
ARHGAP21 Is Involved in the Carcinogenic Mechanism of Cholangiocarcinoma: A Study Based on Bioinformatic Analyses and Experimental Validation
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatics Data
2.2. Human Protein Atlas
2.3. cBioPortal for Cancer Genomics
2.4. GEPIA Online Website
2.5. LinkedOmics Database
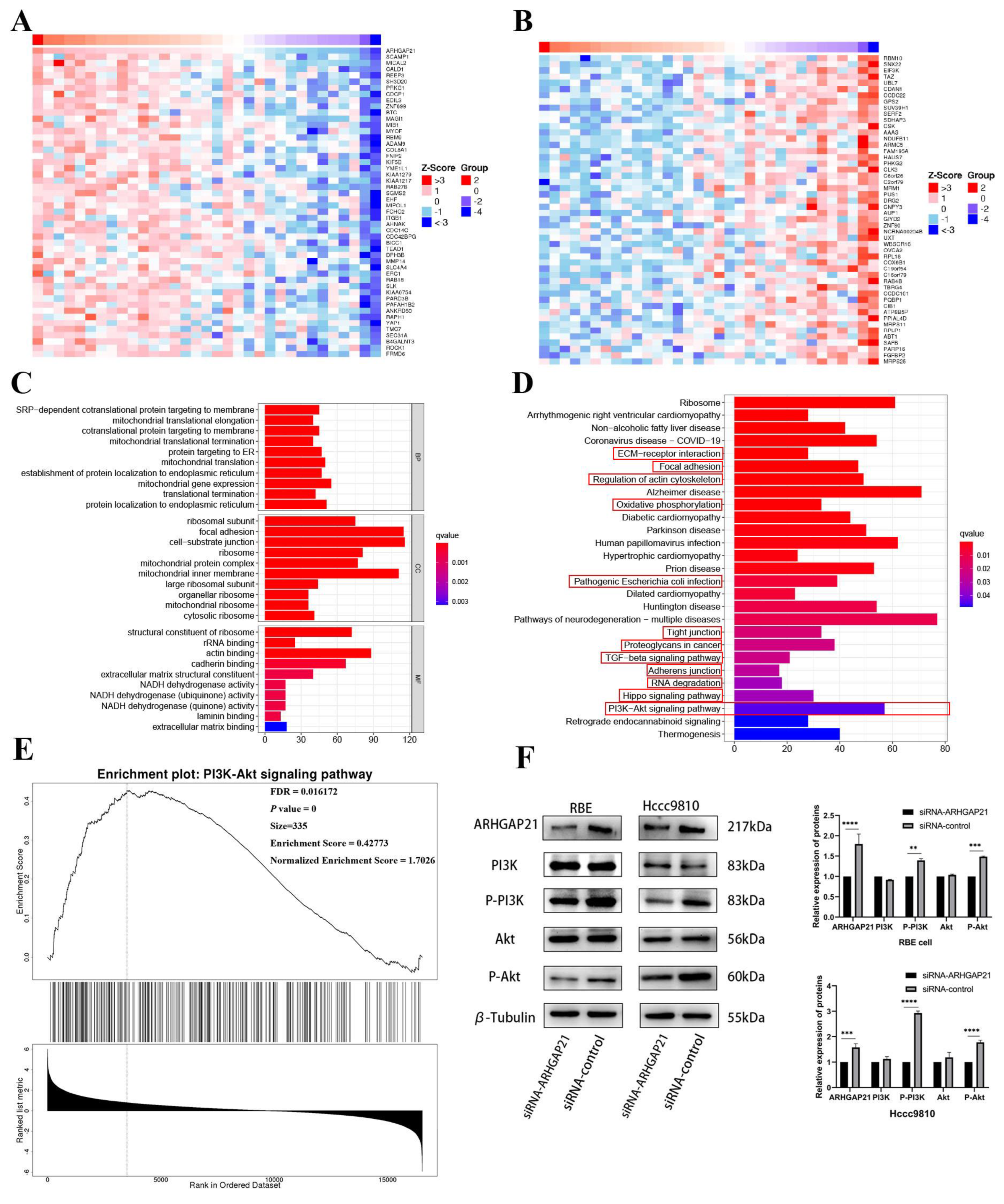
2.6. GO Enrichment Analysis and KEGG Enrichment Analysis
2.7. Cell Lines and Cell Culture
2.8. RNA Extraction and qRT-PCR Analysis
2.9. Cell Transfection
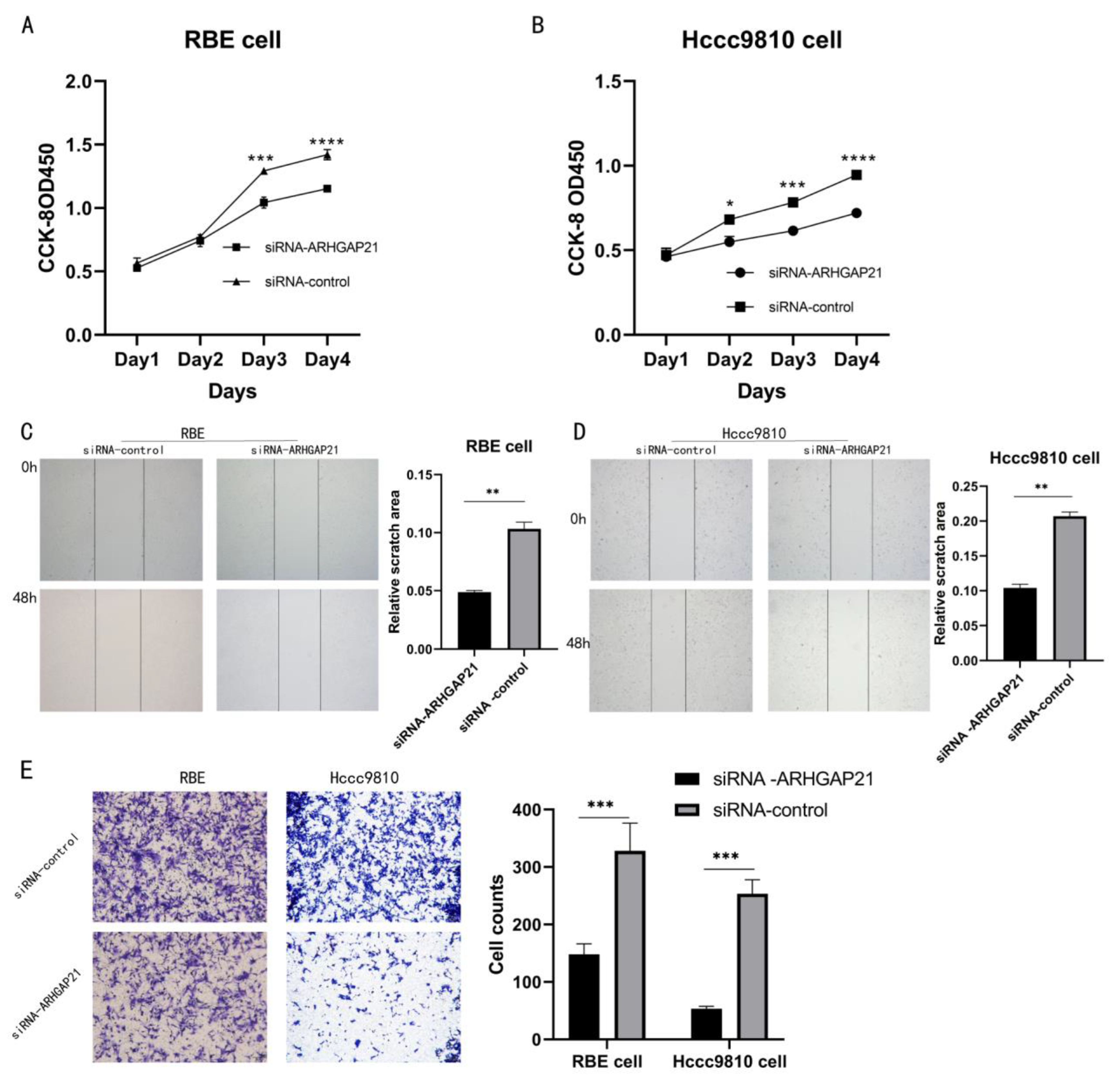
2.10. Cell Counting Kit-8 (CCK-8) Assay
2.11. Cell Scratch Assay
2.12. Transwell Migration Assay
2.13. Western Blot Assay
2.14. TIMER
3. Statistical Analysis
4. Results
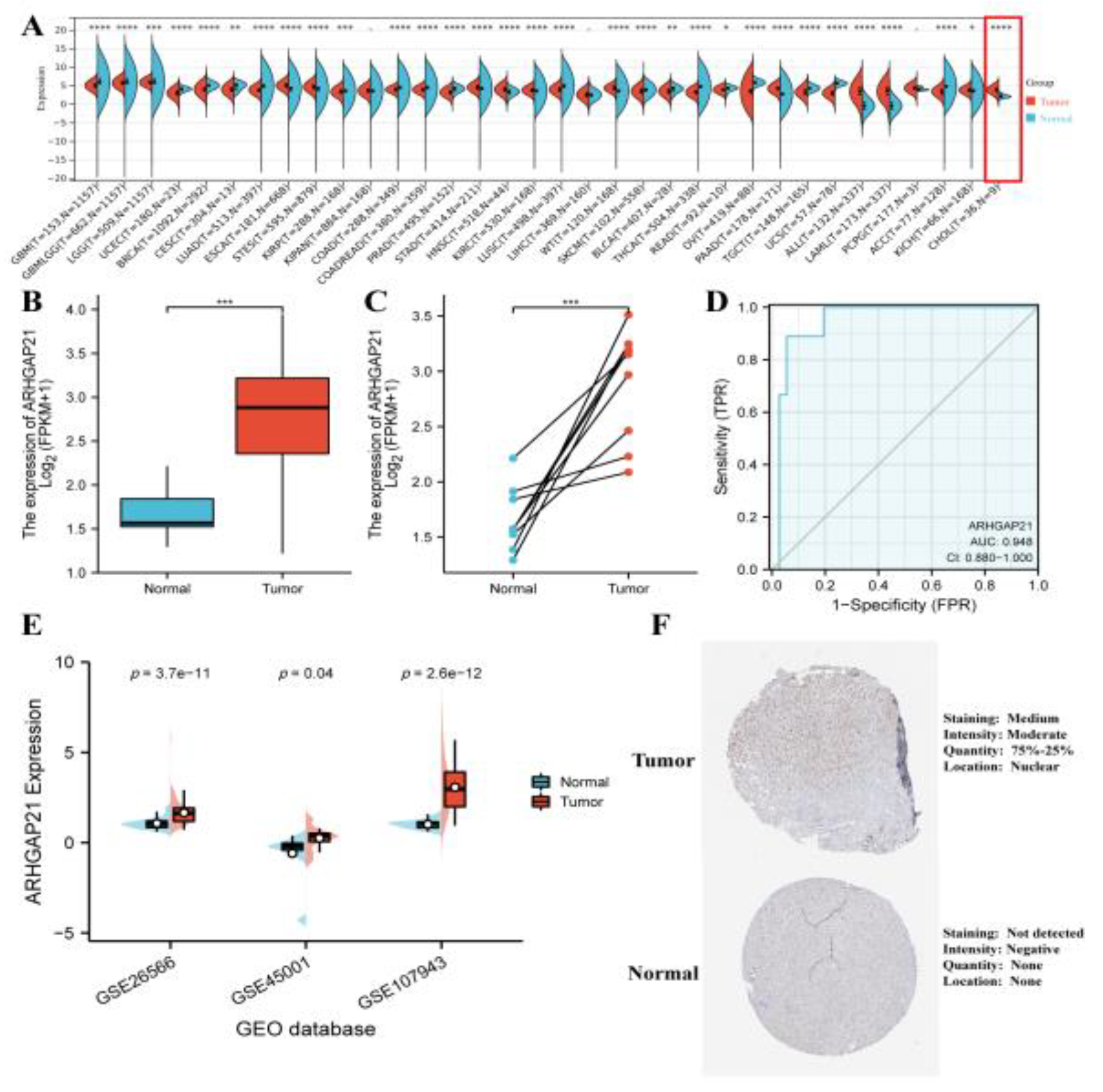
4.1. The Elevated Expression of ARHGAP21 in CCA
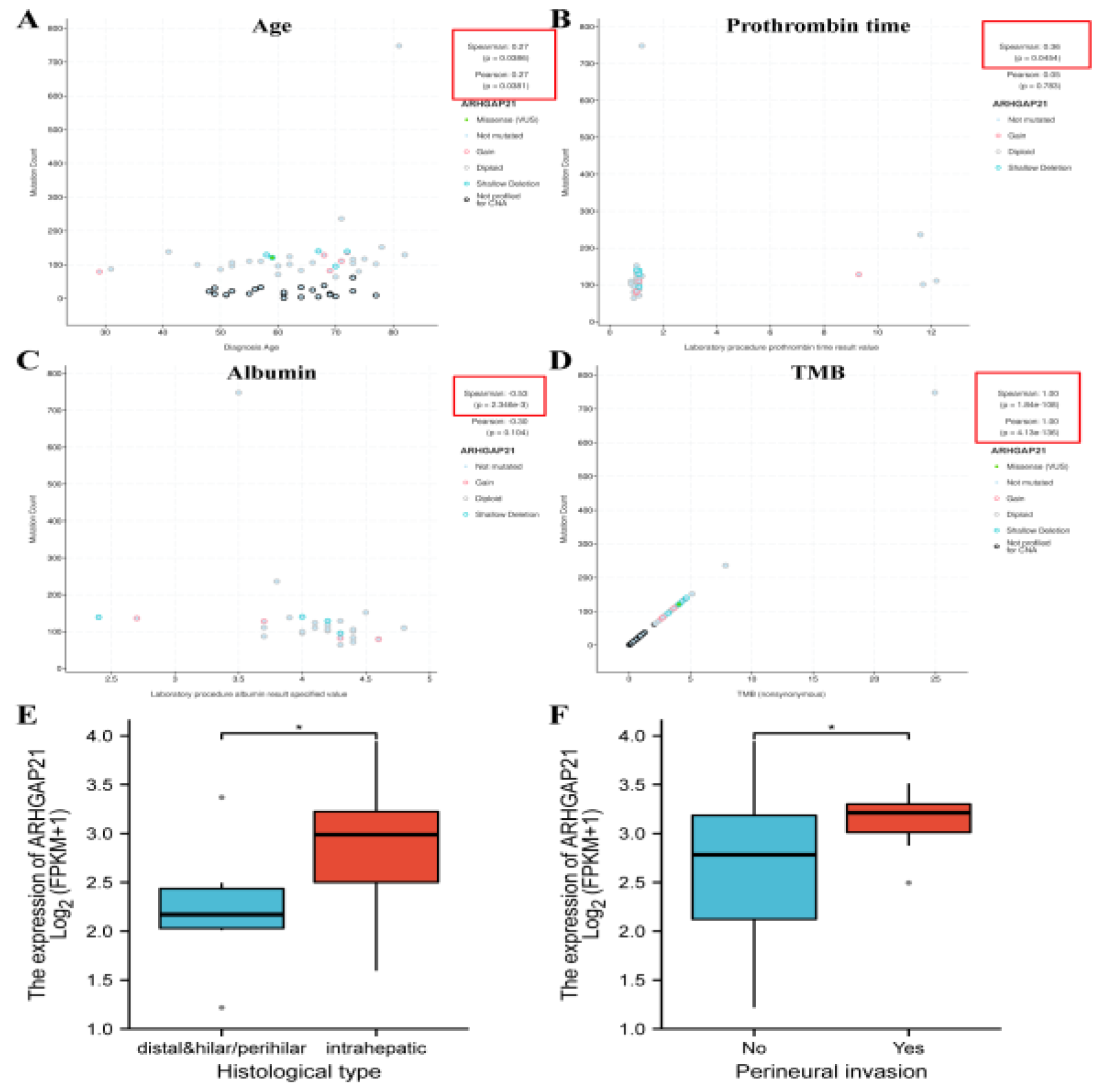
4.2. The Significant Correlation between ARHGAP21 and Clinical Characteristics in CCA Patients
4.3. The Prognostic Significance of ARHGAP21 in CCA Patients with Multiple Clinicopathological Features
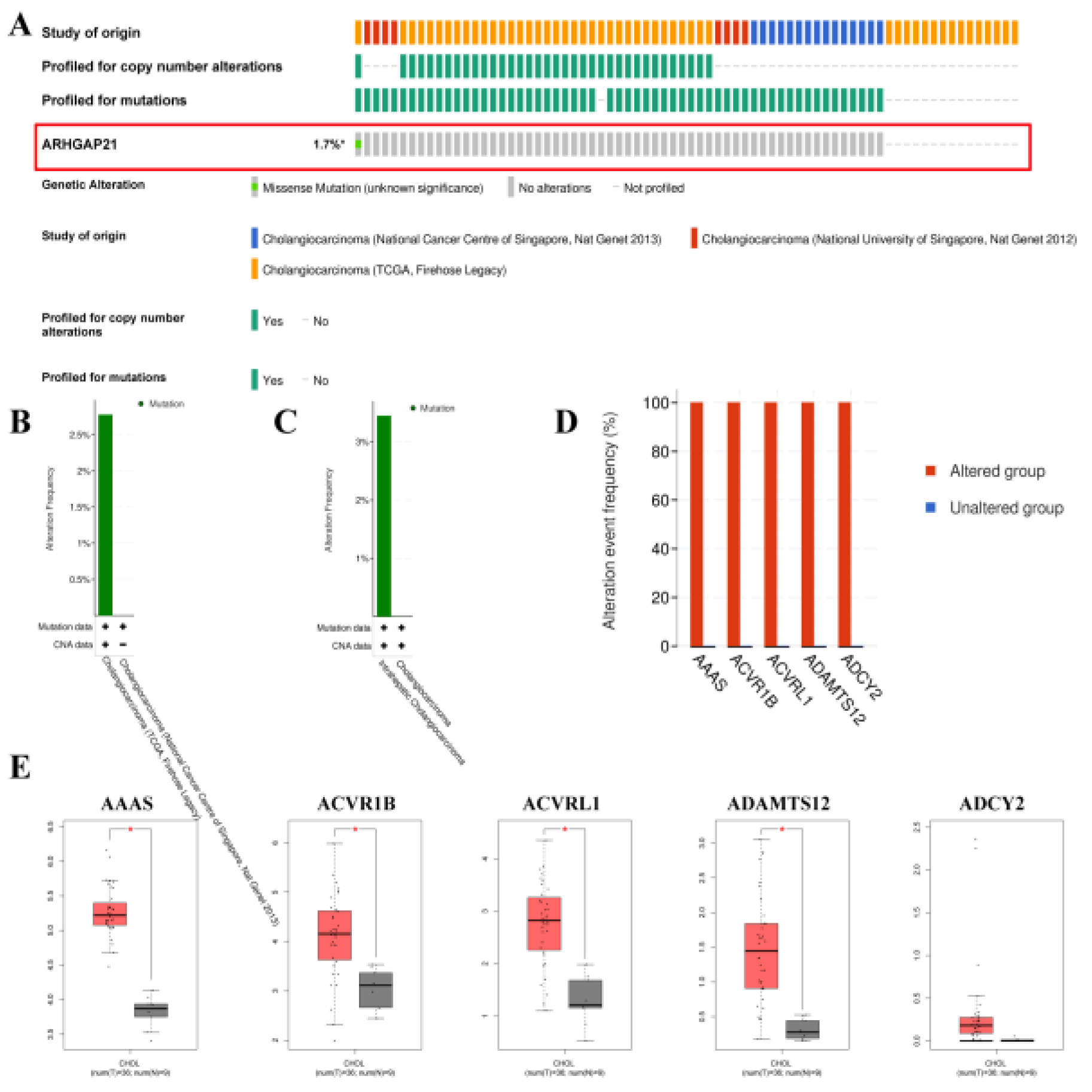
4.4. The Mutations of ARHGAP21 in CCA
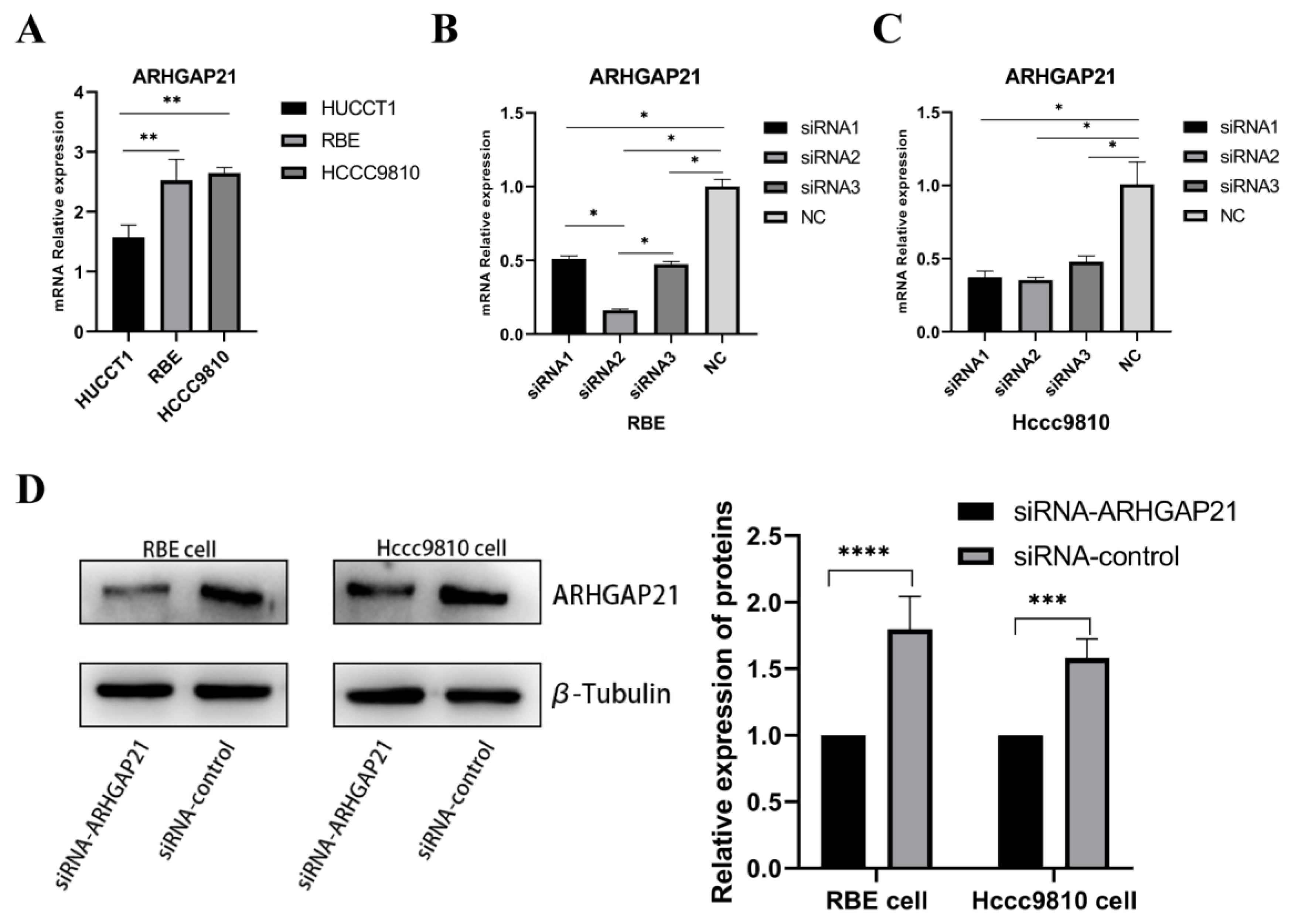
4.5. Detection of ARHGAP21 Expression of CCA Cell Lines and the Transfection Efficiency of siRNA-ARHGAP21
4.6. Downregulation of ARHGAP21 Inhibited the Proliferation and the Migration of CCA Cells
4.7. The Co-Expression Network of ARHGAP21 in CCA
4.8. GO and KEGG Enrichment Analysis
4.9. ARHGAP21 Promotes the Proliferation and Migration via PI3K/Akt Pathway in CCA
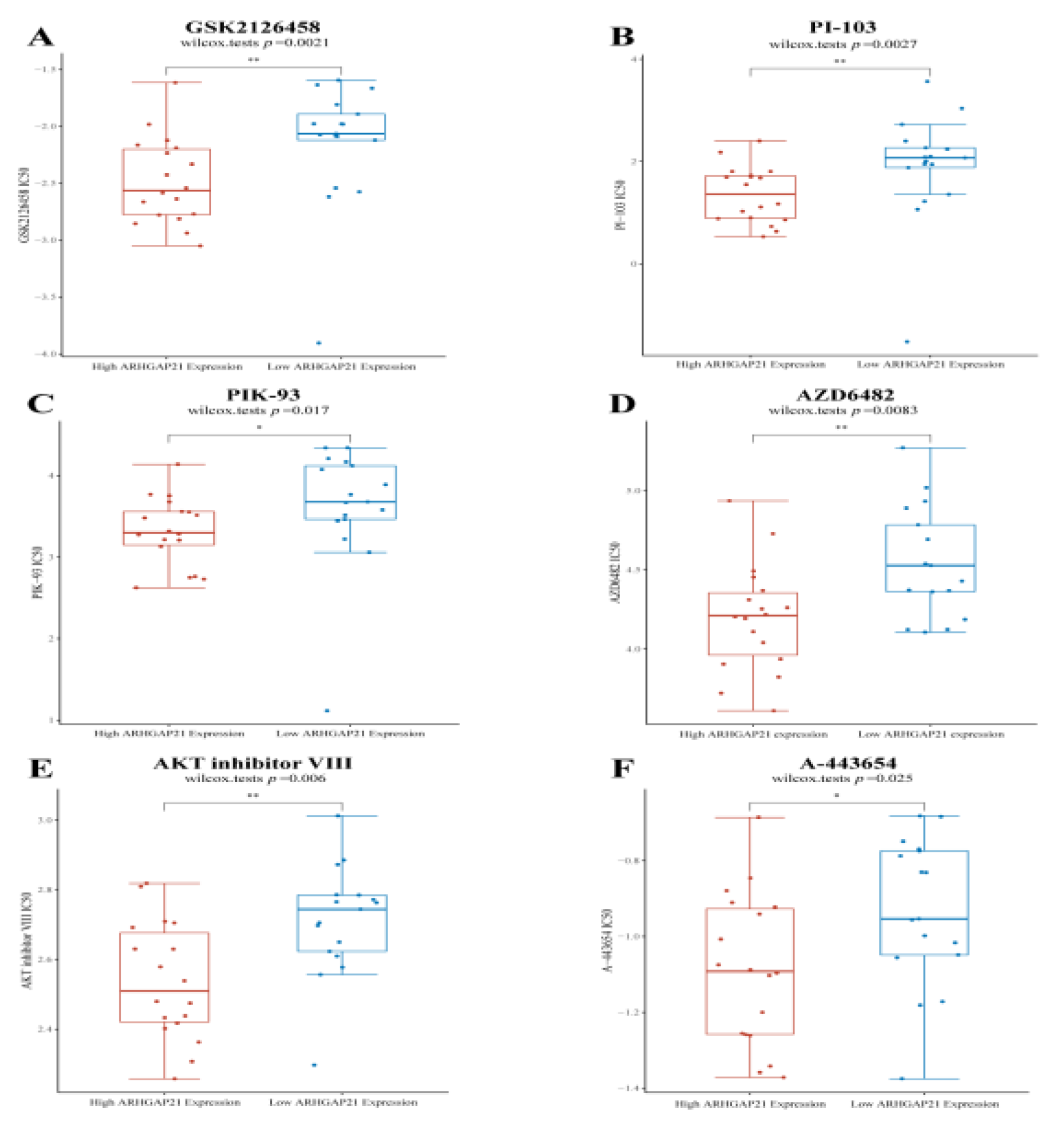
4.10. ARHGAP21 Is Positively Correlated with the PI3K/Akt Signaling Pathway Targeted Drugs
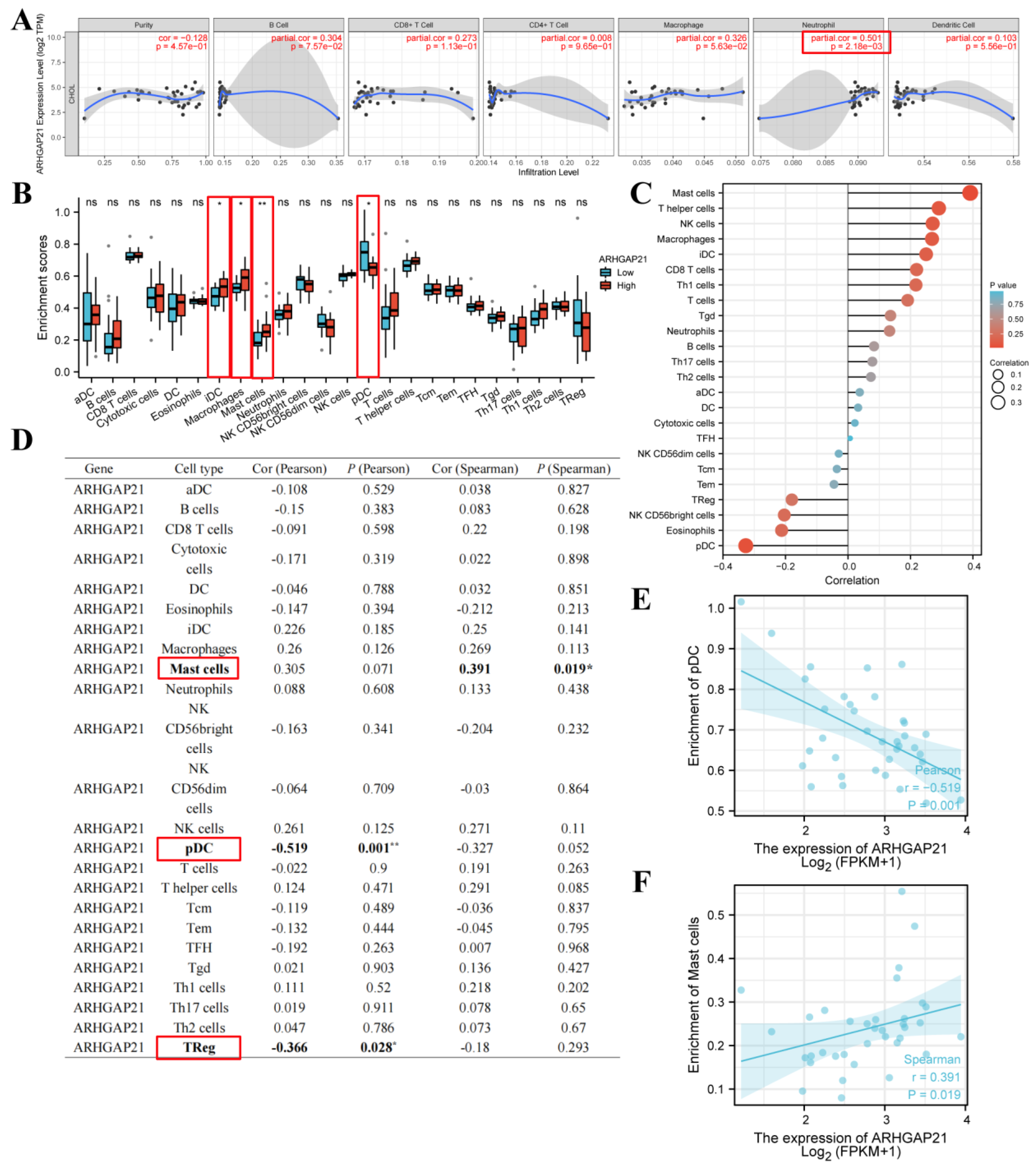
4.11. The Strong Relationship of ARHGAP21 and Infiltrating Immune Cells
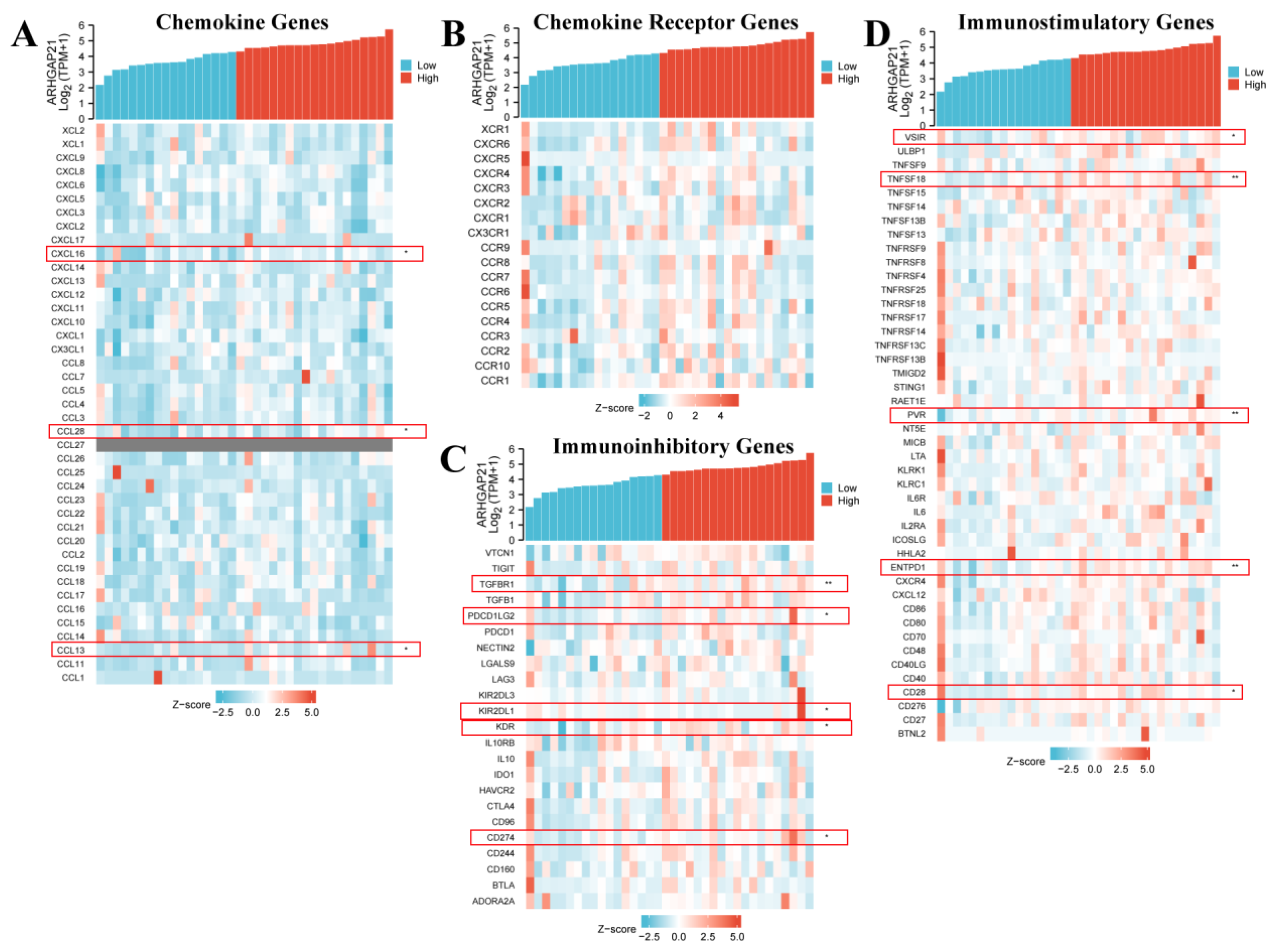
4.12. ARHGAP21 Was Correlated with Immune-related Genes
