar] [CrossRef]
- Olson, J.A. Carotenoids and human health. Arch. Latinoamer. Nutr. 1999, 49, 7–11. [Google Scholar]
- Gyemant, N.; Tanaka, M.; Molnar, P.; Deli, J.; Mandoky, L.; Molnar, J. Reversal of multidrug resistance of cancer cells in vitro: Modification of drug resistance by selected carotenoids. Anticancer Res. 2006, 26, 367–374. [Google Scholar]
- Sato, Y.; Akiyama, H.; Matsuoka, H.; Sakata, K.; Nakamura, R.; Ishikawa, S.; Inakuma, T.; Totsuka, M.; Sugita-Konishi, Y.; Ebisava, M.; Teshima, R. Dietary carotenoids inhibit oral sensitization and the development of food allergy. J. Agric. Food Chem. 2010, 58, 7180–7186. [Google Scholar] [CrossRef]
- Okada, T.; Nakai, M.; Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Suppressive effect of neoxanthin on the differentiation of 3T3-L1 adipose cells. J. Oleo Sci. 2008, 57, 345–351. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; McTavish, H.S.; Dunlap, W.C.; Vesk, M.; Groenewood, K. Occurrence of UVA- and UVB756 absorbing compounds in 152 species (206 Strains) of marine microalgae. Mar. Ecol. Prog. Ser. 1999, 189, 35–51. [Google Scholar] [CrossRef]
- Gwizdala, M.; Wilson, A.; Omairi-Nasser, A.; Kirilovsky, D. Characterization of the Synechocystis PCC 6803 fluorescence recovery protein involved in photoprotection. Biochim. Biophys. Acta 2013, 1827, 348–354. [Google Scholar]
- Alonso, D.L.; Belarbi, E.H.; Rodriguez-Ruiz, J.; Segura, C.I.; Gimenez, A. Acyl lipids of three microalgae. Phytochemistry 1998, 47, 1473–1481. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Macalta, F.X. Effect of culture media on production of polyunsaturated fatty acids by Pavlova lutheri. Cryptogamie Algol. 2000, 21, 59–71. [Google Scholar] [CrossRef]
- Tatsuzawa, H.; Takizawa, E. Changes in lipid and fatty acid composition of Pavlova lutheri. Phytochemistry 1995, 40, 397–400. [Google Scholar] [CrossRef]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Identification of a very long chain polyunsaturated fatty acid [Delta] 4-desaturase from the microalga Pavlova lutheri. FEBS Lett. 2003, 553, 440–444. [Google Scholar] [CrossRef]
- Braud, J.P. Simoultaneous Culture in Pilot Tanks of the Macroalga Chondrus crispus (Gigartinaceae) and the Microalga Odontella aurita (Eupodiscaceae) Producing EPA. In Marine Microorganisms for Industry; Le Gal, Y., Muller-Feuga, A., Eds.; Actes de colloques: Ifremer, France, 1997; pp. 39–47. [Google Scholar]
- Nuno, K.; Villaruel-Lopez, A.; Puebla-Perez, A.M.; Romero-Velarde, E.; Puebla-Mora, A.G.; Ascencio, F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Func. Foods 2013, 5, 106–115. [Google Scholar] [CrossRef]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef]
- Nemoto, N.; Suzuki, S.; Kikuchi, H.; Okabe, H.; Sassa, S.; Sakamoto, S. Ethyl-eicosapentaenoic acid reduces liver lipids and lowers plasma levels of lipids in mice fed high-fat diet. In Vivo 2009, 23, 685–690. [Google Scholar]
- Kajikawa, S.; Harada, T.; Kawashima, A.; Imada, K.; Mizuguchi, K. Highly purified eicosapentaenoic acid prevents the progression of hepatic steatosis by repressing monounsaturated fatty acid synthesis in high-fat/high-sucrose diet-fed mice. Prostagland. Leukotr. Essent. Fatty Acids 2009, 80, 229–238. [Google Scholar] [CrossRef]
- Sano, T.; Tanaka, Y. Effects of dried powdered Chlorella vulgaris on experimental atherosclerosis and alimentary hypercholesterolemia in cholesterol-fed rabbit. Artery 1987, 14, 76–84. [Google Scholar]
- Tabernero, M.; Bermejo, M.L.; Elvira, M.; Caz, V.; Baeza, F.; Senorans , F.J.; de Miguel, E.; Largo, C. Metabolic effect of docosahexaenoic acid supplementation in different doses and formulations (ethyl- and glyceryl-) in hypercholesterolemic rats. J. Func. Foods 2013, in press. [Google Scholar]
- Yashodhara, B.M.; Umakanth, S.; Pappachan, J.M.; Bhat, S.K.; Kamath, R.; Choo, B.H. Omega-3 fatty acids: A comprehensive review of their role in health and disease. Postgrad. Med. J. 2009, 85, 84–90. [Google Scholar] [CrossRef]
- Lee, J.H.; O’Keefe, J.H.; Lavie, C.J.; Harris, W.S. Omega-3 fatty acids: Cardiovascular benefits, sources and sustainability. Nat. Rev. Cardiol. 2009, 6, 753–759. [Google Scholar] [CrossRef]
