2.1. Amino Acid Sequence of BSL Isoforms
The pure lectin active fraction was obtained by ion-exchange chromatography as described by Ainouz and co-worker. The hemagglutinating activity of the lectin was inhibited only by the glycoproteins fetuin, avidin and porcine mucin [
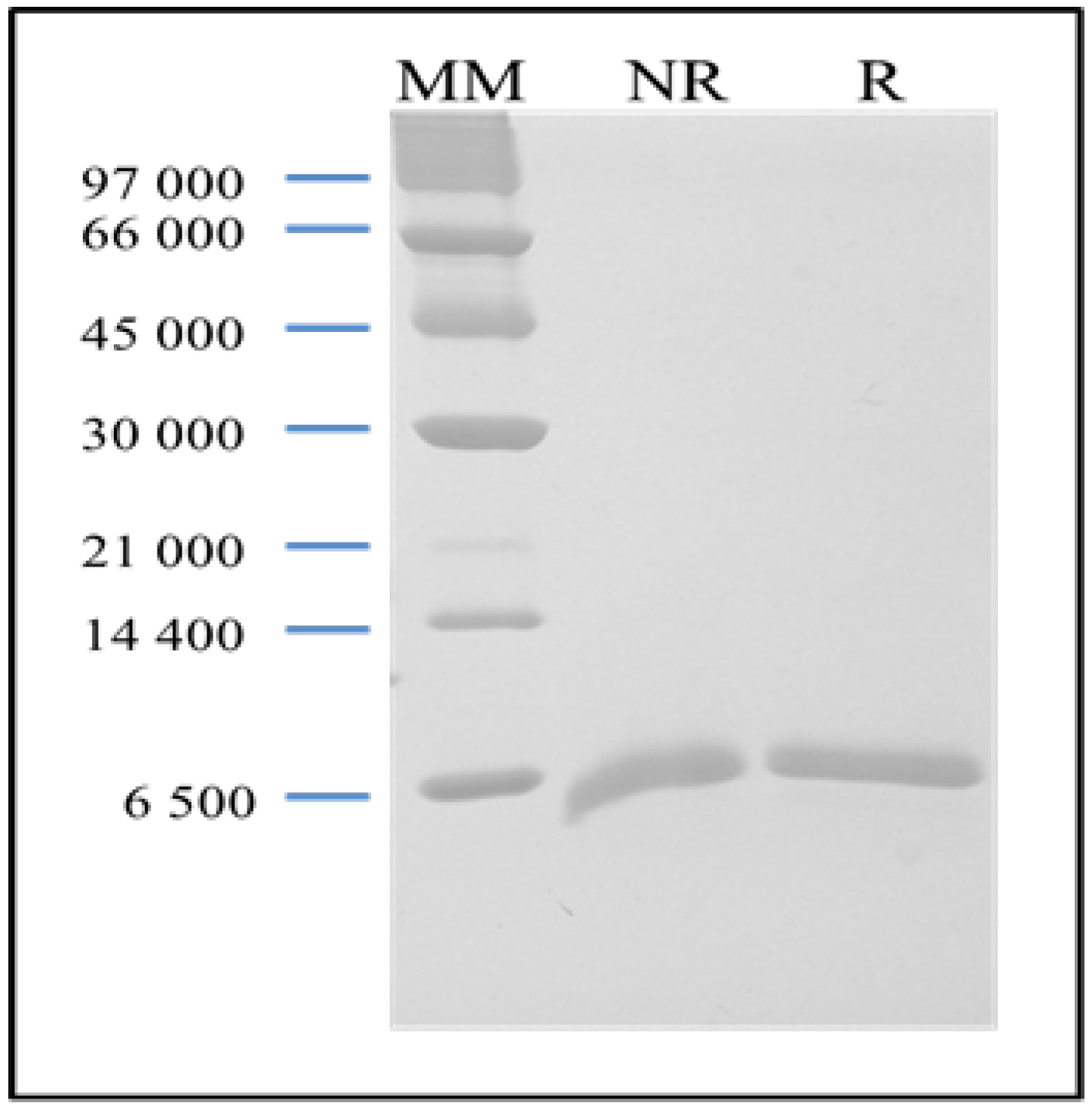
18]. The purified BSL is observed in SDS-PAGE as a broad band with an apparent molecular mass of 9 kDa in the presence and absence of 2-mercaptoethanol (
Figure 1).
Figure 1.
SDS-PAGE of purified BSL. Lane MM: a marker kit including phosphorylase B (97 kDa), BSA (66 kDa), ovalbumin (45 kDa), carbonic anhydrase (30 kDa), trypsinogen (21 kDa), lactalbumin (14.4 kDa) and aprotinin (6.5 kDa). Lane NR: non-reduced BSL. Lane R: reduced BSL.
Figure 1.
SDS-PAGE of purified BSL. Lane MM: a marker kit including phosphorylase B (97 kDa), BSA (66 kDa), ovalbumin (45 kDa), carbonic anhydrase (30 kDa), trypsinogen (21 kDa), lactalbumin (14.4 kDa) and aprotinin (6.5 kDa). Lane NR: non-reduced BSL. Lane R: reduced BSL.
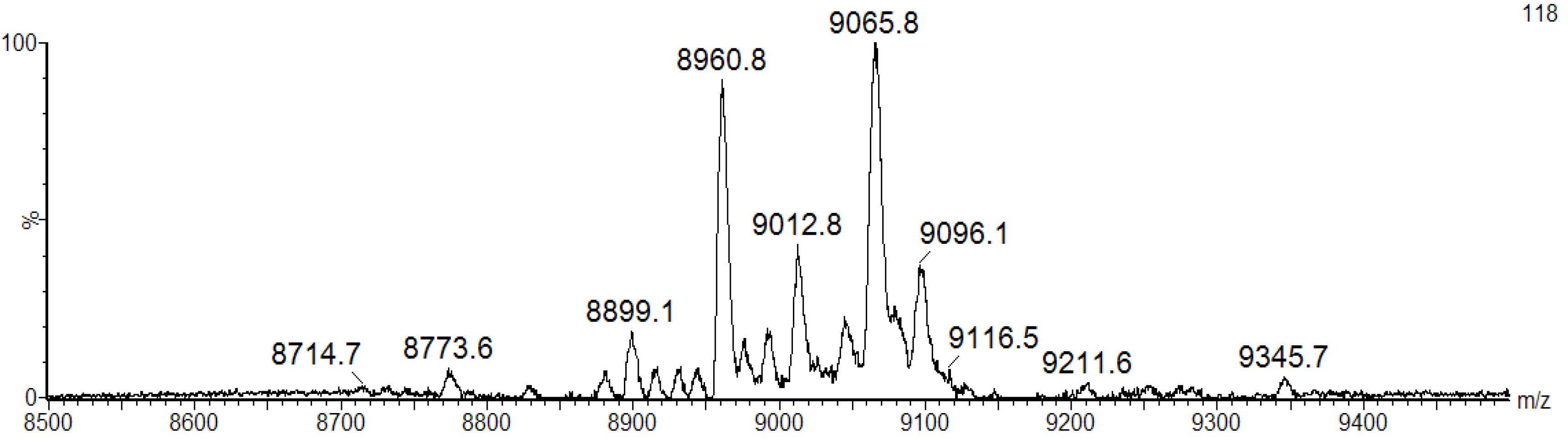
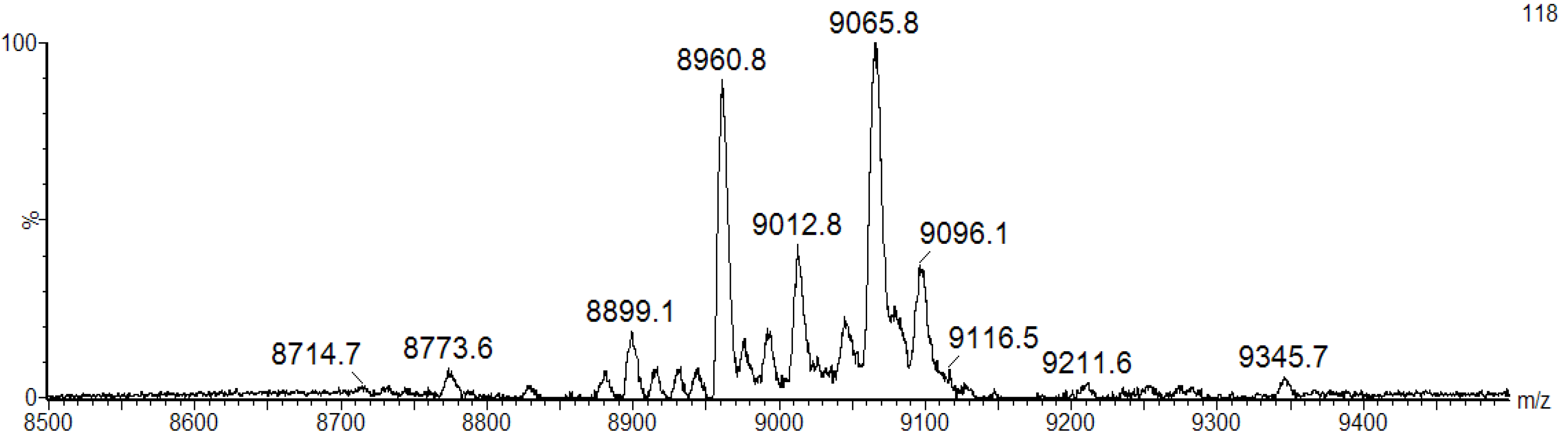
We speculate that the broad band pattern could be due to the presence of isomeric forms of BSL with different molecular mass. To confirm the isoforms, the purified BSL was investigated by matrix-assisted laser desorption ionization mass spectrometry (MALDI-MS). The mass spectrometric analysis revealed that the purified BSL contained a mixture of five isoforms, namely: BSL1, BSL2, BSL3, BSL4 and BSL5 at
m/z 8898.1 ± 2, 8960.8 ± 2, 9012.8 ± 2, 9065.8 ± 2 and 9096.1 ± 2, respectively (
Figure 2).
