
The Role of β1 Integrin/CD29 as a Potential Prognostic Factor for the Risk of Progression to Cervical Carcinoma in HPV-Associated Lesions


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cytofluorimetry
2.2. Immunohistochemistry
2.3. HPV-DNA Test
2.4. Statistical Analysis
3. Results
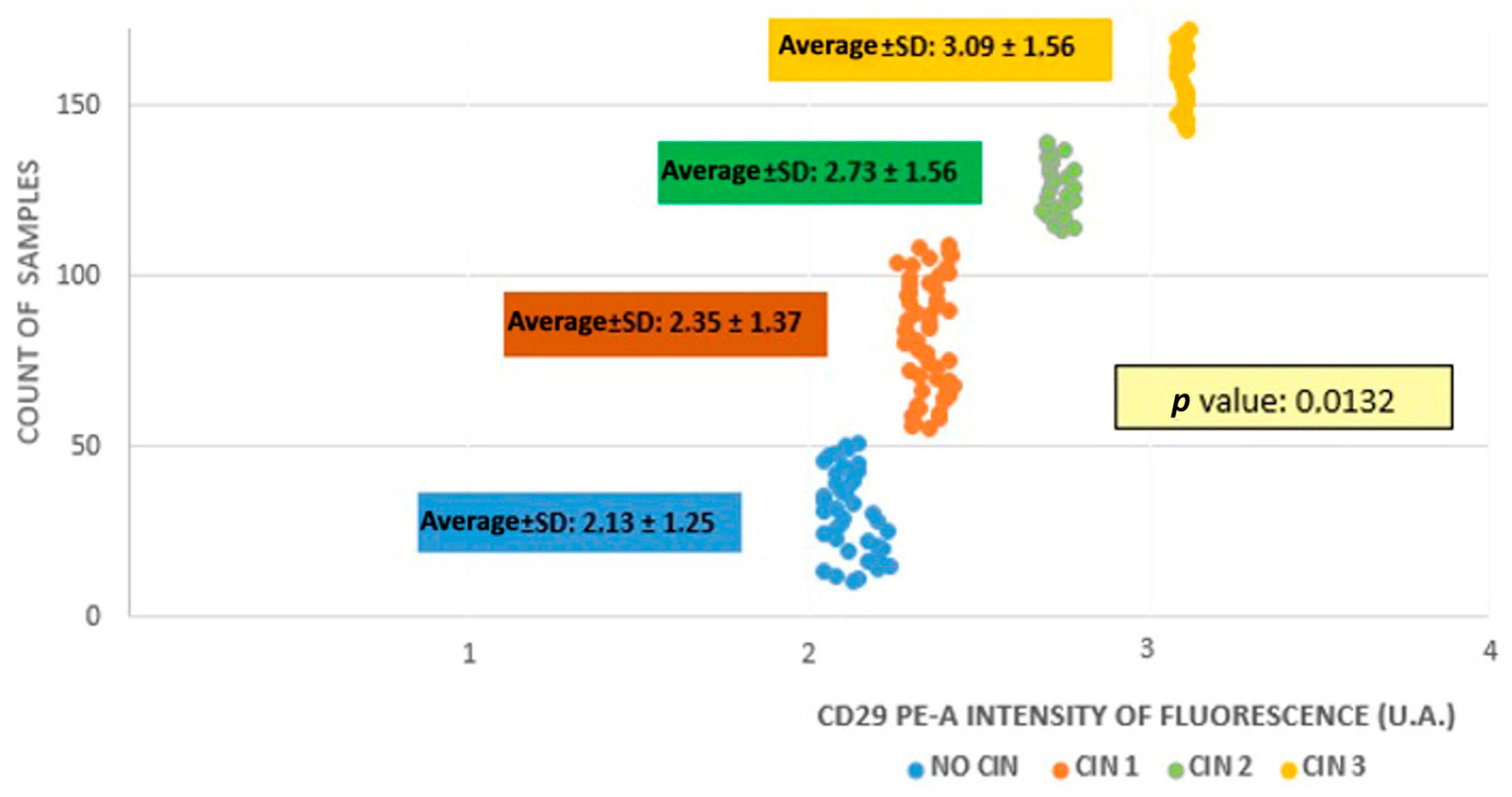
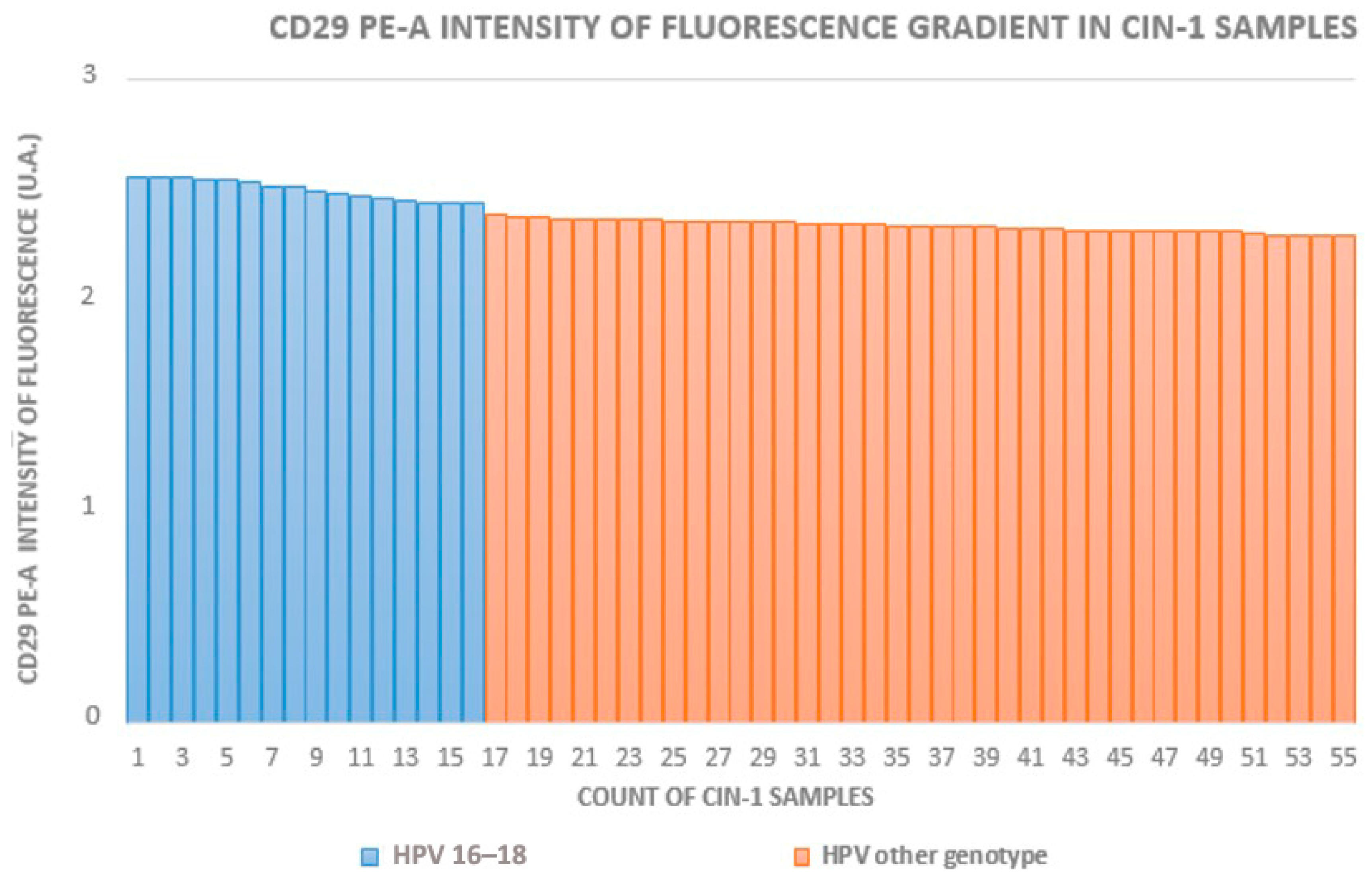
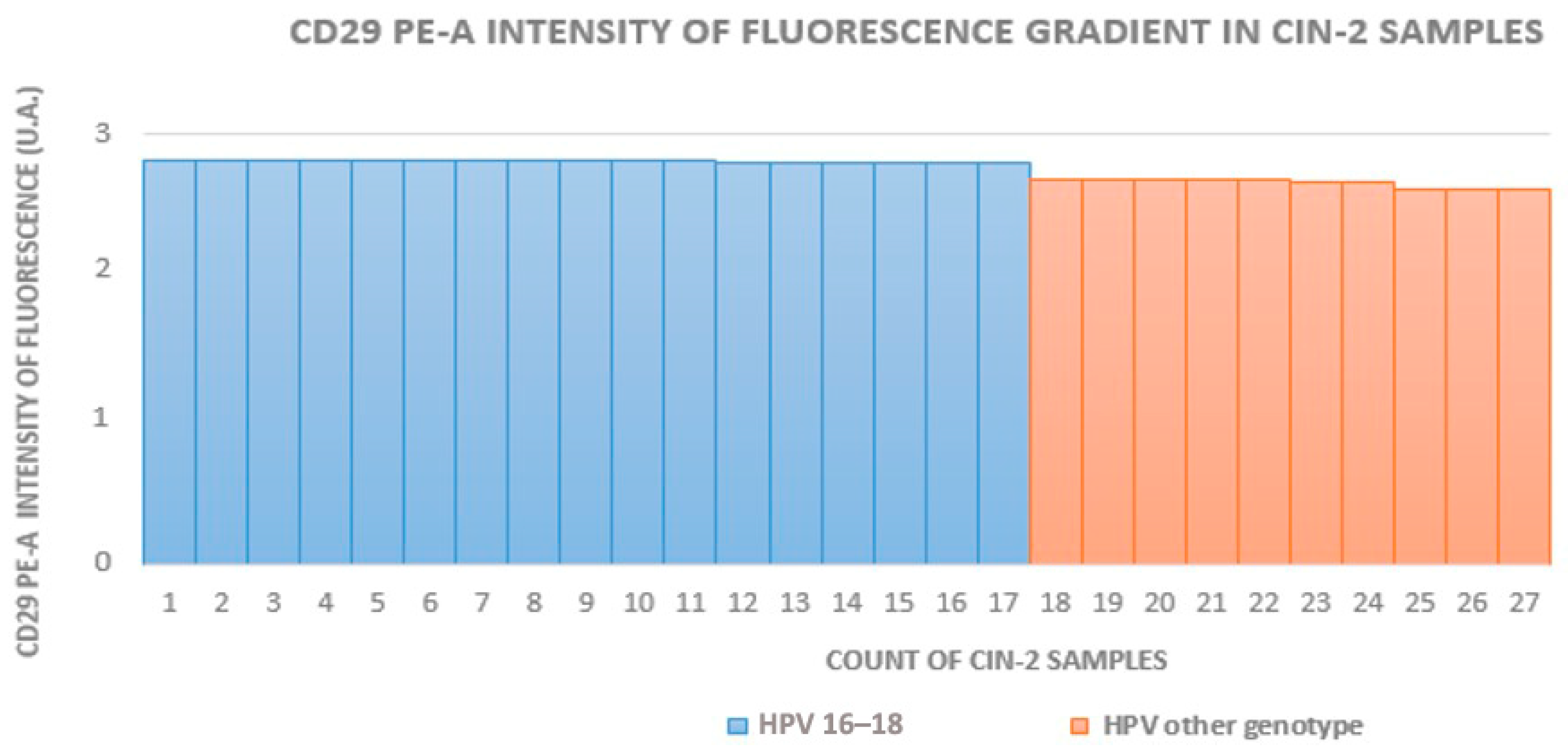
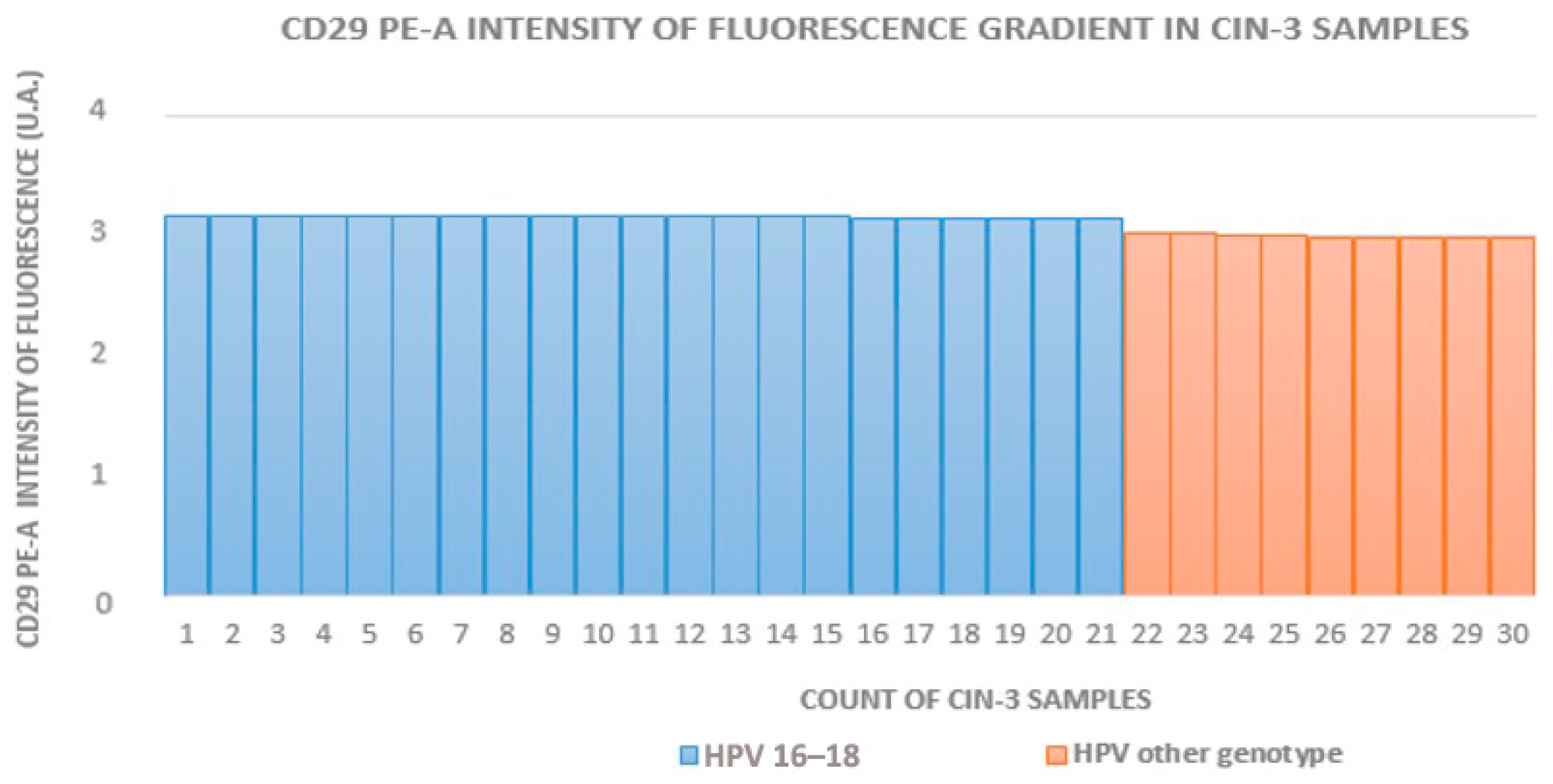
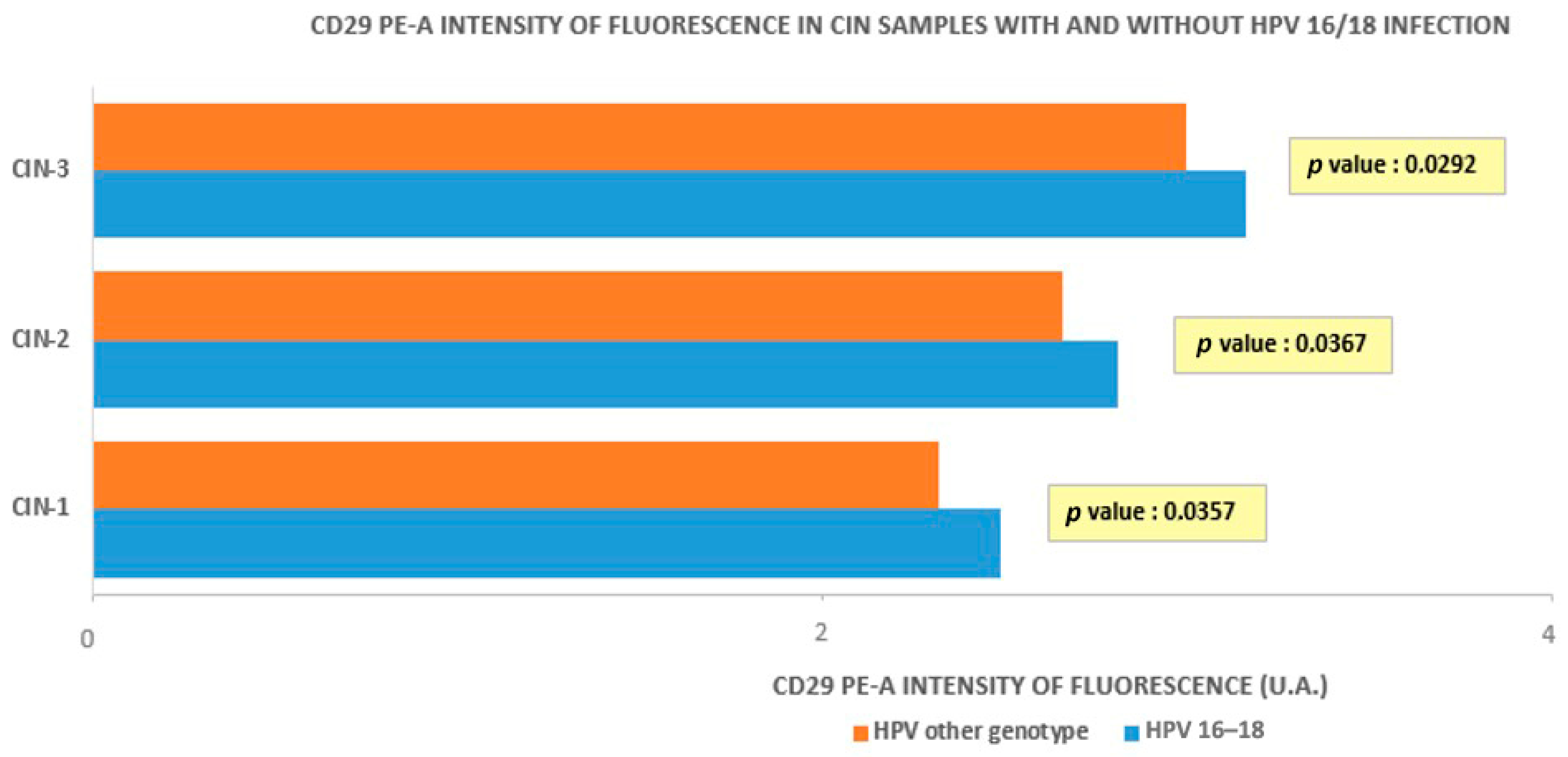
3.1. Cytofluorometry
3.2. Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef]
- Aplin, A.E.; Howe, A.; Alahari, S.K.; Juliano, R.L. Signal transduction and signal modulation by cell adhesion receptors: The role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins. Pharmacol. Rev. 1998, 50, 197–263. [Google Scholar]
- Schwartz, M.A.; Shattil, S.J. Signaling networks linking integrins and rho family GTPases. Trends Biochem. Sci. 2000, 25, 388–391. [Google Scholar] [CrossRef]
- Stromblad, S.; Becker, J.C.; Yebra, M.; Brooks, P.C.; Cheresh, D.A. Suppression of p53 activity and p21WAF1/CIP1 expression by vascular cell integrin alphaVbeta3 during angiogenesis. J. Clin. Investig. 1996, 98, 426–433. [Google Scholar] [CrossRef]
- Assoian, R.K.; Schwartz, M.A. Coordinate signaling by integrins and receptor tyrosine kinases in the regulation of G1 phase cell-cycle progression. Curr. Opin. Genet. Dev. 2001, 11, 48–53. [Google Scholar] [CrossRef]
- Boudreau, N.; Werb, Z.; Bissell, M.J. Suppression of apoptosis by basement membrane requires three-dimensional tissue organization and withdrawal from the cell cycle. Proc. Natl. Acad. Sci. USA 1996, 93, 3509–3513. [Google Scholar] [CrossRef] [PubMed]
