
Lactobacillus paracasei CCFM1229 and Lactobacillus rhamnosus CCFM1228 Alleviated Depression- and Anxiety-Related Symptoms of Chronic Stress-Induced Depression in Mice by Regulating Xanthine Oxidase Activity in the Brain

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Probiotic Strains and Culture Conditions
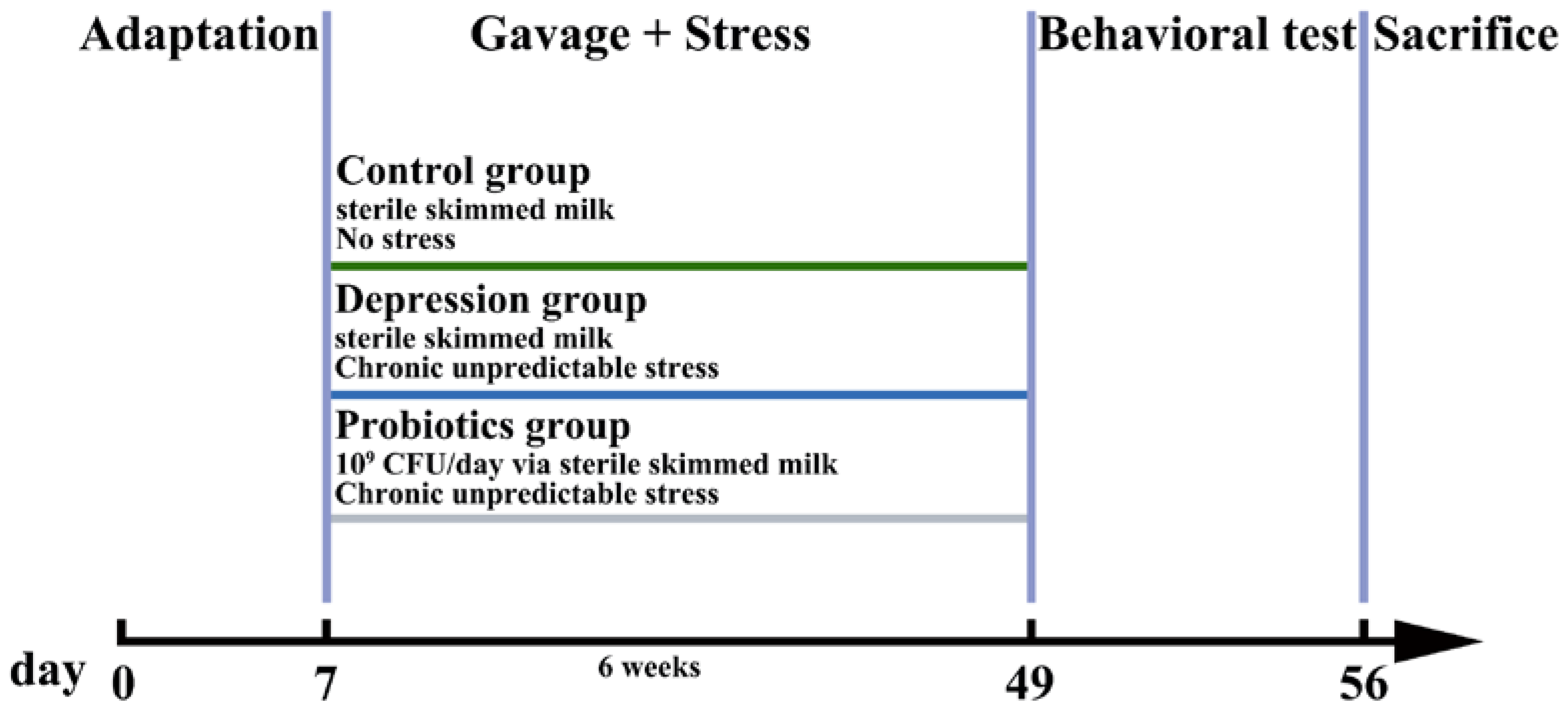