The existence of lectin isoforms was also observed in the lectins isolated from
Bryothamnion triquetrum [
19],
Eucheuma serra [
20] and
Hypnea japonica [
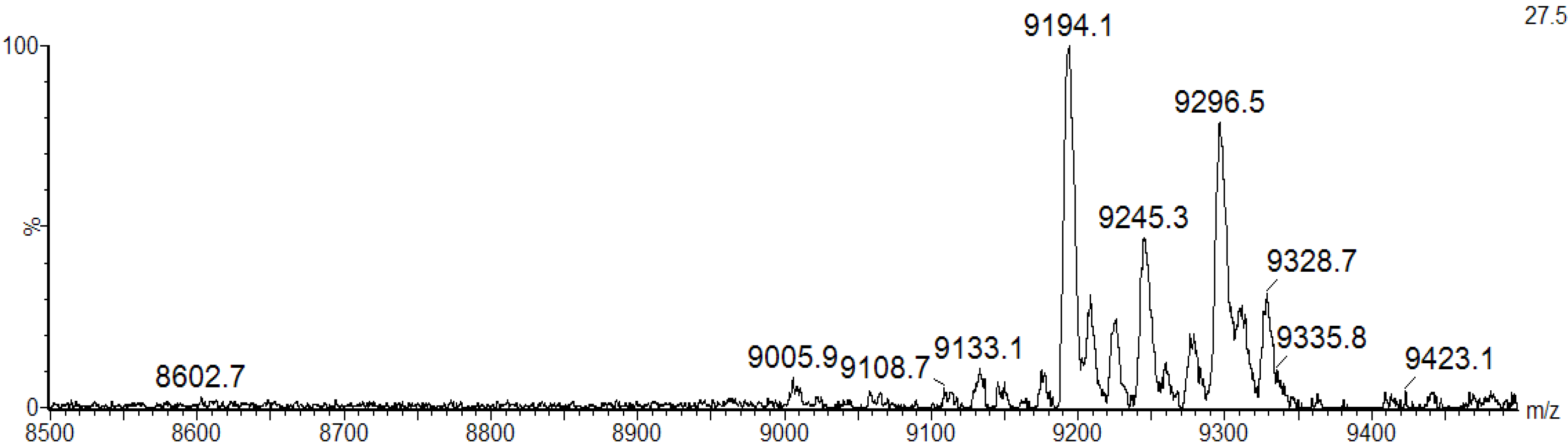
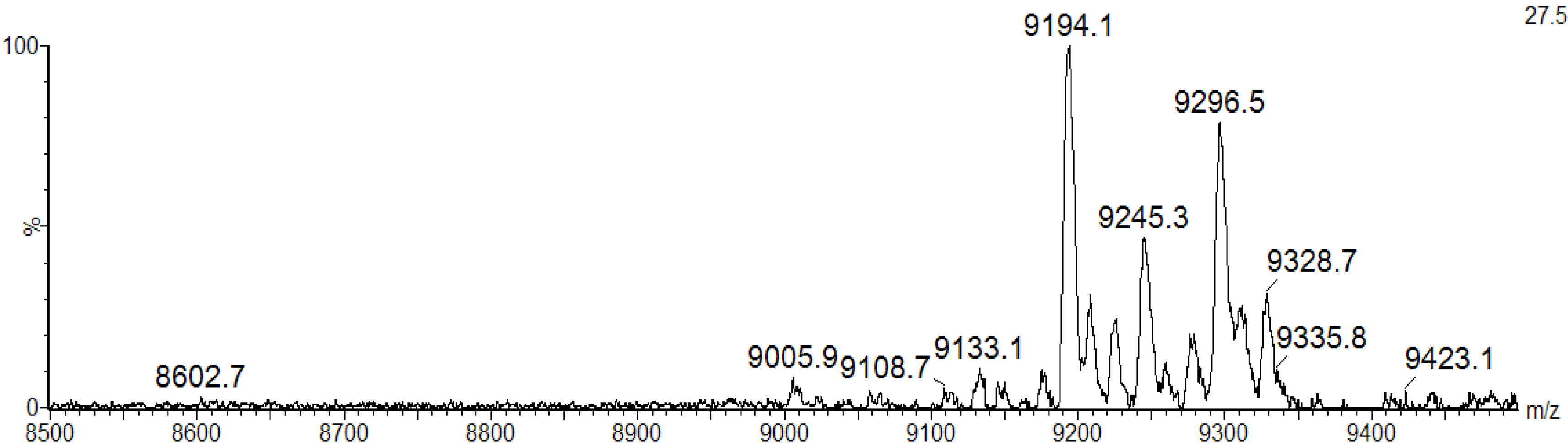
21]. The MALDI-TOF masses of intact BSL did not change upon incubation of the lectin with iodoacetamide under non-reducing conditions, indicating that BSL does not possess free sulfhydryl groups (data not shown). In contrast, a MALDI-TOF-MS analysis of denatured, reduced and carbamidomethylated BSL (CA-BSL) exhibited five major ions at
m/z 9133.1 ± 2, 9194.1 ± 2, 9245.3 ± 2, 9296.5 ± 2 and 9328.7 ± 2, indicating the presence of four cysteine residues involved in two intrachain disulfide bonds (
Figure 3).
Figure 2.
MALDI-TOF-MS analysis of BSL under denaturing conditions. The BSL isoforms exhibit major ion peaks at m/z 9065.8 ± 2 and 8960.8 ± 2 and three ion peaks with minor intensity at m/z 8899.1 ± 2, 9012.8 ± 2 and 9096.1 ± 2.
Figure 2.
MALDI-TOF-MS analysis of BSL under denaturing conditions. The BSL isoforms exhibit major ion peaks at m/z 9065.8 ± 2 and 8960.8 ± 2 and three ion peaks with minor intensity at m/z 8899.1 ± 2, 9012.8 ± 2 and 9096.1 ± 2.
Figure 3.
MALDI-TOF-MS analysis of CA-BSL under denaturing and reducing conditions in the presence of iodoacetamide. The BSL samples treated with dithiothreitol (DTT) and iodoacetamide exhibited increased masses at 232 Daltons (4 × 58.00), suggesting the presence of four cysteine residues engaged in the formation of two intrachain disulfide bonds.