- Stupack, D.G.; Puente, X.S.; Boutsaboualoy, S.; Storgard, C.M.; Cheresh, D.A. Apoptosis of adherent cells by recruitment of caspase-8 to unligated integrins. J. Cell Biol. 2001, 155, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Hansen, R.K.; Radisky, D.; Yoneda, T.; Barcellos-Hoff, M.H.; Petersen, O.W.; Turley, E.A.; Bissell, M.J. Phenotypic reversion or death of cancer cells by altering signaling pathways in three-dimensional contexts. J. Natl. Cancer Inst. 2002, 94, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Cheung, B.C.H.; Chen, X.; Davis, H.J.; Toth, J.; Segall, J.E.; Shenoy, V.B.; Wu, M. CD44 and beta1-integrin are both engaged in cell traction force generation in hyaluronic acid-rich extracellular matrices. bioRxiv 2023. [Google Scholar] [CrossRef]
- Park, C.S.; Yoshihara, H.; Gao, Q.; Qu, C.; Iacobucci, I.; Ghate, P.S.; Connelly, J.P.; Pruett-Miller, S.M.; Wagner, B.; Robinson, C.G.; et al. Stromal-induced epithelial-mesenchymal transition induces targetable drug resistance in acute lymphoblastic leukemia. Cell Rep. 2023, 42, 112804. [Google Scholar] [CrossRef] [PubMed]
- Bayik, D.; Bartels, C.F.; Lovrenert, K.; Watson, D.C.; Zhang, D.; Kay, K.; Lee, J.; Lauko, A.; Johnson, S.; Lo, A.; et al. Distinct Cell Adhesion Signature Defines Glioblastoma Myeloid-Derived Suppressor Cell Subsets. Cancer Res. 2022, 82, 4274–4287. [Google Scholar] [CrossRef]
- Choi, S.; Ismail, A.; Pappas-Gogos, G.; Boussios, S. HPV and Cervical Cancer: A Review of Epidemiology and Screening Uptake in the UK. Pathogens 2023, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J.; Arbyn, M.; Sankaranarayanan, R.; Tsu, V.; Ronco, G.; Mayrand, M.H.; Dillner, J.; Meijer, C.J. Overview of human papillomavirus-based and other novel options for cervical cancer screening in developed and developing countries. Vaccine 2008, 26 (Suppl. S10), K29–K41. [Google Scholar] [CrossRef]