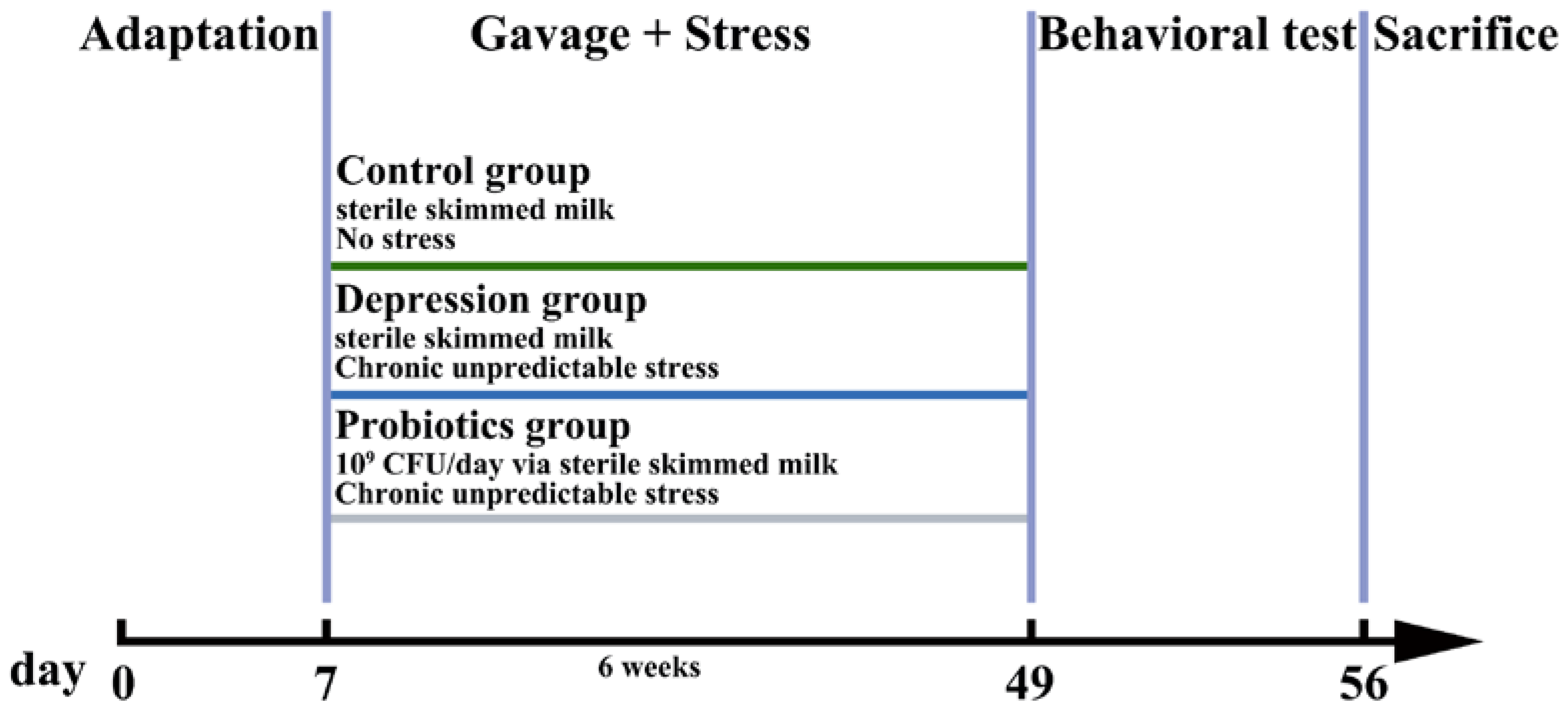
2.2. Animals and Treatment
2.3. Behavioural Tests
2.4. Determination of Neurobiological Factors
2.5. Determination of Neuroregulatory Factors