- Saremi, A.; Arora, R. The utility of omega-3 fatty acids in cardiovascular disease. Am. J. Ther. 2009, 16, 421–436. [Google Scholar] [CrossRef]
- Duda, M.K.; O’Shea, K.M.; Stanley, W.C. Omega-3 polyunsaturated fatty acid supplementattion for the treatment of heart failure: Mechanisms and clinical potential. Cardiovasc. Res. 2009, 84, 33–41. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, H.J.; Lee, H.J.; Kang, M.H.; Park, Y.K. Six-week supplementation with Chlorella has favorable impact on antioxidant status in Korean male smokers. Nutrition 2010, 26, 175–183. [Google Scholar] [CrossRef]
- Haimeur, A.; Ulmann, L.; Mimouni, V.; Guéno, F.; Pineau-Vincent, F.; Meskini, N.; Tremblin, G. The role of Odontella aurita, a marine diatom rich in EPA, as a dietary supplement in dyslipidemia, platelet function and oxidative stress in high-fat fed rats. Lipids Health Dis. 2012, 11, 147–160. [Google Scholar] [CrossRef]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 polyunsaturated fatty acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [CrossRef]
- Cherng, J.Y.; Shih, M.F. Preventing dyslipidemia by Chlorella pyrenoidosa in rat and hamsters after chronic high fat diet treatment. Life Sci. 2005, 76, 3001–3013. [Google Scholar] [CrossRef]
- Valdivielso, P.; Rioja, J.; Garcia-Aras, C.; Sanchez-Chaparro, M.A.; Gonzalez-Santos, P. Omega 3 fatty acids induce a marked reduction of apolipoprotein B48 when added to fluvastatin in patients with type 2 diabetes and mixed hyperlipidemia: A preliminary report. Cardiovasc. Diabetol. 2009, 8, 1–7. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P.; D’Angelo, A.; Salvaseo, S.A.; Ferrari, I.; Fogari, E.; Gravina, A.; Mereu, R.; Randazzo, S.; Cicero, A.F. Effects of long chain omega-3 fatty acids on metalloproteinases and their inhibitors in combined dyslipidemia patients. Expert Opin. Pharmacother. 2009, 10, 1239–1247. [Google Scholar] [CrossRef]
- Adan, Y.; Shibata, K.; Sato, M.; Ikeda, I.; Imaizumi, K. Effects of docosahexaenoic and eicosapentaenoic acid on lipid metabolism, eicosanoid production, platelet aggregation and atherosclerosis in hypercholesterolemic rats. Biosci. Biotechnol. Biochem. 1999, 63, 111–119. [Google Scholar] [CrossRef]
- Ryan, A.S.; Bailey-Hall, E.; Nelson, E.B.; Salem, N., Jr. The hypolipidemic effect of an ethyl ester of algal-docosahexaenoic acid in rats fed a high-fructose diet. Lipids 2009, 44, 817–826. [Google Scholar]
- Okudo, M.; Hasegawa, T.; Sonoda, M.; Okabe, T.; Tanaka, M. The effects of Chlorella on the level of cholesterol in serum and liver. Jpn. J. Nutr. 1975, 33, 3–8. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, F.; Liang, S.Z. Potential production of docosahexaenoic acid by the heterotrophic marine dinoflagellate Crypthecodinium cohnii. Proc. Biochem. 1999, 34, 633–637. [Google Scholar] [CrossRef]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Proc. Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Flickinger, B.D.; Matsuo, N. Nutritional characteristics of DAG Oil. Lipids 2003, 38, 129–132. [Google Scholar] [CrossRef]
- Tada, N.; Shoji, K.; Takeshita, M.; Watanabe, H.; Yoshida, H.; Hase, T.; Matsuo, N.; Tokimitsu, I. Effects of diacylglycerol ingestion on postprandial hyperlipidemia in diabetes. Clin. Chim. Acta 2005, 353, 87–94. [Google Scholar] [CrossRef]
- Takase, H.; Shoji, K.; Hase, T.; Tokimitsu, I. Effect of diacylglycerol on postprandial lipid metabolism in non-diabetic subjects with and without insulin resistance. Atherosclerosis 2005, 180, 197–204. [Google Scholar] [CrossRef]
- Yamamoto, K.; Takeshita, M.; Tokimitsu, I.; Watanabe, H.; Mizuno, T.; Asakawa, H.; Tokunaga, K.; Tatsumi, T.; Okazaki, M.; Yagi, N. Diacylglycerol oil ingestion in type 2 diabetic patients with hypertriglyceridemia. Nutrition 2006, 22, 23–29. [Google Scholar]
- Meng, X.; Zou, D.; Shi, Z.; Duan, Z.; Mao, Z. Dietary diacylglycerol prevents high-fat diet-induced lipid accumulation in rat liver and abdominal adipose tissue. Lipids 2004, 39, 37–41. [Google Scholar] [CrossRef]