Figure 3.
MALDI-TOF-MS analysis of CA-BSL under denaturing and reducing conditions in the presence of iodoacetamide. The BSL samples treated with dithiothreitol (DTT) and iodoacetamide exhibited increased masses at 232 Daltons (4 × 58.00), suggesting the presence of four cysteine residues engaged in the formation of two intrachain disulfide bonds.
These data were confirmed by an analysis of reduced and ethylpyridylated BSL in which the mass increment was 436 Daltons to each native BSL isoform (data not shown). Similar results were obtained for the
Bryothamnion triquetrum lectin (BTL) and the
Hypnea japonica lectin (HJA) [
19,
21]. HML and HCA, lectins isolated from the marine red alga
Hypnea musciformis and
Hypnea cervicornis, respectively, have masses of 9 kDa and contain 14 half-cystines each in their amino acid sequences. Furthermore, they are composed of two similar polypeptide chains linked by S–S bonds [
13]. The presence of cysteinyl residues in BSL lectins may contribute to their extreme thermostability [
22].
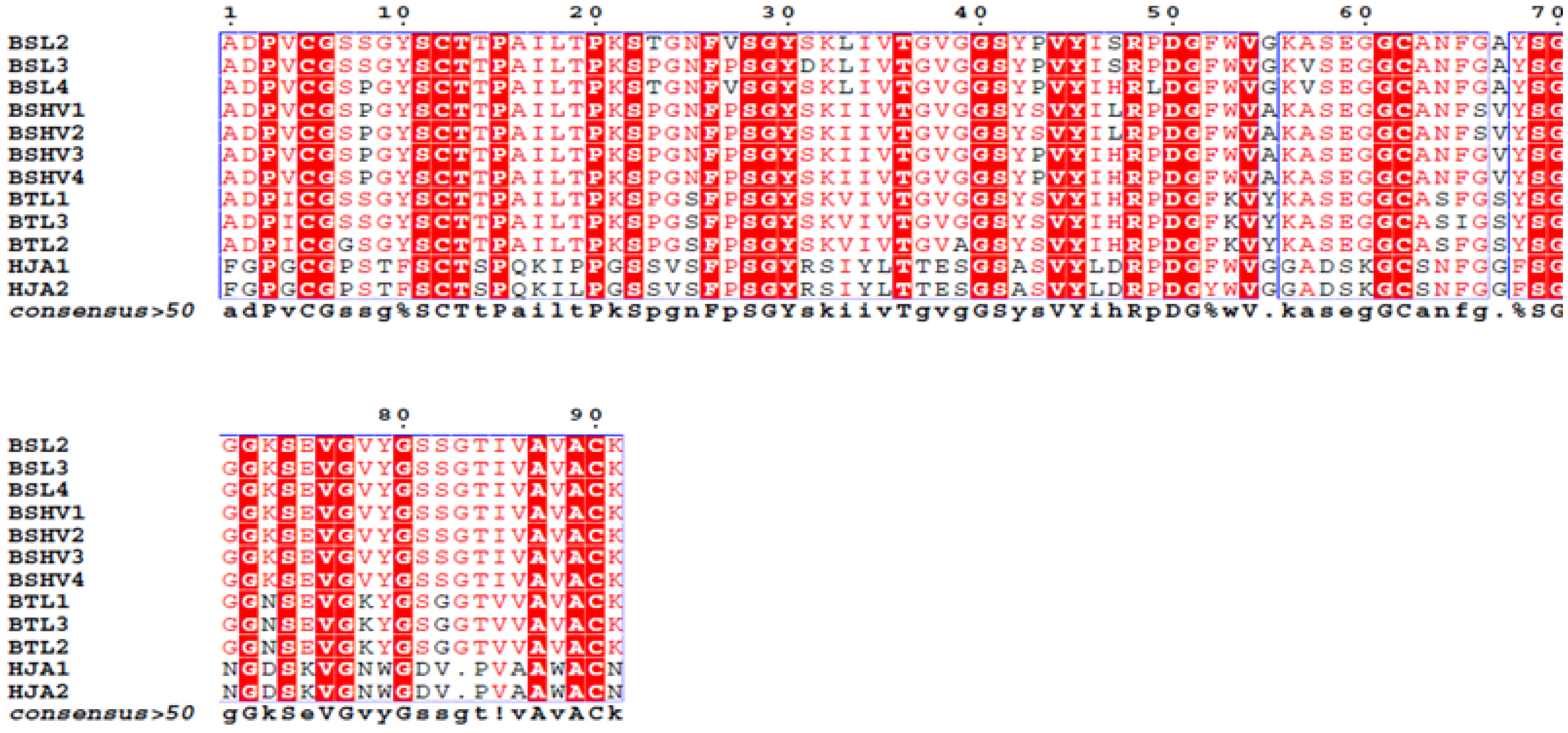
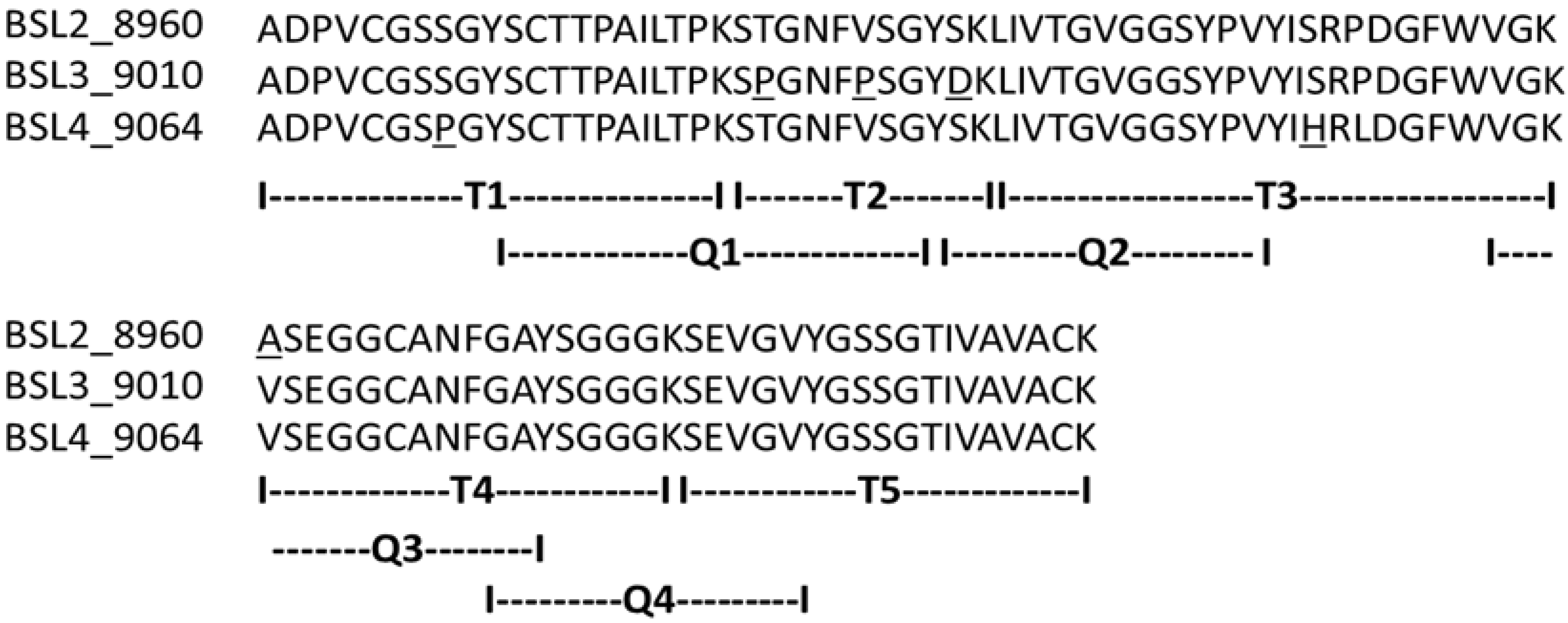
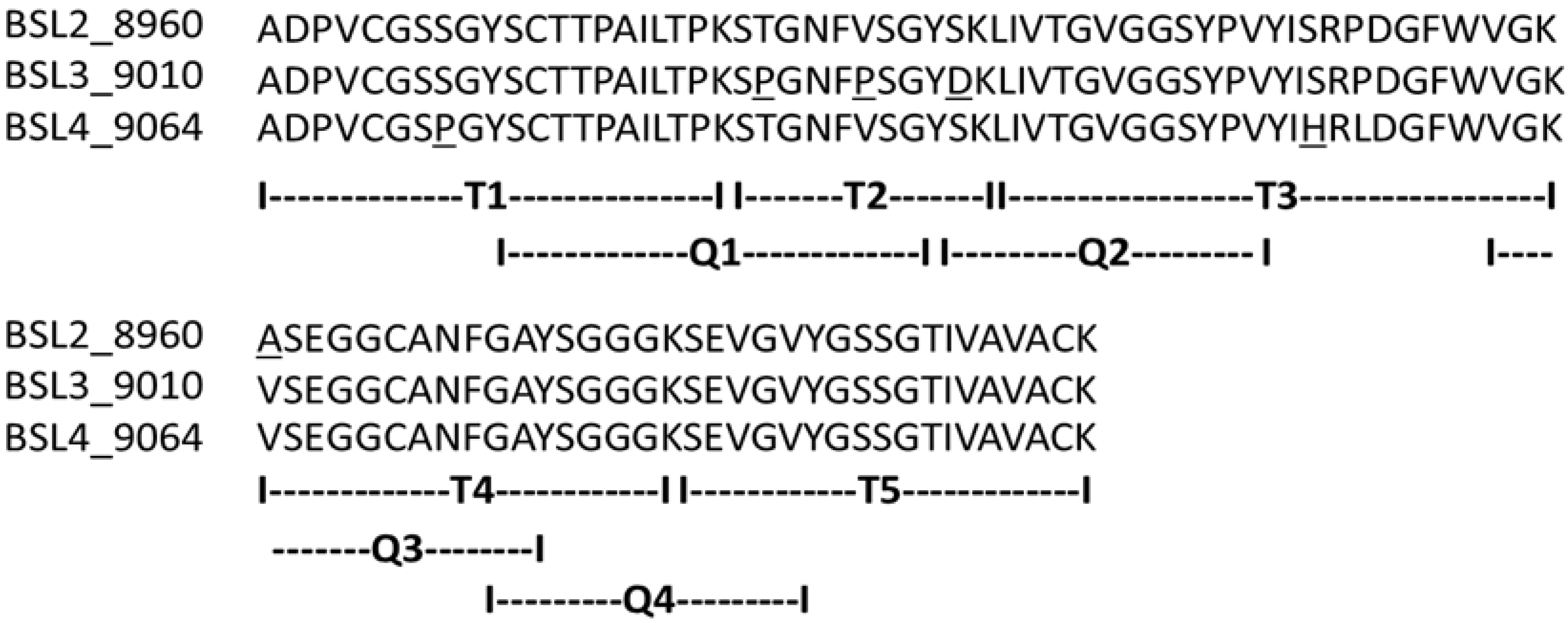
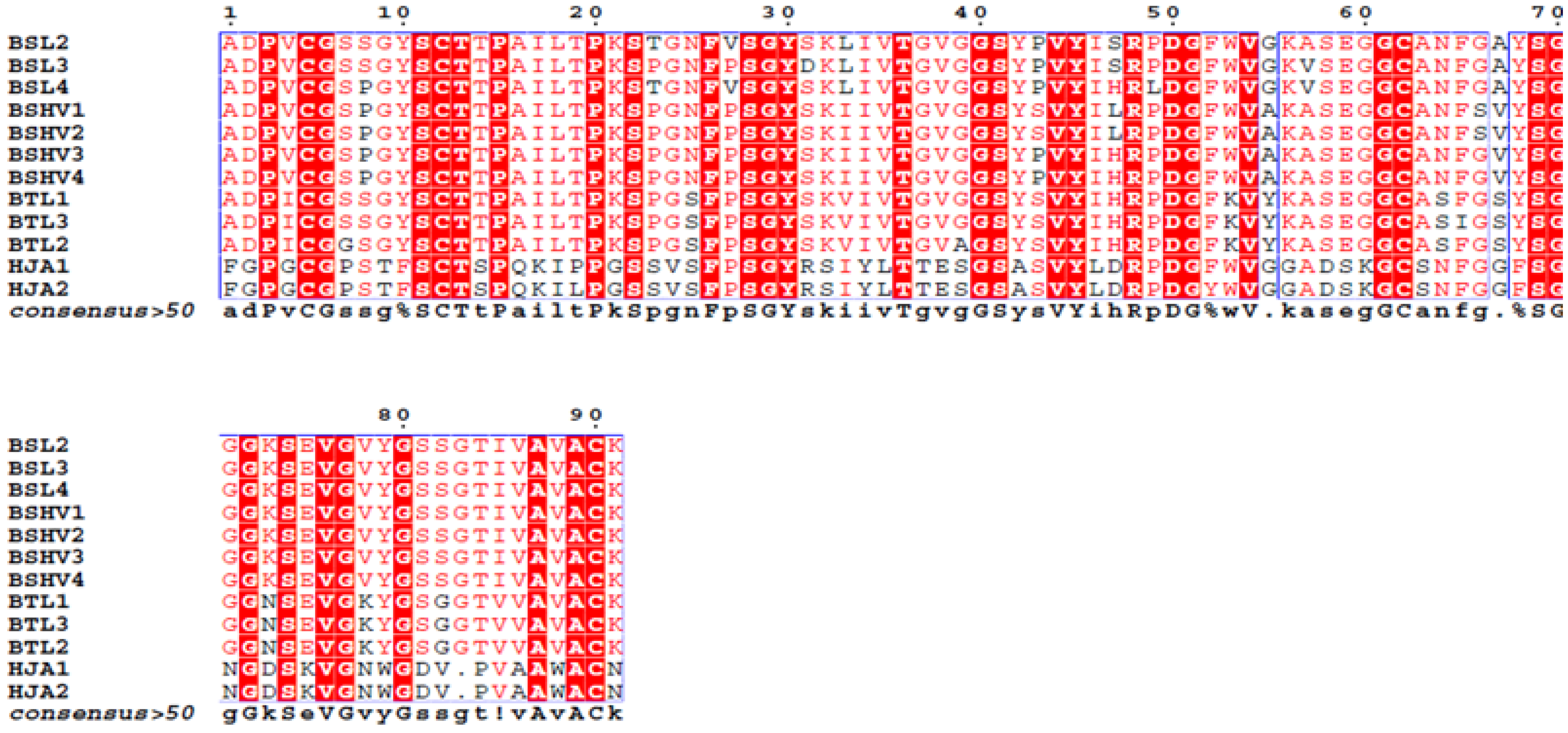
The complete amino acid sequences of BSL2, BSL3 and BSL4 were determined using tandem mass spectrometry (MS/MS) by overlapping the sequences of the peptides generated by digestion of CA-BSL with trypsin and chymotrypsin (
Figure 4,
Table 1). Sequence heterogeneity was observed at positions 8 (S/P), 23 (T/P), 27 (V/P), 31 (D/S), 47 (S/H), 49 (P/L) and 57 (A/V).