2.6. Gut Microbiota Analysis
2.7. Determination of Purine Metabolism Indexes
2.8. Statistical Analysis
3. Results
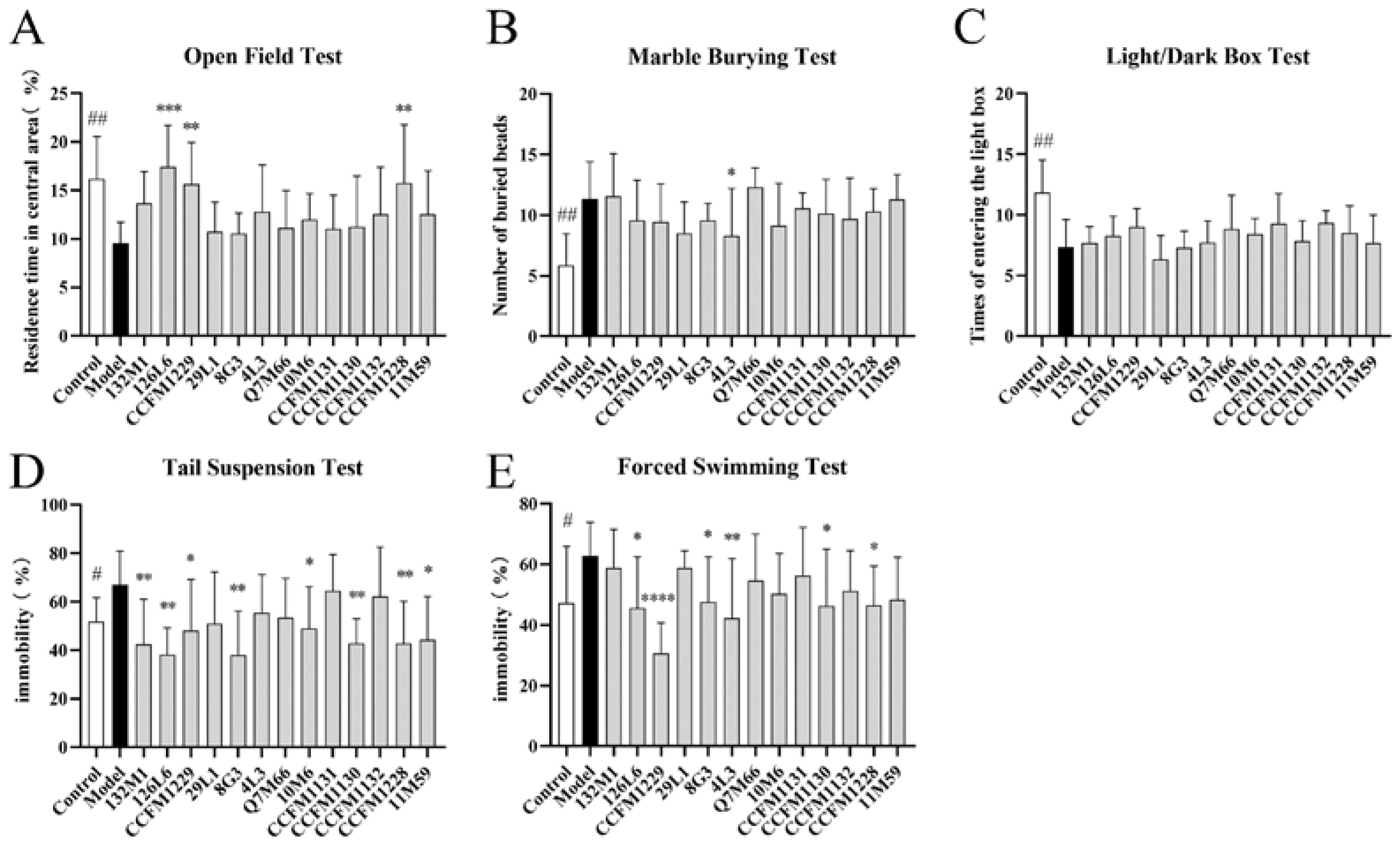