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Razumilava, N.; Gores, G.J. Cholangiocarcinoma. Lancet 2014, 383, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, C.; Dicker, D.; Pain, A.; Hamavid, H.; Moradi-Lakeh, M.; MacIntyre, M.F.; Allen, C.; Hansen, G.; Woodbrook, R.; Wolfe, C.; et al. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505. [Google Scholar] [CrossRef] [PubMed]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Mazzaferro, V.; Gorgen, A.; Roayaie, S.; Droz dit Busset, M.; Sapisochin, G. Liver resection and transplantation for intrahepatic cholangiocarcinoma. J. Hepatol. 2020, 72, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.K.; Bridgewater, J.; Gores, G.J.; Zhu, A.X. Systemic therapies for intrahepatic cholangiocarcinoma. J. Hepatol. 2020, 72, 353–363. [Google Scholar] [CrossRef]
- Sarcognato, S.; Sacchi, D.; Fassan, M.; Fabris, L.; Cadamuro, M.; Zanus, G.; Cataldo, I.; Capelli, P.; Baciorri, F.; Cacciatore, M.; et al. Cholangiocarcinoma. Pathologica 2021, 113, 158–169. [Google Scholar] [CrossRef]
- Rodrigues, P.M.; Olaizola, P.; Paiva, N.A.; Olaizola, I.; Agirre-Lizaso, A.; Landa, A.; Bujanda, L.; Perugorria, M.J.; Banales, J.M. Pathogenesis of Cholangiocarcinoma. Annu. Rev. Pathol. Mech. Dis. 2021, 16, 433–463. [Google Scholar] [CrossRef]
- Sirica, A.E.; Gores, G.J. Desmoplastic stroma and cholangiocarcinoma: Clinical implications and therapeutic targeting. Hepatology 2014, 59, 2397–2402. [Google Scholar] [CrossRef]
- Pichlmayr, R.; Weimann, A.; Klempnauer, J.; Oldhafer, K.J.; Maschek, H.; Tusch, G.; Ringe, B. Surgical reatment in Proximal Bile Duct Cancer: A Single-Center Experience. Ann. Surg. 1996, 224, 628–638. [Google Scholar] [CrossRef]
- Rosa, L.R.O.; Soares, G.M.; Silveira, L.R.; Boschero, A.C.; Barbosa-Sampaio, H.C.L. ARHGAP21 as a master regulator of multiple cellular processes. J. Cell Physiol. 2018, 233, 8477–8481. [Google Scholar] [CrossRef]
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [PubMed]
