1. Introduction
An epitope is a portion of an antigenic protein molecule that can be identified by the immune system of the human body by either B or T lymphocyte cells. The peptides can be interacted with through the T-cell receptors of the immune system. After being bound to at least one Major Histocompatibility Complex (MHC) protein, the antigenic proteins are intracellularly processed and exposed on the surface of the antigen presenting cell, known as the MHC-peptide complex. Since T lymphocytes play an active role in this type of immune response along with the specific MHC molecule from the antigen presenting cell for antigen presentation, this type of epitopes is therefore called a T-cell epitope [
1]. MHC molecules are cell surface glycoproteins, which actively contribute to host immune reactions. These MHC molecules are expressed from human HLA (Human Leukocyte Antigens) gene to participate in adaptive immune system in humans [
2].
The immunogenicity of T cell epitope is dependent on various factors e.g., suitable and actual peptide processing from its protein source, stable small peptide binding with the MHC molecule and lastly, recognition of the MHC-bound peptide molecule by the T cell receptor [
3]. Two types of MHC molecules known as MHC class I molecules normally present peptides containing 8 to 11 amino acids in length, but the peptides binding to MHC class II may have peptide lengths that vary from 12 to 25 amino acids [
4]. MHC class II protein molecules bind with fragments of the oligopeptide, obtained from the proteolytic cleavage of antigenic protein and present them on the cell surface of antigen presenting cells (APCs), to be recognized by CD4+ T cells. When adequate amounts of the epitope are presented, the T cell may generate a specific adaptive immune response for that pathogen through the process of positive selection. Class II MHC molecules are secreted on specialized cells, e.g., professional APCs such as B cells, macrophages and dendritic cells, whereas class I MHCs are expressed on every nucleated cell of the human body [
5].
In an ideal peptide-based vaccine, both B- and T-cell epitopes are present [
2]. B cell and T cell epitopes are recognized by two different pathways. The three-dimensional conformation of the antigenic protein molecule is wholly responsible for recognition by B cells, whereas T cells can recognize an antigen protein molecule only after it has been digested to form a small peptide fragment which must be bound with the major histocompatibility complex (MHC) class II molecule, forming a ternary complex.
Allergic symptoms are one of the most common health problems in the world. Among the four types of hypersensitivity reactions, more than 25% of the world’s population suffers from a type I hypersensitivity reaction. Considering the various causes of allergic reactions, pollen allergens are predicted to be the most potential source of hypersensitivity reaction. Pollinosis due to various pollens results in allergic rhinitis and asthma [
6]. Grass pollen allergens from
Cynedondactylon,
Orzya sativa,
Zea mays etc. are accountable for the allergic reaction in susceptible individuals in different parts of the world [
7]. Allergic diseases can be successfully treated by identifying clinically important allergens. Worldwide, about 400 million people are suffering from hay fever as well as seasonal asthma [
8]. The major causative biomolecules for this allergy are pollen proteins also known as the group-1 grass pollen allergens [
9]. Zea m 1 is found in a class of abundant grass pollen allergens, which are formed by several genes. These proteins can loosen the walls of grass cells, including the maize stigma and style [
10]. In a study, performed in Portugal on thirty-two children that are under 8 years of age, all are given a positive skin-prick test for grass pollen allergens [
11]. 38% of the children are monosensitized to different grass pollen allergens. The decreasing order of sensitization frequency for that pollens are shown in
Table 1.
