Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer


Abstract
1. Introduction
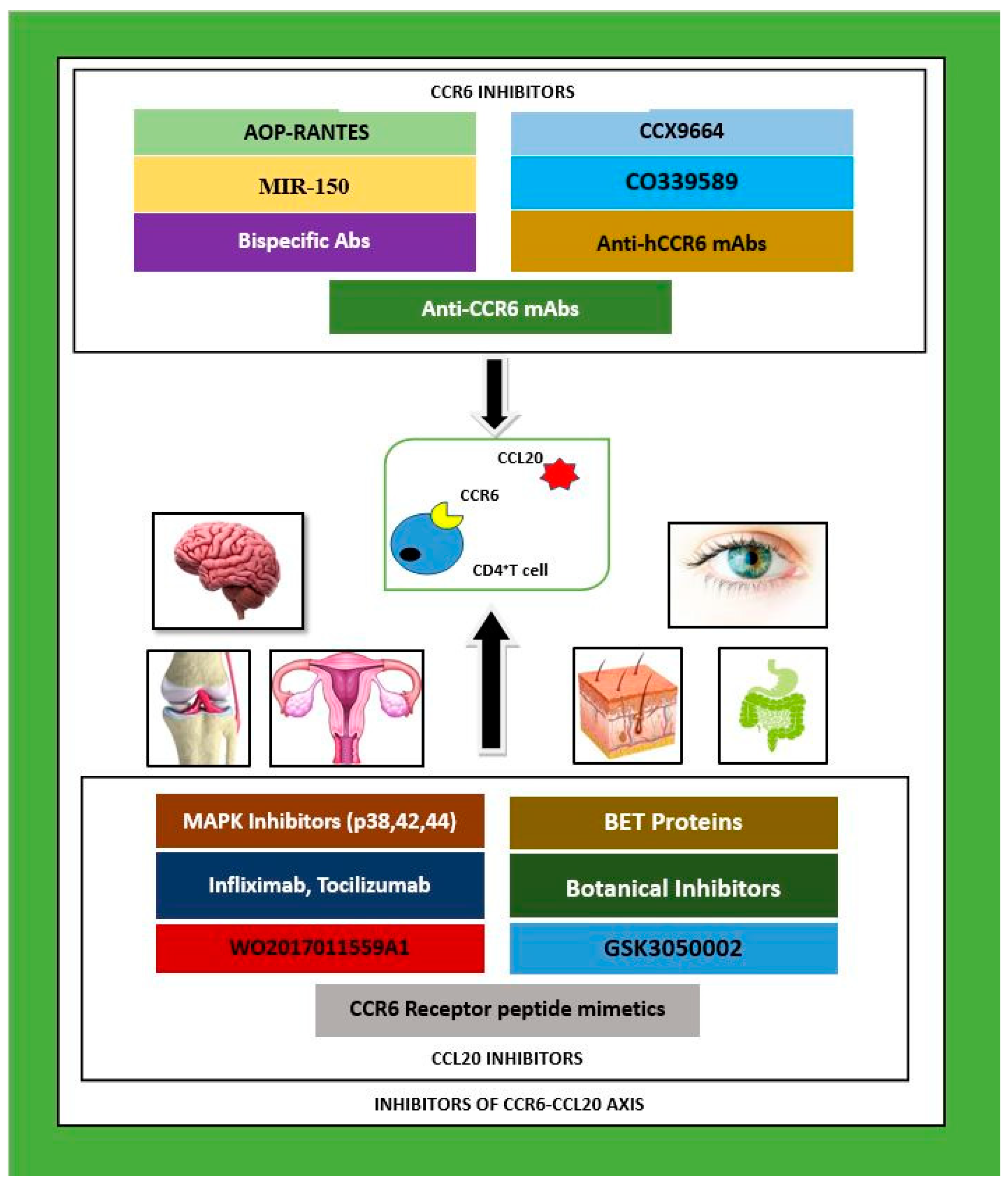
2. Inhibitors of CCR6–CCL20
3. CCR6–CCL20 Inhibition in Multiple Organ Systems
3.1. Nervous System
3.2. Integumentary System
3.3. Skeletal System
3.4. Reproductive System
3.5. Gastrointestinal System
4. Carcinoma Related Studies
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, A.Y.S.; Eri, R.; Lyons ABGrimm, M.C.; Korner, H. CC chemokine ligand CCL20 and its cognate receptor CCR6 in mucosal T cell immunology and inflammatory bowel disease: Odd couple or axis of evil? Front. Immunol. 2013, 4, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O.; Nomiyama, H. The chemokine and chemokine receptor super families and their molecular evolution. Genome Biol. 2006, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Basheer, W.; Kunde, D.; Eri, R. Role of chemokine ligand CCL20 and its receptor CCR6 in intestinal inflammation. Immunol. Infect. Dis. 2013, 1, 30–37. [Google Scholar] [CrossRef]
- Ranasinghe, R.; Eri, R. CCR6–CCL20-Mediated Immunologic Pathways in Inflammatory Bowel Disease. Gastrointest. Disord. 2018, 1, 15–29. [Google Scholar] [CrossRef]
- Ranasinghe, R.; Eri, R. CCR6–CCL20 Axis in IBD: What Have We Learnt in the Last 20 Years? Gastrointest. Disord. 2018, 1, 57–74. [Google Scholar] [CrossRef]
- Lu, M.Y.; Lu, S.S.; Chang, S.L.; Liao, F. The phosphorylation of CCR6 on distinct Ser/Thr residues in the carboxyl terminus differentially regulates biological function. Front. Immunol. 2018, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Scheerens, H.; Hessel, E.; deWaal-Malefyt, R.; Leach, M.W.; Rennick, D. Characterization of chemokines and chemokine receptors in two murine models of inflammatory bowel disease: IL-10-/- mice and Rag-2-/- mice reconstituted with CD4+CD45RBhigh T cells. Eur. J. Immunol. 2001, 31, 1465–1474. [Google Scholar] [CrossRef]
- Teramoto, K.; Miura, S.; Tsuzuki, Y.; Hokari, R.; Watanabe, C.; Inamura, T.; Ogawa, T.; Hosoe, N.; Nagata, H.; Ishii, H.; et al. Increased lymphocyte trafficking to colonic microvessels is dependent on MAdCAM-1 and C-C chemokine mLARC/CCL20 in DSS –induced mice colitis. Clin. Exp. Immunol. 2005, 139, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Atreya, R.; Neurath, M.F. Chemokines in inflammatory bowel diseases. Dig. Dis. 2010, 28, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Pezoldt, J.; Huehn, J. Tissue specific induction of CCR6 and Nrp1 during early CD4+ T cell differentiation. Eur. J. Microbiol. Immunol. (Bp) 2016, 6, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, R.; Eri, R. Pleiotropic immune functions of chemokine receptor 6 in health and disease. Medicines 2018, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Lee AYPhan, T.K.; Hulett MDKörner, H. The relationship between CCR6 and its binding partners: Does the CCR6-CCL20 axis have to be extended? Cytokine 2015, 72, 97–101. [Google Scholar] [CrossRef]