- Barcellos, K.S.A.; Bigarella, C.L.; Wagner, M.V.; Vieira, K.P.; Lazarini, M.; Langford, P.R.; Machado-Neto, J.A.; Call, S.G.; Staley, D.M.; Chung, J.Y.; et al. ARHGAP21 Protein, a New Partner of α-Tubulin Involved in Cell-Cell Adhesion Formation and Essential for Epithelial-Mesenchymal Transition. J. Biol. Chem. 2013, 288, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Lazarini, M.; Traina, F.; Machado-Neto, J.A.; Barcellos, K.S.A.; Moreira, Y.B.; Brandão, M.M.; Verjovski-Almeida, S.; Ridley, A.J.; Saad, S.T.O. ARHGAP21 is a RhoGAP for RhoA and RhoC with a role in proliferation and migration of prostate adenocarcinoma cells. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2013, 1832, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Bigarella, C.L.; Borges, L.; Costa, F.F.; Saad, S.T.O. ARHGAP21 modulates FAK activity and impairs glioblastoma cell migration. Biochim. Biophys. Acta BBA Mol. Cell Res. 2009, 1793, 806–816. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef]
- Wu, P.; Heins, Z.J.; Muller, J.T.; Katsnelson, L.; De Bruijn, I.; Abeshouse, A.; Schultz, N.; Fenyö, D.; Gao, J. Integration and analysis of CPTAC proteomics data in the context of cancer genomics in the cBioPortal. Mol. Cell. Proteom. 2019, 18, 1893–1898. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Massi, D.; Teng, M.W.L.; Mandala, M. PI3K-AKT-mTOR inhibition in cancer immunotherapy, redux. Semin. Cancer Biol. 2018, 48, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Guo, J.; Chen, L.; Yang, W.; Qu, X.; Cheng, Z. ARHGAP10, downregulated in ovarian cancer, suppresses tumorigenicity of ovarian cancer cells. Cell Death Dis. 2016, 7, e2157. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-L.; Yang, F.; Zhang, D.-H.; Hu, C.; Yang, S.; Chen, X.-Q. ARHGAP10 inhibits the epithelial–mesenchymal transition of non-small cell lung cancer by inactivating PI3K/Akt/GSK3β signaling pathway. Cancer Cell Int. 2021, 21, 320. [Google Scholar] [CrossRef]
- Sanchez Bassères, D.; Vedelago Tizzei, E.; Duarte, A.A.; Costa, F.F.; Teresinha Olalla Saad, S. ARHGAP10, a novel human gene coding for a potentially cytoskeletal Rho-GTPase activating protein. Biochem. Biophys. Res. Commun. 2002, 294, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Moujaber, O.; Stochaj, U. The Cytoskeleton as Regulator of Cell Signaling Pathways. Trends Biochem. Sci. 2020, 45, 96–107. [Google Scholar] [CrossRef]
