Deduction of Novel Genes Potentially Involved in the Effects of Very Low Dose Atropine (0.003%) Treatment on Corneal Epithelial Cells Using Next-Generation Sequencing and Bioinformatics Approaches


Abstract
:
1. Introduction
2. Materials and Methods
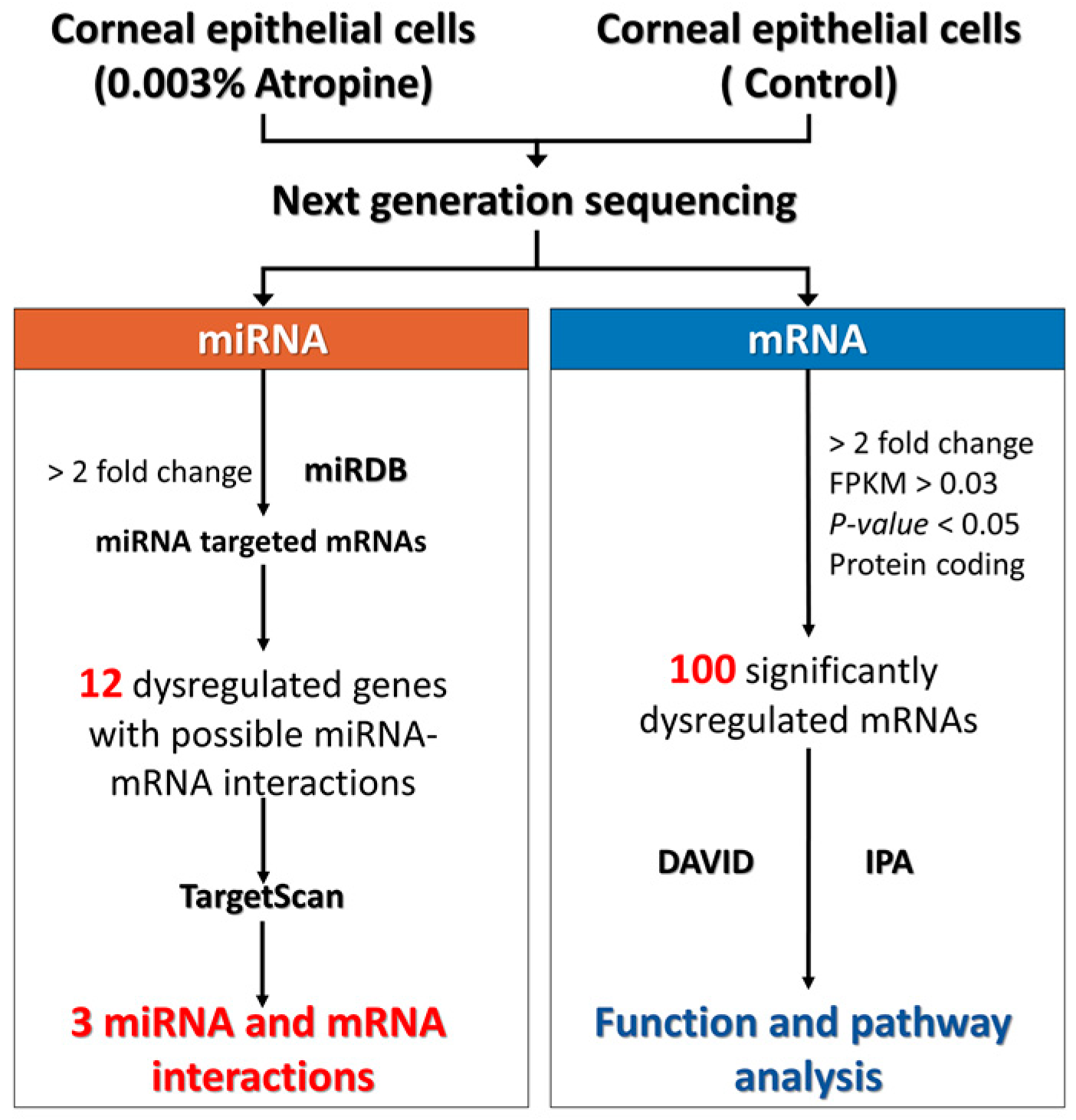
2.1. Study Design
2.2. NGS for miRNA and mRNA Expression Profiles
2.3. Analyses Using miRNA Target Prediction Databases
2.4. Analysis Using DAVID Database
2.5. Analysis Using IPA
3. Results
3.1. Differential Gene Expressions in Corneal Epithelial Cells Treated with Very Low Dose Atropine
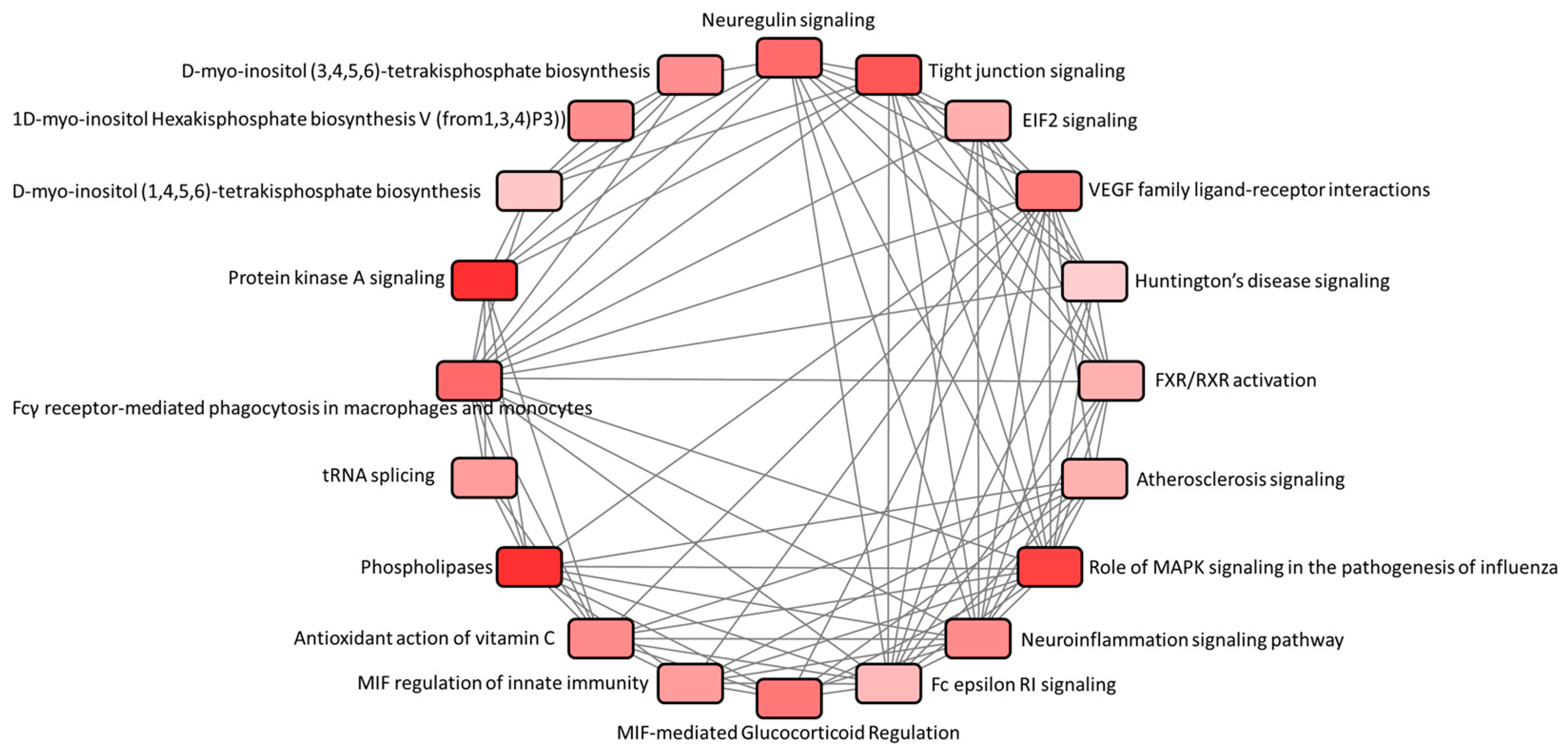
3.2. Function and Pathway Analysis of Dysregulated Genes in Atropine-Treated Corneal Epithelial Cells
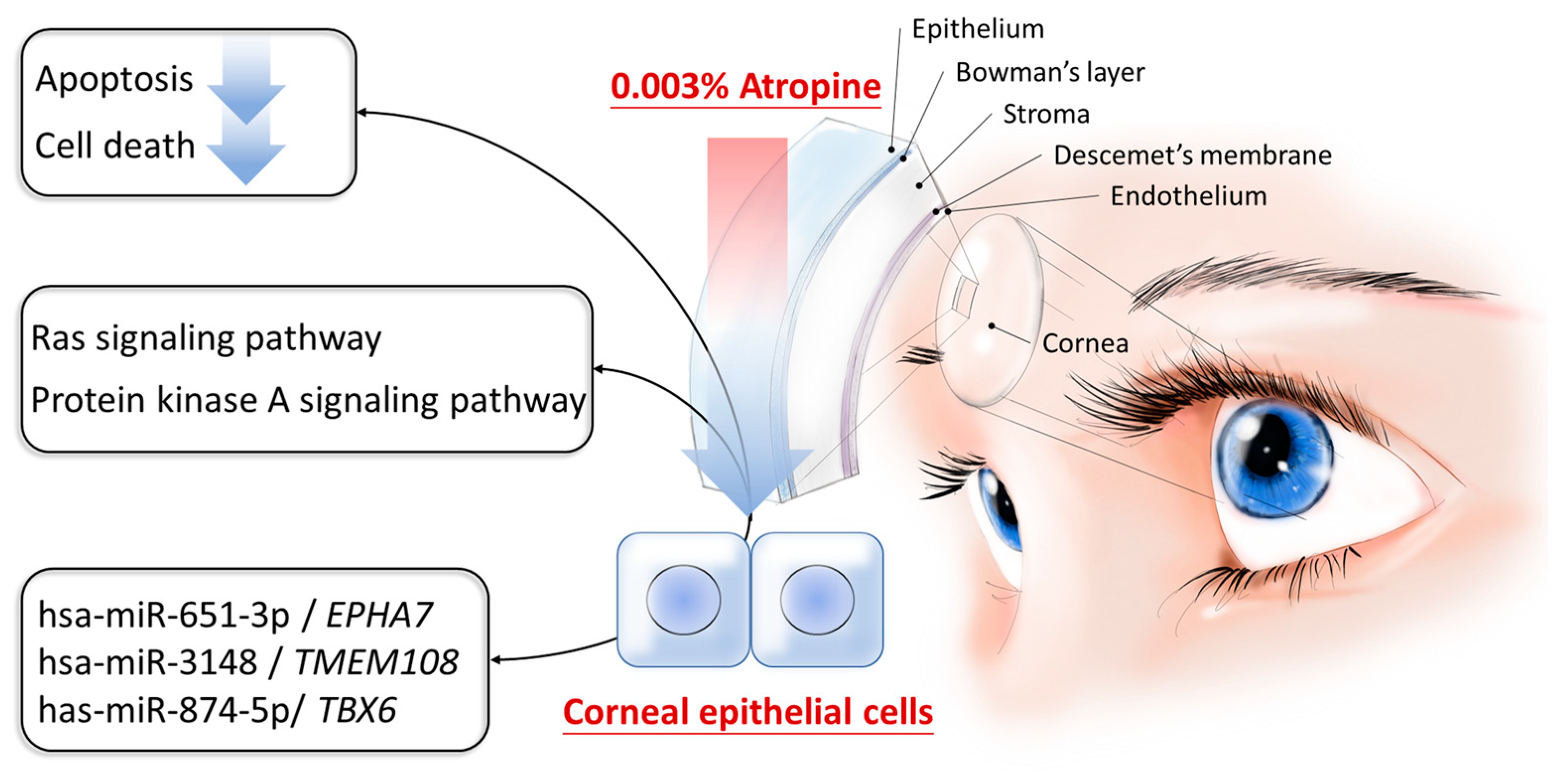
3.3. Potential Dysregulated miRNA-mRNA Interactions in Atropine-Treated Corneal Epithelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gong, Q.; Janowski, M.; Luo, M.; Wei, H.; Chen, B.; Yang, G.; Liu, L. Efficacy and Adverse Effects of Atropine in Childhood Myopia: A Meta-analysis. JAMA Ophthalmol. 2017, 135, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.L.; Wen, Q.; Fan, T.J. Cytotoxicity of atropine to human corneal epithelial cells by inducing cell cycle arrest and mitochondrion-dependent apoptosis. Exp. Toxicol. Pathol. 2015, 67, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, A.; Nishina, S.; Miki, A.; Utsumi, T.; Sugasawa, J.; Hayashi, T.; Sato, M.; Kimura, A.; Fujikado, T. Incidence of side effects of topical atropine sulfate and cyclopentolate hydrochloride for cycloplegia in Japanese children: A multicenter study. Jpn. J. Ophthalmol. 