- Proudfoot, A.E. Chemokine receptors: Multifaceted therapeutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Carson, W.F., 4th; Cavassani, K.A.; Connett, J.M.; Kunkel, S.L. CCR6 as a mediator of immunity in the lung and gut. Exp. Cell Res. 2011, 317, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Comerford, I.; Bunting, M.; Fenix, K.; Haylock-Jacobs, S.; Litchfield, W.; Harata-Lee, Y.; Turvey, M.; Brazzatti, J.; Gregor, C.; Nguyen, P. An immune paradox: How can the same chemokine axis regulate both immune tolerance and activation? CCR6/CCL20, a chemokine axis balancing immunological tolerance and activation in autoimmune disease. Bioessays 2010, 32, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Van Vlerken-Ysla, L.E.; Rios-Doria, J.; Moynihan, J.; Shan, L.; Hollingsworth, R.E.; Herbst, R.; Hurt, E.M. Abstract 4779: Targeting the CCL20-CCR6 axis as a novel opportunity to simultaneously modulate cancer stem cells and the tumor–immune infiltrate by a dual anti-cancer mechanism. Cancer Res. 2017, 77 (Suppl. 13), 4779. [Google Scholar] [CrossRef]
- Lee, A.Y.S.; Korner, H. CCR6 and CCL20: Emerging players in the pathogenesis of rheumatoid arthritis. Immunol. Cell Biol. 2014, 92, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Getschman, A.E.; Imai, Y.; Larsen, O.; Peterson, F.C.; Wu, X.; Rosenkilde, M.M.; Hwang, S.T.; Volkman, B.F. Protein engineering of the chemokine CCL20 prevents psoriasiform dermatitis in an IL-23 –dependent murine model. Proc. Natl. Acad. Sci. USA 2017, 114, 12460–12465. [Google Scholar] [CrossRef] [PubMed]
- Robert, R.; Juglair, L.; Lim, E.X.; Ang, C.; Wang, C.J.H.; Ebert, G.; Dolezal, O.; Mackay, C.R. A fully humanized IgG-like bispecific antibody for effective dual targeting of CXCR3 and CCR6. PLoS ONE 2017, 12, e0184278. [Google Scholar] [CrossRef] [PubMed]
- Klamar, C.R. Development of a Robust and Improved System for Studying Interactions between CCL20 and CCR6 Using Both Recombinant and Chemically Synthesized Rhesus Macaque Chemokines. Master’s Thesis, University of Pittsburgh, Pittsburgh, PA, USA, 2011. [Google Scholar]
- Liston, A.; Kohler, R.E.; Townley, S.; Comerford, I.; Caon, A.C.; Webster, J.; Harrison, J.M.; Swann, J.; Clark-Lewis, I.; Korner, H.; et al. Inhibition of CCR6 function reduces severity of experimental autoimmune encephalomyelitis via effects on the priming phase of the immune response. J. Immunol. 2009, 182, 3121–3130. [Google Scholar] [CrossRef] [PubMed]
