
Full-Length Transcriptome Sequencing-Based Analysis of Pinus sylvestris var. mongolica in Response to Sirex noctilio Venom


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Venom and Plant-Material Collection and Inoculation Trial
2.2. PacBio Iso-Seq Library Preparation and Sequencing
2.3. Iso-Seq Data Analysis
2.4. Illumina Library Preparation and Sequencing
2.5. Gene Functional Annotation
2.6. Quantification of Unigene Expression Levels and Analysis of DEGs
3. Results
3.1. Full-Length Transcriptome Sequencing
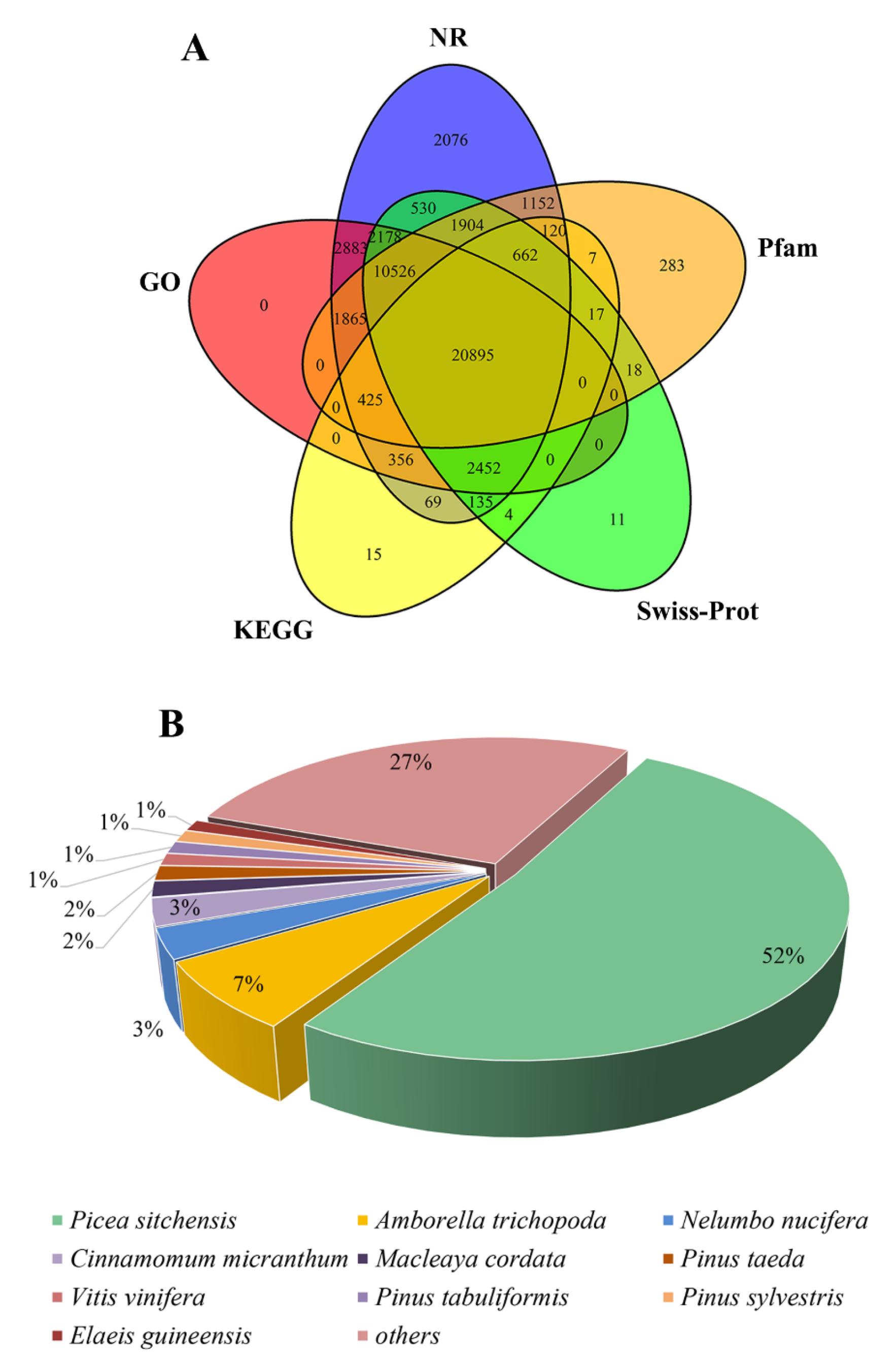
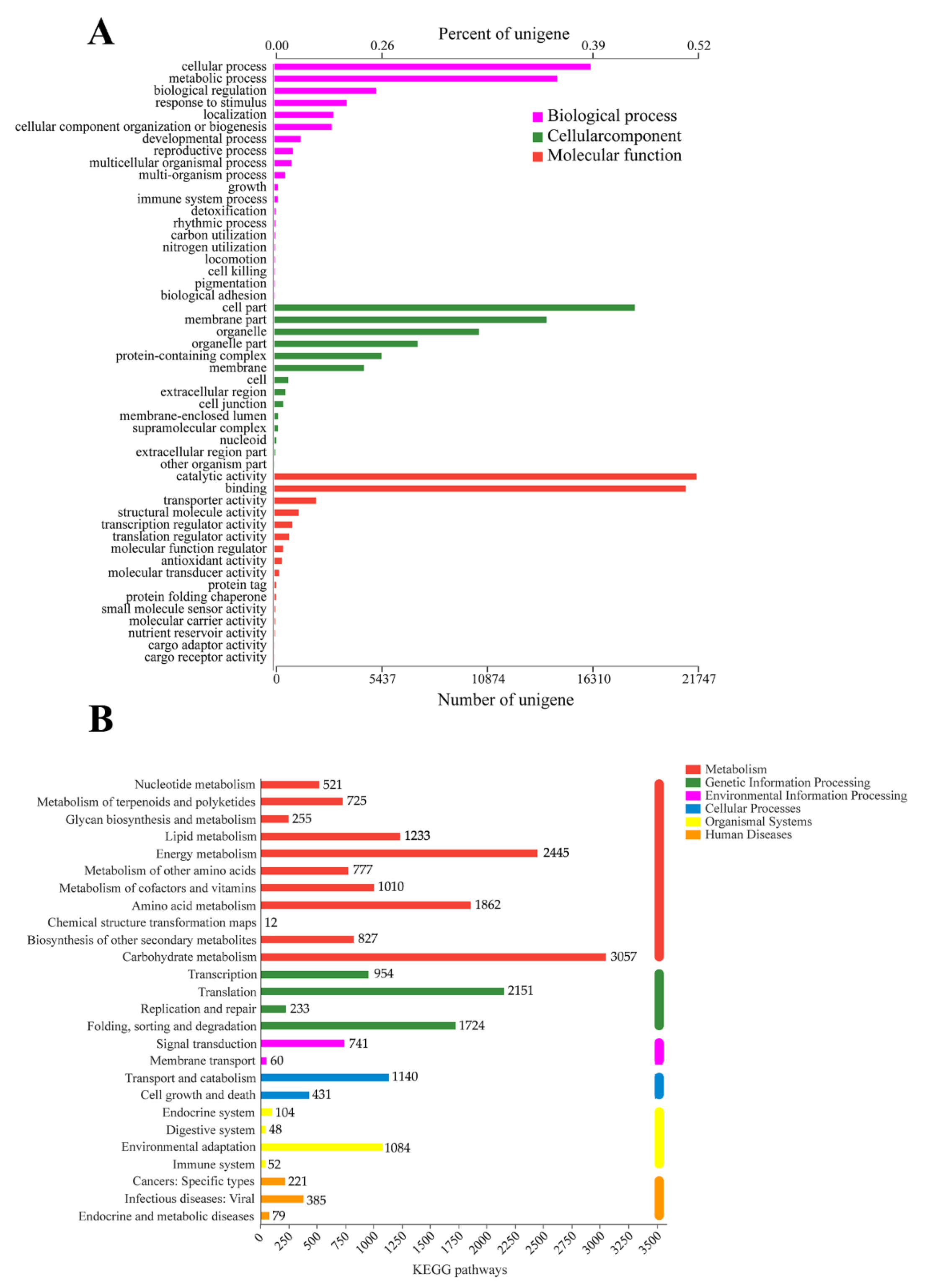
3.2. Functional Annotation of the Full-Length Reference Transcriptome
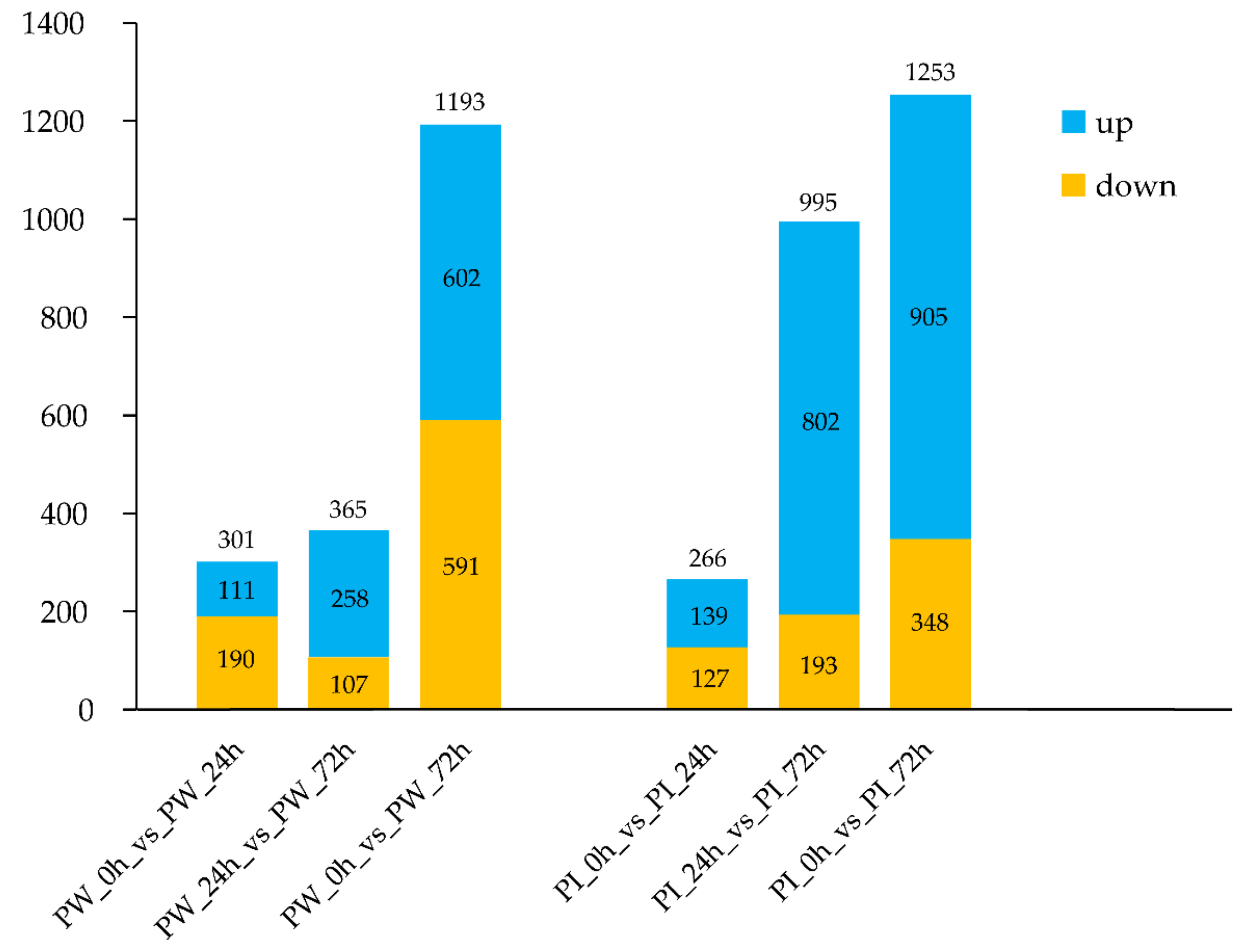
3.3. Identification of DEGs
3.4. Analysis of DEGs in Wounded and Inoculated Mongolian Pine at 72 h
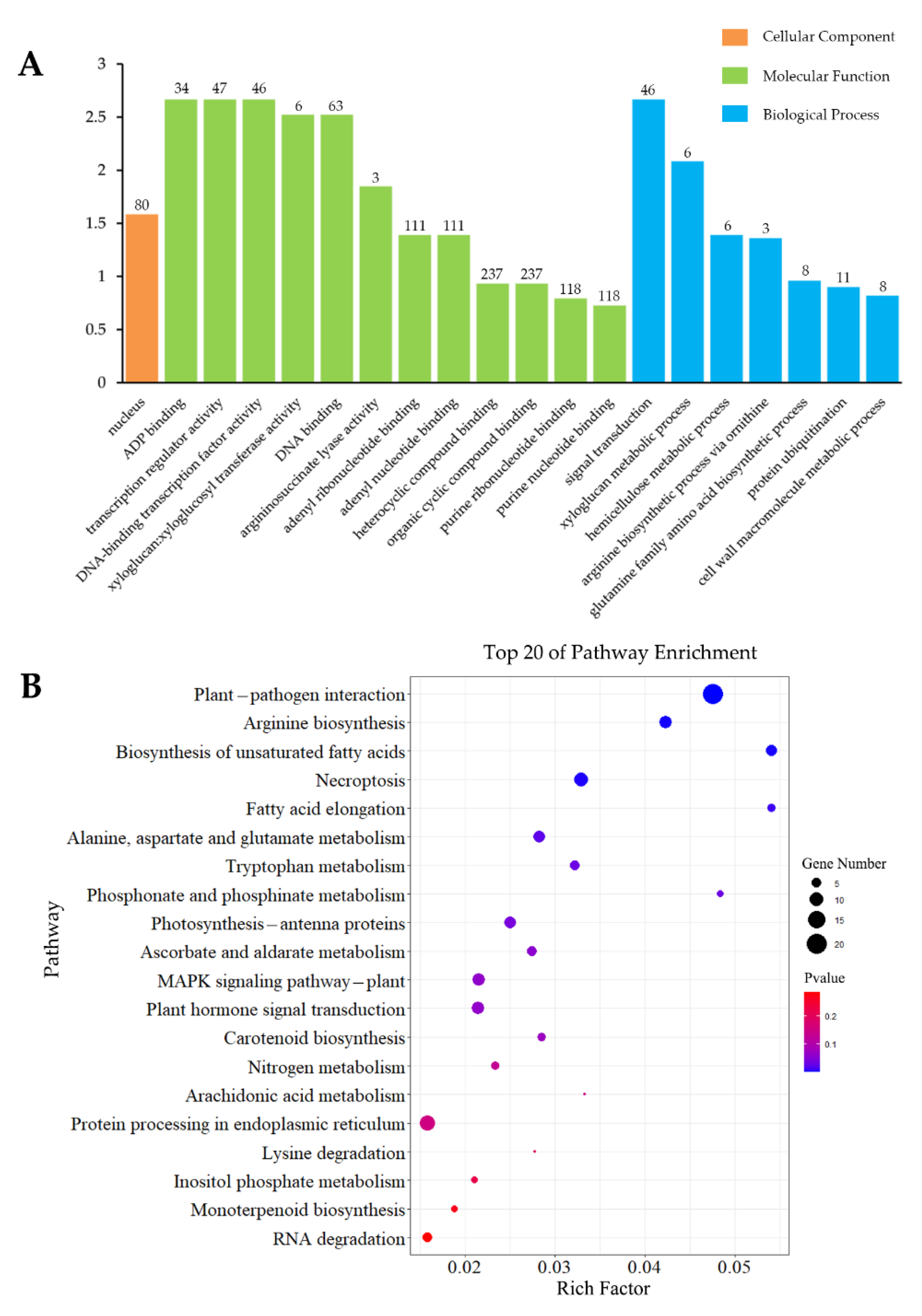
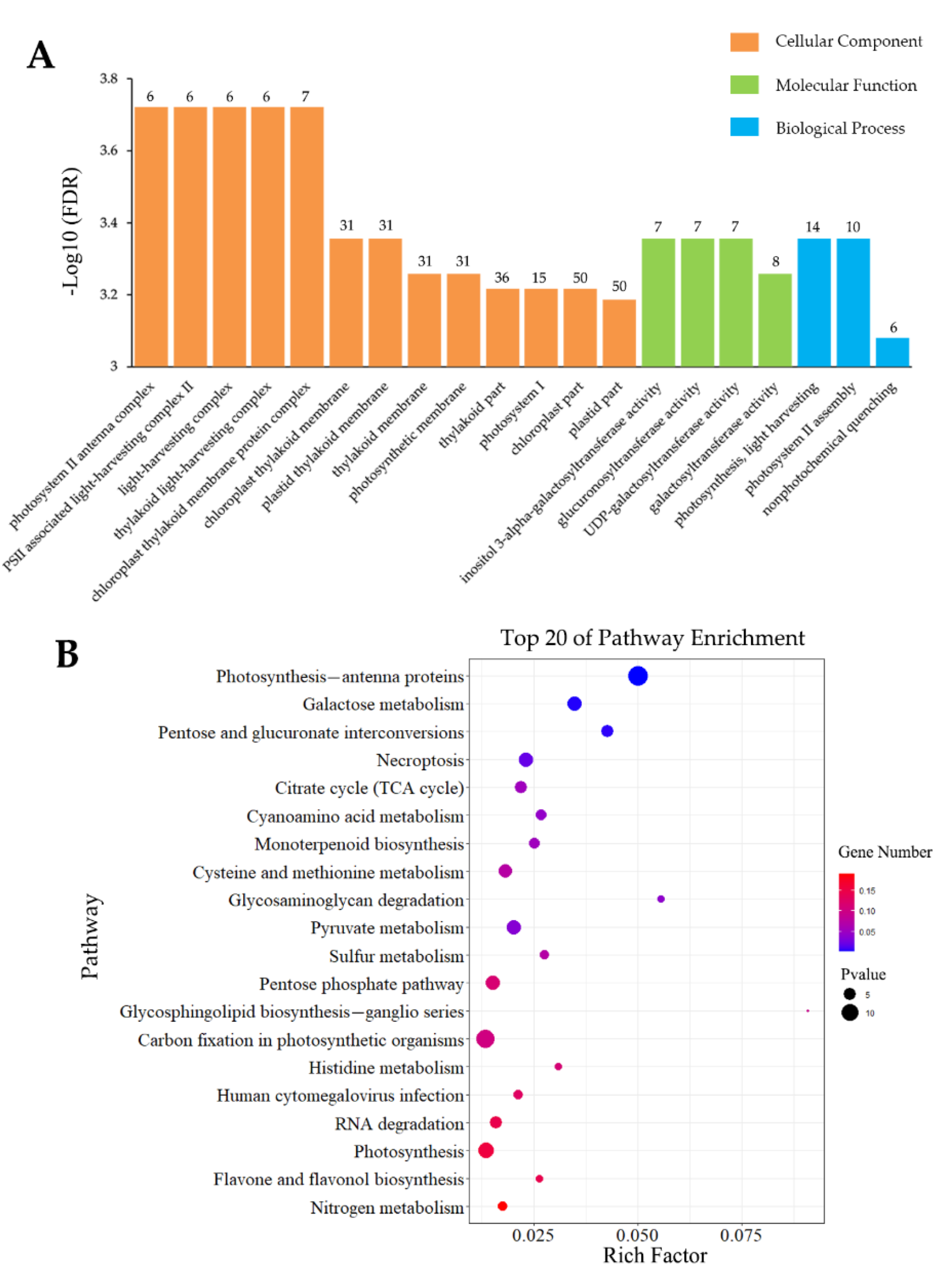
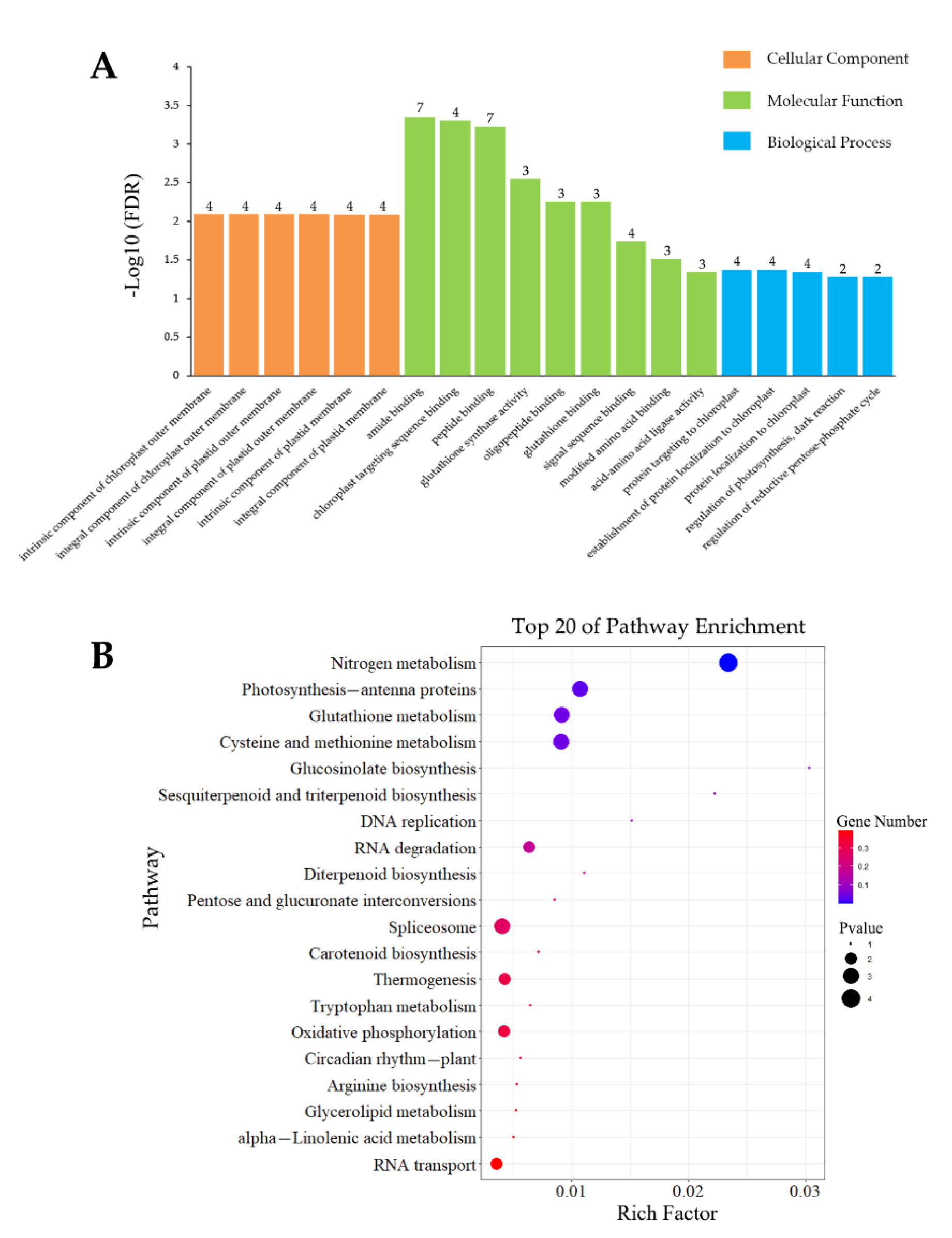
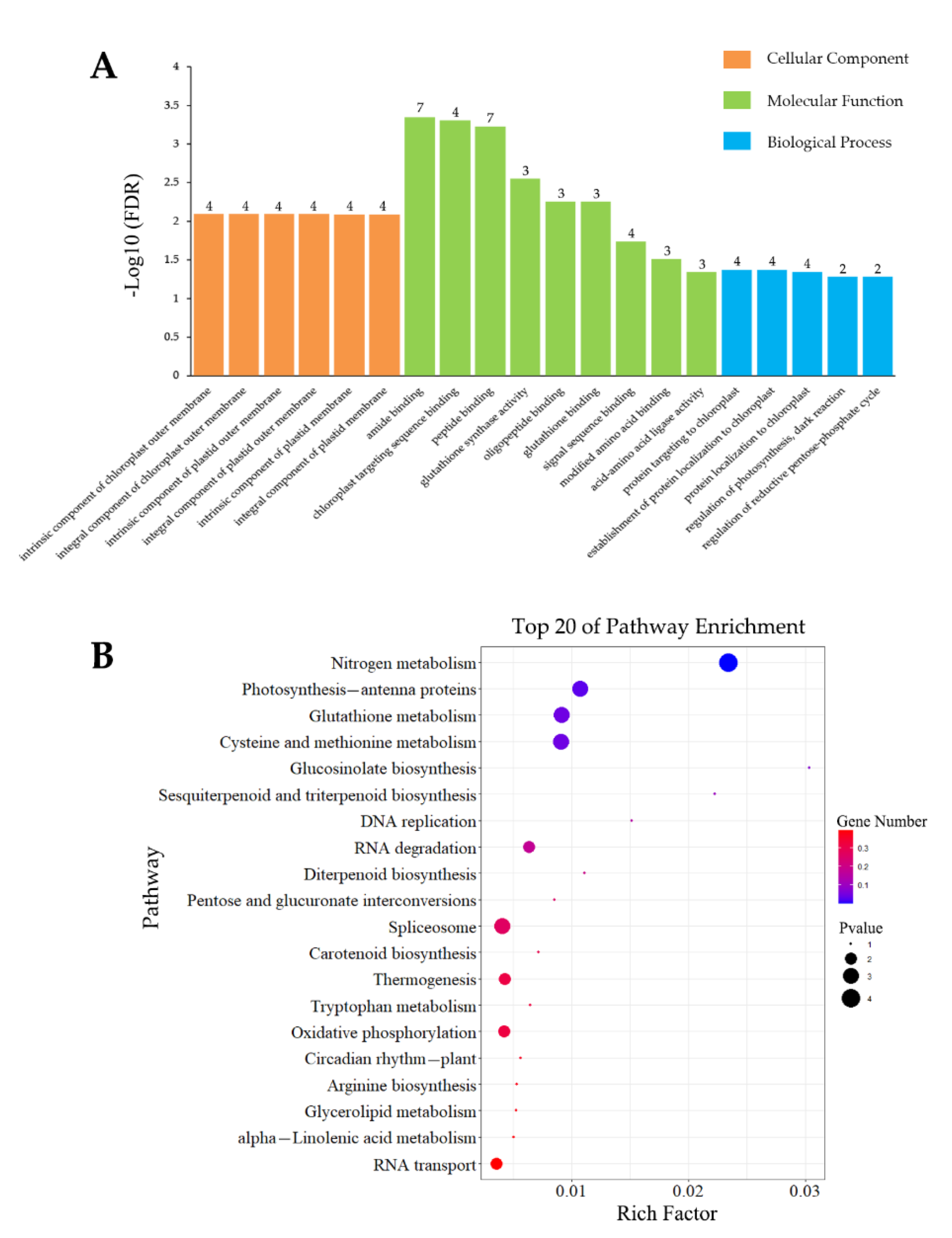
3.4.1. Analysis of Inoculation-Specific DEGs
3.4.2. Analysis of Wounding-Specific DEGs
3.4.3. DEGs Induced by Inoculation and Wounding
3.5. Transcription Factors (TFs) in Response to Inoculation and Wounding in P. sylvestris var. mongolica
4. Discussion
4.1. Signaling Pathways Involved in the Response of P. sylvestris var. mongolica to Wounding and Inoculation
4.2. Expression of Typical Venom-Induced Genes in Mongolian Pine
4.3. Expression of Typical Wounding-Induced Genes in Mongolian Pine
4.4. The Role of TFs in Response to Inoculation and Wounding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, J.-J.; Li, F.-Q.; Xu, M.-L.; Kang, H.-Z.; Wu, X.-Y. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China: An experimental approach. Ann. For. Sci. 2008, 65, 304. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Zhu, J.; Yan, Q.; Li, M.; Yu, G. Comparison of intrinsic water use efficiency between different aged Pinus sylvestris var. mongolica wide windbreaks in semiarid sandy land of northern China. Agrofor. Syst. 2015, 89, 477–489. [Google Scholar] [CrossRef]
- Jiao-Jun, Z.; Zhi-Ping, F.; De-Hui, Z.; Feng-Qi, J.; Takeshi, M. Comparison of stand structure and growth between artificial and natural forests of Pinus sylvestiris var. mongolica on sandy land. J. For. Res. 2003, 14, 103–111. [Google Scholar] [CrossRef]
- Li, D.; Shi, J.; Lu, M.; Ren, L.; Zhen, C.; Luo, Y. Detection and Identification of the Invasive Sirex noctilio (Hymenoptera: Siricidae) Fungal Symbiont, Amylostereum areolatum (Russulales: Amylostereacea), in China and the Stimulating Effect of Insect Venom on Laccase Production by A. areolatum YQL03. J. Econ. Entomol. 2015, 108, 1136–1147. [Google Scholar] [CrossRef]