- Zhang, L.; Luga, V.; Armitage, S.K.; Musiol, M.; Won, A.; Yip, C.M.; Plotnikov, S.V.; Wrana, J.L. A lateral signalling pathway coordinates shape volatility during cell migration. Nat. Commun. 2016, 7, 11714. [Google Scholar] [CrossRef]
- Noorolyai, S.; Shajari, N.; Baghbani, E.; Sadreddini, S.; Baradaran, B. The relation between PI3K/AKT signalling pathway and cancer. Gene 2019, 698, 120–128. [Google Scholar] [CrossRef]
- Lien, E.C.; Dibble, C.C.; Toker, A. PI3K signaling in cancer: Beyond AKT. Curr. Opin. Cell Biol. 2017, 45, 62–71. [Google Scholar] [CrossRef]
- LoRusso, P.M. Inhibition of the PI3K/AKT/mTOR Pathway in Solid Tumors. J. Clin. Oncol. 2016, 34, 3803–3815. [Google Scholar] [CrossRef]
- Akbarzadeh, M.; Mihanfar, A.; Akbarzadeh, S.; Yousefi, B.; Majidinia, M. Crosstalk between miRNA and PI3K/AKT/mTOR signaling pathway in cancer. Life Sci. 2021, 285, 119984. [Google Scholar] [CrossRef]
- Feng, H.; Qiu, L.; Shi, Z.; Sheng, Y.; Zhao, P.; Zhou, D.; Li, F.; Yu, H.; You, Y.; Wang, H.; et al. Modulation of intracellular kinase signaling to improve TIL stemness and function for adoptive cell therapy. Cancer Med. 2022. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, J.; Cai, Y.; Fu, S.; Zhang, N.; Fu, X.; Li, L. IFN-γ-mediated inhibition of lung cancer correlates with PD-L1 expression and is regulated by PI3K-AKT signaling: IFN-γ in lung adenocarcinoma. Int. J. Cancer 2018, 143, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zeng, Y.; Qu, Q.; Zhu, J.; Liu, Z.; Ning, W.; Zeng, H.; Zhang, N.; Du, W.; Chen, C.; et al. PD-L1 induced by IFN-γ from tumor-associated macrophages via the JAK/STAT3 and PI3K/AKT signaling pathways promoted progression of lung cancer. Int. J. Clin. Oncol. 2017, 22, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-Q.; Zhou, Z.-J.; Luo, C.-B.; Xin, H.-Y.; Li, J.; Yu, S.-Y.; Zhou, S.-L. Peritumoral plasmacytoid dendritic cells predict a poor prognosis for intrahepatic cholangiocarcinoma after curative resection. Cancer Cell Int. 2020, 20, 582. [Google Scholar] [CrossRef] [PubMed]