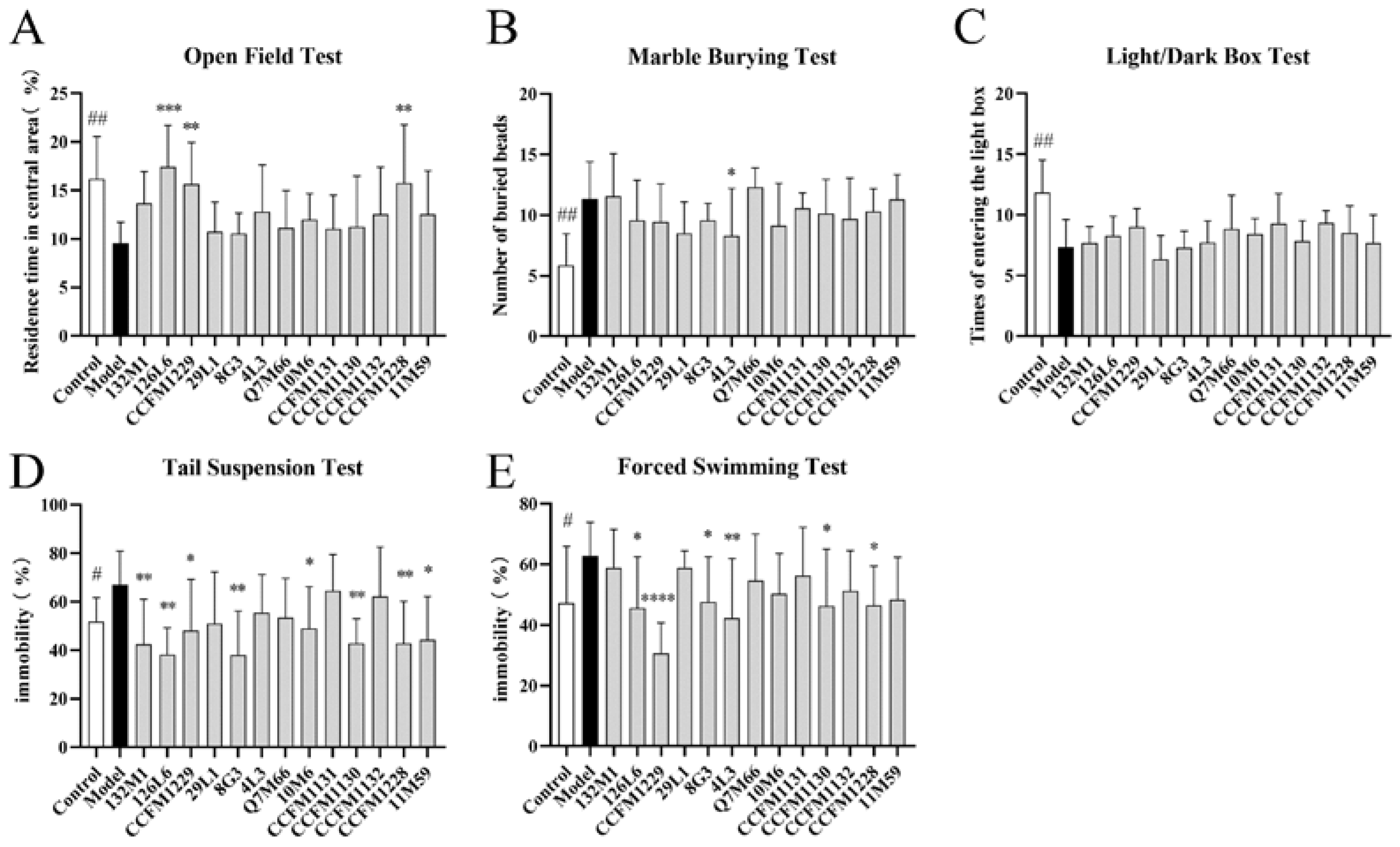
3.1. Some Lactobacillus Strains Can Alleviate Depressive Behaviour in Mice
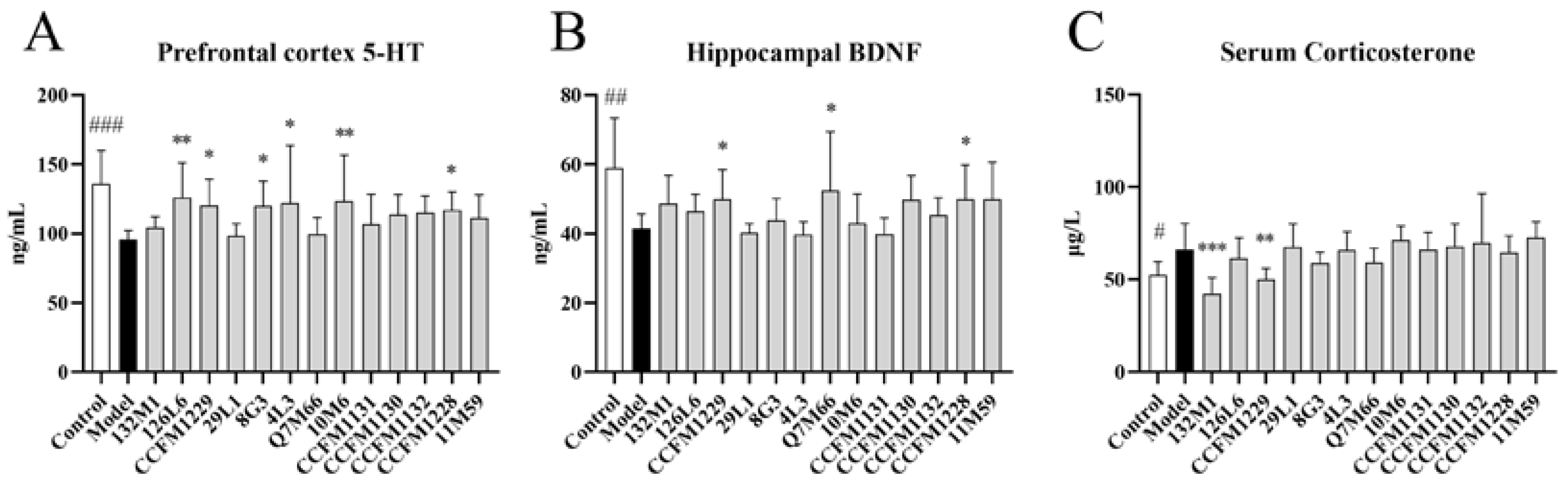
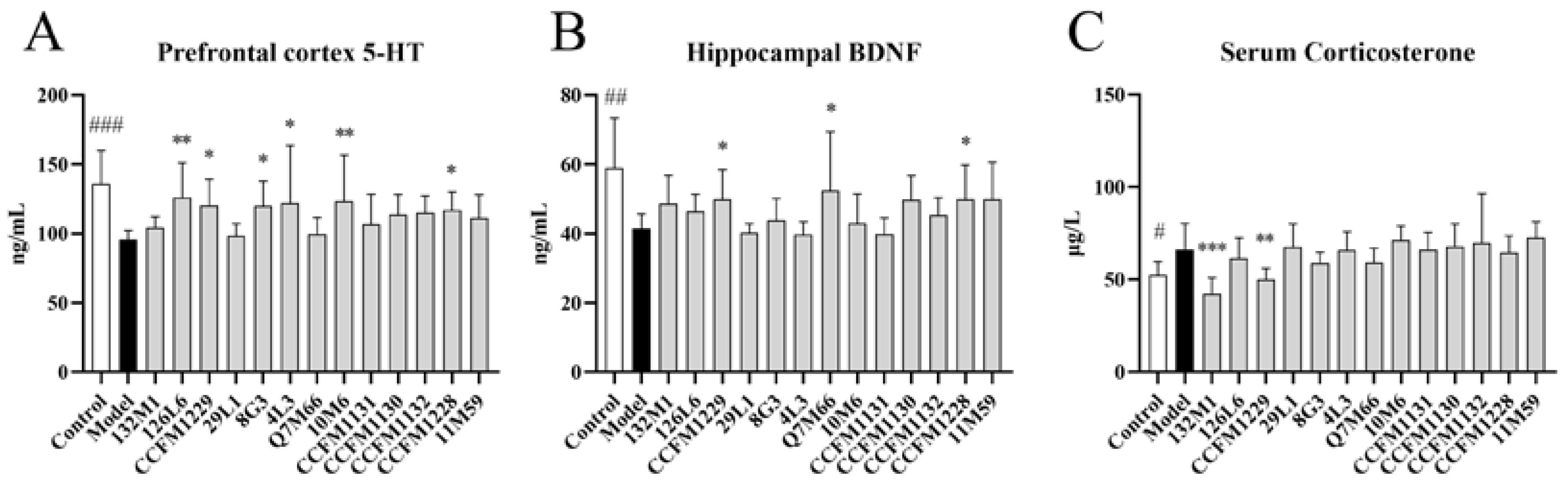