2018, 62, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.C.; Chuang, M.N.; Choi, J.; Chen, H.; Wu, G.; Ohno-Matsui, K.; Jonas, J.B.; Cheung, C.M.G. Update in myopia and treatment strategy of atropine use in myopia control. Eye 2019, 33, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.H.; Hu, W.; Kashgari, G.; Lin, Z.; Nguyen, T.; Doan, M.; Andersen, B. Characterization of enhancers and the role of the transcription factor KLF7 in regulating corneal epithelial differentiation. J. Biol. Chem. 2017, 292, 18937–18950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Galipon, J.; Ishii, R.; Suzuki, Y.; Tomita, M.; Ui-Tei, K. Differential Binding of Three Major Human ADAR Isoforms to Coding and Long Non-Coding Transcripts. Genes 2017, 8, 68. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef]
- Wang, X. Improving microRNA target prediction by modeling with unambiguously identified microRNA-target pairs from CLIP-ligation studies. Bioinformatics 2016, 32, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Bonchev, D. A survey of current software for network analysis in molecular biology. Hum. Genom. 2010, 4, 353–360. [Google Scholar] [CrossRef]
- Wen, Q.; Fan, T.J.; Tian, C.L. Cytotoxicity of atropine to human corneal endothelial cells by inducing mitochondrion-dependent apoptosis. Exp. Biol. Med. 2016, 241, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.F.; Yam, J.C. Low-Concentration Atropine Eye Drops for Myopia Progression. Asia Pac. J. Ophthalmol. 2019. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, K.; Liu, R.B.; Pan, J.H.; Zhang, L.L.; Xu, Z.P.; Lu, X.J. Atropine 0.01% eye drops slow myopia progression: A systematic review and Meta-analysis. Int. J. Ophthalmol. 2019, 12, 1337–1343. [Google Scholar] [CrossRef]