- Kimura, S.; Tsuchiya, H.; Inage, H.; Meguro, S.; Matsuo, N.; Tokimitsu, I. Effects of dietary diacylglycerol on the energy metabolism. Int. J. Vitam. Nutr. Res. 2006, 76, 75–79. [Google Scholar] [CrossRef]
- Hayashi, H.; Tanaka, Y.; Hibino, H.; Umeda, Y.; Kawamitsu, H.; Fujimoto, H.; Amakawa, T. Beneficial effect of salmon roe phosphatidylcholine in chronic liver disease. Curr. Med. Res. Opin. 1999, 15, 177–184. [Google Scholar] [CrossRef]
- Shirai, N.; Higuchi, T.; Suzuki, H. Effect of lipids extracted from a salted herring roe food product on maze-behavior in mice. J. Nutr. Sci. Vitaminol. 2006, 52, 451–456. [Google Scholar] [CrossRef]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Docosahexaenoic acid and eicosapentaenoic acid-enriched phosphatidylcholine liposomes enhance the permeability, transportation and uptake of phospholipids in Caco-2 cells. Mol. Cell. Biochem. 2006, 285, 155–163. [Google Scholar] [CrossRef]
- Saito, H.; Kotani, Y.; Keriko, J.M..; Xue, C.; Taki, K.; Ishihara, K.; Ueda, T.; Myata, S. High levels of n-3 polyunsaturated fatty acids in Euphausia pacifica and its role as a source of docosahexaenoic and ecosapentaenoic acids for higher trophic levels. Mar. Chem. 2002, 78, 9–28. [Google Scholar] [CrossRef]
- Shirouchi, B.; Nagao, K.; Inoue, N.; Ohkubo, T.; Hibino, H.; Yanagita, T. Effect of dietary omega 3 phosphatidylcholine on obesity-related disorders in obese otsuka-long evans tokushima fatty rats. J. Agric. Food. Chem. 2007, 55, 7170–7176. [Google Scholar] [CrossRef]
- Lemaitre-Delaunay, D.; Laville, M.; Pachaiaudi, C.; Pousin, J.; Amstrong, M.; Lagarde, M. Blood compartmental metabolism of docosahexaenoic acid (DHA) in humans after ingestion of a single dose of [13C]DHA in phosphatidylcholine. J. Lipid Res. 1999, 40, 1867–1874. [Google Scholar]
- Akesson, B. Content of phospholipids in human diets studied by the duplicate-portion technique. Br. J. Nutr. 1982, 47, 223–229. [Google Scholar] [CrossRef]
- Ishinaga, M.; Sugiyama, S.; Mochizuki, T. Daily intake of fatty acids, sterols and phospholipids by Japanese women and serum cholesterol. J. Nutr. Sci. Vitaminol. 1994, 40, 557–567. [Google Scholar] [CrossRef]
- Murata, M.; Imaizumi, K.; Sugano, M. Effect of dietary phospholipids and their constituent bases on serum lipids and apolipoproteins in rats. J. Nutr. 1982, 112, 1805–1808. [Google Scholar]
- Imaizumi, K.; Mawatari, K.; Murata, M.; Ikeda, I.; Sugano, M. The contrasting effect of dietary phosphatidylethanolamine and phosphatidylcholine on serum lipoproteins and liver lipids in rats. J. Nutr. 1983, 113, 2403–2411. [Google Scholar]
- Jiang, Y.; Noh, S.K.; Koo, S.I. Egg phosphatidylcholine decreases the lymphatic absorption of cholesterol in rats. J. Nutr. 2001, 131, 2358–2363. [Google Scholar]
- Buang, Y.; Wang, Y.M.; Cha, J.Y.; Nagao, K.; Yanagita, T. Dietary phosphatidylcholine alleviates fatty liver induce by orotic acid. Nutrition 2005, 21, 867–873. [Google Scholar] [CrossRef]
- Stamler, C.J.; Breznan, D.; Neville, T.A.; Viau, F.J.; Camlioglu, E.; Sparks, D.L. Phosphatidylinositol promotes cholesterol transport in vivo. J. Lipid Res. 2000, 41, 1214–1221. [Google Scholar]
- Burgess, J.W.; Boucher, J.; Neville, T.A.; Rouillard, P.; Stamler, C.; Zachariah, S.; Sparks, D.L. Phosphatidylinositol promotes cholesterol transport and excretion. J. Lipid Res. 2003, 44, 1355–1363. [Google Scholar] [CrossRef]
- Burgess, J.W.; Neville, T.A.; Rouillard, P.; Harder, Z.; Beanlands, D.S.; Sparks, D.L. Phosphatidylinositol increases HDL-C levels in humans. J. Lipid Res. 2005, 46, 350–355. [Google Scholar]
- Thompson, P.A.; Bonham, P.; Waite, A.M.; Clementson, L.A.; Cherukuru, N.; Hassler, C.; Doblin, M.A. Contrasting oceanographic conditions and phytoplankton communities on the east and west coasts of Australia. Deep Sea Res. Part II 2011, 58, 645–663. [Google Scholar] [CrossRef]
- Vilchez, C.; Forjan, E.; Cuaresma, M.; Bedmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: Biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef]
- Di Valentin, M.; Büchel, M.; Giacometti, G.M.; Carbonera, D. Chlorophyll triplet quenching by fucoxanthin in the fucoxanthin-chlorophyll protein from the diatom Cyclotella meneghiniana. Biochem. Biophys. Res. Commun. 2012, 427, 637–641. [Google Scholar] [CrossRef]
