

Characterization of Microscopic Multicellular Foci in Grossly Normal Renal Parenchyma of Von Hippel-Lindau Kidney


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Sample Collection
2.2. Immunohistochemistry
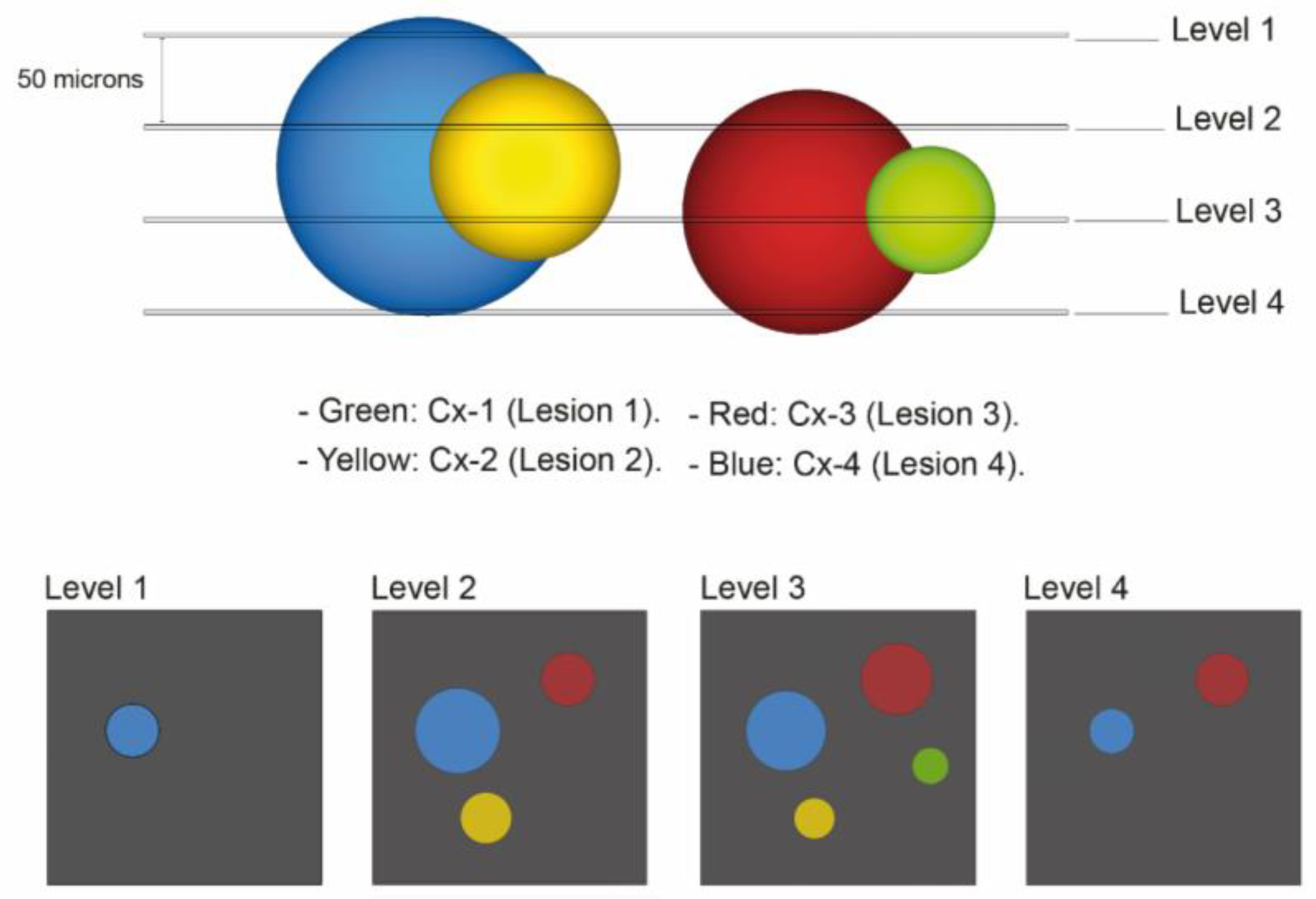
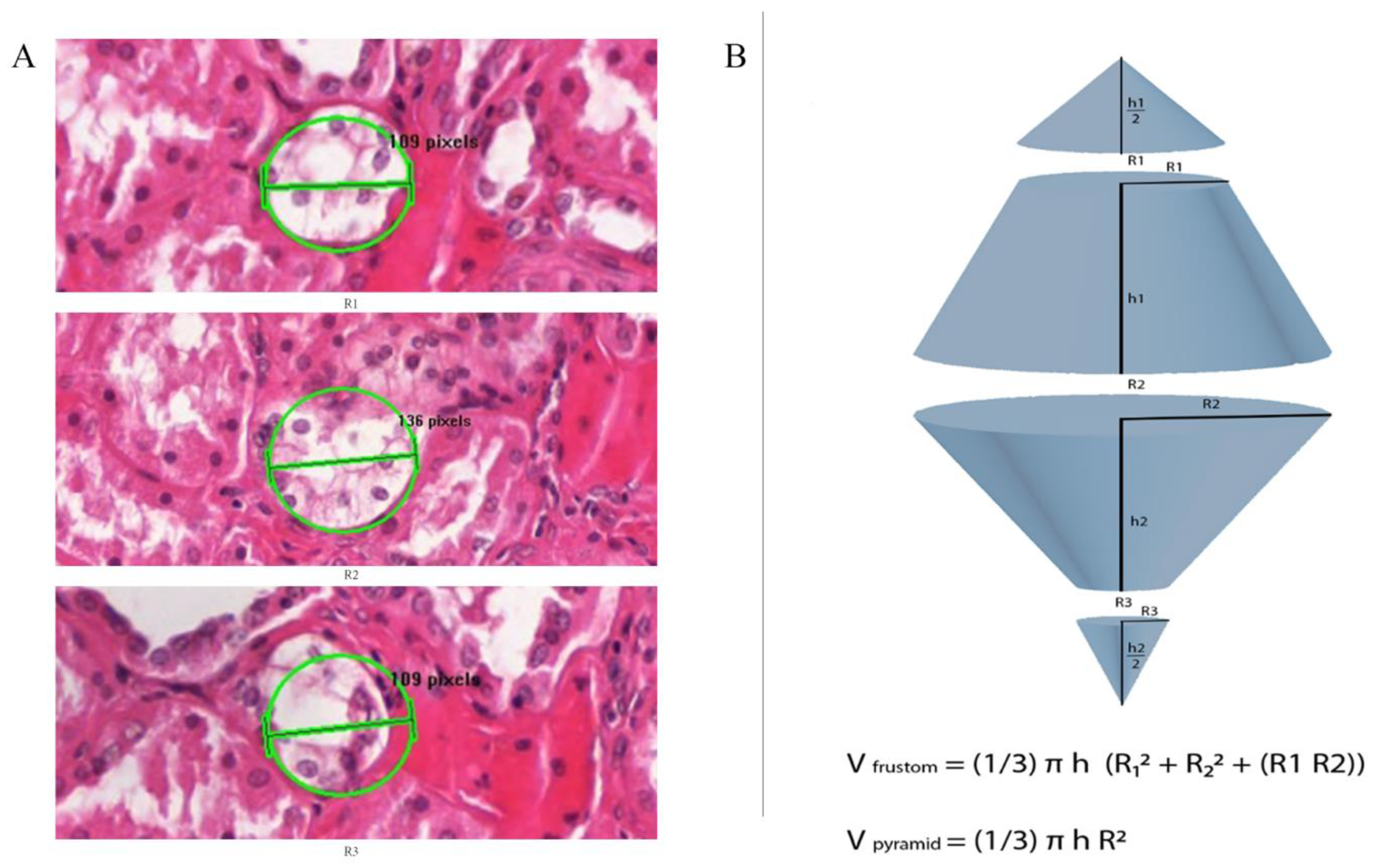
2.3. 3Dimensional Analysis
2.4. First Quantitative Approach Applied
2.5. Second Quantitative Approach Applied
2.6. Third Quantitative Approach Applied
3. Results
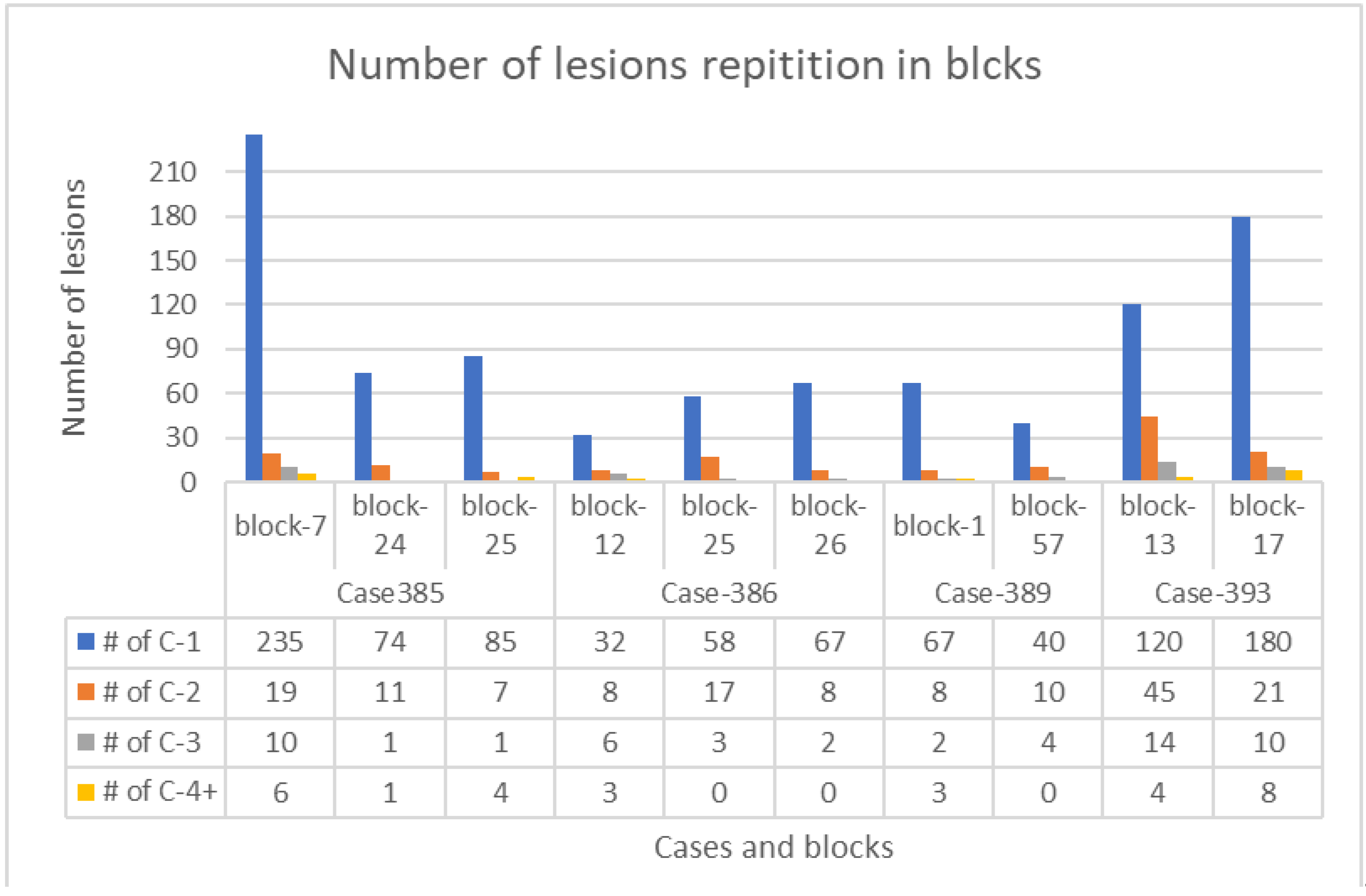
3.1. Lesions Account
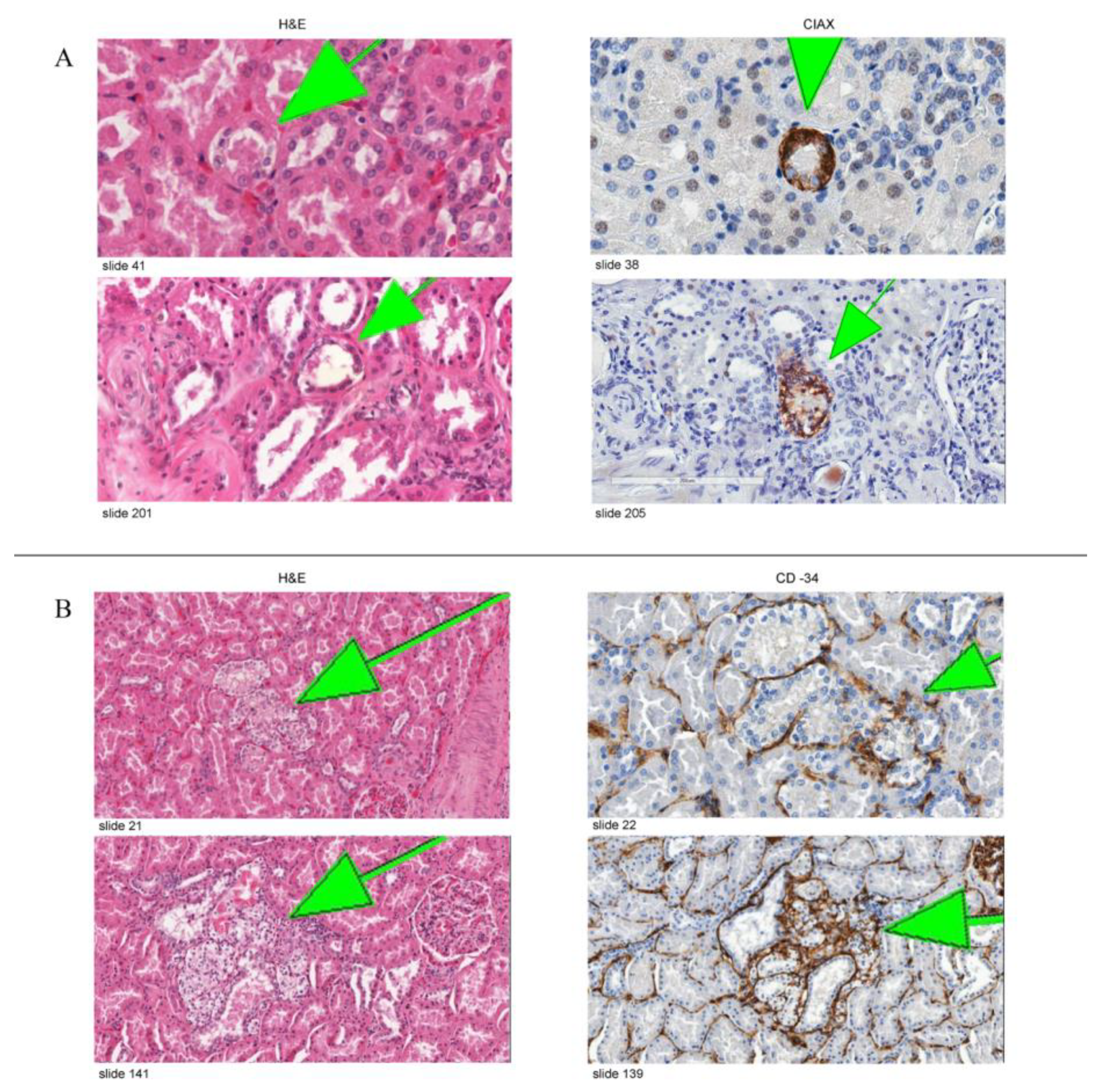
3.2. Immunohistochemical Expression
3.3. Cluster Volumes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation and Annotations
| VHL kidney | Kidney of von Hippel-Lindau disease patient |
| RCCC | Renal clear cell carcinoma |
| CAIX | Carbonic anhydrase IX |
| H&E | Hematoxylin and eosin |
| pVHL | VHL protein |
| VEGF | Vascular endothelial growth factor |
| EPO | Erythropoietin |
| PDGF | Platelet-derived growth factor |
| GLUT-1 | Glucose-transporter 1 |
| HIF | Hypoxia-inducible factor |
References
- Huntoon, K.; Oldfield, E.H.; Lonser, R.R. Dr. Arvid Lindau and discovery of von Hippel-Lindau disease. J. Neurosurg. JNS 2015, 123, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.M.; Cavallerano, J.D.; Silva, P.S. Von hippel-lindau disease: A genetic and clinical review. Semin Ophthalmol. 2013, 28, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Richards, F.M.; Maher, E.R.; Latif, F.; Phipps, M.E.; Tory, K.; Lush, M.; Crossey, P.A.; Oostra, B.; Enblad, P.; Gustavson, K.H. Detailed genetic mapping of the von Hippel-Lindau disease tumour suppressor gene. J. Med. Genet. 1993, 30, 104–107. [Google Scholar] [CrossRef][Green Version]