3.2. The Effects of Lactobacillus on Neurobiology
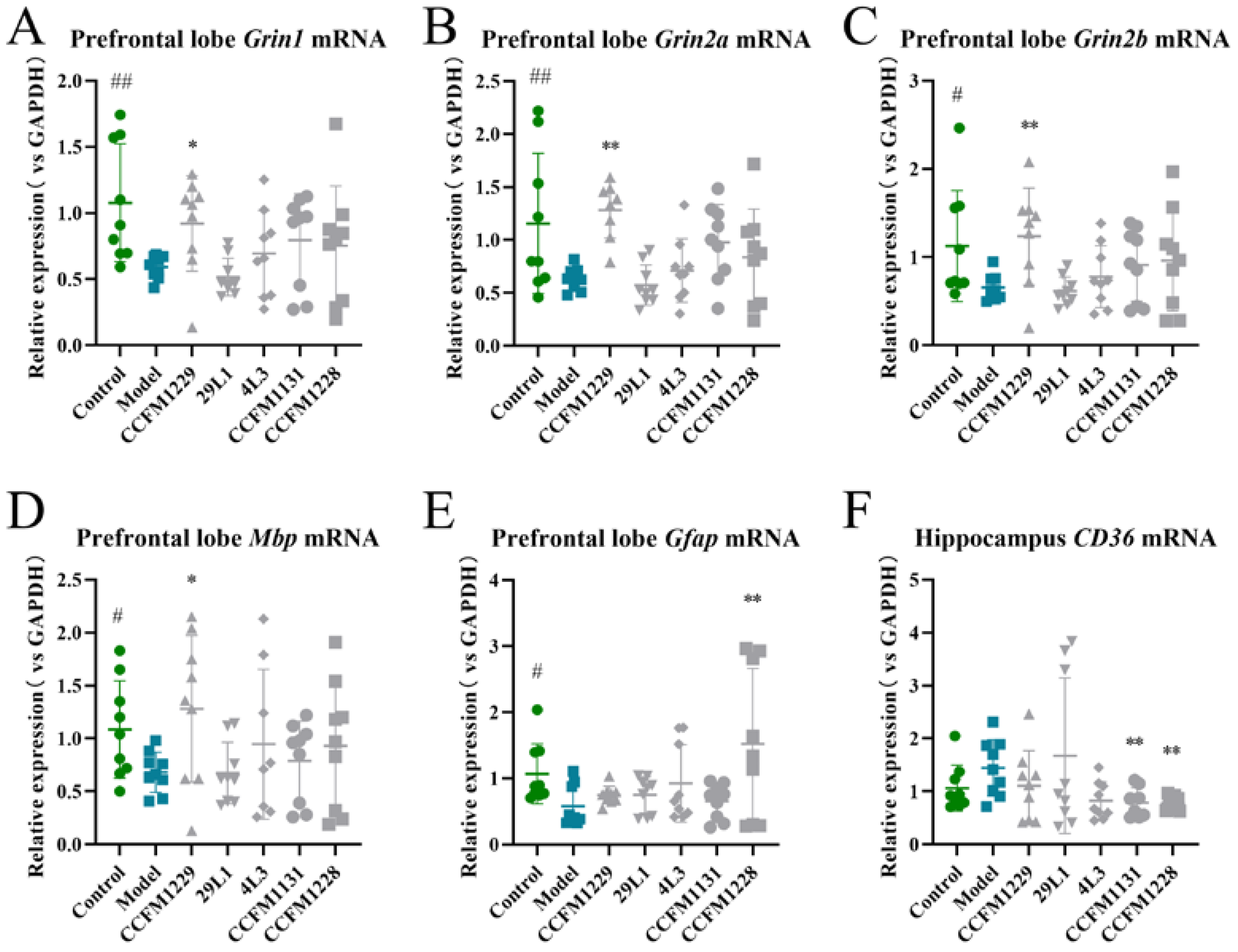
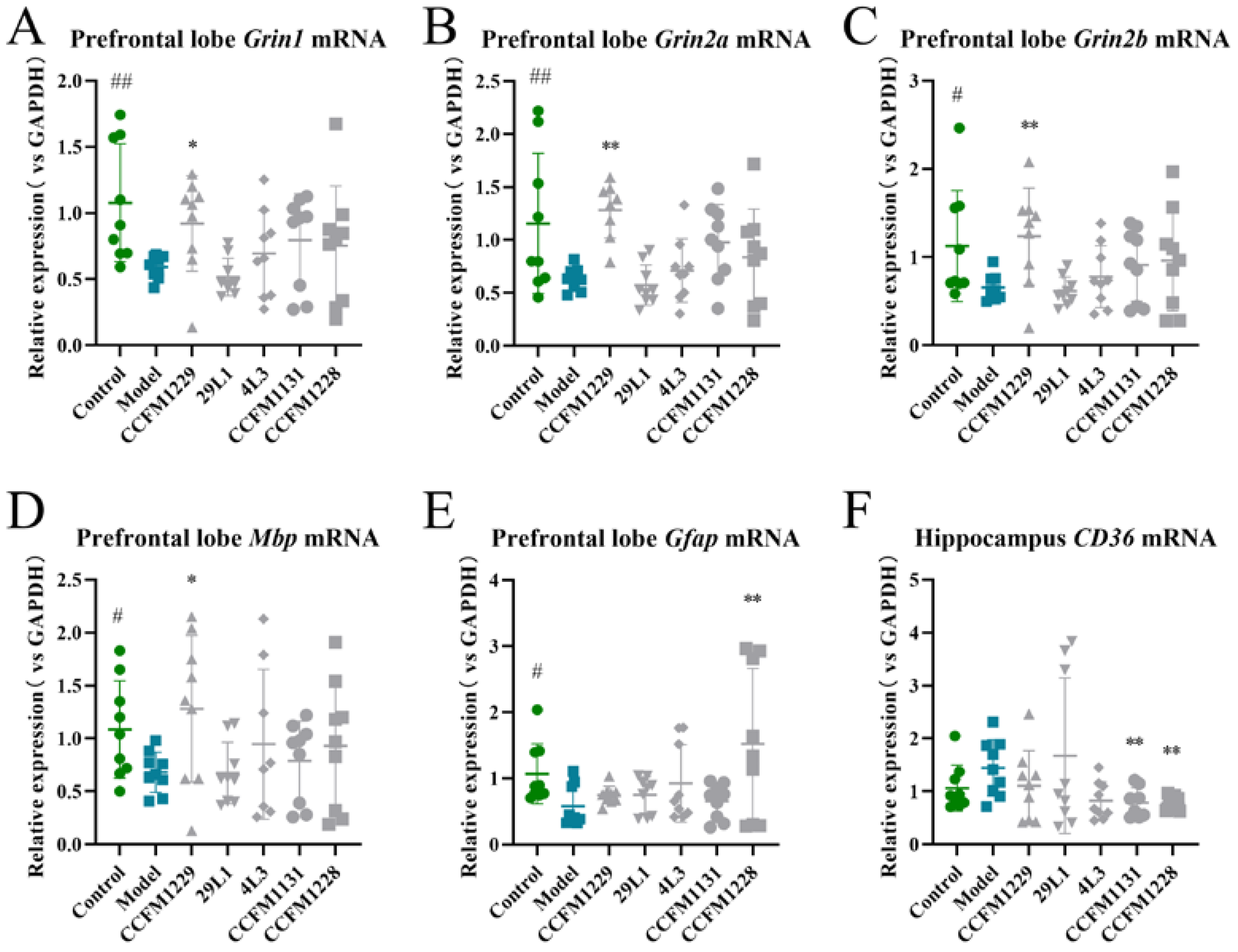
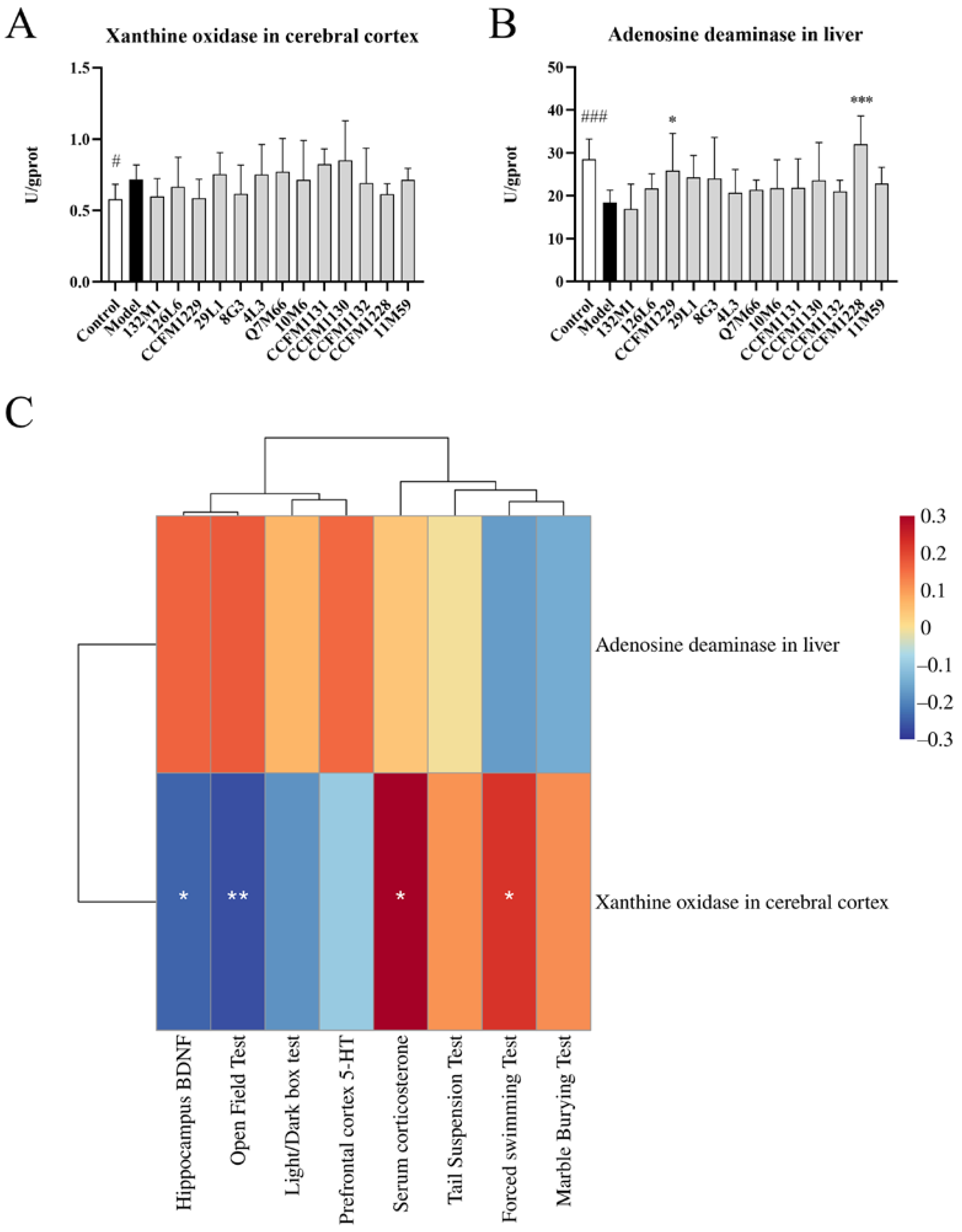