The isolectins from the marine red alga
Bryothamnion seaforthii (BSL) are polypeptides composed of 91 amino acid residues including four half-cystines. Methionine and glutamine were not detected in their sequences. The molecules contained a remarkably high content of Ser (13, 12 and 11 in BSL2, BSL3 and BSL4, respectively), Gly (18 in all) and Val (10 in all), indicating that these three amino acids constitute approximately 44% of the total amino acid residues. The isotope-averaged molecular masses calculated for the BSL2, BSL3 and BSL4 isoforms were 8960.28, 9010.309 and 9064.32, respectively, which is in excellent agreement with the experimentally determined masses. Four half-cystines were found in BSL, and no free sulfhydryl groups were identified (
Figure 3). The ion at
m/z 2289.9487 from the pepsin digestion of BSL corresponds to the peptides
1ADPVCGSPSGY
11 and
53WVGKASEGGCANF
65 linked by a disulfide bond between cysteines 5 and 62. The reduction and alkylation of this peptide results in the dissociation of the disulfide bond and the formation of two ions at
m/z 1430.5880 and 1070.4320 that correspond to the ethylpyridylated peptides WVGKASEGGCANF and ADPVCGSPSGY, respectively. These data suggest that two intrachain disulfide bonds, Cys
5–Cys
62 and Cys
12–Cys
90, are present in BSL.
Figure 4.
Complete amino acid sequence of BSL 2, BSL 3 and BSL 4. The tryptic and chymotryptic peptides are represented by T and Q, respectively. The underlined residues represent micro heterogeneity of the isoform sequences.
Figure 4.
Complete amino acid sequence of BSL 2, BSL 3 and BSL 4. The tryptic and chymotryptic peptides are represented by T and Q, respectively. The underlined residues represent micro heterogeneity of the isoform sequences.
Table 1.
Sequenced peptides and their respective molecular masses.
Table 1.
Sequenced peptides and their respective molecular masses.
| Peptide | Sequence ** | Calculated mass | Experimental mass | Δ Mass (Da) |
|---|
| T1.1 * | ADPVCGSPGYSCTTPAILTPK | 2192.99 | 2192.90 | 0.09 |
| T1.2 * | ADPVCGSSGYSCTTPAILTPK | 2182.88 | 2182.88 | 0.00 |
| T2.1 | SPGNFPSGYDK | 1167.52 | 1167.43 | 0.09 |
| T2.2 | STGNFVSGYSK | 1145.53 | 1145.50 | 0.03 |
| T3.1 | LIVTGVGGSYPVYISRPDGFWVGK | 2566.36 | 2566.30 | 0.06 |
| T3.2 | LIVTGVGGSYPVYIHRLDGFWVGK | 2632.41 | 2632.19 | 0.22 |
| T4.1 * | ASEGGCANFGAYSGGGK | 1589.68 | 1589.64 | 0.04 |
| T4.2 * | VSEGGCANFGAYSGGGK | 1617.64 | 1617.67 | −0.03 |
| T5 * | SEVGVYGSSGTIVAVACK | 1783.82 | 1783.87 | −0.05 |
| Q1 * | SCTTPAILTP KSPGNFPSGY | 2094. 99 | 2094. 93 | 0.06 |
| Q2 | SKIIVTGVGGSYPVY | 1538.83 | 1538.74 | 0.09 |
| Q3 * | VGKASEGGCANFGAY | 1487.64 | 1487.60 | 0.04 |
| Q4 | GAYSGGGKSEVGVY | 1329.62 | 1329.55 | 0.07 |
The alignment of the BSL sequence against the BLAST database reveals 48% identity and 64% similarity between BSL and HJA, a lectin isolated from marine red alga
Hypnea japonica. Compared with the
B. triquetrum lectin (BTL), BSL presents only seven exclusive positions, resulting in 83% identity. Relative to BSH, a hemagglutinin isolated from
B. seaforthii collected on the Venezuela coast [
22], BSL diverges in five exclusive positions beyond the heterogeneities among their own isoforms: 43 (P/S), 47 (S/L), 55 (G/A), 66 (G/S) and 67 (A/V) (
Figure 5).
Figure 5.
Alignment of the family of lectins similar to BSL. BSL2 to BSL4 are isolectins from Bryothamnion seaforthii, BSHV1 to BSHV4 are isolectins from B. seaforthii from the Venezuela coast, BTL1 to BTL3 are isolectins from B. triquetrum, and HJA1 to HJA2 are isolectins from Hypnea japonica.
Figure 5.
Alignment of the family of lectins similar to BSL. BSL2 to BSL4 are isolectins from Bryothamnion seaforthii, BSHV1 to BSHV4 are isolectins from B. seaforthii from the Venezuela coast, BTL1 to BTL3 are isolectins from B. triquetrum, and HJA1 to HJA2 are isolectins from Hypnea japonica.
In recent years, a large number of lectins have been isolated from marine red alga [
23,
24,
25]. These lectins generally have a low molecular weight, an isoelectric point in the range of pH 4–6, no affinity for simple sugars, no requirement for divalent cations for their biological activities and, in some cases, the presence of repeated domains [
24]. Although some of these proteins exhibit potential biological activities [
26,
27,
28,
29,
30], only a few have actually been studied at the structural level. BTL was the first marine algal lectin to have its primary structure determined [
19].
In the same year, Hori
et al. [
21] reported the primary structure of HJA, which shares sequence similarity with BTL and constitutes the first marine red alga lectin family. Based on the identity between HML and HCA and the differences in the amino acid sequences compared with BTL/HJA, Nagano
et al. [
13] suggested that HCA/HML constitute a new lectin family. Conversely, the monomeric lectins isolated from
Eucheuma serra,
E. amakusaensis, and
E. cottonii [
20] have masses of approximately 28 kDa, exist as monomers, and share
N-terminal sequence similarity with the complete amino acid sequence of isolectin 2 from
Eucheuma serra (ESA-2) [
31].
Additionally, the primary structure of ESA-2 contains repeated domains. These data suggest that lectins from the genus Eucheuma can be grouped in a third family of marine red alga lectins. In the present study, five isoforms of BSL, namely BSL1, BSL2, BSL3, BSL4 and BSL5, were identified, and the amino acid sequence of three (BSL2, BSL3, and BSL4) were determined from an MS/MS analysis of tryptic and chymotryptic peptides. BSL exhibited four cysteines in its primary structure that are involved in two intrachain disulfide bonds: Cys
5–Cys
12 and Cys
62–Cys
90. However, in BTL, one of these cysteines can interact with a cysteine present in another polypeptide chain to eventually form a dimer [
19]. In HJA, the disulfide bonds are exclusively involved in intrachain bonds with the pattern Cys
5–Cys
62 and Cys
12–Cys
89[
21]. The data obtained with BSL suggest that the disulfide bonds are also exclusively intrachain with the same pattern observed in HJA. Although the pattern of disulfide bonds between HJA and BSL is different from that presented by BTL, given the similarity between these lectins and the presence of isoforms among other similar biochemical characteristics, the data strongly suggest that BSL is a novel member of the first marine red alga lectin family that includes the isoforms of HJA, BTL and BSH.
2.2. Evaluation of Healing Potential
It was observed that the treatment of wounds with the lectin isolated from the marine red alga Bryothamnion seaforthii (BSL) induced a greater inflammatory process, which is observed during clinical evaluation and by well-evidenced phlogistic signs, such as edema and hyperemia in the first days following treatment, when compared with controls. Such signs are indicative of a pro-inflammatory effect by the lectin tested (data not shown).
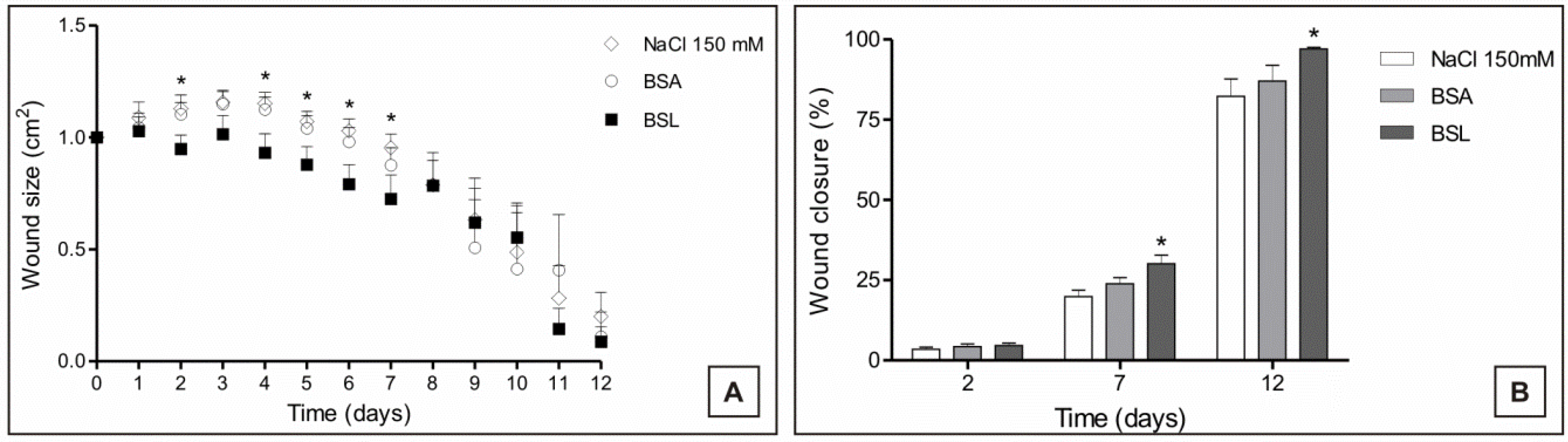
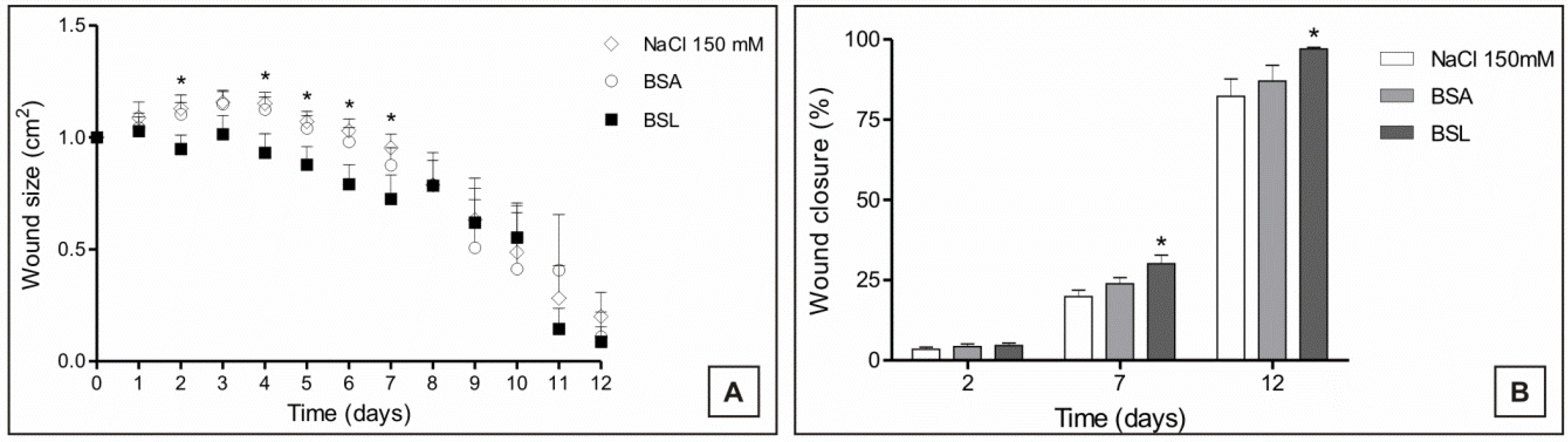
The measurement of the areas of the experimental lesions was performed daily to observe the evolution of healing in the experimental groups. During the inflammatory and proliferative phase, it could be observed that treatment with BSL induced a decrease in the areas of the lesions of the experimental animals in this group (
Figure 6).