- Wang, L.; Ren, L.; Li, C.; Gao, C.; Liu, X.; Wang, M.; Luo, Y. Effects of endophytic fungi diversity in different coniferous species on the colonization of Sirex noctilio (Hymenoptera: Siricidae). Sci. Rep. 2019, 9, 5077. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Q.; Luo, Y. A Maximum Entropy Model Predicts the Potential Geographic Distribution of Sirex noctilio. Forests 2020, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Spradbery, J.P.; Kirk, A.A. Aspects of the ecology of siricid woodwasps (Hymenoptera: Siricidae) in Europe, North Africa and Turkey with special reference to the biological control of Sirex noctilio F. in Australia. Bull. Entomol. Res. 1978, 68, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Kurt, W. Proceedings, 17th US Department of Agriculture Interagency Research Forum on Gypsy Moth and Other Invasive Species 2006; Gen. Tech. Rep. NRS-P-10; US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2007; Volume 10, p. 117. [Google Scholar]
- Coutts, M.; Dolezal, J. Some effects of bark cincturing on the physiology of Pinus radiata, and on Sirex attack. Aust. For. Res. 1966, 2, 17–28. [Google Scholar]
- Borchert, D.; Fowler, G.; Jackson, L. Organism Pest Risk Analysis: Risks to the Conterminous United States Associated with the Woodwasp, Sirex noctilio Fabricius, and the Symbiotic Fungus, Amylostereum areolatum, (Fries: Fries) Boidin; USDA-APHIS-PPQ-EDP: Raleigh, NC, USA, 2007; pp. 1–40. [Google Scholar]
- Corley, J.C.; Lantschner, M.V.; Martínez, A.S.; Fischbein, D.; Villacide, J.M. Management of Sirex noctilio populations in exotic pine plantations: Critical issues explaining invasion success and damage levels in South America. J. Pest Sci. 2019, 92, 131–142. [Google Scholar] [CrossRef]
- Coutts, M.P.; Dolezal, J.E. Polyphenols and Resin in the Resistance Mechanism of Pinus Radiata Attacked by the Wood Wasp, Sirex noctilio, and Its Associated Fungus, leaflet no. 101. Forest Res. Inst. 1966, 1–19. [Google Scholar]
- Coutts, M.P. Rapid physiological change in Pinus radiata following attack by Sirex noctilio and its associated fungus, Amylostereum sp. Aust. J. Sci. 1968, 30, 275–277. [Google Scholar]
- Coutts, M. The Mechanism of Pathogenicity of Sirex noctilio on Pinus Radiata I. Effects of the Symbiotic Fungus Amylostereum sp. (Thelophoraceae). Aust. J. Biol. Sci. 1969, 22, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Coutts, M. The Mechanism of Pathogenicity of Sirex noctilio on Pinus Radiata II. Effects of S. noctilio Mucus. Aust. J. Biol. Sci. 1969, 22, 1153–1162. [Google Scholar] [CrossRef]
- Coutts, M.P. The physiological effects of the mucus secretion of Sirex noctilio on Pinus radiata. Aust. For. Res. 1970, 4, 23–26. [Google Scholar]
- Spradbery, J.P. A comparative study of the phytotoxic effects of siricid woodwasps on conifers. Ann. Appl. Biol. 1973, 75, 309–320. [Google Scholar] [CrossRef]