- Riemma, G.; Schettino, M.T.; Munno, G.M.; Fasulo, D.D.; Sandullo, L.; Amabile, E.; La Verde, M.; Torella, M. Echinacea angustifolia and Echinacea purpurea Supplementation Combined with Vaginal Hyaluronic Acid to Boost the Remission of Cervical Low-Grade Squamous Intraepithelial Lesions (L-SILs): A Randomized Controlled Trial. Medicina 2022, 58, 646. [Google Scholar] [CrossRef]
- Yuan, Y.; Cai, X.; Shen, F.; Ma, F. HPV post-infection microenvironment and cervical cancer. Cancer Lett. 2021, 497, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Liu, L.; Liu, B.; Mao, X.G. Expression of integrin beta1 and its significance in squamous cell carcinoma of the cervix. Mol. Med. Rep. 2014, 9, 2473–2478. [Google Scholar] [CrossRef] [PubMed]
- Colacurci, N.; Schettino, M.T.; Grimaldi, V.; De Luca, F.P.; Mansueto, G.; Costa, D.; Cacciatore, F.; De Franciscis, P.; Napoli, C. Flow Cytometry Characterization of Pluripotent Transmembrane Glycoproteins on Resident Cervix Uteri Cells in Patients Screened for Cervical Cancer. Cancer Investig. 2020, 38, 228–239. [Google Scholar] [CrossRef]
- Riemma, G.; Mainini, G.; Lukanovic, D.; Scalzone, G.; Sandullo, L.; Schettino, M.T.; Vastarella, M.G.; Dominoni, M.; Pieretti, G.; De Franciscis, P.; et al. Efficacy of a Novel Rigenase((R)) and Polyhexanide (Fitostimoline((R)) Septagel) Hydrogel Device for the Treatment of Vulvovaginitis Symptoms: Cross-Sectional Analysis of a National Survey and Prospective Observational Study. Medicina 2023, 59, 2004. [Google Scholar] [CrossRef]
