Behavioral and Biochemical Effects of Mukia madrespatana Following Single Immobilization Stress on Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Plant Material and Chemicals
2.3. Acute Toxicity Test
2.4. Experimental Protocol
2.4.1. Investigation 1: Effect of Various Doses of M. madrespatana to Assess the Exploratory Activity in the Novel and Familiar Environment to Find Out the Potential Dose
2.4.2. Investigation 2: Effect of Potential/Selected Dose (2000 mg/kg/day) of M. madrespatana on Behavioral Deficits, Oxidative Stress, and Memory Function Induced by Single Immobilization Stress
2.4.3. Immobilization Stress Procedure
2.5. Behavioral Tests
2.5.1. Elevated Plus Maze (EPM) Test
2.5.2. Light-Dark Activity (LDA) Test
2.5.3. Forced Swim Test (FST)
2.5.4. Morris Water Maze (MWM) Test
2.6. Biochemical Analysis
2.6.1. Determination of Malondialdehyde (MDA)
2.6.2. Determination of Superoxide Dismutase (SOD) Activity
2.6.3. Determination of Catalase (CAT) Activity
2.6.4. Determination of Glutathione Peroxidase (GPx) Activity
2.6.5. Determination of Acetylcholinesterase (AChE) Activity
2.6.6. Determination of 5-HT and 5-HIAA
2.6.7. Determination of Acetylcholine (ACh)
2.7. Statistical Analysis
3. Results
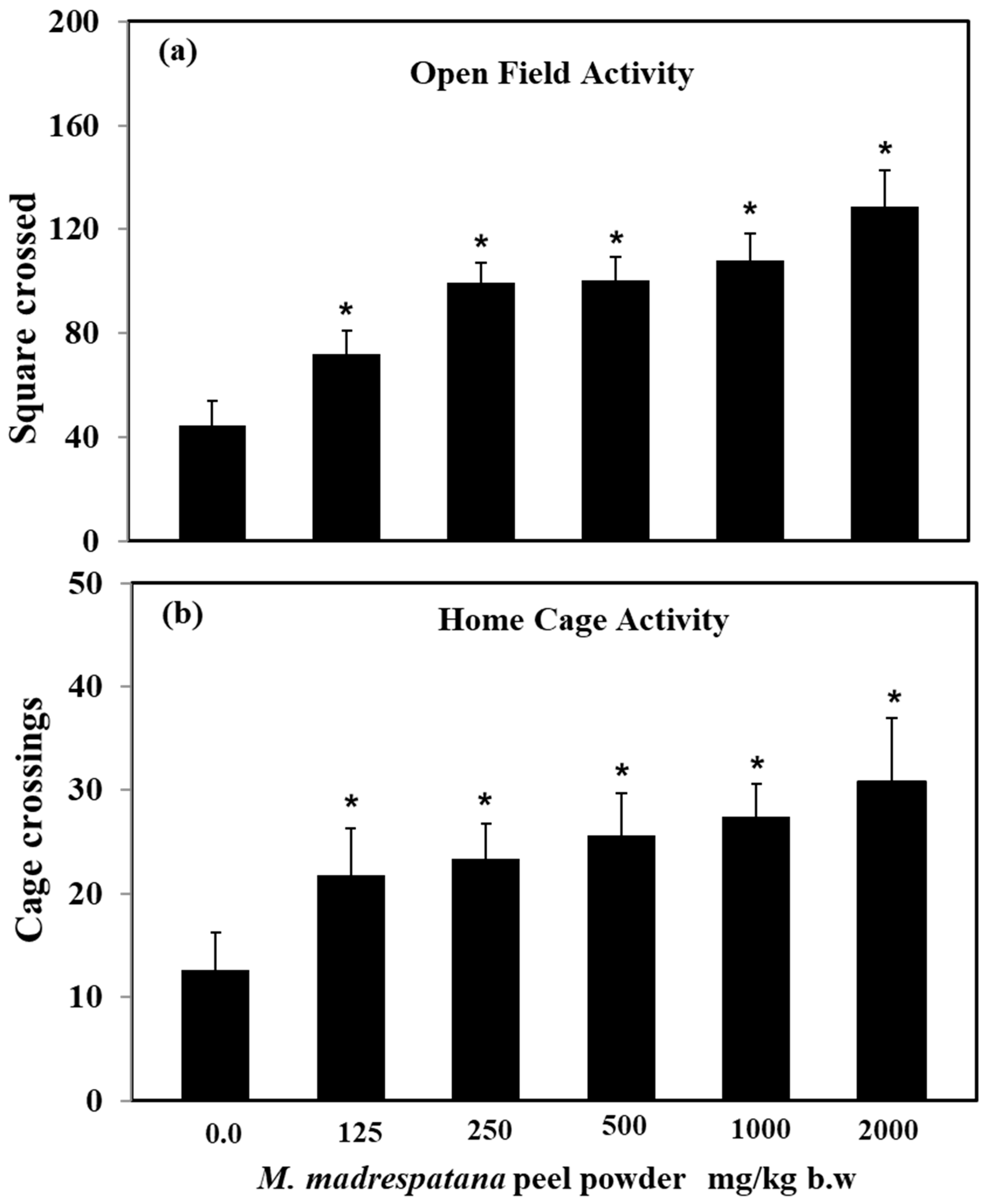
3.1. Various Doses of M. madrespatana Peel
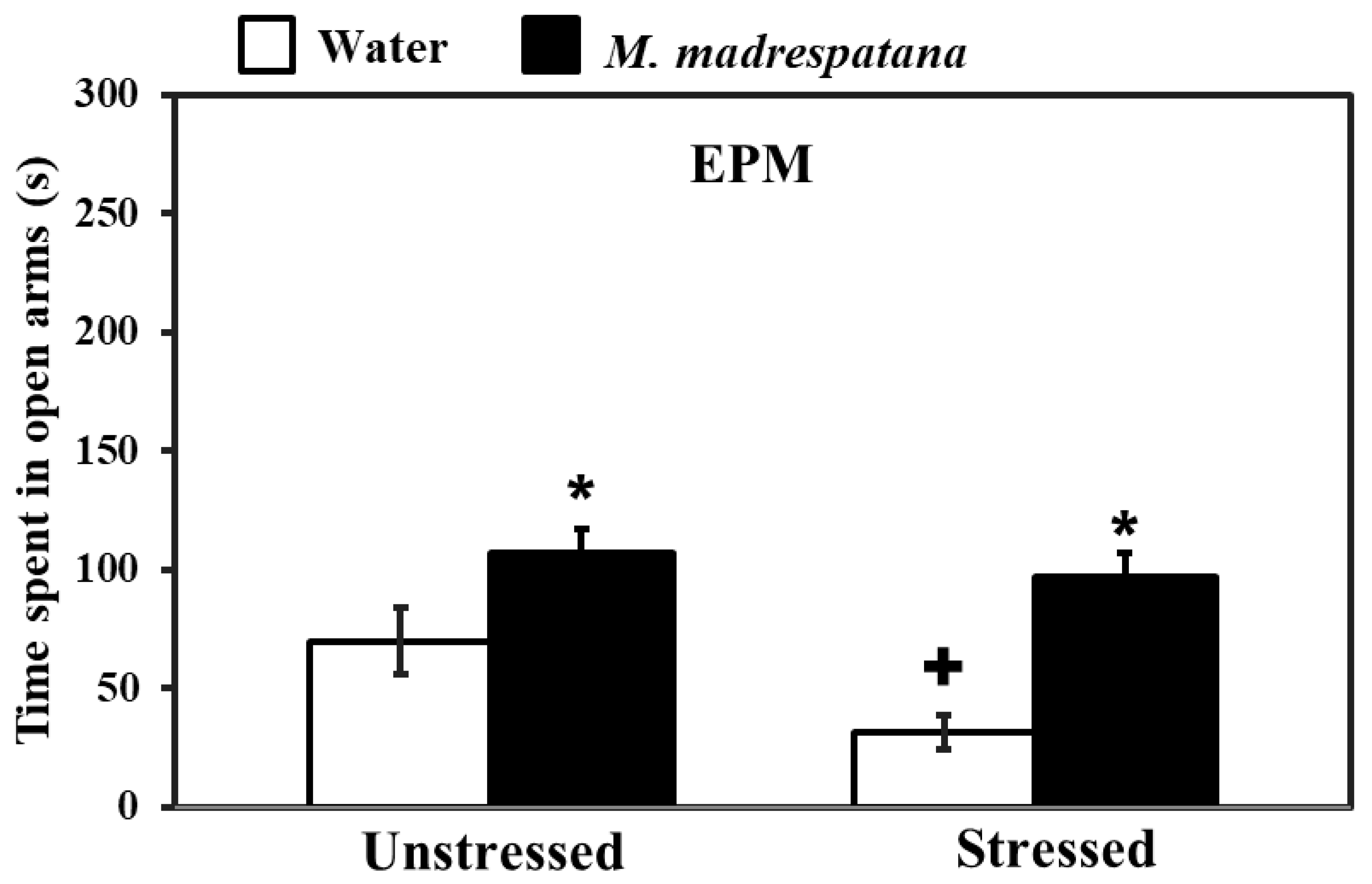
3.2. Elevated Plus-Maze (EPM)
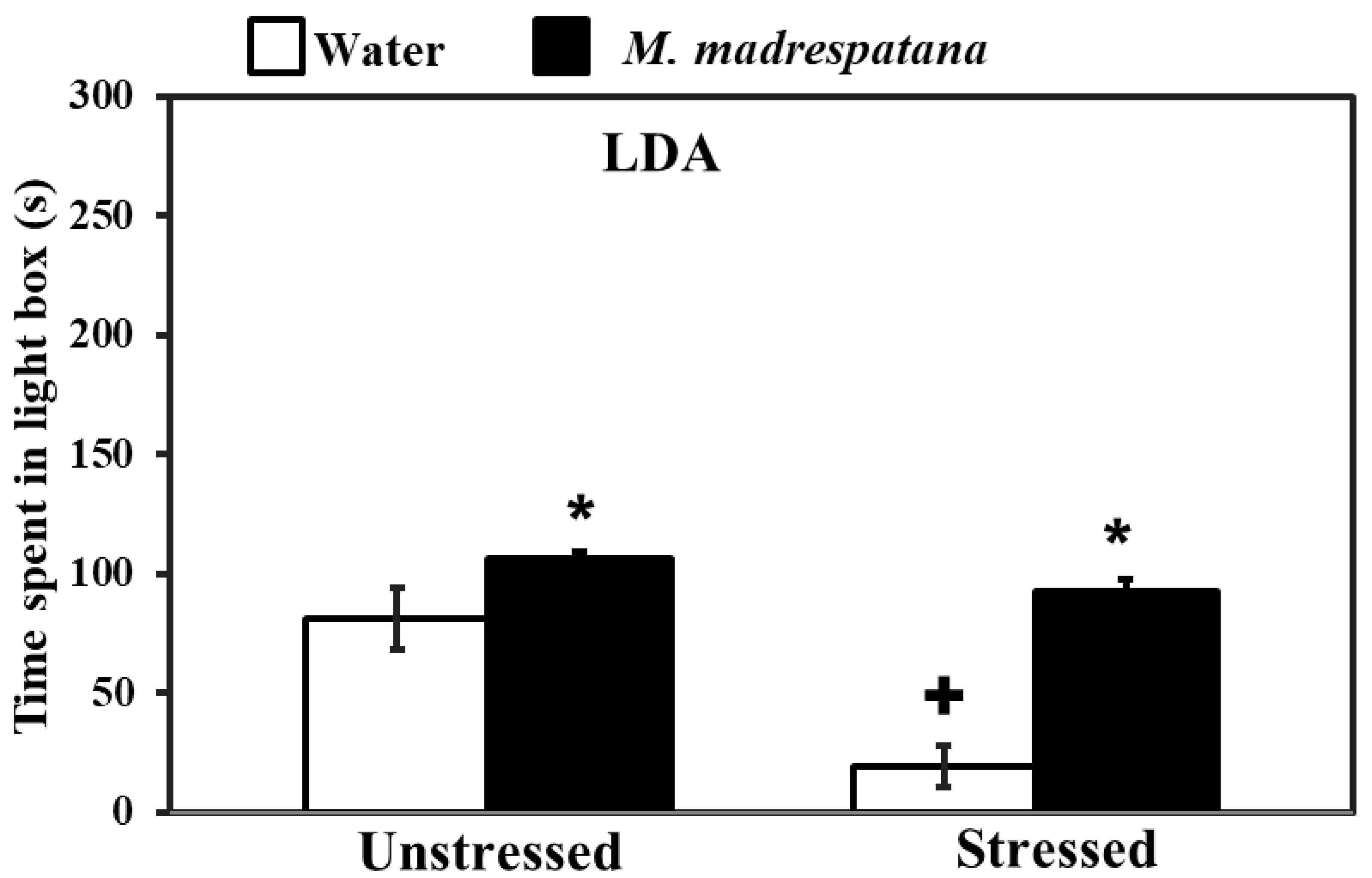
3.3. Light/Dark Box Activity
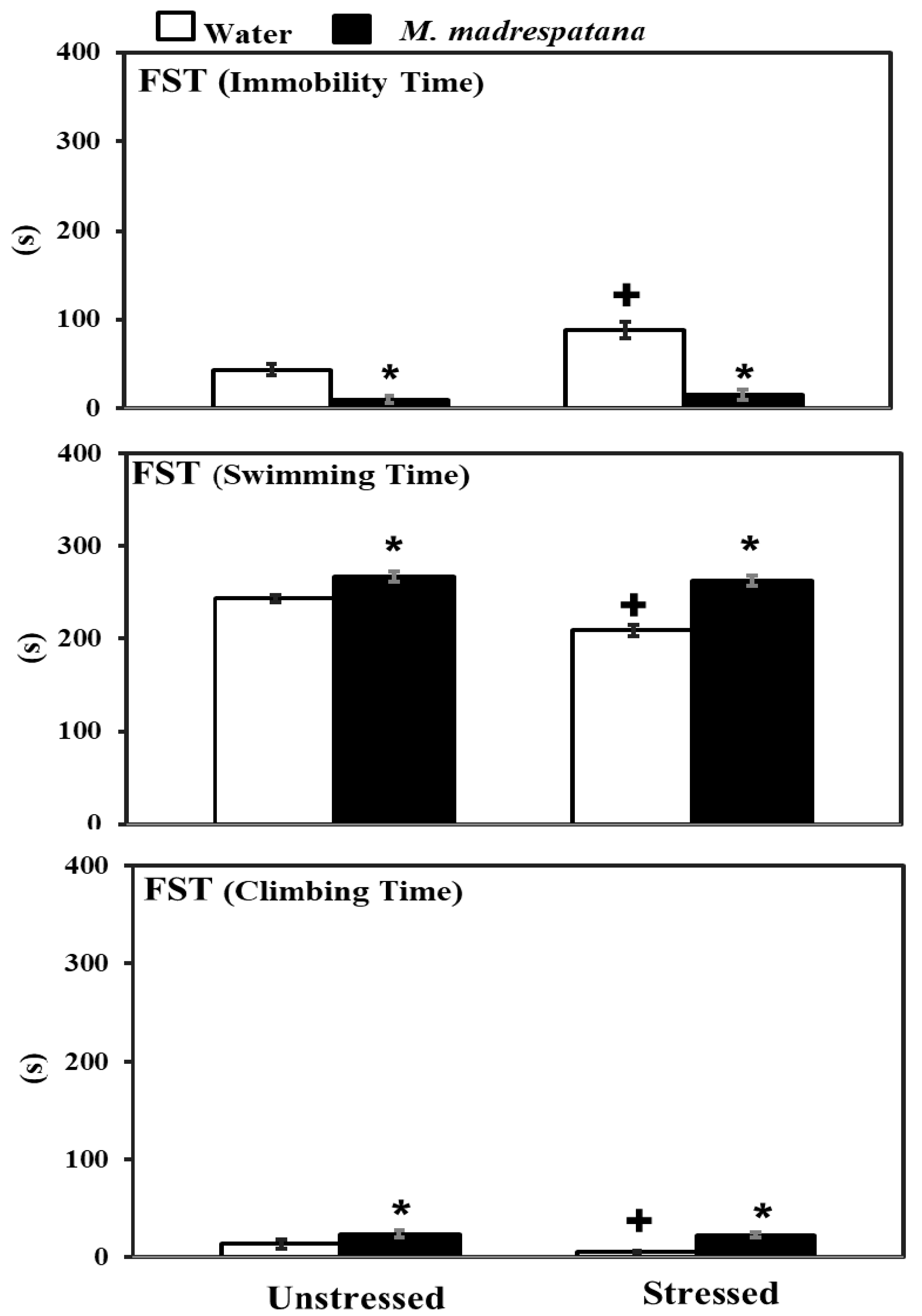
3.4. Forced Swim Test (FST)
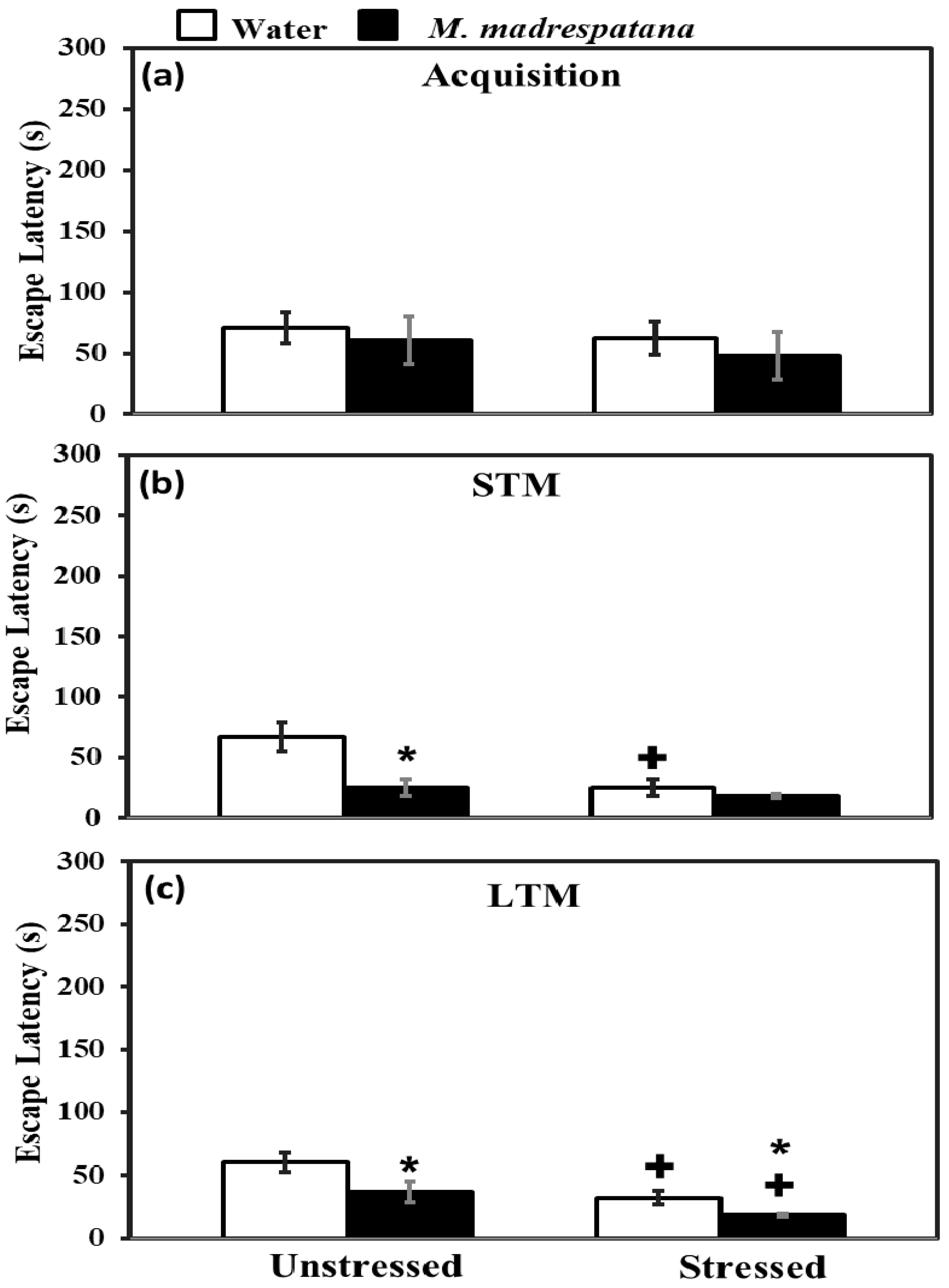
3.5. Morris Water Maze (MWM) Test
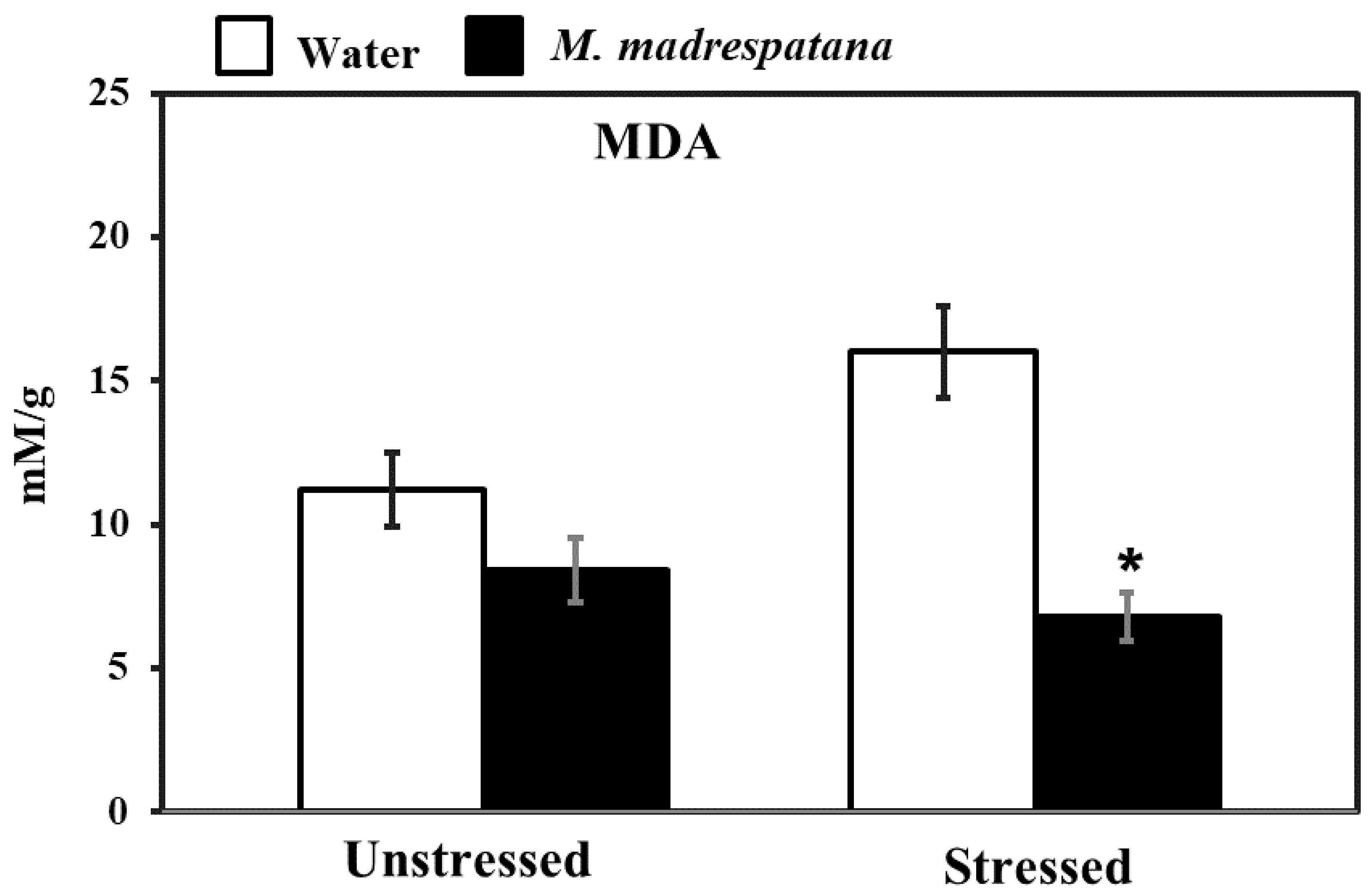
3.6. Assessment of Oxidative Stress
3.7. Determination of Antioxidant Enzymes
3.8. Determination of AChE
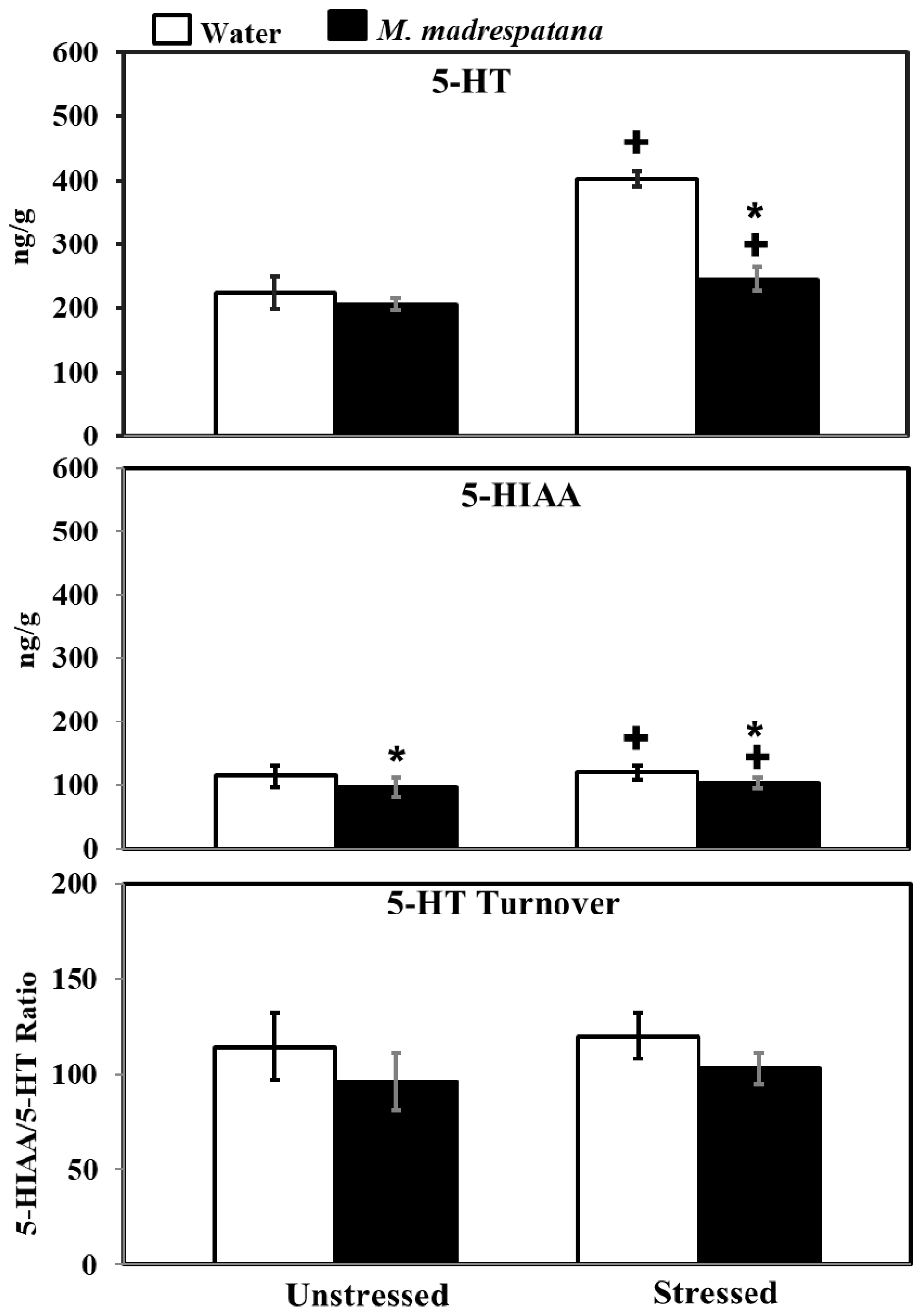
3.9. Determination of 5-Hydroxy Tryptamine (5-HT) and 5-Hydroxy Indole Acetic Acid (5-HIAA) Levels
3.10. Determination of ACh Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chovatiya, R.; Medzhitov, R. Stress, inflammation, and defense of homeostasis. Mol. Cell 2014, 54, 281–288. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Gümüslü, S. Immobilization stress in rat tissues: Alterations in protein oxidation, lipid peroxidation and antioxidant defense system. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 144, 342–347. [Google Scholar] [CrossRef]