Figure 6.
(A) Evolution of the areas; and (B) proportion of experimental closure of skin lesions in mice treated topically with 150 mM NaCl, bovine serum albumin (BSA) and Bryothamnion seaforthii lectin (BSL); * Significant in relation to the 150 mM NaCl group, p < 0.05.
Figure 6.
(A) Evolution of the areas; and (B) proportion of experimental closure of skin lesions in mice treated topically with 150 mM NaCl, bovine serum albumin (BSA) and Bryothamnion seaforthii lectin (BSL); * Significant in relation to the 150 mM NaCl group, p < 0.05.
The percentage of wound closure was evaluated from the exposed areas of each lesion according to the time. BSL treatment induced rapid and more effective healing compared with the controls (BSA and 150 mM NaCl) in which a higher proportion of closure in the animals treated with the lectin was observed on POD (postoperative day) 2, 7 and 12 (
Figure 6). Thus, the greater effectiveness of treatment with BSL can be observed in the proliferative and remodeling phases of the healing process when compared to the controls.
The BSL treatment induced an increased closing in the areas of the lesions in the group treated with BSL from POD 1 to 12, and this was possibly influenced by the contraction process of the wounds during healing.
2.3. Histopathological Assessment
For the histopathological evaluation of the injured tissue, samples were collected from each group on POD 2, 7 and 12 (n = 4 for all groups).
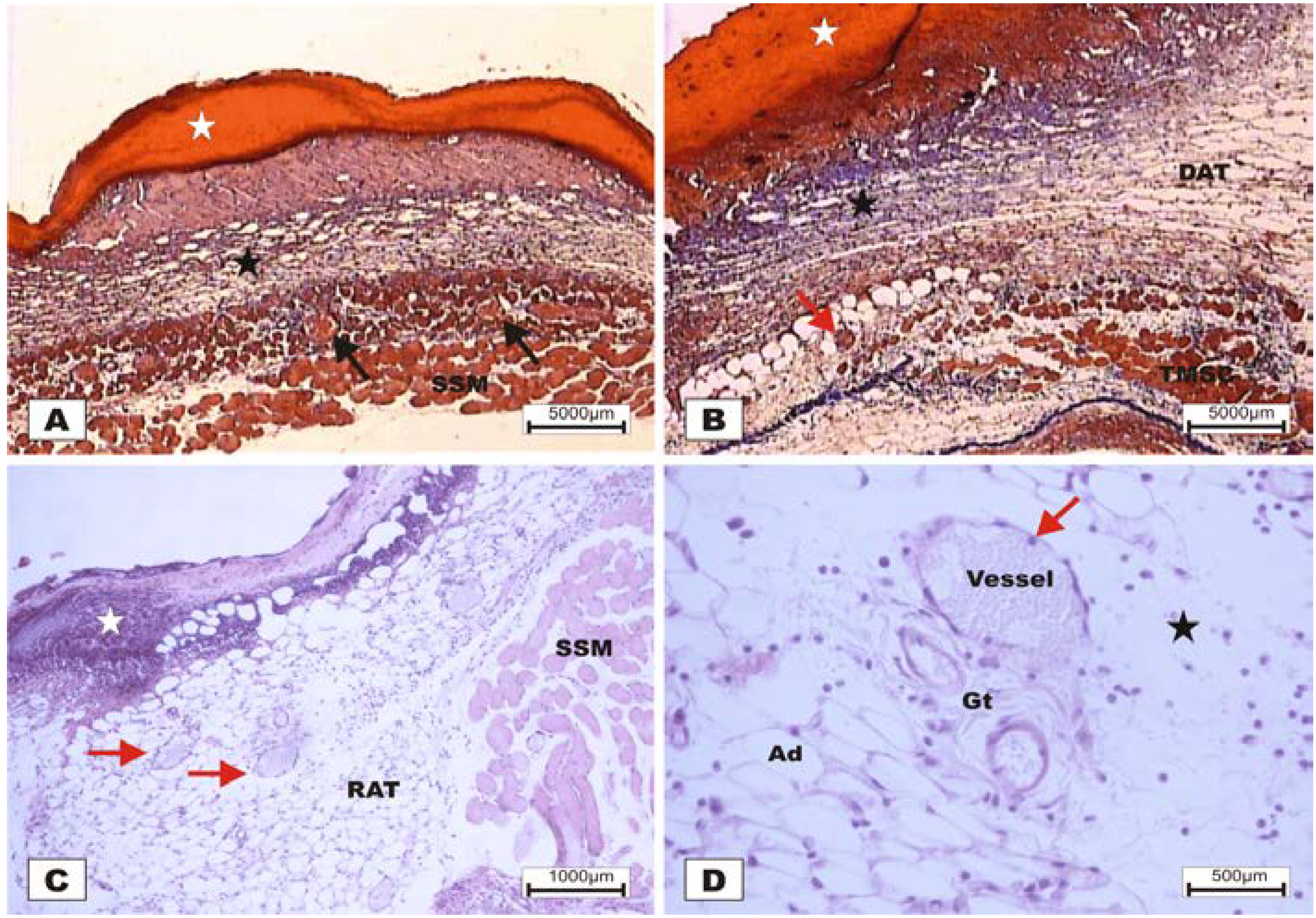
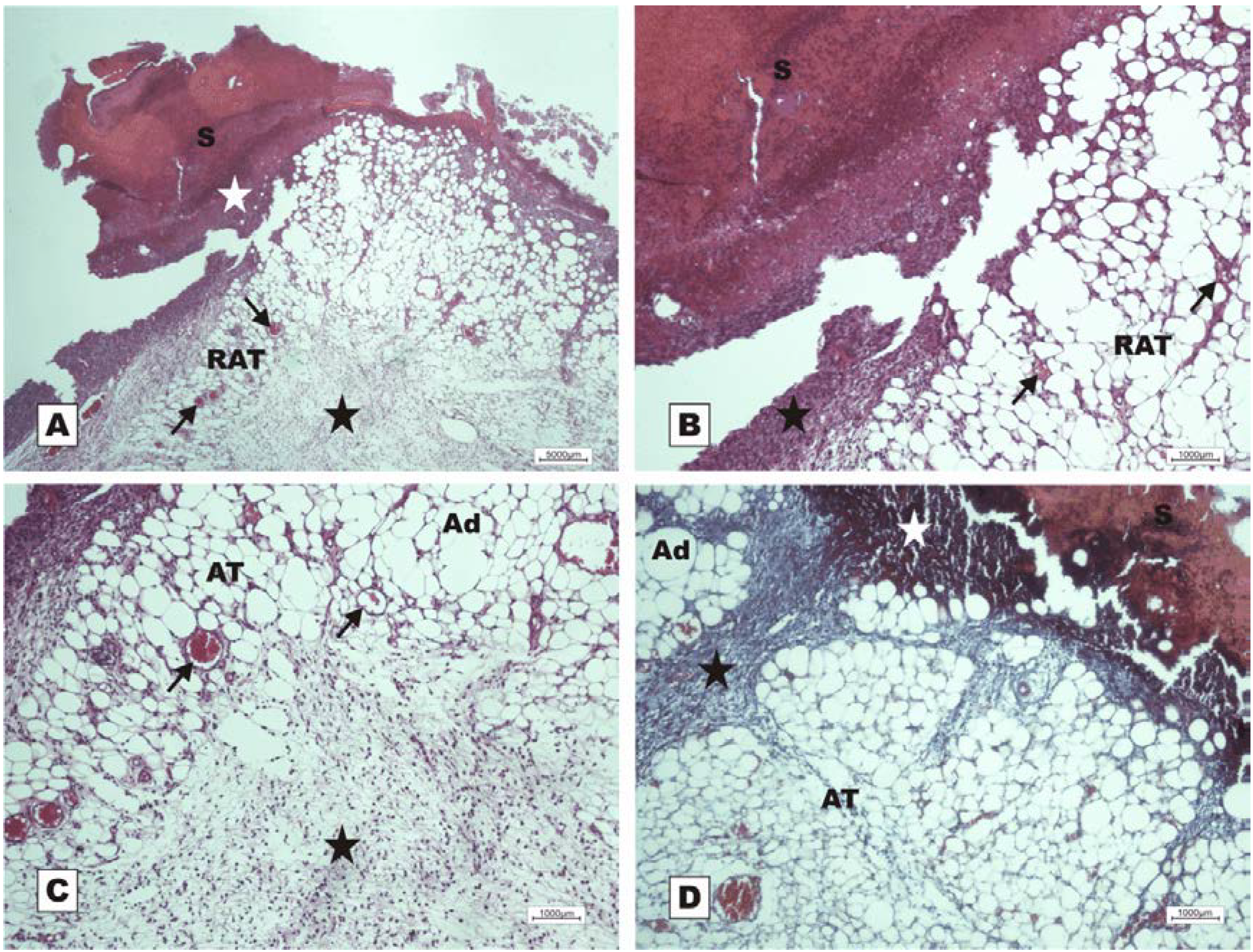
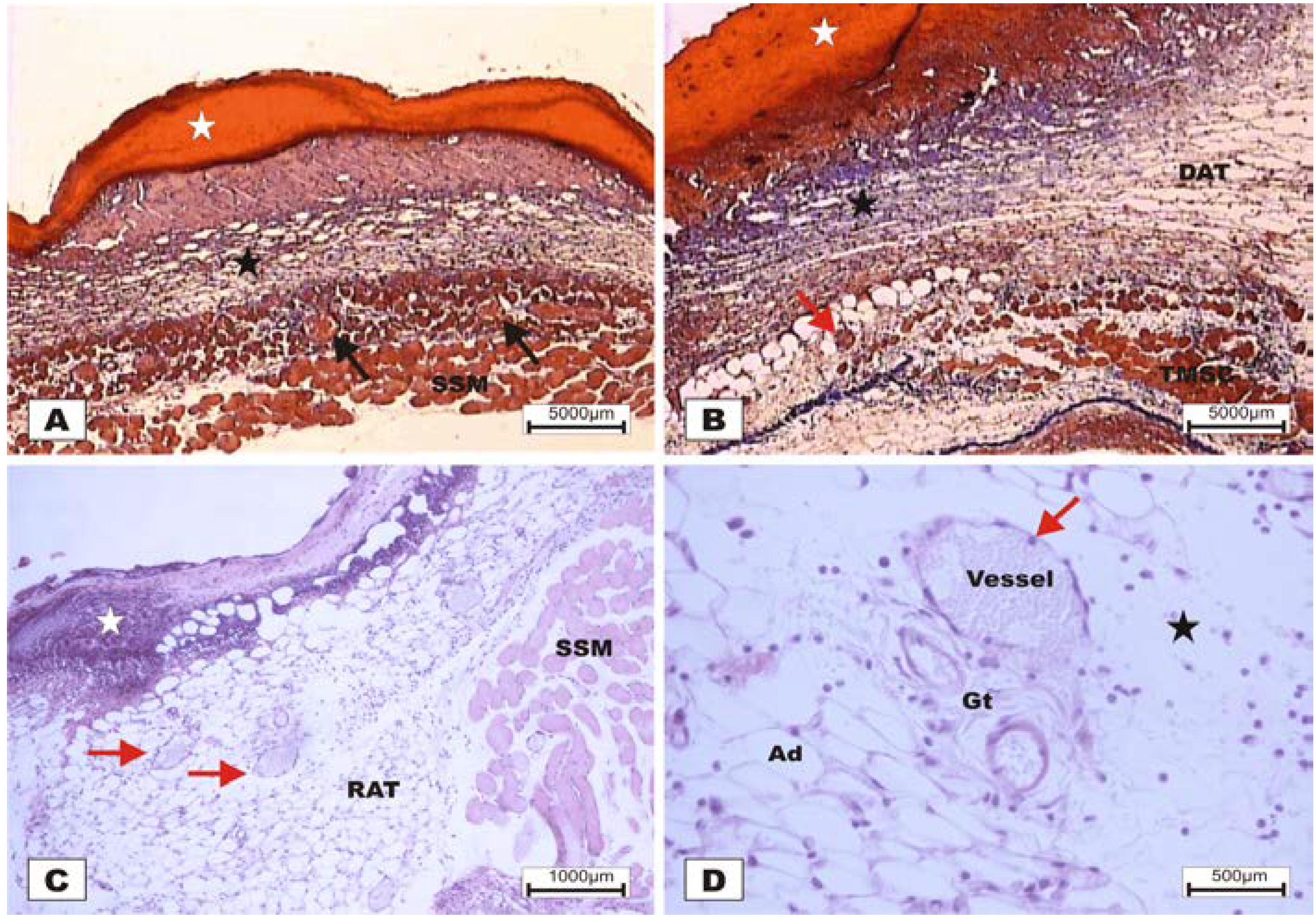
The study revealed that the wounds treated with BSA and BSL exhibited scabs sealing the epithelium opening on POD 2 (
Figure 7). During the histopathological diagnosis, wounds treated with BSL showed intense inflammatory exudates in the reactional adipose tissue, with a mild inflammatory infiltrate in the reticular dermis in addition to the presence of granulation tissue in the reactional adipose tissue (
Figure 7C,D). In wounds treated with BSA, there were intense inflammatory exudates, collagenolysis, and edema of the reticular dermis with the degenerative adipose tissue showing the beginning of a poor healing (
Figure 7A,B).
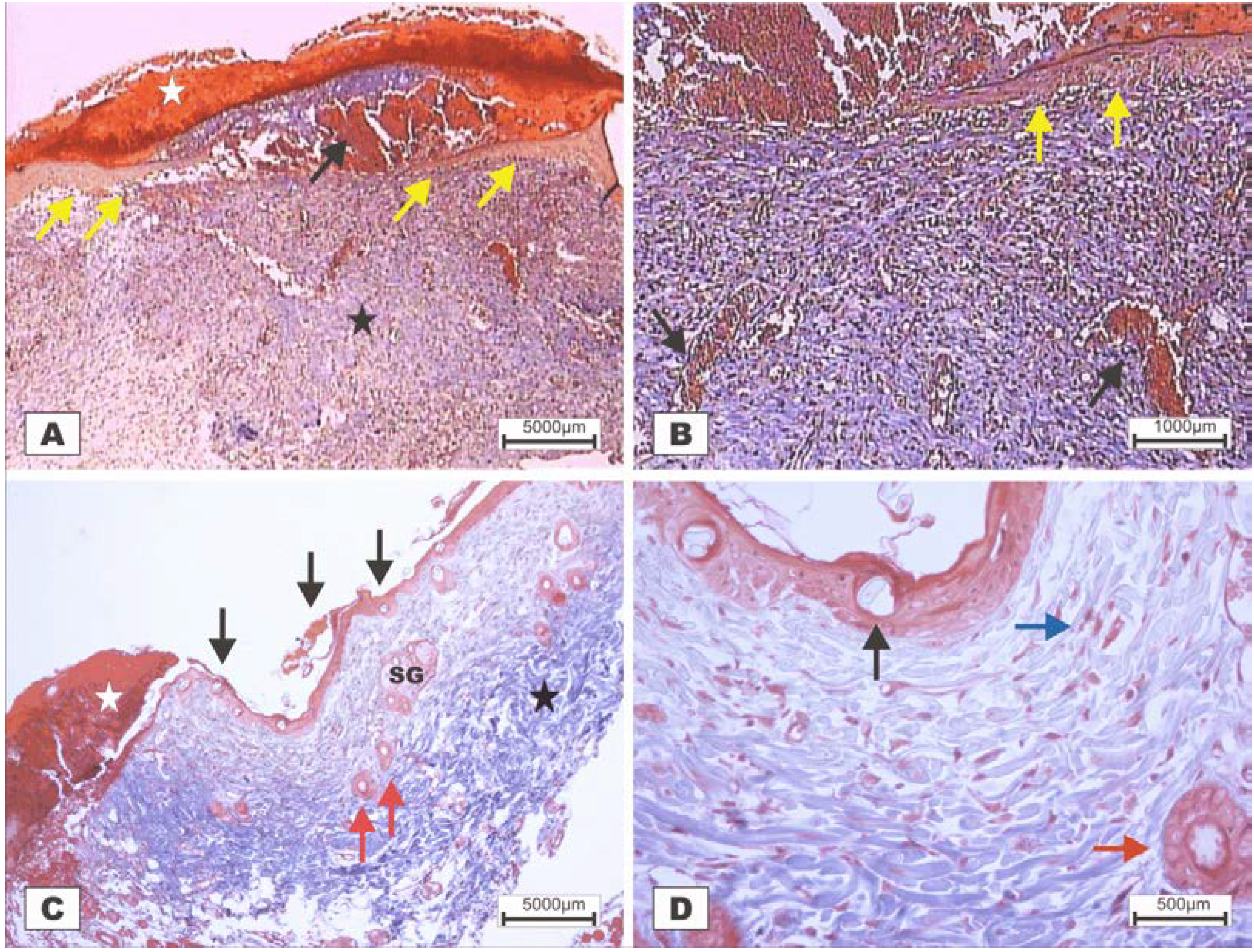