- Hodivala, K.J.; Pei, X.F.; Liu, Q.Y.; Jones, P.H.; Rytina, E.R.; Gilbert, C.; Singer, A.; Watt, F.M. Integrin expression and function in HPV 16-immortalised human keratinocytes in the presence or absence of v-Ha-ras. Comparison with cervical intraepithelial neoplasia. Oncogene 1994, 9, 943–948. [Google Scholar]
- Liu, J.; Tang, L.; Chu, W.; Wei, L. Cellular Retinoic Acid Binding Protein 2 (CRABP2), Up-regulated by HPV E6/E7, Leads to Aberrant Activation of the Integrin beta1/FAK/ERK Signaling Pathway and Aggravates the Malignant Phenotypes of Cervical Cancer. Biochem. Genet. 2023. [Google Scholar] [CrossRef]
- Holloway, A.; Storey, A. A conserved C-terminal sequence of high-risk cutaneous beta-human papillomavirus E6 proteins alters localization and signalling of beta1-integrin to promote cell migration. J. Gen. Virol. 2014, 95, 123–134. [Google Scholar] [CrossRef]
- Lee, C.H.; Chen, J.S.; Sun, Y.L.; Liao, W.T.; Zheng, Y.W.; Chai, C.Z.; Chen, G.S.; Yu, H.S. Defective beta1-integrins expression in arsenical keratosis and arsenic-treated cultured human keratinocytes. J. Cutan. Pathol. 2006, 33, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, F.; Chen, X.Y.; Yan, B.X.; Wang, Z.Y.; Chen, S.Q.; Zheng, M.; Man, X.Y. The epidermal immune microenvironment plays a dominant role in psoriasis development, as revealed by mass cytometry. Cell Mol. Immunol. 2022, 19, 1400–1413. [Google Scholar] [CrossRef]