- Eskandarpour, M.; Alexander, R.; Adamson, P.; Calder, V.L. Pharmacological inhibition of bromodomain proteins suppresses retinal inflammatory disease and downregulates retinal Th 17 cells. J. Immunol. 2017, 198, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.H.; Chang, S.Y.; Lee, H.J.; Choi, S.H. The C-C chemokine receptor 6 (CCR6) is crucial for Th2 –driven allergic conjunctivitis. Clin. Immunol. 2015, 161, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.J.; Ebsworth, K.; Ertl, L.S.; McMahon, J.P.; Newland, D.; Wang, Y.; Liu, S.; Miao, Z.; Dang, T.; Zhang, P.; et al. IL-17-secreting γδT cells are completely dependent upon CCR6 for homing to inflamed skin. J. Immunol. 2017, 199, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Ebsworth, K.; Ertl, L.S.; Wang, H.; Campbell, J.J.; McMahon, J.P.; Zhang, P.; Charo, I.F.; Schall, T.J. 521 Chemokine receptor inhibition as a novel therapeutic approach for psoriasis. In Proceedings of the Annual Meeting of Society for Investigative Dermatology, Portland, OR, USA, 11–14 May 2016. [Google Scholar]
- Campbell, J.J.; Ebsworth, K.; Ertl, L.; McMahon, J.P.; Zhang, P.; Singh, R.; Schall, T.J. 669 chemokine receptor CCR6 antagonist reverses psoriaform dermatitis by preventing accumulation of γδT 17 cells in skin. J. Investig. Dermatol. 2017, 137 (Suppl. 1), S115. [Google Scholar] [CrossRef]
- Bouma, G.; Zamuner, S.; Hicks, K.; Want, A.; Oliveira, J.; Choudhury, A.; Brett, S.; Robertson, D.; Felton, L.; Norris, V. CCL20 neutralization by a monoclonal antibody in healthy subjects selectively inhibits recruitment of CCR6+ cells in an experimental suction blister. Br. J. Clin. Pharmacol. 2017, 83, 1976–1990. [Google Scholar] [CrossRef] [PubMed]
- Dairaghi, D.; Zhang, P.; Leleti, M.; Berahovich, R.; Ebsworth, K.; Ertl, L.; Miao, S.; Miao, Z.; Seitz, L.; Tan, J. et al. In Inhibition of chemokine receptors CCR1 and CCR6 as promising therapies for autoimmune diseases such as rheumatoid arthritis and psoriasis. In Proceedings of the ACR/ARHP Annual Meeting, San Diego, CA, USA, 25–30 October 2013. Abstract No. 1792. [Google Scholar]
- Jaen, J.C.; Dairaghi, D.; Leleti, M.; Powers, M.J.P.; Wang, Y.; Zhang, P.; Schall, T.J. FRI0002 Inhibition of chemokine receptors CCR1 and CCR6 as promising therapies for autoimmune diseases such as rheumatoid arthritis. Ann. Rheumetic Dis. 2013, 72 (Suppl. 3), A369. [Google Scholar] [CrossRef]