- Madden, J.L. Physiological reactions of Pinus radiata to attack by woodwasp, Sirex noctilio F. (Hymenoptera: Siricidae). Bull. Èntomol. Res. 1977, 67, 405–426. [Google Scholar] [CrossRef]
- Iede, E.T.; Zanetti, R. Ocorrência e recomendações para o manejo de Sirex noctilio Fabricius (Hymenoptera, Siricidae) em plantios de Pinus patula (Pinaceae) em Minas Gerais, Brasil. Rev. Bras. Entomol. 2007, 51, 529–531. [Google Scholar] [CrossRef] [Green Version]
- Bordeaux, J.M.; Lorenz, W.W.; Dean, J.F. Biomarker genes highlight intraspecific and interspecific variations in the responses of Pinus taeda L. and Pinus radiata D. Don to Sirex noctilio F. acid gland secretions. Tree Physiol. 2012, 32, 1302–1312. [Google Scholar] [CrossRef] [Green Version]
- Fox, H.; Doron-Faigenboim, A.; Kelly, G.; Bourstein, R.; Attia, Z.; Zhou, J.; Moshe, Y.; Moshelion, M.; David-Schwartz, R. Transcriptome analysis of Pinus halepensis under drought stress and during recovery. Tree Physiol. 2017, 38, 423–441. [Google Scholar] [CrossRef] [Green Version]
- Visser, E.A.; Wegrzyn, J.L.; Myburg, A.A.; Naidoo, S. Defence transcriptome assembly and pathogenesis related gene family analysis in Pinus tecunumanii (low elevation). BMC Genom. 2018, 19, 632. [Google Scholar] [CrossRef]
- Ralph, S.G.; Yueh, H.; Friedmann, M.; Aeschliman, D.; Zeznik, J.A.; Nelson, C.C.; Butterfield, Y.S.N.; Kirkpatrick, R.; Liu, J.; Jones, S.J.M.; et al. Conifer defence against insects: Microarray gene expression profiling of Sitka spruce (Picea sitchensis) induced by mechanical wounding or feeding by spruce budworms (Choristoneura occidentalis) or white pine weevils (Pissodes strobi) reveals large-scale changes of the host transcriptome. Plant Cell Environ. 2006, 29, 1545–1570. [Google Scholar] [CrossRef]
- Li, B.; Ruotti, V.; Stewart, R.M.; Thomson, J.A.; Dewey, C.N. RNA-Seq gene expression estimation with read mapping uncertainty. Bioinformatics 2010, 26, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef]
- Yuan, H.; Yu, H.; Huang, T.; Shen, X.; Xia, J.; Pang, F.; Wang, J.; Zhao, M. The complexity of the Fragaria x ananassa (octoploid) transcriptome by single-molecule long-read sequencing. Hortic. Res. 2019, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Steijger, T.; The RGASP Consortium; Abril, J.F.; Engström, P.G.; Kokocinski, F.; Hubbard, T.J.; Guigó, R.; Harrow, J.; Bertone, P. Assessment of transcript reconstruction methods for RNA-seq. Nat. Methods 2013, 10, 1177–1184. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Ghany, S.E.; Hamilton, M.; Jacobi, J.L.; Ngam, P.; Devitt, N.; Schilkey, F.; Ben-Hur, A.; Reddy, A.S.N. A survey of the sorghum transcriptome using single-molecule long reads. Nat. Commun. 