- González, M.I.; Vannan, D.T.; Eksteen, B.; Flores-Sotelo, I.; Reyes, J.L. Mast Cells in Immune-Mediated Cholangitis and Cholangiocarcinoma. Cells 2022, 11, 375. [Google Scholar] [CrossRef]
- Tamma, R.; Annese, T.; Ruggieri, S.; Brunetti, O.; Longo, V.; Cascardi, E.; Mastropasqua, M.G.; Maiorano, E.; Silvestris, N.; Ribatti, D. Inflammatory cells infiltrate and angiogenesis in locally advanced and metastatic cholangiocarcinoma. Eur. J. Clin. Investig. 2019, 49, e13087. [Google Scholar] [CrossRef]
- Korbecki, J.; Bajdak-Rusinek, K.; Kupnicka, P.; Kapczuk, P.; Simińska, D.; Chlubek, D.; Baranowska-Bosiacka, I. The Role of CXCL16 in the Pathogenesis of Cancer and Other Diseases. Int. J. Mol. Sci. 2021, 22, 3490. [Google Scholar] [CrossRef]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.-P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and Treg cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef]
- He, B.; Xu, T.; Pan, B.; Pan, Y.; Wang, X.; Dong, J.; Sun, H.; Xu, X.; Liu, X.; Wang, S. Polymorphisms of TGFBR1, TLR4 are associated with prognosis of gastric cancer in a Chinese population. Cancer Cell Int. 2018, 18, 191. [Google Scholar] [CrossRef]
- Levovitz, C.; Chen, D.; Ivansson, E.; Gyllensten, U.; Finnigan, J.P.; Alshawish, S.; Zhang, W.; Schadt, E.E.; Posner, M.R.; Genden, E.M.; et al. TGFβ Receptor 1: An Immune Susceptibility Gene in HPV-Associated Cancer. Cancer Res. 2014, 74, 6833–6844. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, Z.; Hu, R.; Wang, Y.; Wang, Y.; Su, Z.; Zhang, X.; Yang, J.; Mei, M.; Ren, Y.; et al. PD-L2 glycosylation promotes immune evasion and predicts anti-EGFR efficacy. J. Immunother. Cancer 2021, 9, e002699. [Google Scholar] [CrossRef] [PubMed]
- Dermani, F.K.; Samadi, P.; Rahmani, G.; Kohlan, A.K.; Najafi, R. PD-1/PD-L1 immune checkpoint: Potential target for cancer therapy. J. Cell Physiol. 2019, 234, 1313–1325. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D.; et al. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef]