- Hirata, T.; Osuga, Y.; Takamura, M.; Kodama, A.; Hirota, Y.; Koga, K.; Yoshino, O.; Harada, M.; Takemura, Y.; Yano, T.; et al. Recruitment of CCR6—Expressing Th17 cells by CCL20 secreted from IL-1 beta, TNF- alpha, and IL-17A-stimulated endometriotic stromal cells. Endocrinology 2010, 151, 5468–5476. [Google Scholar] [CrossRef] [PubMed]
- Blazquez, A.B.; Knight, A.K.; Getachew, H.; Bromberg, J.S.; Lira, S.A.; Mayer, L.; Berin, M.C. A functional role for CCR6 on pro-allergic T cells in the gastrointestinal tract. Gastroenterology 2010, 138, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Yang, H.; Tartar, D.M.; Gao, B.; Luo, X.; Ye, S.Q.; Zaghouani, H.; Fang, D. Prevention and treatment of diabetes with resveratrol in a non-obese mouse model of type 1 diabetes. Diabetologia 2011, 54, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ke, F.; Xu, Z.; Liu, Z.; Zhang, L.; Yan, S.; Wang, Z.; Wang, H.; Wang, H. CCR6 is a prognostic marker for overall survival in patients with colorectal cancer and its overexpression enhances metastasis in vivo. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Marsigliante, S.; Vetrugno, C.; Muscella, A. CCL20 induces migration and proliferation on breast epithelial cells. J. Cell Physiol. 2013, 228, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Kitadate, A.; Ito, M.; Abe, F.; Nara, M.; Watanabe, A.; Takahashi, N.; Miyagaki, T.; Sugaya, M.; Tagawa, H. Disruption of CCL20-CCR6 interaction inhibits metastasis of advanced cutaneous T cell lymphoma. Oncotarget 2016, 7, 13563–13574. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Teshima, K.; Ikeda, S.; Kitadate, A.; Watanabe, A.; Nara, M.; Yamashita, J.; Ohshima, K.; Sawada, K.; Tagawa, H. MicroRNA-150 inhibits tumor invasion and metastasis by targeting the chemokine receptor CCR6, in advanced cutaneous T-cell lymphoma. Blood 2014, 123, 1499–1511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Hu, Z.J.; Meng, A.H.; Duan, G.C.; Zhao, Q.T.; Yang, J. Role of CCL20/CCR6 and the ERK signaling pathway in lung adenocarcinoma. Oncol. Lett. 2017, 14, 8183–8189. [Google Scholar] [CrossRef] [PubMed]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Kryczek, I.; Nagarsheth, N.; Zhao, L.; Wei, S.; Wang, W.; Sun, Y.; Zhao, E.; Vatan, L.; Szeliga, W.; et al. Epigenetic silencing of TH1 -type chemokines shapes tumor immunity and immunotherapy. Nature 2015, 527, 249–253. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Inhibitor | Name of Inhibitor | Nature of Inhibitor | Immune Cells/Cytokines Involved | Experimental Outcome | Name of Disease/Model | Ref |
|---|---|---|---|---|---|---|
| Anti-hCCR6 mAbs | CCR6 Inhibitor | TH17, B cells | Reduced inflammatory cell infiltration | Experimental Autoimmune Encephalitis (EAE) | [20] | |