2016, 7, 11706. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Tseng, E.; Regulski, M.; Clark, T.A.; Hon, T.; Jiao, Y.; Lu, Z.; Olson, A.; Stein, J.C.; Ware, D. Unveiling the complexity of the maize transcriptome by single-molecule long-read sequencing. Nat. Commun. 2016, 7, 11708. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.-T.; Gernandt, D.S.; Wehenkel, C.; Xia, X.-M.; Wei, X.-X.; Wang, X.-Q. Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proc. Natl. Acad. Sci. USA 2021, 118, e2022302118. [Google Scholar] [CrossRef]
- Wachowiak, W.; Trivedi, U.; Perry, A.; Cavers, S. Comparative transcriptomics of a complex of four European pine species. BMC Genom. 2015, 16, 234. [Google Scholar] [CrossRef] [Green Version]
- Vornam, B.; Leinemann, L.; Peters, F.S.; Wolff, A.; Leha, A.; Salinas, G.; Schumacher, J.; Gailing, O. Response of Scots pine (Pinus sylvestris) seedlings subjected to artificial infection with the fungus Sphaeropsis sapinea. Plant Mol. Biol. Rep. 2019, 37, 214–223. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, K.; Pei, D.; Yu, D.; Zhang, J.; Li, X.; Chen, G.; Yang, H.; Zhou, W.; Li, C. ShORR-1, a Novel Tomato Gene, Confers Enhanced Host Resistance to Oidium neolycopersici. Front. Plant Sci. 2019, 10, 1400. [Google Scholar] [CrossRef] [Green Version]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Peng, Y.; Van Wersch, R.; Zhang, Y. Convergent and Divergent Signaling in PAMP-Triggered Immunity and Effector-Triggered Immunity. Mol. Plant Microbe Interact. 2018, 31, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, W.; Bhattacharjee, S. Effector-Triggered Immunity Signaling: From Gene-for-Gene Pathways to Protein-Protein Interaction Networks. Mol. Plant Microbe Interact. 2012, 25, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.K.; Prajapati, R.; Krishna, D.; Divakaran, K.; Pandey, Y.; Reichelt, M.; Mathew, M.; Boland, W.; Mithöfer, A.; Vadassery, J. The Ca2+ Channel CNGC19 Regulates Arabidopsis Defense Against Spodoptera Herbivory. Plant Cell 2019, 31, 1539–1562. [Google Scholar] [CrossRef]
- Cheval, C.; Aldon, D.; Galaud, J.-P.; Ranty, B. Calcium/calmodulin-mediated regulation of plant immunity. Biochim. Biophys. Acta 2013, 1833, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Vandelle, E.; Vannozzi, A.; Wong, D.; Danzi, D.; Digby, A.-M.; Santo, S.D.; Astegno, A. Identification, characterization, and expression analysis of calmodulin and calmodulin-like genes in grapevine (Vitis vinifera) reveal likely roles in stress responses. Plant Physiol. Biochem. 2018, 129, 221–237. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; Ganthaler, A.; Stöggl, W.; Kranner, I.; Schüler, S.; Ertl, R.; Schlosser, S.; George, J.-P.; Mayr, S. RNA-Seq and secondary metabolite analyses reveal a putative defence-transcriptome in Norway spruce (Picea abies) against needle bladder rust (Chrysomyxa rhododendri) infection. BMC Genom. 2020, 21, 336. [Google Scholar] [CrossRef]