- Spikes, J.D.; Bommer, J.C. Chlorophyll and Related Pigments as Photosensitizers in Biological Medicine. In Chlorophylls; Scheer, H., Ed.; CRC Press: Boca Raton, FL, USA, 1991; pp. 1024–1181. [Google Scholar]
- Tripathy, B.; Chakraborty, N. 5-Aminolevulinic acid induced photodynamic damage of the photosynthetic electron transport chain of cucumber (Cucumis sativus L) cotyledons. Plant Physiol. 1991, 96, 761–767. [Google Scholar] [CrossRef]
- Rebeiz, C.A.; Nandihalli, U.B.; Reddy, K. Photodynamic Herbicides and Chlorophyll Biosynthesis Inhibitors. In Herbicides; Baker, N.R., Percival, M., Eds.; Elsevier: Amsterdam, the Netherlands, 1995; pp. 173–208. [Google Scholar]
- Ricchelli, F. Photophysical properties of porphyrins in biological membranes. J. Photochem. Photobiol. B 1995, 29, 109–118. [Google Scholar] [CrossRef]
- Ebermann, R.; Alth, G.; Kreitner, M.; Kubin, A. Natural products derived from plants and potential drugs for the photodynamic destruction of tumor cells. J. Photochem. Photobiol. B 1996, 36, 956–697. [Google Scholar]
- Solymosi, K.; Schoefs, B. Etioplast and etio-chloroplast formation under natural conditions: The Dark side of chlorophyll biosynthesis in angiosperms. Photosynth. Res. 2010, 105, 145–166. [Google Scholar]
- Czarnecki, O.; Grimm, B. Post-translational control of tetrapyrrole biosynthesis in plants, algae, and cyanobacteria. J. Exp. Bot. 2012, 63, 1675–1687. [Google Scholar] [CrossRef]
- Murthy, K.; Vanitha, A.; Rajesha, J.; Swamy, M.; Sowmya, P.; Ranishankar, G.A. In vivo antioxidant activity of carotenoids from Dunalliela salina, a green microalga. Life Sci. 2005, 76, 1381–1390. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Gressel, J.; Avron, M. Massive accumulation of phytoene induced by norflurazon in Dunaliella bardawil (Chlorophyceae) prevents recovery from photoinhibition. J. Phycol. 1987, 23, 176–181. [Google Scholar]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylidès, C.; Havaux, M. Carotenoid oxidation products are stress signals that mediate gene responses to singlet oxygen in plants. Proc. Natl. Acad. Sci.USA 2012, 109, 5535–5540. [Google Scholar]
- Tardy, F.; Havaux, M. Photosynthesis, chlorophyll fluorescence, light-harvesting system and photoinhibition resistance of a zeaxanthin-accumulating mutant of Arabidopsis thaliana. J. Photochem. Photobiol. 1996, 34, 87–94. [Google Scholar] [CrossRef]
- Lohr, M.; Schwender, J.; Polle, J.E.W. Isoprenoid biosynthesis in eukaryotic phototrophs: A spotlight on algae. Plant Sci. 2012, 185–186, 9–22. [Google Scholar] [CrossRef]
- Kana, T.M.; Glibert, P.M.; Goericke, R.; Welschmeyer, N.A. Zeaxanthin and β-carotene in Synechococcus WH7803 respond differently to irradiance. Limnol. Oceanogr. 1998, 33, 1623–1627. [Google Scholar]
- Masamoto, K.; Zsiros, O.; Gombos, Z. Accumulation of zeaxanthin in cytoplasmic membranes of the cyanobacterium Synechococcus sp. strain PCC 7942 grown under high light condition. J. Plant Physiol. 1999, 155, 136–138. [Google Scholar]
- Pfannschmidt, T. Plastidial retrograde signalling—A true plastid factor or just metabolite signatures? Trends Plant Sci. 2010, 15, 427–435. [Google Scholar]
- Foyer, C.H.; Neukermans, J.; Queval, G.; Noctor, G.; Harbinson, J. Photosynthetic control of electron transport and the regulation of gene expression. J. Exp. Bot. 2012, 63, 1637–1661. [Google Scholar] [CrossRef]
- Coesel, S.; Obornik, M.; Valera, J.; Falciatore, A.; Bowler, C. Evolutionary origins and functions of the carotenoids biosynthetic pathway in marine diatoms. PloS One 2008, 3, e2896. [Google Scholar] [CrossRef]
- Frommolt, R.; Werner, S.; Paulsen, H.; Goss, R.; Wilhem, C.; Zauner, S.; Maier, U.G.; Grossman, A.R.; Bhattacharya, D.; Lohr, M. Ancient recruitment by chromists of green algal genes encoding enzymes for carotenoid biosynthesis. Mol. Biol. Evol. 2008, 25, 2653–2667. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Biosynthesis, accumulation and emission of carotenoids, alpha-tocopherol, plastoquinone, and isoprene in leaves under high potosynthetic irradiance. Photosynth. Res. 2007, 92, 163–179. [Google Scholar] [CrossRef]