- Miyata, S.; Taniguchi, M.; Koyama, Y.; Shimizu, S.; Tanaka, T.; Yasuno, F.; Yamamoto, A.; Iida, H.; Kudo, T.; Katayama, T.; et al. Association between chronic stress-induced structural abnormalities in Ranvier nodes and reduced oligodendrocyte activity in major depression. Sci. Rep. 2016, 6, 23084. [Google Scholar] [CrossRef] [PubMed]
- Vale, W.W.; Spiess, J.; Rivier, C.; Rivier, J. Characterization of a 41-residue ovine hypothalamic peptide that differential environmental modulations on locomotor activity, exploration and spatial behaviour in young and old rats. Physiol. Behav. 1981, 59, 265–271. [Google Scholar]
- Musazzi, L.; Milanese, M.; Farisello, P.; Zappettini, S.D. Acute stress increases depolarization-evoked glutamate release in the rat prefrontal/frontal cortex: The dampening action of antidepressants. PLoS ONE 2010, 5, e8566. [Google Scholar] [CrossRef]
- Dong, K.; Wu, M.; Liu, X.; Huang, Y.; Zhang, D.; Wang, Y.; Yan, L.J.; Shi, D. Glutaredoxins concomitant with optimal ROS activate AMPK through S-glutathionylation to improve glucose metabolism in type 2 diabetes. Free Radic. Biol. Med. 2016, 101, 334–347. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Afshari, R.; Farkhondeh, T.; Karimnezhad, F. Effects of buprenorphine on balance of oxidant/antioxidant system in the different ages of male rat liver. J. Biochem. Mol. Toxicol. 2015, 29, 249–253. [Google Scholar] [CrossRef]
- Ali, S.N.; Ahmad, M.K.; Mahmood, R. Sodium chlorate, A herbicide and major water disinfectant byproduct, generates reactive oxygen species and induces oxidative damage in human erythrocytes. Environ. Sci. Pollut. Res. Int. 2017, 24, 1898–1909. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Samini, F.; Farkhondeh, T. Chrysin treatment improves diabetes and its complications in streptozotocin-induced diabetic rats. Can. J. Physiol. Pharmacol. 2016, 94, 388–393. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.; Yu, M.; Xu, Y.; Ma, R.; Yu, Z. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol. Biochem. 