Figure 7.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 2 (Stain: hematoxylin-eosin and Masson’s trichrome). (A) (BSA). Note the presence of a scab (white star) covering the wound and intense inflammatory exudate. Below the exudate, a collagenolysis area and edema of the reticular dermis are observed (black star). Immediately below the dermis is the subcutaneous muscle layer with congested vessels (arrows). A 4× objective was used; (B) (BSA). Note the presence of a thicker crust (white star) covering the wound bed and a less-intense inflammatory exudate. Below the exudate, a collagenolysis area, edema of the reticular dermis and intense inflammatory infiltrate are observed (black star) with degeneration fatty tissue (DAT). Note the muscle fibers spaced by muscle tissue and fat cells with congested vessels near the collagenolysis area (red arrow). A 4× objective was used; (C) (BSL). A thick scab covers the wound bed (white star). Note the congested vessels of the granulation tissue (arrow) in the reactional adipose tissue (RAT). A 10× objective was used; (D) (BSL). Detail from the previous photo showing vessels of the granulation tissue in areas of intense inflammatory exudate (star). Observe the possible diapedesis of the neutrophil PMNs into the inflamed area (red arrow). A 40× objective was used.
Figure 7.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 2 (Stain: hematoxylin-eosin and Masson’s trichrome). (A) (BSA). Note the presence of a scab (white star) covering the wound and intense inflammatory exudate. Below the exudate, a collagenolysis area and edema of the reticular dermis are observed (black star). Immediately below the dermis is the subcutaneous muscle layer with congested vessels (arrows). A 4× objective was used; (B) (BSA). Note the presence of a thicker crust (white star) covering the wound bed and a less-intense inflammatory exudate. Below the exudate, a collagenolysis area, edema of the reticular dermis and intense inflammatory infiltrate are observed (black star) with degeneration fatty tissue (DAT). Note the muscle fibers spaced by muscle tissue and fat cells with congested vessels near the collagenolysis area (red arrow). A 4× objective was used; (C) (BSL). A thick scab covers the wound bed (white star). Note the congested vessels of the granulation tissue (arrow) in the reactional adipose tissue (RAT). A 10× objective was used; (D) (BSL). Detail from the previous photo showing vessels of the granulation tissue in areas of intense inflammatory exudate (star). Observe the possible diapedesis of the neutrophil PMNs into the inflamed area (red arrow). A 40× objective was used.