- Klodawska, K.; Malec, P.; Kis, M.; Gombos, Z.; Strzalka, K. EPR study of thylakoid membrane dynamics in mutants of the carotenoid biosynthesis pathway of Synechocystis sp. PCC6803*. Acta Biochim. Pol. 2012, 59, 87–90. [Google Scholar]
- Mouget, J.L.; Tremblin, G.; Morant-Manceau, A.; Morancais, M.; Robert, J.M. Long-tterm photoacclimation of Haslea ostrearia (Bacillariophyta): Effect of irradiance on growth rates, pigment content and photosynthesis. Eur. J. Phycol. 1999, 34, 109–115. [Google Scholar]
- Kulk, G.; van de Poll, W.H.; Visser, R.J.W.; Buma, A.G.J. Distinct differences in photoacclimation potential between prokaryotic and eukaryotic oceanic phytoplankton. J. Exp. Mar. Biol. Ecol. 2011, 398, 63–72. [Google Scholar] [CrossRef]
- Pattanaik, B.; Whitaker, M.J.; Montgomery, B.L. Light quantity affects the regulation of cell shape in Fremyella diplosiphon. Front Microbiol. 2012, 3, 170–175. [Google Scholar]
- Demers, S.; Roy, S.; Gognon, R.; Vignault, C. Rapid light induced changes in cell fluorescence 986 and in xanthophylls-cycle pigments of Alexandrium excavatum (Dinophyceae) and Thalassiosira pseudonana (Bacillaryophyceae): A photoprotection mechanism. Mar. Ecol. Prog. Ser. 1991, 76, 185–196. [Google Scholar] [CrossRef]
- Recht, M.; Mouget, J.L.; Morant-Manceau, A.; Rosa, P.; Tremblin, G. Long-term acclimation to UV radiation: Effects on growth, photosynthesis and carbonic anhydrase activity in marine diatoms. Bot. Mar. 2005, 48, 407–420. [Google Scholar]
- Rijstenbil, J.W. UV- and salinity-induced oxidative effects in the marine diatom Cylindrotheca closterium during simulated emersion. Mar. Biol. 2005, 147, 1063–1073. [Google Scholar] [CrossRef]
- Huang, J.J.; Chueng, P.C.K. +UVA treatment increases the degree of insaturation in microalga fatty acids and total carotenoid content in Nitschia closterium (Bacillariophyceae) and Isochrysis zhangjiangensis (Chrysophyceae). Food Chem. 2011, 129, 783–791. [Google Scholar] [CrossRef]
- Shpilyov, A.V.; Zinchenko, V.V.; Grimm, B.; Lokstein, H. Chlorophyll a phytylation is required for the stability of photosystems I and II in the cyanobacterium Synechocystis sp. PCC 6803. Plant J. 2013, 73, 336–346. [Google Scholar]
- Kepekçi, R.A.; Saygideger, S.D. Enhancement of phenolic compound production in Spirulina platensis by two-step batch mode cultivation. J. Appl. Phycol. 2012, 24, 897–905. [Google Scholar] [CrossRef]
- Recht, M.; Morant-Manceau, A.; Tremblin, G. Carbon fixation and anhydrase carbonic activity in Haslea ostrearia (Bacillariophyceae) in relation to growth irradiance. Photosynthetica 2008, 46, 56–62. [Google Scholar] [CrossRef]
- Bhandari, R.; Sharma, K.P. High-light-induced changes on photosynthesis, pigments, sugars, lipids and antioxidant enzymes in freshwater (Nostoc spongiaeforme) and marine (Phormidium corium) cyanobacteria. Photochem. Photobiol. 2006, 82, 702–710. [Google Scholar] [CrossRef]
- Li, Z.; Ma, X.; Zhang, C. A novel potential source of β-carotene: Eustigmatos cf. polyphem (Eustigmatophyceae) and pilot β-carotene production in bubble column and flat panel photobioreactors. Biores. Technol. 2012, 117, 257–263. [Google Scholar] [CrossRef]
- Lamers, P.P.; Janssen, M.; de Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in nitrogen-starved Dunalliela salina, a unicellular green microalga. J. Biotechnol. 2012, 162, 21–27. [Google Scholar] [CrossRef]
- Simionato, D.; Block, M.A.; la Rocca, N.; Jouhet, J.; Maréchal, E.; Finazzi, G.; Morosinotto, T. Response of Nannochloropsis gaditana to nitrogen starvation includes a de novo biosynthesis of triacylglycerols, a decrease of chloroplast galactolipids and a reorganization of the photosynthetic apparatus. Eukarot. Cell 2013. [Google Scholar] [CrossRef]
- Riyahi, J.; Haouazine, Y.; Akallal, R.; Mouradi, A.; Creach, A.; Givernaud, R.; Mouradi, A. Influence des nitrates, de la salinité et du stress lumineux sur la teneur en acides gras et en β-carotène de Dunaliella salina. (in French). Bull. Soc. Pharm. Bordeaux 2007, 146, 235–250. [Google Scholar]