2016, 104, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Borji, A.; Tabasi, S.H. Effects of Cichorium intybus linn on blood glucose, lipid constituents and selected oxidative stress parameters in streptozotocin-induced diabetic rats. Cardiovasc. Hematol. Disord. Drug Targets 2013, 13, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, A.; Rabiei, Z.; Rafieian, M.; Shirzad, H. A review study on medicinal plants affecting amnesia through cholinergic system. J. Herbmed. Plarmacol. 2012, 1, 3–9. [Google Scholar]
- Rahnama, S.; Rabiei, Z.; Alibabaei, Z.; Mokhtari, S.; Rafieian-kopaei, M.; Deris, F. Antiamnesic activity of Citrus aurantium flowers extract against scopolamine-induced memory impairments in rats. Neurol. Sci. 2014, 36, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Haq, M.Z.; Riaz, M.; Saad, B. Anthocyanins and Human Health, Springer Briefs in Food, Health and Nutrition; Springer: New York, NY, USA, 2016; pp. 1–19. [Google Scholar]
- Mamun-or-Rashid, A.N.M.; Hossain, M.S.; Hassan, N.; Dash, B.K.; Sapon, M.A.; Sen, M.K. A review on medicinal plants with antidiabetic activity. Pharmacogn. Phytochem. 2014, 3, 149–159. [Google Scholar]
- Thabrew, M.I.; Gove, C.D.; Hughes, R.D. Protection against galactosamine and tert-butyl hydroperoxide induced hepatocyte damage by Melothria maderaspatana extract. Phytother. Res. 1995, 9, 513–517. [Google Scholar] [CrossRef]
- Petrus, A.J.A. Mukia maderaspatana (L.) M. Roemer-A review of its global distribution, phytochemical profile and antioxidant capacity. Asian J. Chem. 2012, 24, 23612368. [Google Scholar]
- Priya, G.S.; Radhika, R.; Siddhuraju, P. Antioxidant and antimicrobial activity of traditional indian leavesy vegetables: Mukia maderaspatana and Solanum trilobatum. Int. J. Pharm. Pharm. Sci. 2012, 4, 513–521. [Google Scholar]
- Ramakrishnamacharya, C.H.; Krishnaswamy, M.R.; Rao, R.B.; Vishwanathan, S. Anti-inflammatory efficacy of Melothria maderaspatana in active rheumatoid arthritis. Clin. Rheumatol. 1996, 15, 214–215. [Google Scholar] [CrossRef]
- Raja, B.; Kaviarasan, K.; Arjunan, M.M.; Pugalendi, K.V. Effect of Melothria maderaspatana leaf-tea consumption on blood pressure, lipid profile, anthropometry, fibrinogen, bilirubin and albumin levels in patients with hypertension. J. Altern. Complement. Med. 2007, 13, 349–354. [Google Scholar] [CrossRef]