- Orosa, B.; Yates, G.; Verma, V.; Srivastava, A.K.; Srivastava, M.; Campanaro, A.; De Vega, D.; Fernandes, A.; Zhang, C.; Lee, J.; et al. SUMO conjugation to the pattern recognition receptor FLS2 triggers intracellular signalling in plant innate immunity. Nat. Commun. 2018, 9, 5185. [Google Scholar] [CrossRef]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [Green Version]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-Produced Mycotoxins in Plant-Pathogen Interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Geissler, N.; Hussin, S.; Koyro, H.-W. Interactive effects of NaCl salinity and elevated atmospheric CO2 concentration on growth, photosynthesis, water relations and chemical composition of the potential cash crop halophyte Aster tripolium L. Environ. Exp. Bot. 2009, 65, 220–231. [Google Scholar] [CrossRef]
- Zhang, L.-T.; Zhang, Z.-S.; Gao, H.-Y.; Xue, Z.-C.; Yang, C.; Meng, X.-L.; Meng, Q.-W. Mitochondrial alternative oxidase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species in Rumex K-1 leaves. Physiol. Plant. 2011, 143, 396–407. [Google Scholar] [CrossRef]
- Bhat, F.A.; Ganai, B.A.; Uqab, B. Carbonic anhydrase: Mechanism, structure and importance in higher plants. Asian J. Plant Sci. Res. 2017, 7, 17–23. [Google Scholar]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond Glycolysis: GAPDHs Are Multi-functional Enzymes Involved in Regulation of ROS, Autophagy, and Plant Immune Responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [Green Version]
- Islam, T.; Manna, M.; Kaul, T.; Pandey, S.; Reddy, C.S.; Reddy, M.K. Genome-Wide Dissection of Arabidopsis and Rice for the Identification and Expression Analysis of Glutathione Peroxidases Reveals Their Stress-Specific and Overlapping Response Patterns. Plant Mol. Biol. Rep. 2015, 33, 1413–1427. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Z.; Ding, A.; Kong, Y. Genome-Wide Identification and Expression Profiling Analysis of the Xyloglucan Endotransglucosylase/Hydrolase Gene Family in Tobacco (Nicotiana tabacum L.). Genes 2018, 9, 273. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Ren, L.; Wang, M.; Wang, Z.; Fu, N.; Wang, H.; Wang, X.; Ao, T.; Du, W.; Zheng, Z.; et al. Proteo-Transcriptomic Characterization of Sirex nitobei (Hymenoptera: Siricidae) Venom. Toxins 2021, 13, 562. [Google Scholar] [CrossRef]