- Saxena, P.; Bassi, A. Removal nutrient from hydroponic greenhouse effluent by alkali precipitation and algae cultivation method. J. Chem. Technol. Biotechnol. 2013, 88, 858–863. [Google Scholar] [CrossRef]
- Lemoine, Y.; Schoefs, B. Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef]
- Kim, D.K.; Hong, S.J.; Bae, J.H.; Yim, N.; Jin, E.S.; Lee, C.G. Transcriptomic analysis of Haematococcus lacustris during astaxanthin accumulation under high irradiance and nutrient starvation. Biotech. Bioprocess. Eng. 2011, 16, 698–705. [Google Scholar] [CrossRef]
- Martin, F. Optimization of Photobioreactor for Astaxanthin Production in Chlorella zofingiensis. M.Sc. Thesis, National University of Singapore, Singapore, 2011. [Google Scholar]
- Wayama, M.; Ota, S.; Matsuura, H.; Nango, N.; Hirata, A.; Kawano, S. Three-dimensional ultrastructural study of oil and astaxanthin accumulation during encystment in the green alga Haematococcus pluvialis. PloS One 2013, 8, e53618. [Google Scholar]
- Milledge, J.J. Commercial application of microalgae other than as biofuels: A brief review. Rev. Environ. Sci. Biotechnol. 2011, 10, 31–41. [Google Scholar] [CrossRef]
- Singh, S.; Kate, B.N.; Banecjee, U.C. Bioactive compounds from cyanobacteria and microalgae: An overview. Crit. Rev. Biotechnol. 2005, 25, 73–95. [Google Scholar] [CrossRef]
- Nikkinen, H.-L.; Hakkila, K.; Gunnelius, L.; Huokko, T.; Pollari, M.; Tyystjärvi, T. The SigB σ factor regulates multiple salt acclimation responses of the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2012, 158, 514–523. [Google Scholar]
- Bano, F.; Zutshi, S.; Fatma, T. Chromium (VI) induced oxidative stress in Hapalosiphon fontinalis. World J. Microbiol. Biotechnol. 2012, 28, 2505–2511. [Google Scholar] [CrossRef]
- Singh, P.; Tiwari, A.; Singh, S.P.; Asthana, R.K. Desiccation induced changes in osmolytes production and the antioxidative defense in the cyanobacterium Anabaena sp. PCC 7120. Physiol. Mol. Biol. Plants 2013, 19, 61–68. [Google Scholar]
- Jiang, H.; Gao, K. Effects of lowering temperature during culture on the production of polyunsaturated fatty acids in the marine diatom Phaeodactylum tricornutum (Bacillariophyceae). J. Phycol. 2004, 40, 651–654. [Google Scholar] [CrossRef]
- Zhu, C.J.; Lee, Y.K.; Chao, T.M. Effects of temperature and growth phase on lipid and biochemical composition of Isochrysis galbana TK1. J. Appl. Phycol. 1997, 9, 451–457. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.V.; Lambrinidis, G.; Parry, D.L. Effect of temperature on growth, chemical composition and fatty acid composition of tropical australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- De Castro Araujo, S.; Tavano Garcia, V.M. Growth and biochemical composition of the diatom Chaetoceros cf. wighamii brightwell under different temperature, salinity and carbon dioxide levels. I. Protein, carbohydrates and lipids. Aquaculture 2005, 246, 405–412. [Google Scholar] [CrossRef]
- Rousch, J.M.; Bingham, S.E.; Sommerfeld, M.R. Changes in fatty acid profiles of thermo-intolerant and thermo-tolerant marine diatoms during temperature stress. J. Exp. Mar. Biol. Ecol. 2003, 295, 145–156. [Google Scholar] [CrossRef]
- Gacheva, G.; Gigova, L.; Ivanova, N.; Iliev, I.; Toshkova, R.; Gardeva, E.; Kussovski, V.; Najdenski, H. Suboptimal growth temperatures enhance the biological activity of cultured cyanobacterium Gloeocapsa sp. J. Appl. Phycol. 2013, 25, 183–194. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Mimouni, V.; Ulmann, L.; Tremblin, G. Combined effects of irradiance level and carbon source on fatty acid and lipid class composition in the microalga Pavlova lutheri commonly used in mariculture. J. Exp. Mar. Biol. Ecol. 2009, 369, 136–143. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Ulmann, L.; Mimouni, V.; Tremblin, G. Use of radiolabelled substrates to determine the desaturase and elongase activities involved in eicosapentaenoic acid and docosahexaenoic acid biosynthesis in the marine microalga Pavlova lutheri. Phytochemistry 2013, 90, 43–49. [Google Scholar]