- Gou, Q.; Dong, C.; Xu, H.; Khan, B.; Jin, J.; Liu, Q.; Shi, J.; Hou, Y. PD-L1 degradation pathway and immunotherapy for cancer. Cell Death Dis. 2020, 11, 955. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Pan, Y.; Fei, Q.; Lin, Y.; Zhou, Y.; Liu, Y.; Guan, H.; Yu, X.; Lin, X.; Lu, F.; et al. Prognostic significance and therapeutic potential of the immune checkpoint VISTA in pancreatic cancer. J. Cancer Res. Clin. Oncol. 2021, 147, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Tai, R.; Wu, Y.; Yang, S.; Wang, J.; Yu, X.; Lei, L.; Shan, Z.; Li, N. The expression and immunoregulation of immune checkpoint molecule VISTA in autoimmune diseases and cancers. Cytokine Growth Factor Rev. 2020, 52, 1–14. [Google Scholar] [CrossRef]
- Baltz, K.M.; Krusch, M.; Bringmann, A.; Brossart, P.; Mayer, F.; Kloss, M.; Baessler, T.; Kumbier, I.; Peterfi, A.; Kupka, S.; et al. Cancer immunoediting by GITR (glucocorticoid-induced TNF-related protein) ligand in humans: NK cell/tumor cell interactions. FASEB J. 2007, 21, 2442–2454. [Google Scholar] [CrossRef]
- Zhou, X.; Du, J.; Wang, H.; Chen, C.; Jiao, L.; Cheng, X.; Zhou, X.; Chen, S.; Gou, S.; Zhao, W.; et al. Repositioning liothyronine for cancer immunotherapy by blocking the interaction of immune checkpoint TIGIT/PVR. Cell Commun. Signal 2020, 18, 142. [Google Scholar] [CrossRef]
- Bastid, J.; Cottalorda-Regairaz, A.; Alberici, G.; Bonnefoy, N.; Eliaou, J.-F.; Bensussan, A. ENTPD1/CD39 is a promising therapeutic target in oncology. Oncogene 2013, 32, 1743–1751. [Google Scholar] [CrossRef]
- Hui, E.; Cheung, J.; Zhu, J.; Su, X.; Taylor, M.J.; Wallweber, H.A.; Sasmal, D.K.; Huang, J.; Kim, J.M.; Mellman, I.; et al. T cell costimulatory receptor CD28 is a primary target for PD-1–mediated inhibition. Science 2017, 355, 1428–1433. [Google Scholar] [CrossRef]
- Xia, S.; Chen, Q.; Niu, B. CD28: A New Drug Target for Immune Disease. Curr. Drug Targets 2020, 21, 589–598. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | Sequence (5′–3′) |
|---|---|
| ARHGAP21 F | TTGAGCAAACAGCAAACCAG |
| ARHGAP21 R | GCAACATCTGTTGGTGATGG |