3.3. Unique Neuroregulatory Effects of Lactobacillus paracasei CCFM1229 and Lactobacillus rhamnosus CCFM1228
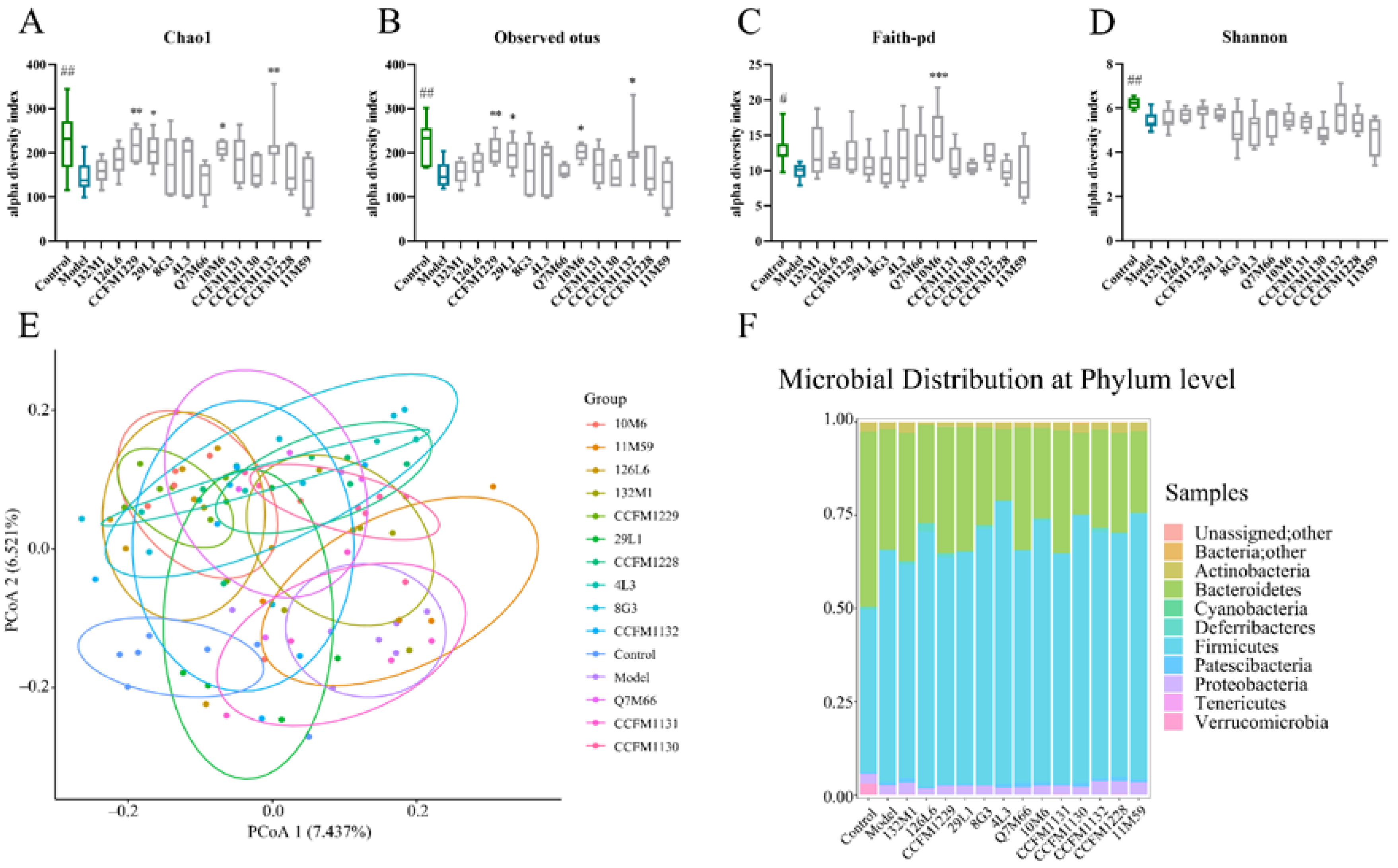
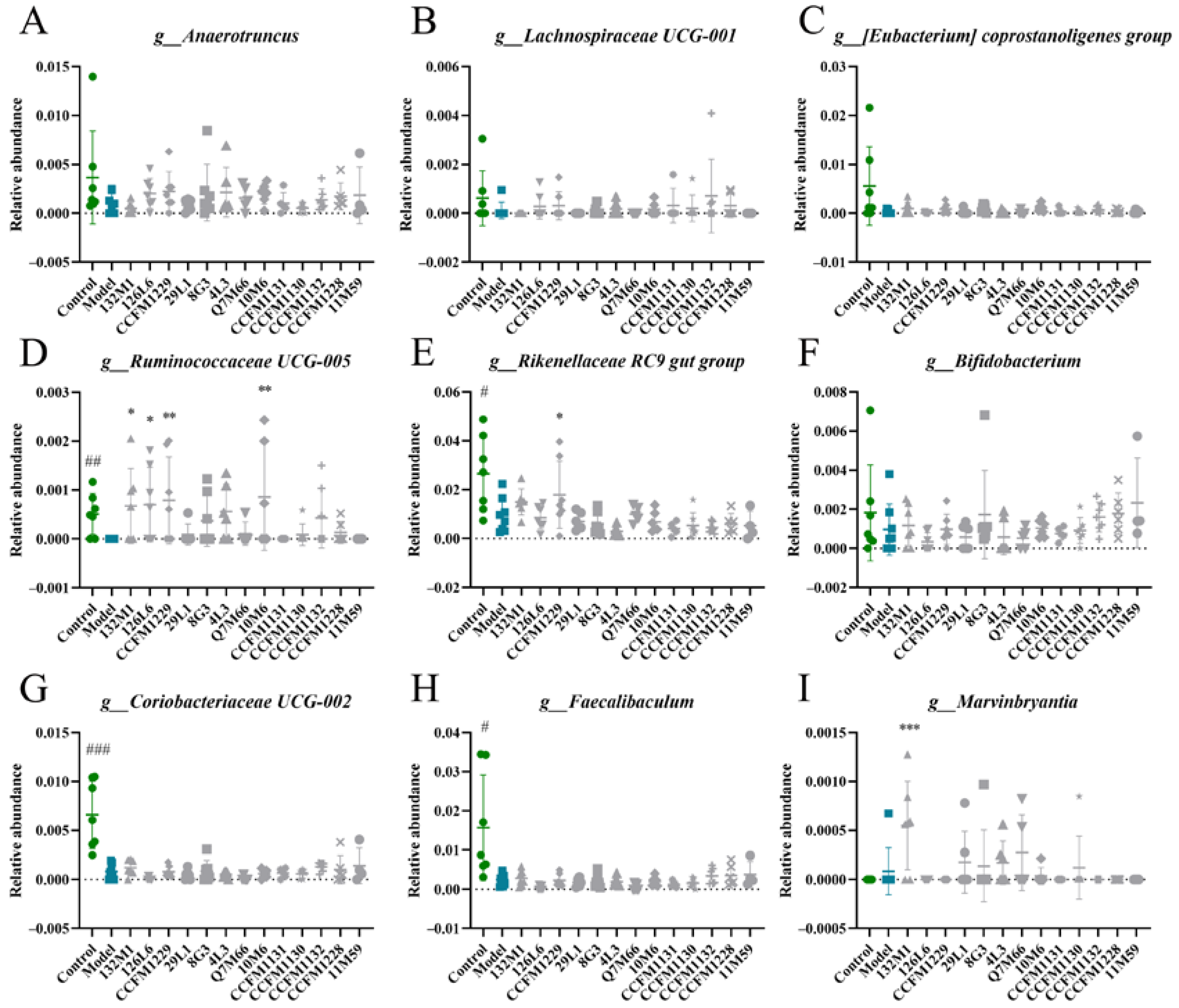