- Cerri, A.; Favre, A.; Giunta, M.; Corte, G.; Grossi, C.E.; Berti, E. Immunohistochemical localization of a novel beta 1 integrin in normal and pathologic squamous epithelia. J. Investig. Dermatol. 1994, 102, 247–252. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jones, J.; Sugiyama, M.; Watt, F.M.; Speight, P.M. Integrin expression in normal, hyperplastic, dysplastic, and malignant oral epithelium. J. Pathol. 1993, 169, 235–243. [Google Scholar] [CrossRef]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-regulated FAK-Src signaling in normal and cancer cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef]
- Damgaard, R.; Jenkins, D.; Stoler, M.H.; de Koning, M.C.; van de Sandt, M.M.; Lycke, K.D.; Kahlert, J.; Gravitt, P.E.; Quint, W.V.; Steiniche, T.; et al. HPV genotypes and risk of persistence and progression in women undergoing active surveillance for cervical intraepithelial neoplasia grade 2. Am. J. Obstet. Gynecol. 2024; ahead of print. [Google Scholar] [CrossRef]
- de Sanjose, S.; Quint, W.G.; Alemany, L.; Geraets, D.T.; Klaustermeier, J.E.; Lloveras, B.; Tous, S.; Felix, A.; Bravo, L.E.; Shin, H.R.; et al. Human papillomavirus genotype attribution in invasive cervical cancer: A retrospective cross-sectional worldwide study. Lancet Oncol. 2010, 11, 1048–1056. [Google Scholar] [CrossRef]
- Berraho, M.; Amarti-Riffi, A.; El-Mzibri, M.; Bezad, R.; Benjaafar, N.; Benideer, A.; Matar, N.; Qmichou, Z.; Abda, N.; Attaleb, M.; et al. HPV and cofactors for invasive cervical cancer in Morocco: A multicentre case-control study. BMC Cancer 2017, 17, 435. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, S.; Li, X.; Wu, Z.; Zhao, Y.; Cui, J.; Liu, B.; Chen, F.; Zhang, X.; Qiao, Y.; et al. The features of high-risk human papillomavirus infection in different female genital sites and impacts on HPV-based cervical cancer screening. Virol. J. 2023, 20, 116. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Song, G.B.; Qin, J.; Luo, Q.; Shen, X.D.; Yan, R.B.; Cai, S.X. Adhesion of different cell cycle human hepatoma cells to endothelial cells and roles of integrin beta1. World J. Gastroenterol. 2005, 11, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Selivanova, G.; Ivaska, J. Integrins and mutant p53 on the road to metastasis. Cell 2009, 139, 1220–1222. [Google Scholar] [CrossRef] [PubMed]
- Daves, M.H.; Hilsenbeck, S.G.; Lau, C.C.; Man, T.K. Meta-analysis of multiple microarray datasets reveals a common gene signature of metastasis in solid tumors. BMC Med. Genom. 2011, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, X.; Xie, Q.; Sun, H.; Huang, K.; Chen, H.; Wang, W.; Zhou, B.; Wei, X.; Zeng, D.; et al. CD146 interaction with integrin beta1 activates LATS1-YAP signaling and induces radiation-resistance in breast cancer cells. Cancer Lett. 2022, 546, 215856. [Google Scholar] [CrossRef] [PubMed]
- Rotman, J.; den Otter, L.A.S.; Bleeker, M.C.G.; Samuels, S.S.; Heeren, A.M.; Roemer, M.G.M.; Kenter, G.G.; Zijlmans, H.; van Trommel, N.E.; de Gruijl, T.D.; et al. PD-L1 and PD-L2 Expression in Cervical Cancer: Regulation and Biomarker Potential. Front. Immunol. 2020, 11, 596825. [Google Scholar] [CrossRef]
- Heeren, A.M.; Punt, S.; Bleeker, M.C.; Gaarenstroom, K.N.; van der Velden, J.; Kenter, G.G.; de Gruijl, T.D.; Jordanova, E.S. Prognostic effect of different PD-L1 expression patterns in squamous cell carcinoma and adenocarcinoma of the cervix. Mod. Pathol. 2016, 29, 753–763. [Google Scholar] [CrossRef]
- Zagorianakou, N.; Mitrogiannis, I.; Konis, K.; Makrydimas, S.; Mitrogiannis, L.; Makrydimas, G. The HPV-DNA Test in Pregnancy: A Review of the Literature. Cureus 2023, 15, e38619. [Google Scholar] [CrossRef]