- Daum, B.; Nicastro, D.; Austin, J.; McIntosh, J.R.; Kühlbrandt, W. Arrangement of Photosystem II and ATP Synthase in Chloroplast Membranes of Spinach and Pea. Plant Cell 2010, 22, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.P.; Boekema, E.J. Supramolecular organization of thylakoid membrane proteins in green plants. Biochim. Biophys. Acta 2005, 1706, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Green, B.R.; Durnford, D.G. The Chlorophyll-Carotenoid Proteins of Oxygenic Photosynthesis. Annu. Rev. Plant Biol. 1996, 47, 685–714. [Google Scholar] [CrossRef]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Caffarri, S.; Frigerio, S.; Olivieri, E.; Righetti, P.G.; Bassi, R. Differential accumulation of Lhcb gene products in thylakoid membranes of Zea mays plants grown under contrasting light and temperature conditions. Proteomics 2005, 5, 758–768. [Google Scholar] [CrossRef]
- Ganeteg, U.; Kulheim, C.; Andersson, J.; Jansson, S. Is Each Light-Harvesting Complex Protein Important for Plant Fitness? Plant Physiol. 2004, 134, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Humbeck, K.; Krupinska, K. The abundance of minor chlorophyll a/b-binding proteins CP29 and LHCI of barley (Hordeum vulgare L.) during leaf senescence is controlled by light. J. Exp. Bot. 2003, 54, 375–383. [Google Scholar] [CrossRef]
- Staneloni, R.J.; Rodriguez-Batiller, M.J.; Casal, J.J. Abscisic Acid, High-Light, and Oxidative Stress Down-Regulate a Photosynthetic Gene via a Promoter Motif Not Involved in Phytochrome-Mediated Transcriptional Regulation. Mol. Plant 2008, 1, 75–83. [Google Scholar] [CrossRef]
- Jung, C.; Müller, A.E. Flowering time control and applications in plant breeding. Trends Plant Sci. 2009, 14, 563–573. [Google Scholar] [CrossRef]
- Kiranmai, K.; Gunupuru, L.R.; Nareshkumar, A.; Reddy, V.A.; Lokesh, U.; Pandurangaiah, M.; Venkatesh, B.; Kirankumar, T.V.; Sudhakar, C. Expression Analysis of WRKY Transcription Factor Genes in Response to Abiotic Stresses in Horsegram (Macrotyloma uniflorum (Lam.) Verdc.). Am. J. Mol. Biol. 2016, 6, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cheng, J.; Sun, X.; Zhao, T.; Li, M.; Wang, Q.; Li, S.; Xin, H. Overexpression of VaWRKY14 increases drought tolerance in Arabidopsis by modulating the expression of stress-related genes. Plant Cell Rep. 2018, 37, 1159–1172. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2016, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]
- Okamuro, J.K.; Caster, B.; Villarroel, R.; Van Montagu, M.; Jofuku, K.D. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 7076–7081. [Google Scholar] [CrossRef] [Green Version]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 1998, 379, 633–646. [Google Scholar] [CrossRef]