- Zhao, F.; Ma, J.X. Will the long-term use of atropine eye drops in children increase the risk of dry eye? Med. Hypotheses 2019, 132, 109331. [Google Scholar] [CrossRef]
- Figueroa, J.D.; Benton, R.L.; Velazquez, I.; Torrado, A.I.; Ortiz, C.M.; Hernandez, C.M.; Diaz, J.J.; Magnuson, D.S.; Whittemore, S.R.; Miranda, J.D. Inhibition of EphA7 up-regulation after spinal cord injury reduces apoptosis and promotes locomotor recovery. J. Neurosci. Res. 2006, 84, 1438–1451. [Google Scholar] [CrossRef]
- Xiang, C.; Lv, Y.; Wei, Y.; Wei, J.; Miao, S.; Mao, X.; Gu, X.; Song, K.; Jia, S. Effect of EphA7 Silencing on Proliferation, Invasion and Apoptosis in Human Laryngeal Cancer Cell Lines Hep-2 and AMC-HN-8. Cell. Physiol. Biochem. 2015, 36, 435–445. [Google Scholar] [CrossRef]
- Yu, Z.; Lin, D.; Zhong, Y.; Luo, B.; Liu, S.; Fei, E.; Lai, X.; Zou, S.; Wang, S. Transmembrane protein 108 involves in adult neurogenesis in the hippocampal dentate gyrus. Cell Biosci. 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Jiao, H.F.; Sun, X.D.; Bates, R.; Xiong, L.; Zhang, L.; Liu, F.; Li, L.; Zhang, H.S.; Wang, S.Q.; Xiong, M.T.; et al. Transmembrane protein 108 is required for glutamatergic transmission in dentate gyrus. Proc. Natl. Acad. Sci. USA 2017, 114, 1177–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruzat, A.; Witkin, D.; Baniasadi, N.; Zheng, L.; Ciolino, J.B.; Jurkunas, U.V.; Chodosh, J.; Pavan-Langston, D.; Dana, R.; Hamrah, P. Inflammation and the nervous system: The connection in the cornea in patients with infectious keratitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5136–5143. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.L.; Cooper-Morgan, A.; Harrelson, Z.; Papaioannou, V.E. Critical role for Tbx6 in mesoderm specification in the mouse embryo. Mech. Dev. 2003, 120, 837–847. [Google Scholar] [CrossRef]
- Haginiwa, S.; Sadahiro, T.; Kojima, H.; Isomi, M.; Tamura, F.; Kurotsu, S.; Tani, H.; Muraoka, N.; Miyake, N.; Miyake, K.; et al. Tbx6 induces cardiomyocyte proliferation in postnatal and adult mouse hearts. Biochem. Biophys. Res. Commun. 2019, 513, 1041–1047. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated Genes |
| RPL34, ACACA, CUTA, AP2A2, ITPK1, IFI27, ERMARD, ZNF578, ZNF487, TBCE, HHAT, CBS, PCOLCE2, CTIF, UBR7, NEK10, KCNJ5, FAM156A, NAPEPLD, IRF7, PTEN, C16orf45, SMN2, CFB, PYGM, PLA2G3, PRSS27, RPS18, GGT5, GBA, PYCR3, CALML6, KIF6, NRG4, ARHGAP15, ART3, JAML, TIE1, ARHGAP11B, LGR5 |
| Downregulated Genes |
| SQSTM1, SYNRG, PTPRK, COG5, SLC25A26, PSMC4, CEP170, TC2N, FOPNL, NSF, AKT3, SLX1B, FAM120B, STAT4, PIF1, IQANK1, PON1, FRG1, LEKR1, AC145212.1, STK33, CRIP1, PLA2G4C, LIMS4, CDC20, HORMAD1, GGN, FOXA1, CIT, APBB1, BUB1B-PAK6, TBX3, SLAIN1, UGT2B10, IFI27L1, CCHCR1, FAM111B, DHRS11, SMN1, POLQ, RPL7A, ZNF100, PQLC2L, RASA3, OXER1, FOLH1, PLEKHG4, BEGAIN, RNF150, NCALD, OCLN, MCM10, CHST6, DUSP8, COL14A1, PDE9A, ZKSCAN7, HSPA6, FSTL4, PRLR |
| Canonical Pathways | −log10(p-Value) | Ratio | z-Score | Molecules |
|---|---|---|---|---|
| Protein Kinase A Signaling | 2.69 | 0.0182 | 0.378 | NAPEPLD, HHAT, PYGM, DUSP8, PTPRK, PDE9A, PTEN |
| Neuroinflammation Signaling Pathway | 1.87 | 0.0165 | 0.447 | IRF7, KCNJ5, PLA2G4C, AKT3, PLA2G3 |