- Srilatha, B.R.; Ananda, S. Antidiabetic effects of Mukia maderaspatana and its phenolics: An in-vitro study on gluconeogenesis and glucose uptake in rat tissues. Pharm. Biol. 2014, 52, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Bocarsly, M.E.; Barson, J.R.; Hauca, J.M.; Hoebel, B.G.; Leibowitz, S.F.; Avena, N.M. Effects of perinatal exposure to palatable diets on body weight and sensitivity to drugs of abuse in rats. Physiol. Behav. 2012, 107, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Hemalatha, S.; Wahi, A.K.; Singh, P.N.; Chansouria, J.P.N. Evaluation of anti-hyperglycemic and free radical scavenging activity of Melothria maderaspatana Linn. in streptozotocin-induced diabetic rats. Afr. J. Pharm. Pharmacol. 2010, 4, 817–822. [Google Scholar]
- Pandey, D.; Pandey, S.; Hemalatha, S. Hypolipidemic activity of aqueous extract of Melothria maderaspatana. Pharmacol. Online 2010, 3, 76–83. [Google Scholar]
- Vadivelan, R.; Dhanabal, S.P.; Mohan, P.; Shanish, A.; Elango, K.; Suresh, B. Antidiabetic activity of Mukia maderaspatana (L.) Roem in alloxan induced diabetic rats. Res. J. Pharmacol. Pharmacodyn. 2010, 2, 78–80. [Google Scholar]
- Saher, S.; Ali, S.A.; Sakle, N.S.; Khan, S.W. Allium cepa attenuates ischemia reperfusion induced brain injury. Int. J. Pharmaceutical. Res. Allied. Sci. 2015, 4, 59–68. [Google Scholar]
- Samad, N.; Tariq, S.; Mahmood, A.; Imran, I.; Alqahtani, F. Reversal of repeated noise stress-induced behavioral, cognitive, neuroendocrine response and oxidative parameters by Abelmoschus esculentus (Okra) root powder in rats. Pak. J. Pharm. Sci. 2020, 33, 1221–1232. [Google Scholar]
- Tabassum, S.; Ahmad, S.; Madiha, S.; Khaliqm, S.; Shahzadm, S.; Batool, Z.; Haider, S. Impact of oral supplementation of Glutamate and GABA on memory performance and neurochemical profile in hippocampus of rats. Pak. J. Pharm. Sci. 2017, 30, 1013–1021. [Google Scholar]
- Samad, N.; Parveen, T.; Haider, S.; Haleem, D.J. Attenuation of restraint induced behavioral deficits by buspirone and propranolol in rats. J. Coll. Physicians. Surg. Pak. 2005, 15, 795–798. [Google Scholar]