- Magnani, E.; Sjölander, K.; Hake, S. From Endonucleases to Transcription Factors: Evolution of the AP2 DNA Binding Domain in Plants[W]. Plant Cell 2004, 16, 2265–2277. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Meeley, R.; Hake, S. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development 2008, 135, 3013–3019. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, H.; Zhu, J.; Zhang, J.; Zhang, X.; Wang, F.; Tang, Y.; Mei, B.; Xu, Z.; Song, R. An expression analysis of 57 transcription factors derived from ESTs of developing seeds in Maize (Zea mays). Plant Cell Rep. 2010, 29, 545–559. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Front. Plant Sci. 2019, 10, 228. [Google Scholar]
- Hao, D.; Ohme-Takagi, M.; Sarai, A. Unique Mode of GCC Box Recognition by the DNA-binding Domain of Ethylene-responsive Element-binding Factor (ERF Domain) in Plant. J. Biol. Chem. 1998, 273, 26857–26861. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Ju, H.; Zhou, G.; Zhu, C.; Erb, M.; Wang, X.; Wang, P.; Lou, Y. An EAR-motif-containing ERF transcription factor affects herbivore-induced signaling, defense and resistance in rice. Plant J. 2011, 68, 583–596. [Google Scholar] [CrossRef]
- Khare, D.; Mitsuda, N.; Lee, S.; Song, W.; Hwang, D.; Ohme-Takagi, M.; Martinoia, E.; Lee, Y.; Hwang, J. Root avoidance of toxic metals requires the GeBP-LIKE 4 transcription factor in Arabidopsis thaliana. New Phytol. 2016, 213, 1257–1273. [Google Scholar] [CrossRef] [Green Version]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Number |
|---|---|
| Subread Reads | 6,588,055 |
| Average Subread Length | 3625 |
| Subread N50 | 4581 |
| Total CCS | 199,841 |
| Average CCS Length (bp) | 4250 |
| FL Reads | 163,130 |
| FL Mean Length (bp) | 4232 |
| FLNC Reads | 162,036 |
| FLNC Mean Length (bp) | 4184 |
| Total Unigenes | 52,963 |
| Average Unigene Length (bp) | 1801 |
| TF number | 1017 |
| TF Family | Number of TFs | ||
|---|---|---|---|
| Inoculation-Specific | Wounding-Specific | Co-Induced | |
| AP2/ERF | 46 | 4 | 4 |
| LOB | 1 | 1 | 1 |
| MYB_superfamily | 1 | 3 | 1 |
| C2H2 | 1 | N/A | N/A |
| C3H | 1 | N/A | N/A |
| GeBP | 1 | N/A | N/A |
| GRAS | 1 | N/A | N/A |
| NAC | N/A | 1 | N/A |
| Gene_ID | Swiss-Prot | Log2FC(PI_72 h/CK_72 h) | p-Value |
|---|---|---|---|
| transcript_26258 | Probable disease-resistance protein At1g15890 Arabidopsis thaliana | 1.84 | 2.31 × 10−6 |
| transcript_64802 | Calmodulin-like protein 3 Arabidopsis thaliana | 4.40 | 3.95 × 10−7 |
| transcript_66692 | Putative calmodulin-3 (Fragment) Solanum tuberosum | 4.78 | 2.69 × 10−8 |
| transcript_11085 | Calmodulin-like protein 2 Arabidopsis thaliana | 4.36 | 1.26 × 10−15 |
| transcript_52365 | LRR receptor-like serine/threonine-protein kinase FLS2 Arabidopsis thaliana | 4.24 | 5.83 × 10−5 |
| transcript_35906 | Heat-shock protein 83 Ipomoea nil | 2.25 | 2.47 × 10−5 |
| transcript_25250 | Heat-shock protein 83 Ipomoea nil | 4.02 | 3.52 × 10-4 |