| GAPDH F | TGAAGGTCGGAGTCAACGGATTTGGT |
| GAPDH R | CATGTGGGCCATGAGGTCCACCAC |
| Clinical Characteristic | OS | DSS | ||||
|---|---|---|---|---|---|---|
| N | HR | p | N | HR | p | |
| T stage | ||||||
| T1 | 19 | 0.32 (0.06−1.60) | 0.167 | 19 | 0.32 (0.06−1.60) | 0.167 |
| T2 | 12 | 0.11 (0.01−0.94) | 0.044 | 12 | 0.13 (0.01−1.22) | 0.074 |
| T3 | 5 | - | - | 5 | - | - |
| T1 + T2 | 31 | 0.25 (0.08−0.77) | 0.016 | 31 | 0.28 (0.09−0.90) | 0.033 |
| T2 + T3 | 17 | 0.30 (0.08−1.19) | 0.087 | 17 | 0.39 (0.09−1.66) | 0.202 |
| N stage | ||||||
| N0 | 26 | 0.22 (0.05−1.03) | 0.055 | 26 | 0.22 (0.05−1.03) | 0.055 |
| N1 | 5 | - | - | 5 | - | - |
| N0 + N1 | 31 | 0.25 (0.07−0.90) | 0.034 | 31 | 0.27 (0.07−0.99) | 0.049 |
| M stage | ||||||
| M0 | 28 | 0.41 (0.12−1.34) | 0.14 | 28 | 0.41 (0.12−1.34) | 0.14 |
| M1 | 5 | - | - | 5 | - | - |
| M0 + M1 | 33 | 0.28 (0.09−0.87) | 0.028 | 33 | 0.30 (0.10−0.95) | 0.041 |
| TNM stage | ||||||
| 1 | 19 | 0.32 (0.06−1.60) | 0.167 | 19 | 0.32 (0.06−1.60) | 0.167 |
| 2 | 9 | - | - | 9 | - | - |
| 3 | 1 | - | - | 1 | - | - |
| 4 | 7 | - | - | 7 | - | - |
| 1 + 2 | 28 | 0.23 (0.06−0.87) | 0.03 | 28 | 0.26 (0.07−0.96) | 0.043 |
| 3 + 4 | 8 | - | - | 8 | - | - |
| Gender | ||||||
| Male | 16 | 0.44 (0.08−2.27) | 0.325 | 16 | 0.500 (0.09−2.76) | 0.428 |
| Female | 20 | 0.35 (0.09−1.38) | 0.135 | 20 | 0.41 (0.10−1.64) | 0.205 |
| Race | ||||||
| Asian | 3 | - | - | 3 | - | - |
| Black | 2 | - | - | 2 | - | - |
| White | 31 | 0.30 (0.09−0.94) | 0.039 | 31 | 0.35 (0.11−1.14) | 0.082 |
| Age | ||||||
| ≤65 | 17 | 0.37 (0.09−1.50) | 0.164 | 17 | 0.37 (0.09−1.50) | 0.164 |
| >65 | 19 | 0.24 (0.05−1.20) | 0.083 | 19 | 0.33 (0.06−1.80) | 0.199 |
| BMI | - | 16 | - | - | ||
| ≤25 | 10 | 0.33 (0.06−1.88) | 0.212 | 10 | 0.42 (0.07−2.57) | 0.345 |
| >25 | 25 | 0.39 (0.10−1.55) | 0.183 | 25 | 0.45 (0.11−1.83) | 0.265 |
| Histological type | ||||||
| distal | 2 | - | - | 2 | - | - |
| perihilar | 4 | - | - | 4 | - | - |
| intrahepatic | 30 | 0.35 (0.11−1.11) | 0.075 | 30 | 0.38 (0.12−1.24) | 0.109 |
| CA19-9 | ||||||