![Marinedrugs 10 01936 g007]()
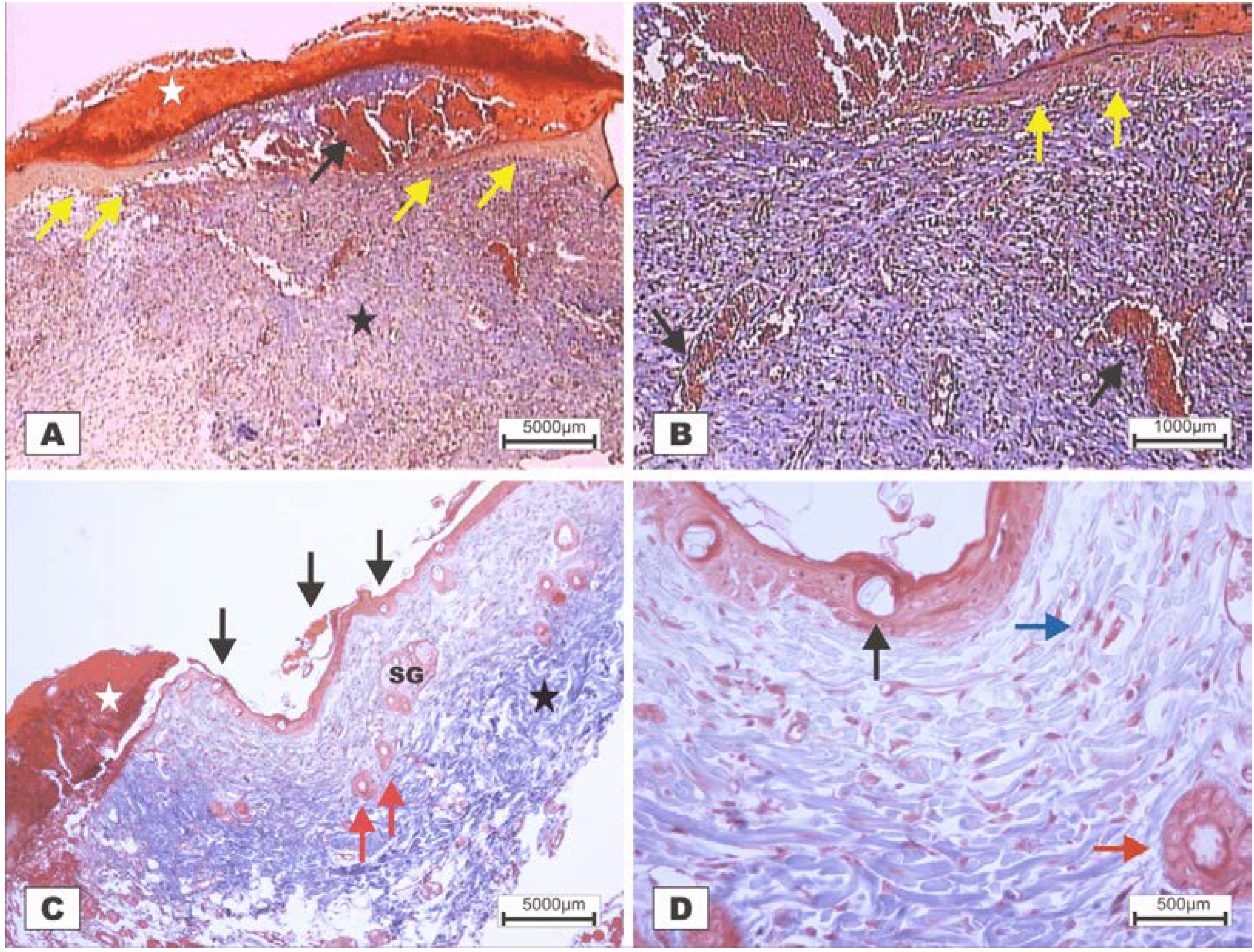
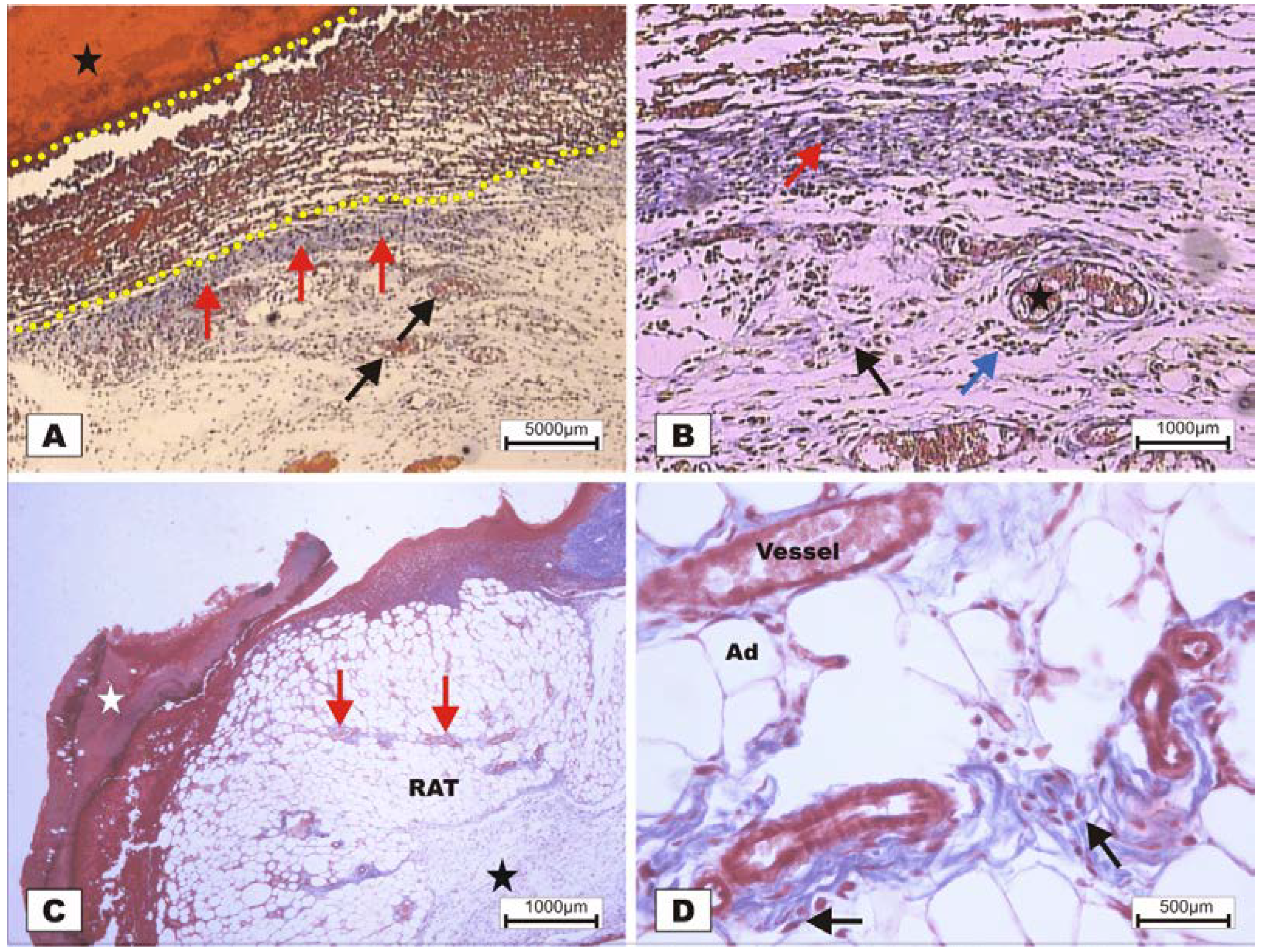
On POD 7, densification of collagen fibers in the reticular dermis, which is primarily surrounded by the vessels of the granulation tissue in adipose tissue with the presence of discrete reactional inflammatory exudates, was observed (
Figure 8C,D). In the group treated with BSA (
Figure 8A,B), intense inflammatory exudates and early fibroblastic proliferation with moderate collagen synthesis were still observed, demonstrating a less evolved healing process when compared with the wounds treated with BSL.
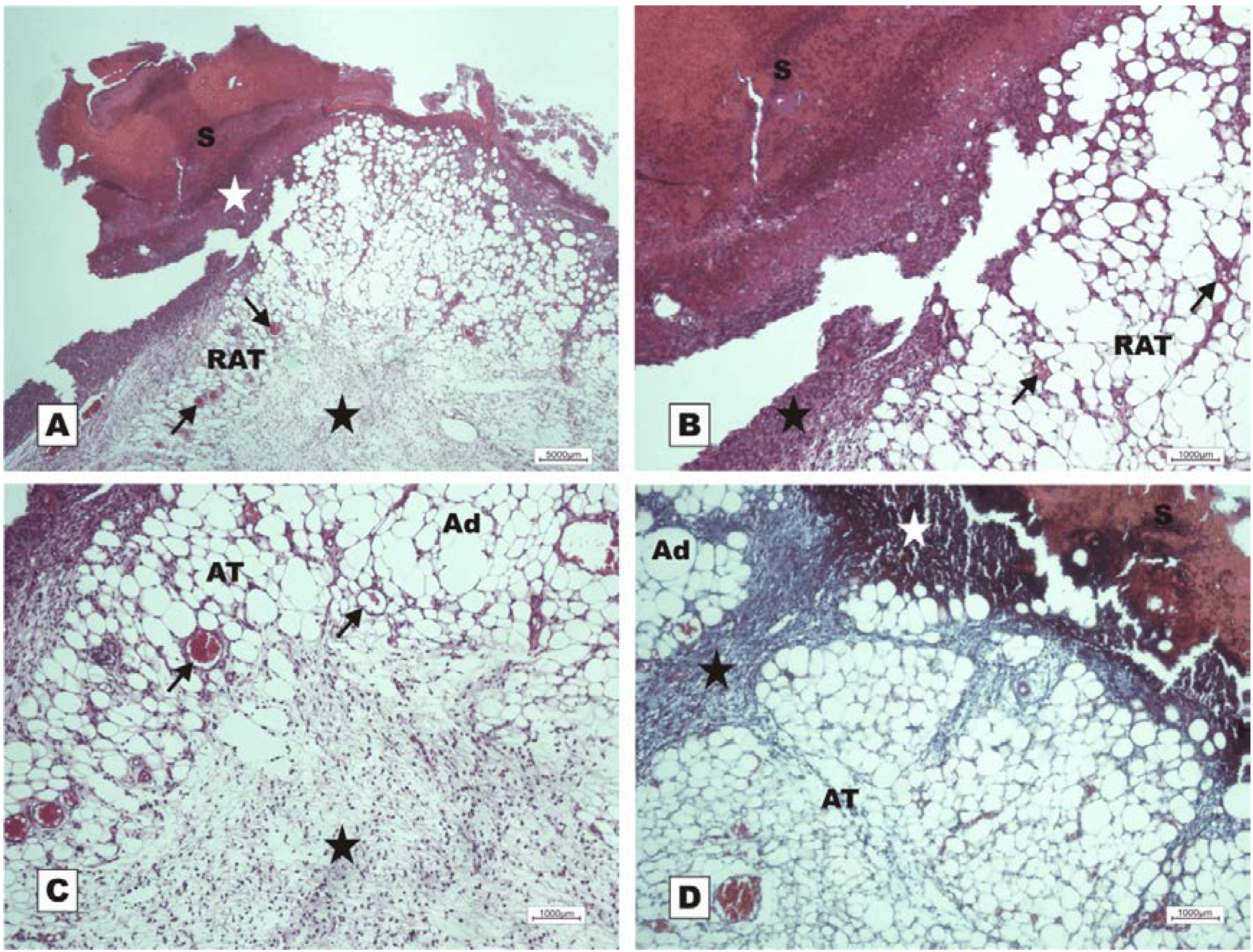
Figure 8.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 7 (Stain: Masson’s trichrome). (A) (BSA). Note the presence of a scab (star) covering the wound bed and intense inflammatory exudate (delimited by dashed yellow lines). Below the exudate, observe the area of granulation tissue with congested vessels (arrow) and intense fibroblastic proliferation. Immediately beneath the inflammatory exudate, note the bluish band resulting from the synthesis of collagen (red arrows). A 4× objective was used; (B) (BSA). Details of the granulation tissue. Note the fibroblastic proliferation (arrow) and mild inflammatory infiltrate around the vessels (blue arrow). Note the collagen above the range of the granulation tissue (red arrow). Vessels of the granulation tissue are observed (star). A 10× objective was used; (C) (BSL). Overview of the injured area. Note the presence of a crust sealing the epithelial opening (white star). Vessels of the granulation tissue in the reactional adipose tissue (RAT) surrounded by thickening collagen fibers (arrows) resulted from intense fibroblastic synthesis. An area with a mild inflammatory exudate is observed (black star). A 10× objective was used; (D) (BSL). Detail from the previous photo showing the area of granulation tissue vessels surrounded by active collagen. Note the active fibroblasts in the area (arrows). A 40× objective was used.