- Samad, N.; Saleem, A. Administration of Allium cepa L. bulb attenuates stress-produced anxiety and depression and improves memory in male mice. Metab. Brain Dis. 2018, 33, 271–281. [Google Scholar] [CrossRef]
- Porsolt, R.; Enna, S.J.; Malick, J.B.; Richelson, E. Behavioral despair. In Antidepressants: Neurochemical, Behavioral and Clinical Perspectives; Raven Press New York: New York, NY, USA, 1981; pp. 121–139. [Google Scholar]
- Haider, S.; Naqvi, F.; Batool, Z.; Tabassum, S.; Perveen, T.; Saleem, S.; Haleem, D.J. Decreased Hippocampal 5-HT and DA Levels Following Sub-Chronic Exposure to Noise Stress: Impairment in both Spatial and Recognition Memory in Male Rats. Sci. Pharm. 2012, 80, 1001–1011. [Google Scholar] [CrossRef]
- Chow, C.K.; Tappel, A.L. An enzymatic protective mechanism against lipid peroxidation damage to lungs of ozone-exposed rats. Lipids 1972, 7, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Naskar, S.; Islam, A.; Mazumder, U.K.; Saha, P.; Haldar, P.K.; Gupta, M. In Vitro and In Vivo Antioxidant Potential of Hydromethanolic Extract of Phoenix dactylifera Fruits. J. Sci. Res. 2010, 2, 144–157. [Google Scholar] [CrossRef]
- Pari, L.; Latha, M. Protective role of Scorparia dulcis plant extract on brain antioxidant status and lipid peroxidation in STZ diabetic male wistar rats. BMC 2004, 6, 16. [Google Scholar]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 114–120. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Liaquat, L.; Sadir, S.; Batool, Z.; Tabassum, S.; Shahzad, S.; Afzal, A.; Haider, S. Acute aluminum chloride toxicity revisited: Study on DNA damage and histopathological, biochemical and neurochemical alterations in rat brain. Life Sci. 2019, 217, 202–211. [Google Scholar] [CrossRef]
- Samad, N.; Saleem, A.; Yasmin, F.; Shehzad, M.A. Quercetin protects against stress-induced anxiety- and depression-like behavior and improves memory in male mice. Physiol. Res. 2018, 67, 795–808. [Google Scholar] [CrossRef]