- Guiheneuf, F.; Mimouni, V.; Ulmann, L.; Tremblin, G. Environmental factors affecting growth and omega 3 fatty acid composition in Skeletonema costatum. The influences of irradiance and carbon source. Diatom Res. 2008, 23, 93–103. [Google Scholar]
- Fabregas, J.; Maseda, A.; Dominquez, A.; Otero, A. The cell composition of Nannochloropsis sp. Changes Under different irradiances in semicontinuous culture. World J. Microbiol. Biotechnol. 2004, 20, 31–35. [Google Scholar]
- Manoharan, K.; Lee, T.K.; Cha, J.M.; Kim, J.H.; Lee, W.S.; Chang, M.; Park, C.W.; Cho, J.H. Acclimation of Prorocentrum minimum (Dinophyceae) to prolonged darkness by use of an alternative carbon source from triacylglycerides and galactolipids. J. Phycol. 1999, 35, 287–292. [Google Scholar] [CrossRef]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty acid and lipid content of marine microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar]
- Alonso, D.L.; Belarbi, E.H.; Fernández-Sevilla, J.M.; Rodríguez-Ruiz, J.; Grima, E.M. Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum tricornutum. Phytochemistry 2000, 54, 461–471. [Google Scholar]
- Muradyan, E.A.; Klyachko-Gurvich, G.L.; Tsoglin, L.N.; Sergeyenko, T.V.; Pronina, N.A. Changes in lipid metabolism during adaptation of the Dunaliella salina photosynthetic apparatus to high CO2 concentration. Russ. J. Plant Physiol. 2004, 51, 53–62. [Google Scholar] [CrossRef]
- Azachi, M.; Sadka, A.; Fisher, M.; Goldshlag, P.; Gokhman, I.; Zamir, A. Salt Induction of fatty acid elongase and membrane lipid modifications in the extreme halotolerant alga Dunaliella salina. Plant Physiol. 2002, 129, 1320–1329. [Google Scholar] [CrossRef]
- Thompson, G.A., Jr. Lipids and membrane function in green Algae. Biochim. Biophys. Acta 1996, 1302, 17–45. [Google Scholar]
- Cho, S.H.; Thompson, G.A., Jr. On the metabolic relationships between monogalactosyldiacylglycerol and digalactosyldiacylglycerol molecular species in Dunaliella salina. J. Biol. Chem. 1987, 262, 7586–7593. [Google Scholar]
- Sato, N.; Seyama, Y.; Murata, N. Lipid-linked desaturation of palmitic acid in monogalactosyl diacylglycerol in the blue-green alga (Cyanobacterium) Anabaena variabilis studied in vivo. Plant Cell. Physiol. 1986, 27, 819–835. [Google Scholar]
- Domergue, F.; Spiekermann, P.; Lerchl, J.; Beckmann, C.; Kilian, O.; Kroth, P.G.; Boland, W.; Zähringer, U.; Heinz, E. New insight into Phaeodactylum tricornutum fatty acid metabolism. Cloning and functional characterization of plastidial and microsomal δ12-fatty acid desaturases. Plant Physiol. 2003, 131, 1648–1660. [Google Scholar] [CrossRef]
- Eichenberger, W.; Gribi, C. Lipids of Pavlova lutheri: Cellular site and metabolic role of DGCC. Phytochemistry 1997, 8, 1561–1567. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Ulmann, L.; Tremblin, G.; Mimouni, V. Light-dependent utilization of two radiolabelled carbon sources, sodium bicarbonate and sodium acetate. Relationships with the long chain polyunsaturated fatty acid synthesis in the microalga Pavlova lutheri (Haptophyta). Eur. J. Phycol. 2011, 46, 143–152. [Google Scholar] [CrossRef]
- Meireles, L.A.; Guedes, A.C.; Malcata, F.X. Lipid class composition of the microalga Pavlova lutheri: Eicosapentaenoic and docosahexaenoic acids. J. Agric. Food Chem. 2003, 51, 2237–2241. [Google Scholar] [CrossRef]
- Khozin, I.; Adlerstein, D.; Bigogno, C.; Heimer, Y.M.; Cohen, Z. Elucidation of the biosynthesis of eicosapentaenoic acid in the microalga Porphyridium cruentum. 2. Studies with radiolabeled precursors. Plant Physiol. 1997, 114, 223–230. [Google Scholar]
- Khozin-Goldberg, I.; Bigogno, C.; Cohen, Z. Salicylhydroxamic acid inhibits delta 6 desturation in the microalga Porphyridium cruentum. Biol. Lipids 1999, 1439, 384–394. [Google Scholar]
- Khozin-Goldberg, I.; Didi-Cohen, S.; Shayakhmetova, I.; Cohen, Z. Biosynthesis of eicosapentaenoic acid (EPA) in the freshwater eustigmatophyte Monodus subterraneus (Eustigmatophyceae). J. Phycol. 2002, 38, 745–756. [Google Scholar] [CrossRef]
- Schneider, J.C.; Livne, A.; Sukenik, A. A mutant of Nannochloropsis deficient in eicosapentaenoic acid production. Phytochemistry 1995, 40, 807–814. [Google Scholar] [CrossRef]