| abnormal | 16 | 0.91 (0.20−4.10) | 0.9 | 16 | 1.17 (0.23−5.89) | 0.849 |
| normal | 14 | 0.10 (0.01−0.90) | 0.04 | 14 | 0.12 (0.01−1.03) | 0.054 |
| Vascular invasion | ||||||
| yes | 5 | - | - | 5 | - | - |
| no | 29 | 0.26 (0.07−0.96) | 0.042 | 29 | 0.29 (0.08−1.07) | 0.063 |
| Perineural invasion | ||||||
| Yes | 7 | - | - | 7 | - | - |
| No | 26 | 0.19 (0.04−0.88) | 0.034 | 26 | 0.21 (0.04−0.98) | 0.047 |
| Gene Type | Gene | Spearman | |
|---|---|---|---|
| cor | p | ||
| Chemokine genes | CXCL16 | 0.34 | 0.043 |
| CCL28 | 0.403 | 0.015 | |
| CCL13 | 0.359 | 0.031 | |
| Immunoinhibitory genes | TGFBR1 | 0.53 | 0.001 |
| PDCD1LG2 | 0.425 | 0.01 | |
| KIR2DL1 | −0.385 | 0.02 | |
| KDR | 0.381 | 0.023 | |
| CD274 | 0.372 | 0.026 | |
| Immunostimulatory genes | VSIR | 0.395 | 0.018 |
| TNFSF18 | 0.493 | 0.002 | |
| PVR | 0.484 | 0.003 | |
| ENTPD1 | 0.45 | 0.006 | |
| CD28 | 0.388 | 0.02 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wu, S.; Wang, G.; Yang, Z.; Zhang, Y.; Zhu, C.; Qin, X. ARHGAP21 Is Involved in the Carcinogenic Mechanism of Cholangiocarcinoma: A Study Based on Bioinformatic Analyses and Experimental Validation. Medicina 2023, 59, 139. https://doi.org/10.3390/medicina59010139
Wang Z, Wu S, Wang G, Yang Z, Zhang Y, Zhu C, Qin X. ARHGAP21 Is Involved in the Carcinogenic Mechanism of Cholangiocarcinoma: A Study Based on Bioinformatic Analyses and Experimental Validation. Medicina. 2023; 59(1):139. https://doi.org/10.3390/medicina59010139
Chicago/Turabian StyleWang, Zhihuai, Siyuan Wu, Gaochao Wang, Zhen Yang, Yinjie Zhang, Chunfu Zhu, and Xihu Qin. 2023. "ARHGAP21 Is Involved in the Carcinogenic Mechanism of Cholangiocarcinoma: A Study Based on Bioinformatic Analyses and Experimental Validation" Medicina 59, no. 1: 139. https://doi.org/10.3390/medicina59010139
APA StyleWang, Z., Wu, S., Wang, G., Yang, Z., Zhang, Y., Zhu, C., & Qin, X. (2023). ARHGAP21 Is Involved in the Carcinogenic Mechanism of Cholangiocarcinoma: A Study Based on Bioinformatic Analyses and Experimental Validation. Medicina, 59(1), 139. https://doi.org/10.3390/medicina59010139