- Kaelin, W.G., Jr. Molecular basis of the VHL hereditary cancer syndrome. Nat. Rev. Cancer. 2002, 2, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Knudson Alfred, G. Mutation and Cancer: Statistical Study of Retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Latif, F.; Tory, K.; Gnarra, J.; Yao, M.; Duh, F.M.; Orcutt, M.L.; Stackhouse, T.; Kuzmin, I.; Modi, W.; Lerman, M.I. Identification of the von Hippel-Lindau disease tumor suppressor gene. Science 1993, 260, 1317–1320. [Google Scholar] [CrossRef] [PubMed]
- Lonser, R.R.; Glenn, G.M.; Walther, M.; Chew, E.Y.; Libutti, S.K.; Linehan, W.M.; Oldfield, E.H. von Hippel-Lindau disease. Lancet 2003, 361, 2059–2567. [Google Scholar] [CrossRef]
- Glasker, S.; Tran, M.G.; Shively, S.B.; Ikejiri, B.; Lonser, R.R.; Maxwell, P.H.; Zhuang, Z.; Oldfield, E.H.; Vortmeyer, A.O. Epididymal cystadenomas and epithelial tumourlets: Effects of VHL deficiency on the human epididymis. J. Pathol. 2006, 210, 32–41. [Google Scholar] [CrossRef]
- Mandriota, S.J.; Turner, K.J.; Davies, D.R.; Murray, P.G.; Morgan, N.V.; Sowter, H.M.; Wykoff, C.C.; Maher, E.R.; Harris, A.L.; Ratcliffe, P.J.; et al. HIF activation identifies early lesions in VHL kidneys: Evidence for site-specific tumor suppressor function in the nephron. Cancer Cell 2002, 1, 459–468. [Google Scholar] [CrossRef]
- Neumann, H.P.; Eggert, H.R.; Weigel, K.; Friedburg, H.; Wiestler, O.D.; Schollmeyer, P. Hemangioblastomas of the central nervous system. A 10-year study with special reference to von Hippel-Lindau syndrome. J. Neurosurg. 1989, 70, 24–30. [Google Scholar] [CrossRef]