- Haigh, W.G.; Yoder, T.F.; Ericson, L.; Pratum, T.; Winget, R.R. The characterisation and cyclic production of a highly unsaturated homoserine lipid in Chlorella minutissima. Biochim. Biophys. Acta 1996, 1299, 183–190. [Google Scholar]
- Khozin-Goldberg, I.; Yu, H.Z.; Adlerstein, D.; Didi-Cohen, S.; Heimer, Y.M.; Cohen, Z. Triacylglycerols of the red microalga Porphyridium cruentum can contribute to the biosynthesis of eukaryotic galactolipids. Lipids 2000, 35, 881–889. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; de Rijk, T.C.; Eggink, G.; Sijtsma, L. Optimisation of docosahexaenoic acid production in batch cultivation by Crypthecodinium cohnii. J. Biotechnol. 1999, 70, 185–192. [Google Scholar] [CrossRef]
- Olaizola, M. Commercial development of microalgal biotechnology: From the test tube to the market place. Biomol. Eng. 2003, 20, 459–466. [Google Scholar] [CrossRef]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Chen, C.Y.; Kao, P.C.; Tsai, C.J.; Lee, D.J.; Chang, J.S. Engineering strategies for simultaneous enhancement of C-phycocyanin production and CO2 fixation with Spirulina platensis. Bioresour. Technol. 2013. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Fernandez Sevilla, J.M.; Molina Grima, E.; Perez Parra, J.; Acien Fernandes, F.G.; Magan, C.J.J.; Friedl, T. Novel microalgal species and their use for animal and/or human consumption and in the production of carotenoids. European Patent EP1849858, 21 September 2011. [Google Scholar]
- Blackburn, S.I.; Volkman, J.K. Microalgae: A Renewable Source of Bioproducts. In Food and Industrial Bioproducts and Bioprocessing; Dunford, N.T., Ed.; Wiley-Blackwell: New York, NY, USA, 2012; pp. 221–241. [Google Scholar]
- Kopecky, J.; Stys, D.; Loetz, K.; Poulz, O.; Schoefs, B. Microalgae as a source for secondary carotenoid production. A screening study. Arch. Hydrobiol. Algol. Stud. 2000, 98, 153–167. [Google Scholar]
- Bourre, J.M. Acides gras oméga-3, structures et fonctions du cerveau. (in French). Rev. Neurol. 2007, 163, 243–244. [Google Scholar] [CrossRef]
- Gong, Y.; Hu, H.; Gao, Y.; Xu, X.; Gao, H. Microalgae as platforms for production of recombinant proteins and valuable compounds: Progress and prospects. J. Ind. Microbiol. Biotechnol. 2011, 38, 1879–1890. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, F.; Benavides, J.; Ritopalomares, M. Scaling-up of a B-phycoerythrin production and purification bioprocess involving aqueous two-phase systems: Practical experiences. Process Biochem. 2013, 48, 738–745. [Google Scholar] [CrossRef]
- Paniagua-Michel, J.; Olmos-Soto, J.; Acosta-Ruiz, M. Pathways of carotenoid biosynthesis in bacteria and microalgae. Methods Mol. Biol. 2012, 892, 1–12. [Google Scholar] [CrossRef]
- Huang, J.; Zhong, Y.; Sandmann, G.; Liu, J.; Chen, F. Cloning and selection of carotenoid ketolase genes for the engineering of high-yield astaxanthin in plants. Planta 2012, 236, 691–699. [Google Scholar] [CrossRef]
- Blatti, J.L.; Michaud, J.; Burkart, M.D. Engineering fatty acid biosynthesis in microalgae for sustainable biodiesel. Curr. Opin. Chem. Biol. 2013, 17, 496–505. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Heydarizadeh, P.; Poirier, I.; Loizeau, D.; Ulmann, L.; Mimouni, V.; Schoefs, B.; Bertrand, M. Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids. Mar. Drugs 2013, 11, 3425-3471. https://doi.org/10.3390/md11093425
Heydarizadeh P, Poirier I, Loizeau D, Ulmann L, Mimouni V, Schoefs B, Bertrand M. Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids. Marine Drugs. 2013; 11(9):3425-3471. https://doi.org/10.3390/md11093425
Chicago/Turabian StyleHeydarizadeh, Parisa, Isabelle Poirier, Damien Loizeau, Lionel Ulmann, Virginie Mimouni, Benoît Schoefs, and Martine Bertrand. 2013. "Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids" Marine Drugs 11, no. 9: 3425-3471. https://doi.org/10.3390/md11093425
APA StyleHeydarizadeh, P., Poirier, I., Loizeau, D., Ulmann, L., Mimouni, V., Schoefs, B., & Bertrand, M. (2013). Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids. Marine Drugs, 11(9), 3425-3471. https://doi.org/10.3390/md11093425