- Leeson, S.; Alalade, R.; Singh, N.; Nieminen, P.; Cruickshank, M.; Carcopino, X.; Bergeron, C. Options for triage and implications for colposcopists within European HPV-based cervical screening programmes. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 258, 332–342. [Google Scholar] [CrossRef]
- Ronco, G.; Dillner, J.; Elfstrom, K.M.; Tunesi, S.; Snijders, P.J.; Arbyn, M.; Kitchener, H.; Segnan, N.; Gilham, C.; Giorgi-Rossi, P.; et al. Efficacy of HPV-based screening for prevention of invasive cervical cancer: Follow-up of four European randomised controlled trials. Lancet 2014, 383, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Maver, P.J.; Poljak, M. Primary HPV-based cervical cancer screening in Europe: Implementation status, challenges, and future plans. Clin. Microbiol. Infect. 2020, 26, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Calabro, G.E.; Riccardi, M.T.; D’Ambrosio, F.; Castagna, C.; Sapienza, M.; Millevolte, R.; Pellacchia, A.; Ricciardi, R.; de Vincenzo, R.P.; de Waure, C. Cervical cancer elimination in Italy: Current scenario and future endeavors for a value based prevention. Front. Public. Health 2022, 10, 1010237. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (N) | % (n/N) Negative Samples | % (n/N) Samples 1+ (Weak Intensity) | % (n/N) Samples 2+ (Medium Intensity) | % (n/N) Samples 3+ (High Intensity) | |
|---|---|---|---|---|---|
| Group A (CIN1) | 55 | 65.5% (36/55) | 12.7% (7/55) | 12.7% (7/55) | 9.1% (5/55) |
| Group B (CIN2) | 27 | 33.3% (9/27) | 11.1% (3/27) | 22.3% (6/27) | 33.3% (9/27) |
| Group C (CIN3) | 30 | 13.3% (4/30) | 13.3% (4/30) | 40% (12/30) | 33.4% (10/30) |