- Daglia, M.; Di Lorenzo, A.; Nabavi, S.F.; Sureda, A.; Khanjani, S.; Moghaddam, A.H.; Braidy, N.; Nabavi, S.M. Improvement of Antioxidant Defences and Mood Status by Oral GABA Tea Administration in a Mouse Model of Post-Stroke Depression. Nutrients 2017, 9, 446. [Google Scholar] [CrossRef]
- Samad, N.; Jabeen, S.; Imran, I.; Zulfiqar, I.; Bilal, K. Protective effect of gallic acid against arsenic-induced anxiety-/depression- like behaviors and memory impairment in male rats. Metab. Brain Dis. 2019, 34, 1091–1102. [Google Scholar] [CrossRef]
- Cho, S.H.; Heo, S.J.; Yang, H.W.; Ko, E.Y.; Jung, M.S.; Cha, S.H.; Ahn, G.; Jeon, Y.J.; Kim, K.N. Protective Effect of 3-Bromo-4,5-Dihydroxybenzaldehyde from Polysiphonia morrowii Harvey against Hydrogen Peroxide-Induced Oxidative Stress In Vitro and In Vivo. J. Microbiol. Biotechnol. 2019, 29, 1193–1203. [Google Scholar] [CrossRef]
- Miłkowska, P.; Popko, K.; Demkow, U.; Wolańczyk, T. Pro-inflammatory Cytokines in Psychiatric Disorders in Children and Adolescents: A Review. Adv. Exp. Med. Biol. 2017, 1021, 73–80. [Google Scholar]
- Gadek-Michalska, A.; Tadeusz, J.; Rachwalska, P.; Bugajski, J. Cytokines, prostaglandins and nitric oxide in the regulation of stress-response systems. Pharmacol. Rep. 2013, 65, 1655–1662. [Google Scholar] [CrossRef]
- Shehu, A.; Magaji, M.G.; Yau, J.; Ahmed, A. Methanol stem bark extract of Adansonia digitata ameliorates chronic unpredictable mild stress-induced depression-like behavior: Involvement of the HPA axis, BDNF, and stress biomarkers pathways. J. Basic Clin. Physiol. Pharmacol. 2019, 30, 3. [Google Scholar] [CrossRef] [PubMed]
- Samad, N.; Perveen, T.; Haider, S.; Haleem, M.; Haleem, D.J. Inhibition of restraint-induced neuroendocrine and serotonergic responses by buspirone in rats. Pharmacol. Rep. 2006, 58, 636–642. [Google Scholar] [PubMed]
- Perveen, T.; Emad, S.; Haider, S.; Sadaf, S.; Qadeer, S.; Batool, Z.; Sarfaraz, Y.; Sheikh, S. Role of Cyclooxygenase Inhibitors in Diminution of Dissimilar Stress-induced Depressive Behavior and Memory Impairment in Rats. Neuroscience 2018, 370, 121–129. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef]
- Samad, N.; Muneer, A.; Ullah, N.; Zaman, A.; Ayaz, M.M.; Ahmad, I. Banana fruit pulp and peel involved in antianxiety and antidepressant effects while invigorate memory performance in male mice: Possible role of potential antioxidants. Pak. J. Pharm. Sci. 2017, 30, 989–995. [Google Scholar]