| Anti-CCR6 Abs | CO339589 | CCR6 inhibitor | NK cells PBMC | Reduced disease | RA, psoriasis, MS | [30] |
| Anti CCL20 Abs | WO2017011559A1 | CCL20 inhibitor | TH17, Treg | Inhibits CSC activity, tumorigenesis | Carcinoma | [17] |
| GSK3050002 | CCL20 inhibitor | TH17 | Reduced cell chemotaxis | Suction blister in skin | [28] | |
| CCL20 inhibitor | γδ T cells | Reduced immune cell infiltration to skin | Imiquimod (IMQ) induced psoriasiform dermatitis | [19] | ||
| Anti-TNF-α Abs | Infliximab Tocilizumab | Indirect CCL20 inhibitor | Synoviocytes | Inhibits TNF-α induced CCL20 upregulation | RA | [18] |
| Inhibitors of p38, 42, 44 | MAPK Inhibitors | Indirect CCL20 inhibitor | TH17 | Reduced pro-inflammatory cytokines, suppressed CCL20 | Ovarian endometritis | [31] |
| Partial Agonists | AOP-RANTES | Partial CCR6 inhibitor | TH17 | Reduced inflammatory cell chemotaxis | Psoriasiform dermatitis | [19] |
| Bispecific Abs | Humanised IgG-like BsAb | CCR6 and CXCR3 inhibitors | TH17 | Blocks cell chemotaxis | Murine model | [20] |
| BET Proteins | Gene regulator | TH17, Treg | Reduced inflammation | Posterior uveitis | [23] | |
| Small Molecule Inhibitors | CCX9664 | CCR6 inhibitor | TH17 | Reduced disease | Rheumatoid arthritiis (RA) | [29] |
| Botanical Inhibitors | EGCG, Gallotannin | CCL20 inhibitor | TH17 | Inhibited cell chemotaxis | Rhesus Macaque model | [21] |
| Sphingosine 1 PO4 R agonist | FTY720 | CD4+T Cells | Inhibited egress of lymphocytes from LNs | Allergic diarrhea | [32] | |
| microRNA | MiR-150 | CCR6 inhibitor | CTCL cells | Reduced metastasis | Cutaneous T cell lymphoma (CTCL) | [37] |
| Polyphenol compound | Resveratrol | Sirtuin-1 activator | TH17, macrophages | Reduced disease, inflammatory cytokines | Type-1 diabetes, RA, lupus, EAE | [33] |
| CCR6R Peptide Mimetics | ECL-2 | CCL20 onhibitor | CCR6+ cells | Reduced cell chemotaxis | Rhesus Macaque model | [21] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranasinghe, R.; Eri, R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina 2018, 54, 88. https://doi.org/10.3390/medicina54050088
Ranasinghe R, Eri R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina. 2018; 54(5):88. https://doi.org/10.3390/medicina54050088
Chicago/Turabian StyleRanasinghe, Ranmali, and Rajaraman Eri. 2018. "Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer" Medicina 54, no. 5: 88. https://doi.org/10.3390/medicina54050088
APA StyleRanasinghe, R., & Eri, R. (2018). Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina, 54(5), 88. https://doi.org/10.3390/medicina54050088