| Group D (squamous metaplasia with HPV+) | 42 | 83.4% (35/42) | 12% (5/42) | 4.8% (2/42) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schettino, M.T.; Preti, E.P.; Vietri, V.; Agrillo, N.; Iavazzo, N.; Fasulo, D.D.; De Franciscis, P.; Campitiello, M.R.; Vastarella, M.G.; Riemma, G.; et al. The Role of β1 Integrin/CD29 as a Potential Prognostic Factor for the Risk of Progression to Cervical Carcinoma in HPV-Associated Lesions. Medicina 2024, 60, 364. https://doi.org/10.3390/medicina60030364
Schettino MT, Preti EP, Vietri V, Agrillo N, Iavazzo N, Fasulo DD, De Franciscis P, Campitiello MR, Vastarella MG, Riemma G, et al. The Role of β1 Integrin/CD29 as a Potential Prognostic Factor for the Risk of Progression to Cervical Carcinoma in HPV-Associated Lesions. Medicina. 2024; 60(3):364. https://doi.org/10.3390/medicina60030364
Chicago/Turabian StyleSchettino, Maria Teresa, Eleonora Petra Preti, Valeria Vietri, Nadia Agrillo, Nicola Iavazzo, Diego Domenico Fasulo, Pasquale De Franciscis, Maria Rosaria Campitiello, Maria Giovanna Vastarella, Gaetano Riemma, and et al. 2024. "The Role of β1 Integrin/CD29 as a Potential Prognostic Factor for the Risk of Progression to Cervical Carcinoma in HPV-Associated Lesions" Medicina 60, no. 3: 364. https://doi.org/10.3390/medicina60030364
APA StyleSchettino, M. T., Preti, E. P., Vietri, V., Agrillo, N., Iavazzo, N., Fasulo, D. D., De Franciscis, P., Campitiello, M. R., Vastarella, M. G., Riemma, G., Gardella, B., & Murina, F. (2024). The Role of β1 Integrin/CD29 as a Potential Prognostic Factor for the Risk of Progression to Cervical Carcinoma in HPV-Associated Lesions. Medicina, 60(3), 364. https://doi.org/10.3390/medicina60030364