- Vortmeyer, A.O.; Yuan, Q.; Lee, Y.S.; Zhuang, Z.; Oldfield, E.H. Developmental effects of von Hippel-Lindau gene deficiency. Ann. Neurol. 2004, 55, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Glasker, S.; Vergauwen, E.; Koch, C.A.; Kutikov, A.; Vortmeyer, A.O. Von Hippel-Lindau disease: Current challenges and future prospects. OncoTargets Ther. 2020, 13, 5669–5690. [Google Scholar] [CrossRef] [PubMed]
- Lubensky, I.A.; Pack, S.; Ault, D.; Vortmeyer, A.O.; Libutti, S.K.; Choyke, P.L.; Walther, M.M.; Linehan, W.M.; Zhuang, Z. Multiple neuroendocrine tumors of the pancreas in von Hippel-Lindau disease patients: Histopathological and molecular genetic analysis. Am. J. Pathol. 1998, 153, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Vortmeyer, A.O.; Alomari, A.K. Pathology of the Nervous System in Von Hippel-Lindau Disease. J. Kidney Cancer VHL 2015, 2, 114–129. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Anglard, P.; Tory, K.; Brauch, H.; Weiss, G.H.; Latif, F.; Merino, M.J.; Lerman, M.I.; Zbar, B.; Linehan, W.M. Molecular analysis of genetic changes in the origin and development of renal cell carcinoma. Cancer Res. 1991, 51, 1071–1077. [Google Scholar] [PubMed]
- Seizinger, B.R.; Rouleau, G.A.; Ozelius, L.J.; Lane, A.H.; Farmer, G.E.; Lamiell, J.M.; Haines, J.; Yuen, J.W.M.; Collins, D.; Majoor-Krakauer, D.; et al. Von Hippel-Lindau disease maps to the region of chromosome 3 associated with renal cell carcinoma. Nature 1988, 332, 268–269. [Google Scholar] [CrossRef] [PubMed]
- Tory, K.; Brauch, H.; Linehan, M.; Barba, D.; Oldfield, E.; Filling-Katz, M.; Seizinger, B.; Nakamura, Y.; White, R.; Marshall, F.F.; et al. Specific genetic change in tumors associated with von Hippel-Lindau disease. J. Natl. Cancer Inst. 1989, 81, 1097–1101. [Google Scholar] [CrossRef]