To prevent the type I hypersensitivity reaction, drugs known as mast cell stabilizers e.g., synthetic, semi synthetic and natural stabilizers, can be used as therapeutic agents. These mast cell stabilizers obtained from natural resources can be classified into different groups such as flavonoids, coumarins, phenols [
12], terpenoids, alkaloids etc. Considering their mechanisms of action, mast cell stabilizers can be classified as Ca
2+channel blocking agents, suppressors of gene expression of genes (tumor necrosis factor alpha, different interleukins), inhibitors for phosphorylation reactions in MAPK, ERK, JNK Gab2 signaling pathways, down regulators of enzyme histidine dicarboxylase, suppressors of mRNA of CD23, inhibitors of COX2/5 lipooxygenase enzyme, inhibitors of prostaglandin D
2 synthesis, LTC
4 inhibitors, and Spleen tyrosin kinase enzyme inhibitors [
13]. Explorations for alternative types of therapeutics in allergic reactions are explained in my earlier work where, a specific sense siRNA is explored as anti-allergic therapeutic during an immediate type of hypersensitivity reaction, caused by the Zea m 1 pollen allergenic protein [
14]. Presently, in allergen-specific immunotherapy (AIT), the disease-causing allergens are used for a disease-modifying treatment of allergy. The molecular allergen characterization process is applied, to produce allergy vaccines with the recombinant allergens, peptides and genes synthesizing allergens. The B-cell epitope technique is also another promising method used to identify the antigenic determinants or epitopes present in the antigenic proteins [
15]. Epitope based vaccine design is a state-of-art method, because it is very specific, is able to evade undesirable immune reactions, has the power to create long lasting immunity, and at the same time, it is cheap in price. This method has been applied to treat various diseases like tuberculosis [
1], Nipah virus infection [
16] etc. In this study, an epitope-based peptide vaccine design method is studied for Zea m 1 pollen allergen, with various T cell epitope prediction methods, followed by molecular docking technique. Prediction methods and docking experiments are performed to design peptide-based vaccines for an allergic reaction caused by Zea m 1 pollen allergen.
4. Discussion
Allergens cause type I hypersensitivity reactions mediated by immunoglobulin E (IgE) molecule. IgE biosynthesis, also known as sensitization, may be caused by airborne allergens, food allergens, drug allergens and occupational allergens. Modern clinical therapeutics for allergic reactions includes a combination of patient awareness, allergenic molecule avoidance, pharmacotherapy, and allergy immunotherapy. Allergy immunotherapy is a type of treatment targeting the basic immunological molecules and immunological pathways involved in allergic reaction and resulting in the activation of immunological tolerance by reducing IgE molecule reactivity and retaining T cell molecule reactivity [
24].A vast array of structurally altered allergens has been created, including allergenic oligopeptides, chemically modified allergoids, adjuvant-bound allergens, and nanoparticle-based allergy vaccines. In allergen-specific immunotherapy (AIT), repeated doses of sensitive allergens are used for desensitization or hypo-sensitization of allergic patients. Several herbal and natural products are reported to regulate antigen-IgE mediated allergic responses [
25,
26]. There are other types of immunotherapeutic strategies that have also reported which use idiotype and anti-idiotype antibody interaction in vaccine production [
27] and epitope-paratope peptide modulation [
28,
29]. Different bioinformatic algorithms, as well as computational methods [
30] are used for identifying biological functions of peptides.