- Haleem, D.J.; Samad, N.; Perveen, T.; Haider, S.; Haleem, M.A. Role of serotonin-1A receptors in restraint-induced behavioral deficits and adaptation to repeated restraint stress in rats. Int. J. Neurosci. 2007, 117, 243–257. [Google Scholar] [CrossRef]
- Saravanan, S.; Manokaran, S.; Chand, S. Anxiolytic Activity of Hydro-alcoholic Extract of Mukia maderaspatana Linn. Leaves on Experimental Animals. Jordan J. Pharm. Sci. 2012, 5, 43–50. [Google Scholar]
- Petrus, A.J.A. Mukia maderaspatana (Linn.) M. Roemer: A potentially antidiabetic and vasoprotective functional leafy-vegetable. Phcog. J. 2012, 4, 1–12. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Neeta, D.; Sarkar, M.K. Inhibition of carrageenin-induced pedal oedema in rats by immobilization stress. Res. Exp. Med. 1987, 187, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Haleem, D.J. Behavioral deficits and exaggerated feedback control over raphe-hippocampal serotonin neurotransmission in restrained rats. Pharmacol. Rep. 2011, 63, 888–897. [Google Scholar] [CrossRef]
- Samad, N.; Batool, F.; Haleem, D.J. Neurochemical and behavioral effects of 8-OH-DPAT following exposure to restraint stress in rats. Pharmacol. Rep. 2007, 59, 173–180. [Google Scholar]
- Kraus, C.; Castrén, E.; Kasper, S.; Lanzenberger, R. Serotonin and neuroplasticity-Links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef]
- Pehrson, A.L.; Leiser, S.C.; Gulinello, M.; Dale, E.; Li, Y.; Waller, J.A.; Sanchez, C. Treatment of cognitive dysfunction in major depressive disorder—a review of the preclinical evidence for efficacy of selective serotonin reuptake inhibitors, serotonin norepinephrine reuptake inhibitors and the multimodal-acting antidepressant vortioxetine. Eur. J. Pharmacol. 2015, 753, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Samad, N.; Imran, I.; Zulfiqar, I.; Bilal, K. Ameliorative effect of lithium chloride against d-galactose induced behavioral and memory impairment, oxidative stress and alteration in serotonin function in rats. Pharmacol. Rep. 2019, 71, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Parameswari, K.; Ananthi, T. Physico-Chemical and Phytochemical Analysis of Mukia maderaspatana L. Res. J. Sci. Technol. 2013, 5, 272–274. [Google Scholar]