Figure 8.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 7 (Stain: Masson’s trichrome). (A) (BSA). Note the presence of a scab (star) covering the wound bed and intense inflammatory exudate (delimited by dashed yellow lines). Below the exudate, observe the area of granulation tissue with congested vessels (arrow) and intense fibroblastic proliferation. Immediately beneath the inflammatory exudate, note the bluish band resulting from the synthesis of collagen (red arrows). A 4× objective was used; (B) (BSA). Details of the granulation tissue. Note the fibroblastic proliferation (arrow) and mild inflammatory infiltrate around the vessels (blue arrow). Note the collagen above the range of the granulation tissue (red arrow). Vessels of the granulation tissue are observed (star). A 10× objective was used; (C) (BSL). Overview of the injured area. Note the presence of a crust sealing the epithelial opening (white star). Vessels of the granulation tissue in the reactional adipose tissue (RAT) surrounded by thickening collagen fibers (arrows) resulted from intense fibroblastic synthesis. An area with a mild inflammatory exudate is observed (black star). A 10× objective was used; (D) (BSL). Detail from the previous photo showing the area of granulation tissue vessels surrounded by active collagen. Note the active fibroblasts in the area (arrows). A 40× objective was used.
![Marinedrugs 10 01936 g008]()
At POD 12, the BSL-treated animals experienced restructuring of the epithelium covering with little production of keratin, formation of cutaneous immature annexes, and active collagen in the region of the reticular dermis with presence of fibroblastic activity (
Figure 9C,D). In the lesions treated with BSA over the same period, an epithelial scab sealing the epithelium opening, a mild inflammatory infiltrate beneath the crust and progression of the epithelial lining were observed (
Figure 9A,B).
Figure 9.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 12 (Stain: Masson’s trichrome). (A) (BSA). Presence of a scab (white star) and an inflammatory exudate beneath the scab (arrow). Note the progression of the luminal epithelium (yellow arrows) below the crust and inflammatory exudate. Dermis with less collagen deposition and the presence of active granulation tissue (black star) in a regression process is observed. A 4× objective was used; (B) (BSA). Details of the previous photo. Observe the granulation tissue characterized by the presence of vessels (black arrows) and young collagen in the extracellular matrix with inflammatory infiltrate. Note the projection of the epithelium beneath the crust (yellow arrows). A 10× objective was used; (C) (BSL). Note also the newly formed epithelium coated with little production of keratin (black arrows). The presence of a fragment of crust on the edge of the injured area (white star), the formation of skin appendages as sebaceous glands (SG), and the remaining vessels of the granulation tissue can be observed (red arrows). Note the active collagen in the reticular dermis (black star). A 4× objective was used; (D) (BSL). Details of the previous photo showing the presence of cutaneous annexes in the area of the newly formed epithelium (black arrow), fibroblasts in the papillary dermis area (blue arrow) and characteristic neoangiogenesis vessels in the granulation tissue (red arrow). A 40× objective was used.
Figure 9.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of BSA and BSL in POD 12 (Stain: Masson’s trichrome). (A) (BSA). Presence of a scab (white star) and an inflammatory exudate beneath the scab (arrow). Note the progression of the luminal epithelium (yellow arrows) below the crust and inflammatory exudate. Dermis with less collagen deposition and the presence of active granulation tissue (black star) in a regression process is observed. A 4× objective was used; (B) (BSA). Details of the previous photo. Observe the granulation tissue characterized by the presence of vessels (black arrows) and young collagen in the extracellular matrix with inflammatory infiltrate. Note the projection of the epithelium beneath the crust (yellow arrows). A 10× objective was used; (C) (BSL). Note also the newly formed epithelium coated with little production of keratin (black arrows). The presence of a fragment of crust on the edge of the injured area (white star), the formation of skin appendages as sebaceous glands (SG), and the remaining vessels of the granulation tissue can be observed (red arrows). Note the active collagen in the reticular dermis (black star). A 4× objective was used; (D) (BSL). Details of the previous photo showing the presence of cutaneous annexes in the area of the newly formed epithelium (black arrow), fibroblasts in the papillary dermis area (blue arrow) and characteristic neoangiogenesis vessels in the granulation tissue (red arrow). A 40× objective was used.
![Marinedrugs 10 01936 g009]()
In the control group treated with 150 mM NaCl, there was a delay in wound healing in which, at POD 7, acute inflammation and granulation tissue at the beginning of proliferation were observed (
Figure 10).
Figure 10.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of 150 mM NaCl in POD 7 (Stain: hematoxylin-eosin and Masson’s trichrome). (A) (NaCl). Open wound with a thick scab and dystrophic calcification (white star) sealing the epithelial opening (S). Observe the beginning of granulation tissue formation originating from the reactional adipose tissue (RAT), and note the presence of congested vessels (arrows) and the area of inflammation (star). A 4× objective was used; (B) (NaCl). Details showing the congested vessels in fat tissue (arrows) and the inflammatory infiltrate beneath the crust (star). A 10× objective was used; (C) (NaCl). Details of the area of acute inflammation in the degenerative adipose tissue (star) and the granulation tissue vessels above the inflamed area (arrows). A 10× objective was used; (D) (NaCl). Observe the inflammatory exudate beneath the crust (white star). There is poor collagen deposition and poor formation of granulation tissue in the dermis (black star) between the areas of adipose tissue. A 10× objective was used.
Figure 10.
Photomicrographs of surgically induced skin wounds in mice treated with a topical administration of 150 mM NaCl in POD 7 (Stain: hematoxylin-eosin and Masson’s trichrome). (A) (NaCl). Open wound with a thick scab and dystrophic calcification (white star) sealing the epithelial opening (S). Observe the beginning of granulation tissue formation originating from the reactional adipose tissue (RAT), and note the presence of congested vessels (arrows) and the area of inflammation (star). A 4× objective was used; (B) (NaCl). Details showing the congested vessels in fat tissue (arrows) and the inflammatory infiltrate beneath the crust (star). A 10× objective was used; (C) (NaCl). Details of the area of acute inflammation in the degenerative adipose tissue (star) and the granulation tissue vessels above the inflamed area (arrows). A 10× objective was used; (D) (NaCl). Observe the inflammatory exudate beneath the crust (white star). There is poor collagen deposition and poor formation of granulation tissue in the dermis (black star) between the areas of adipose tissue. A 10× objective was used.
![Marinedrugs 10 01936 g010]()
Wound healing is characterized as a complex process that results in contraction and wound closure, as well as restoration of the epithelium as a functional coating [
32]. Thus, the repair of skin wounds is accompanied by an ordered sequence of biological events starting with the closure of the wound and advancing to the remodeling of damaged tissue [
33].
The use of natural products to improve and accelerate the healing process has been widely envisioned because delayed skin healing, such as a problem in tissue repair in individuals suffering from diabetes or leprosy, is a major economic implication in medicine. This economic factor is a result of large drug spending over a long period that patients who suffer from chronic wounds or ulcers spend in hospitals [
1,
6,
34]. Thus, agents that decrease the time of the tissue repair process are desired to contribute to a faster and more effective healing process [
35].
Lectins are a structurally heterogeneous group of proteins of non-immune origin that have the characteristic to bind carbohydrates reversibly with high selectivity [
36]. Lectins are very versatile proteins and have many direct applications in biological and biomedical research [
37].
The field of application of lectins in the treatment of wounds is unexplored. However, important studies have shown that these biomolecules have healing potential in the treatment of acute wounds [
11,
12].
The biological application of lectins from marine algae deserves special interest because these lectins are lower molecular weight proteins than plant lectins. Thus, it is expected that these marine algae lectins are much less antigenic than others [
38]. Additionally, lectins from marine algae seem to express their biological activities in a stronger manner than lectins from other sources. Furthermore, they possess great molecular stability due to several disulfide bridges and show high specificity for complex carbohydrates and glycoconjugates, especially for mucins [
13,
21,
39].
The lectin isolated from the marine red alga
Bryothamnion seaforthii has been shown to have several biological activities, as described in the literature [
14,
27,
40,
41], and may be considered to be an important tool for biomedical applications.
Our findings demonstrate the potential pro-healing activity of the BSL, which was responsible for acceleration in the healing of skin wounds treated with this lectin. Thus, the histopathological findings showed that the lectin used in this study exerts its activity on the migration of polymorphonuclear cells to the injured site, confirming the data already published [
11] that suggest that the lectin of
Artocarpus integrifolia (KM+) induced a more effective healing of the corneal epithelium via a mechanism that also involves the migration of neutrophils.
It is possible that BSL is also able to act on the activation and proliferation of fibroblasts because most of the lectin-treated animals exhibited strongly active collagen in the dermis. However, future experiments are needed to better characterize the action of the BSL on fibroblasts.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}