- Zbar, B.; Brauch, H.; Talmadge, C.; Linehan, M. Loss of alleles of loci on the short arm of chromosome 3 in renal cell carcinoma. Nature 1987, 327, 721–724. [Google Scholar] [CrossRef]
- Choyke, P.L.; Glenn, G.M.; Walther, M.M.; Zbar, B.; Weiss, G.H.; Alexander, R.B.; Hayes, W.S.; Long, J.P.; Thakore, K.N.; Linehan, W.M. The natural history of renal lesions in von Hippel-Lindau disease: A serial CT study in 28 patients. AJR Am. J. Roentgenol. 1992, 159, 1229–1234. [Google Scholar] [CrossRef]
- Solomon, D.; Schwartz, A. Renal pathology in von Hippel-Lindau disease. Hum. Pathol. 1988, 19, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Walther, M.M.; Lubensky, I.A.; Venzon, D.; Zbar, B.; Linehan, W.M. Prevalence of microscopic lesions in grossly normal renal parenchyma from patients with von Hippel-Lindau disease, sporadic renal cell carcinoma and no renal disease: Clinical implications. J. Urol. 1995, 154, 2010–2014, discussion 4–5. [Google Scholar] [CrossRef] [PubMed]
- Mubarak, M.; Al-Gharaibeh, N.; Sommaruga, S.; Li, J.; Vortmeyer, A.O. Histological Tracking into the Third Dimension: Evolution of Early Tumorigenesis in VHL Kidney. J. Kidney Cancer VHL 2021, 8, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.V.; Kuzmin, I.; Wei, M.H.; Pack, S.; Geil, L.; Johnson, B.E.; Stanbridge, E.J.; Lerman, M.I. Down-regulation of transmembrane carbonic anhydrases in renal cell carcinoma cell lines by wild-type von Hippel-Lindau transgenes. Proc. Natl. Acad. Sci. USA 1998, 95, 12596–12601. [Google Scholar] [CrossRef] [PubMed]
- Wykoff, C.C.; Beasley, N.J.; Watson, P.H.; Turner, K.J.; Pastorek, J.; Sibtain, A.; Wilson, G.D.; Turley, H.; Talks, K.L.; Maxwell, P.H.; et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 2000, 60, 7075–7083. [Google Scholar]