- Lian, W.; Fang, J.; Xu, L.; Zhou, W.; Kang, D.; Xiong, W.; Jia, H.; Liu, A.L.; Du, G.H. DL0410 Ameliorates memory and cognitive impairments induced by scopolamine via increasing cholinergic neurotransmission in mice. Molecules 2017, 22, 410. [Google Scholar] [CrossRef]
- Imperato, A.; Puglisi-Allegra, S.; Casolini, P.; Angelucci, L. Changes in brain dopamine and acetylcholine release during and following stress are independent of the pituitary-adrenocortical axis. Brain Res. 1991, 538, 111–117. [Google Scholar] [CrossRef]
- Kaufer, D.; Friedman, A.; Seidman, S.; Soreq, H. Acute stress facilitates long-lasting changes in cholinergic gene expression. Nature 1998, 393, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Naqvi, F.; Batool, Z.; Tabassum, S.; Sadir, S.; Liaquat, L.; Naqvi, F.; Zuberi, N.A.; Shakeel, H.; Perveen, T. Pretreatment with curcumin attenuates anxiety while strengthens memory performance after one short stress experience in male rats. Brain Res. Bull. 2015, 115, 1–8. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samad, N.; Ali, A.; Yasmin, F.; Ullah, R.; Bari, A. Behavioral and Biochemical Effects of Mukia madrespatana Following Single Immobilization Stress on Rats. Medicina 2020, 56, 350. https://doi.org/10.3390/medicina56070350
Samad N, Ali A, Yasmin F, Ullah R, Bari A. Behavioral and Biochemical Effects of Mukia madrespatana Following Single Immobilization Stress on Rats. Medicina. 2020; 56(7):350. https://doi.org/10.3390/medicina56070350
Chicago/Turabian StyleSamad, Noreen, Amna Ali, Farzana Yasmin, Riaz Ullah, and Ahmed Bari. 2020. "Behavioral and Biochemical Effects of Mukia madrespatana Following Single Immobilization Stress on Rats" Medicina 56, no. 7: 350. https://doi.org/10.3390/medicina56070350
APA StyleSamad, N., Ali, A., Yasmin, F., Ullah, R., & Bari, A. (2020). Behavioral and Biochemical Effects of Mukia madrespatana Following Single Immobilization Stress on Rats. Medicina, 56(7), 350. https://doi.org/10.3390/medicina56070350