| Categories | Functions | Diseases or Functions Annotation | p-Value | Activation z-Score | Molecules |
|---|---|---|---|---|---|
| Cell death and survival | Cell death | Cell death of colorectal cancer cell lines | 0.0116 | −0.468 | SQSTM1, PTEN, AP2A2, RPL34, CDC20, LGR5 |
| Cell death and survival | Apoptosis | Apoptosis | 0.0252 | −0.103 | SQSTM1, IFI27L1, PTEN, AKT3, MCM10, OCLN, IFI27, CRIP1, CDC20, CCHCR1, ITPK1, FOXA1, AP2A2, CIT, TBX3, APBB1, ACACA, SMN1/SMN2, LGR5 |
| Pathway | p-Value | Genes |
|---|---|---|
| Glucagon signaling pathway | 0.018 | PYGM, ACACA, CALML6, AKT3 |
| Estrogen signaling pathway | 0.018 | KCNJ5, HSPA6, CALML6, AKT3 |
| Ras signaling pathway | 0.038 | CALML6, PLA2G4C, PLA2G3, RASA3, AKT3 |
| Insulin signaling pathway | 0.043 | PYGM, ACACA, CALML6, AKT3 |
| Arachidonic acid metabolism | 0.046 | GGT5, PLA2G4C, PLA2G3 |
| Hepatitis B | 0.048 | STAT4, IRF7, PTEN, AKT3 |
| Predicted Consequential Pairing of Target Region (Top) and miRNA (Bottom) | Context++ Score | Context++ Score Percentile | Weighted Context++ Score | miRDB | |
|---|---|---|---|---|---|
| Position 1034–1040 of EPHA7 3′ UTR miR-651-3p | 5′ … UGCAGACAUCCUCCAUUUCCUUU… | | | | | | | 3′ GAAAAUCCUAUGUGAAAGGAAA | −0.02 | 37 | 0.00 | 97 |
| Position 3122–3128 of EPHA7 3′ UTR miR-651-3p | 5′ …AUAGUAUUCUUCCUGUUUCCUUU… | | | | | | | 3′ GAAAAUCCUAUGUGAAAGGAAA | −0.02 | 37 | 0.00 | 97 |
| Position 1932-1938 of TMEM108 3′ UTR miR-3148 | 5′ …CACUUUAGGCCACAG - - UUUUUCCU… | | | | | | | | | | 3′ UUCGUGUGUGGUCAAAAAAGGU | −0.02 | 38 | −0.02 | 99 |
| Position 204-210 of TBX6 3′ UTR miR-874-5p | 5′ …UGCUGGGAGUCCAGCUGGGGCCG… | | | | | | | 3′ AGAAUGGGACCACGCACCCCGGC | −0.10 | 57 | −0.10 | 94 |