- Ebert, B.L.; Firth, J.D.; Ratcliffe, P.J. Hypoxia and mitochondrial inhibitors regulate expression of glucose transporter-1 via distinct Cis-acting sequences. J. Biol. Chem. 1995, 270, 29083–29089. [Google Scholar] [CrossRef]
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Aimee, Y.Y.; et al. Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev. 1998, 12, 149–162. [Google Scholar] [CrossRef]
- Dickenmann, M.; Oettl, T.; Mihatsch, M.J. Osmotic nephrosis: Acute kidney injury with accumulation of proximal tubular lysosomes due to administration of exogenous solutes. Am. J. Kidney Dis. 2008, 51, 491–503. [Google Scholar] [CrossRef]
- Moreau, J.F.; Droz, D.; Sabto, J.; Jungers, P.; Kleinknecht, D.; Hinglais, N.; Michel, J.R. Osmotic Nephrosis Induced by Water-Soluble Triiodinated Contrast Media in Man. A Retrospective Study of 47 Cases. Radiology 1975, 115, 329–336. [Google Scholar] [CrossRef]
- Zhou, C.; Vink, R.; Byard, R.W. Hyperosmolarity Induces Armanni-Ebstein-like Renal Tubular Epithelial Swelling and Cytoplasmic Vacuolization. J. Forensic. Sci. 2017, 62, 229–232. [Google Scholar] [CrossRef]
- Benej, M.; Pastorekova, S.; Pastorek, J. Carbonic anhydrase IX: Regulation and role in cancer. Subcell Biochem. 2014, 75, 199–219. [Google Scholar] [PubMed]
- Shively, S.B.; Beltaifa, S.; Gehrs, B.; Duong, H.; Smith, J.; Edwards, N.A.; Lonser, R.R.; Raffeld, M.; Vortmeyer, A.O. Protracted haemangioblastic proliferation and differentiation in von Hippel-Lindau disease. J. Pathol. 