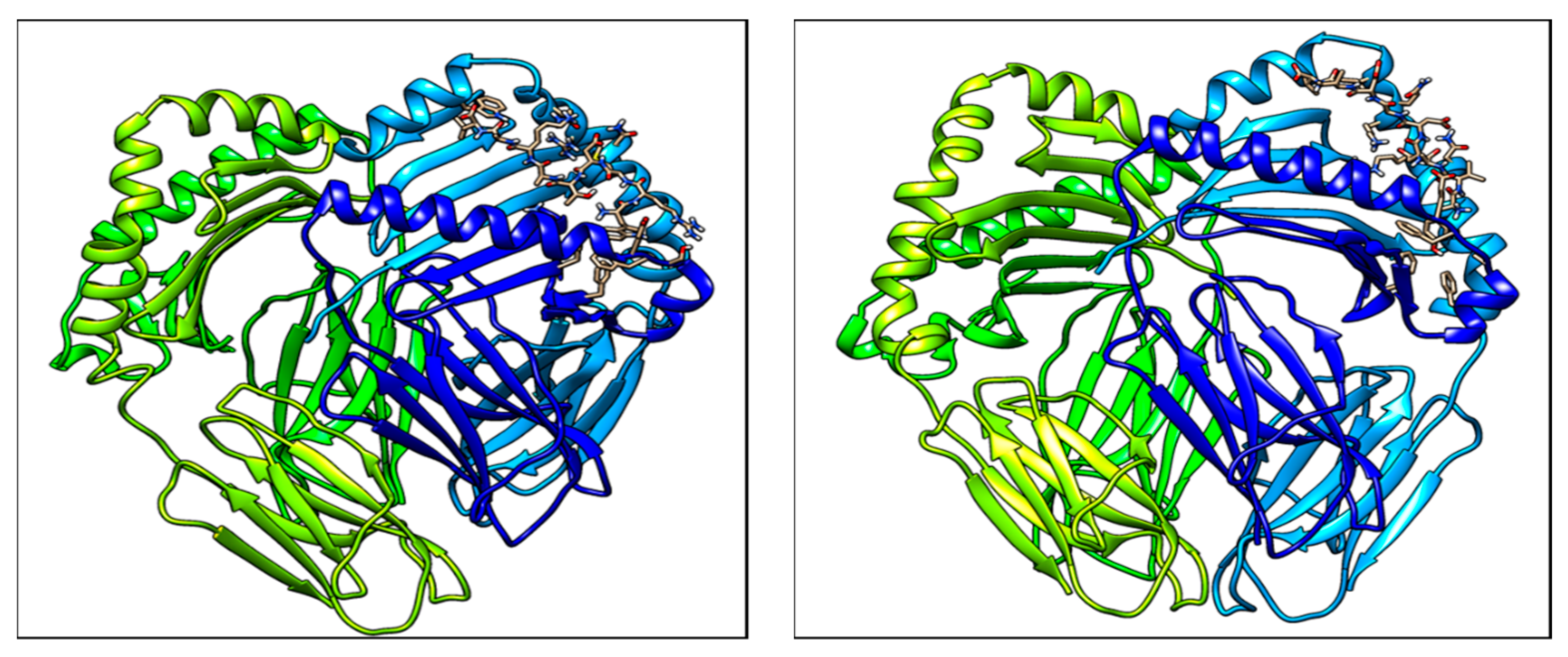
Epitope-based vaccines are short oligopeptides derived from antigen that are used after antigen presentation to T-cells to prevent diseases like type I hypersensitivity. During antigen presentation, epitopes are bound with major histocompatibility complex (MHC) protein molecules. Peptide vaccines based on multiple T-cell epitopes can be administered for the rational use of immunogens among distinct ethnic populations, while providing numerous potential advantages over conventional vaccines. The advantages are more accurate regulation of the immune response activation, concentrating on most appropriate antigenic regions of a group of proteins (which are conserved and immunodominant in nature), as well as having advantages for production and biosafety. CD4+ T-cell epitopes display an important role in epitope-based vaccine design. The help of these cells is indispensable for the production of strong humoral and cytotoxic CD8+ T-cell responses. But the immune response to T-cell epitopes is limited only by HLA proteins. As a result, the HLA selectivity for T-cell epitopes develops a major constraint for epitope-based vaccine design, for genetically varied human populations. Two important factors cause major problems in epitope-based vaccine design. The most common problem is that MHC class II alleles are synthesized in different amounts in different ethnicities such as Asian and European populations. The second problem is that MHC class II genes are the most polymorphic genes in nature found in the human genome. Since the experimental testing of large sets of peptides of MHC molecules is very time-consuming as well as costly, in silico methods are more sought after methods to overcome these problems. Criteria-based analysis predicts two epitopes on this pollen allergen: YVADDGDIV and WRMDTAKAL. The T cell epitopes identified in this study provide insight into a peptide-based vaccine for type I hypersensitivity reaction induced by the Zea m 1 pollen allergen.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}