| Position 269-276 of TBX6 3′ UTR miR-874-5p | 5′ …CUCUGCCUGGCCAAAUGGGGCCA… | | | | | | | | | | | | 3′ AGAAUGGGACCACGC - ACCCCGGC | −0.47 | 98 | −0.47 | 94 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-A.; Hsiao, Y.-T.; Lin, H.-C.; Jian, S.-F.; Chen, Y.-J.; Kuo, P.-L. Deduction of Novel Genes Potentially Involved in the Effects of Very Low Dose Atropine (0.003%) Treatment on Corneal Epithelial Cells Using Next-Generation Sequencing and Bioinformatics Approaches. Medicina 2019, 55, 589. https://doi.org/10.3390/medicina55090589
Chang W-A, Hsiao Y-T, Lin H-C, Jian S-F, Chen Y-J, Kuo P-L. Deduction of Novel Genes Potentially Involved in the Effects of Very Low Dose Atropine (0.003%) Treatment on Corneal Epithelial Cells Using Next-Generation Sequencing and Bioinformatics Approaches. Medicina. 2019; 55(9):589. https://doi.org/10.3390/medicina55090589
Chicago/Turabian StyleChang, Wei-An, Yu-Ting Hsiao, Hsien-Chung Lin, Shu-Fang Jian, Yi-Jen Chen, and Po-Lin Kuo. 2019. "Deduction of Novel Genes Potentially Involved in the Effects of Very Low Dose Atropine (0.003%) Treatment on Corneal Epithelial Cells Using Next-Generation Sequencing and Bioinformatics Approaches" Medicina 55, no. 9: 589. https://doi.org/10.3390/medicina55090589
APA StyleChang, W.-A., Hsiao, Y.-T., Lin, H.-C., Jian, S.-F., Chen, Y.-J., & Kuo, P.-L. (2019). Deduction of Novel Genes Potentially Involved in the Effects of Very Low Dose Atropine (0.003%) Treatment on Corneal Epithelial Cells Using Next-Generation Sequencing and Bioinformatics Approaches. Medicina, 55(9), 589. https://doi.org/10.3390/medicina55090589