2008, 216, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Vortmeyer, A.O.; Tran, M.G.; Zeng, W.; Gläsker, S.; Riley, C.; Tsokos, M.; Ikejiri, B.; Merrill, M.J.; Raffeld, M.; Zhuang, Z.; et al. Evolution of VHL tumourigenesis in nerve root tissue. J. Pathol. 2006, 210, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Frew, I.J.; Moch, H. A clearer view of the molecular complexity of clear cell renal cell carcinoma. Ann. Rev. Pathol. 2015, 10, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.J.; Turajlic, S.; Rowan, A.; Nicol, D.; Farmery, J.H.R.; O’Brien, T.; Martincorena, I.; Tarpey, P.; Angelopoulos, N.; Yates, L.R.; et al. Timing the landmark events in the evolution of clear cell renal cell cancer: TRACERx Renal. Cell 2018, 173, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Meister, M.; Choyke, P.; Anderson, C.; Patel, U. Radiological evaluation, management, and surveillance of renal masses in von Hippel-Lindau disease. Clin. Radiol. 2009, 64, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Spowart, J.E. Automated serial sectioning for 3-D analysis of microstructures. Scr. Mater. 2006, 55, 5–10. [Google Scholar] [CrossRef]
- Kudo, Y.; Shimada, Y.; Matsubayashi, J.; Kitamura, Y.; Makino, Y.; Maehara, S.; Hagiwara, M.; Park, J.; Yamada, T.; Takeuchi, S.; et al. Artificial intelligence analysis of three-dimensional imaging data derives factors associated with postoperative recurrence in patients with radiologically solid-predominant small-sized lung cancers. Eur. J. Cardiothorac. Surg. 2022, 61, 751–760. [Google Scholar] [CrossRef]
- Vortmeyer, A.O.; Weil, R.J.; Zhuang, Z. Proteomic applications for differential diagnosis of histologically identical tumors. Neurology 2003, 61, 1626–1627. [Google Scholar] [CrossRef]