| Annotation | Gene_ID | Log2FC(PI_72h/CK_72h) |
|---|---|---|
| GAPDH | transcript_53182 | 7.7805 |
| transcript_44911 | 4.5701 | |
| transcript_1666 | 3.1995 | |
| GPX | transcript_43923 | 2.1892 |
| transcript_54716 | 1.5788 | |
| CAT | transcript_40212 | 3.5800 |
| XTH2 | transcript_48021 | 4.9165 |
| transcript_59698 | 3.7113 | |
| transcript_7449 | 3.3294 | |
| transcript_52674 | 2.7133 | |
| transcript_7326 | 2.5641 | |
| UGT | transcript_70636 | 2.6561 |
| transcript_40291 | 1.7841 | |
| transcript_41406 | 1.6345 | |
| putative truncated TIR-NBS-LRR protein | transcript_20771 | 2.7826 |
| transcript_19948 | 2.1045 | |
| transcript_19947 | 1.9758 | |
| transcript_21018 | 1.7106 | |
| transcript_20174 | 1.5549 | |
| polyubiquitin | transcript_65149 | 9.5582 |
| transcript_55582 | 2.1239 | |
| transcript_5903 | −1.554 | |
| transcript_6253 | 7.3522 | |
| PDR protein 1-like | transcript_37577 | 6.2179 |
| Hs1pro-1 | transcript_23455 | 2.1044 |
| transcript_65743 | 2.1014 |
| Annotation | Gene_ID | Log2FC(PW_72 h/CK_72 h) |
|---|---|---|
| Lhcb5 protein | transcript_58449 | 10.3402 |
| transcript_61176 | 9.1811 | |
| transcript_71238 | 5.2818 | |
| transcript_8414 | 2.5689 | |
| transcript_67898 | −7.5324 | |
| transcript_15721 | −9.1712 | |
| LHC protein, partial | transcript_73156 | 7.4302 |
| transcript_67949 | −2.3693 | |
| transcript_51999 | −2.3040 | |
| transcript_47491 | −2.4656 | |
| OEE | transcript_7777 | −4.6929 |
| transcript_5485 | −2.0221 | |
| Lhca4 protein | transcript_49364 | −2.3063 |
| HD16 | transcript_28611 | −7.1177 |
| transcript_31667 | −9.0054 | |
| transcript_17461 | −9.0169 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, C.; Ren, L.; Wang, M.; Wang, Z.; Fu, N.; Wang, H.; Shi, J. Full-Length Transcriptome Sequencing-Based Analysis of Pinus sylvestris var. mongolica in Response to Sirex noctilio Venom. Insects 2022, 13, 338. https://doi.org/10.3390/insects13040338
Gao C, Ren L, Wang M, Wang Z, Fu N, Wang H, Shi J. Full-Length Transcriptome Sequencing-Based Analysis of Pinus sylvestris var. mongolica in Response to Sirex noctilio Venom. Insects. 2022; 13(4):338. https://doi.org/10.3390/insects13040338
Chicago/Turabian StyleGao, Chenglong, Lili Ren, Ming Wang, Zhengtong Wang, Ningning Fu, Huiying Wang, and Juan Shi. 2022. "Full-Length Transcriptome Sequencing-Based Analysis of Pinus sylvestris var. mongolica in Response to Sirex noctilio Venom" Insects 13, no. 4: 338. https://doi.org/10.3390/insects13040338
APA StyleGao, C., Ren, L., Wang, M., Wang, Z., Fu, N., Wang, H., & Shi, J. (2022). Full-Length Transcriptome Sequencing-Based Analysis of Pinus sylvestris var. mongolica in Response to Sirex noctilio Venom. Insects, 13(4), 338. https://doi.org/10.3390/insects13040338