- Al-Gharaibeh, N.S.; Temm, C.J.; Shively, S.B.; Vortmeyer, A.O. Kidney in VHL disease: Early cl ear cell proliferation occurs in the distal tubular system. Oncol. Rep. 2022, 48, 222. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case 385 | Case 393 | Case 389 | Case 386 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Block #7 | Block #24 | Block #25 | Block #13 | Block #17 | Block #1 | Block #57 | Block #12 | Block #25 | Block #26 |
| 270 | 87 | 97 | 183 | 219 | 80 | 54 | 49 | 78 | 77 |
| Type of Lesions | Lesion 1 | Lesion 2 | Lesion 3 | Lesion 4 | Total |
|---|---|---|---|---|---|
| Number of lesions in all slides | 958 | 154 | 53 | 29 | 1194 |
| Percentage of each type from total | 80.2% | 12.9% | 4.4% | 2.5% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Gharaibeh, N.S.; Shively, S.B.; Vortmeyer, A.O. Characterization of Microscopic Multicellular Foci in Grossly Normal Renal Parenchyma of Von Hippel-Lindau Kidney. Medicina 2022, 58, 1725. https://doi.org/10.3390/medicina58121725
Al-Gharaibeh NS, Shively SB, Vortmeyer AO. Characterization of Microscopic Multicellular Foci in Grossly Normal Renal Parenchyma of Von Hippel-Lindau Kidney. Medicina. 2022; 58(12):1725. https://doi.org/10.3390/medicina58121725
Chicago/Turabian StyleAl-Gharaibeh, Nayef S., Sharon B. Shively, and Alexander O. Vortmeyer. 2022. "Characterization of Microscopic Multicellular Foci in Grossly Normal Renal Parenchyma of Von Hippel-Lindau Kidney" Medicina 58, no. 12: 1725. https://doi.org/10.3390/medicina58121725
APA StyleAl-Gharaibeh, N. S., Shively, S. B., & Vortmeyer, A. O. (2022). Characterization of Microscopic Multicellular Foci in Grossly Normal Renal Parenchyma of Von Hippel-Lindau Kidney. Medicina, 58(12), 1725. https://doi.org/10.3390/medicina58121725