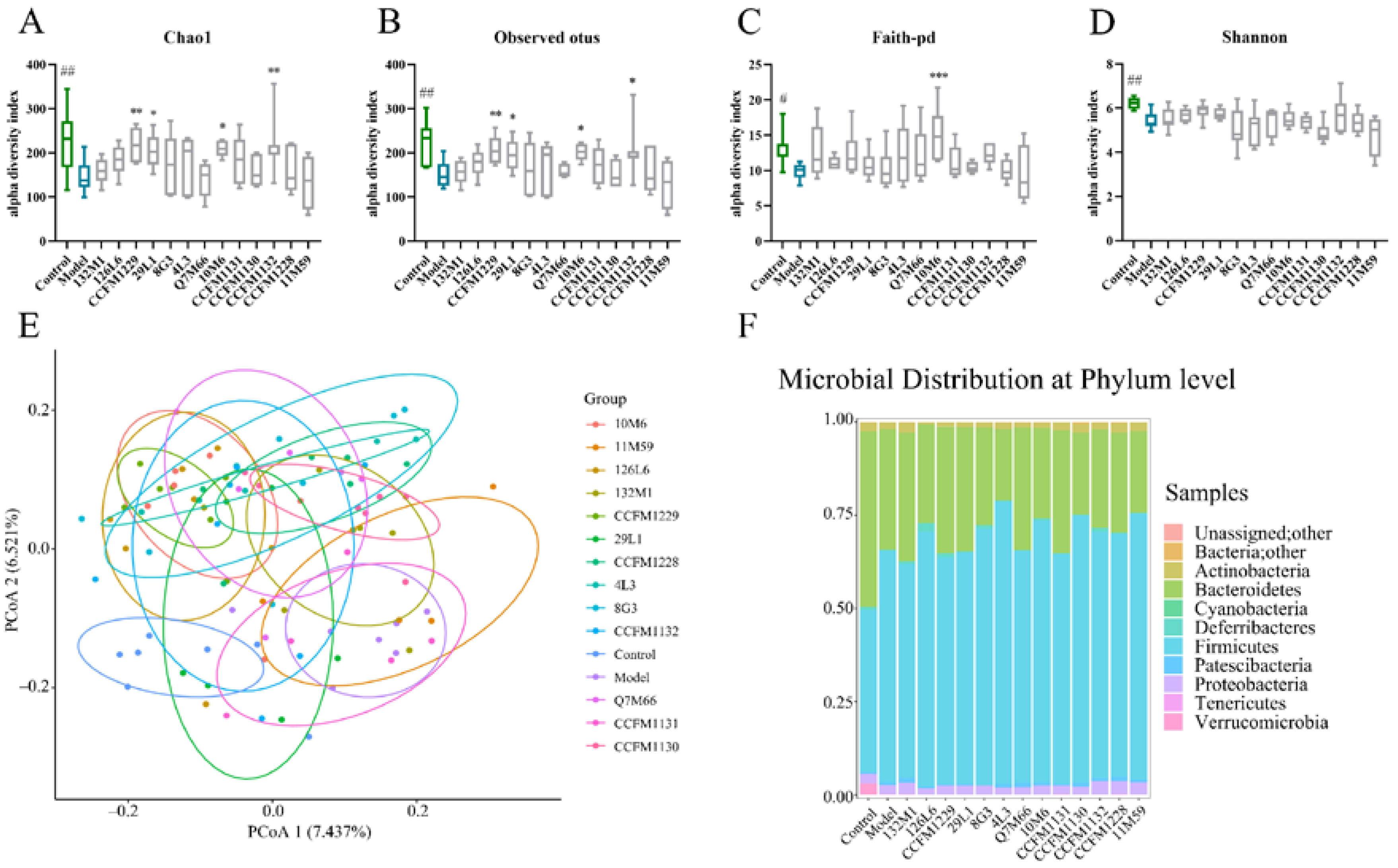
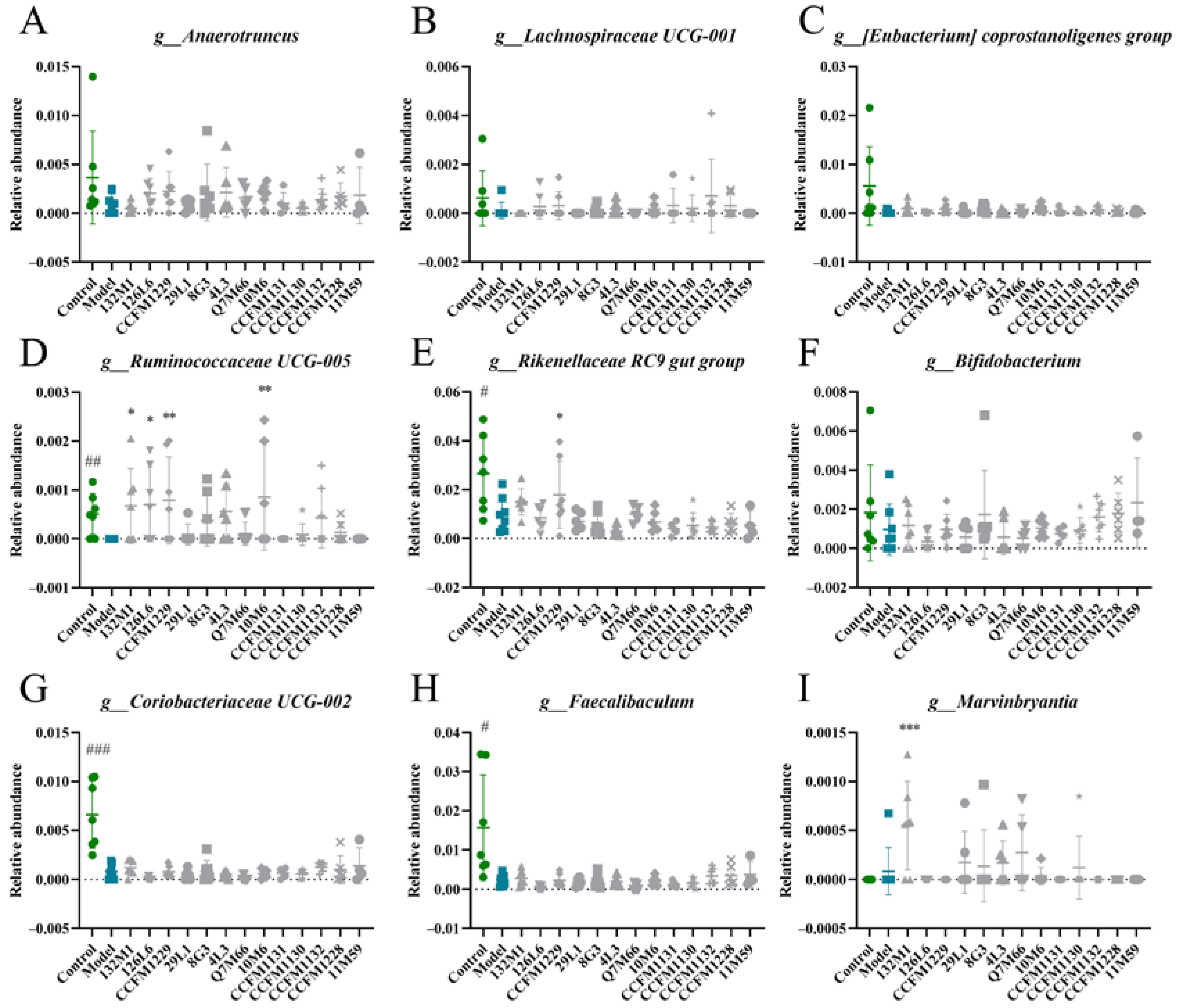
3.4. Effects of Lactobacillus on Microbiome Disorders Caused by Chronic Stress
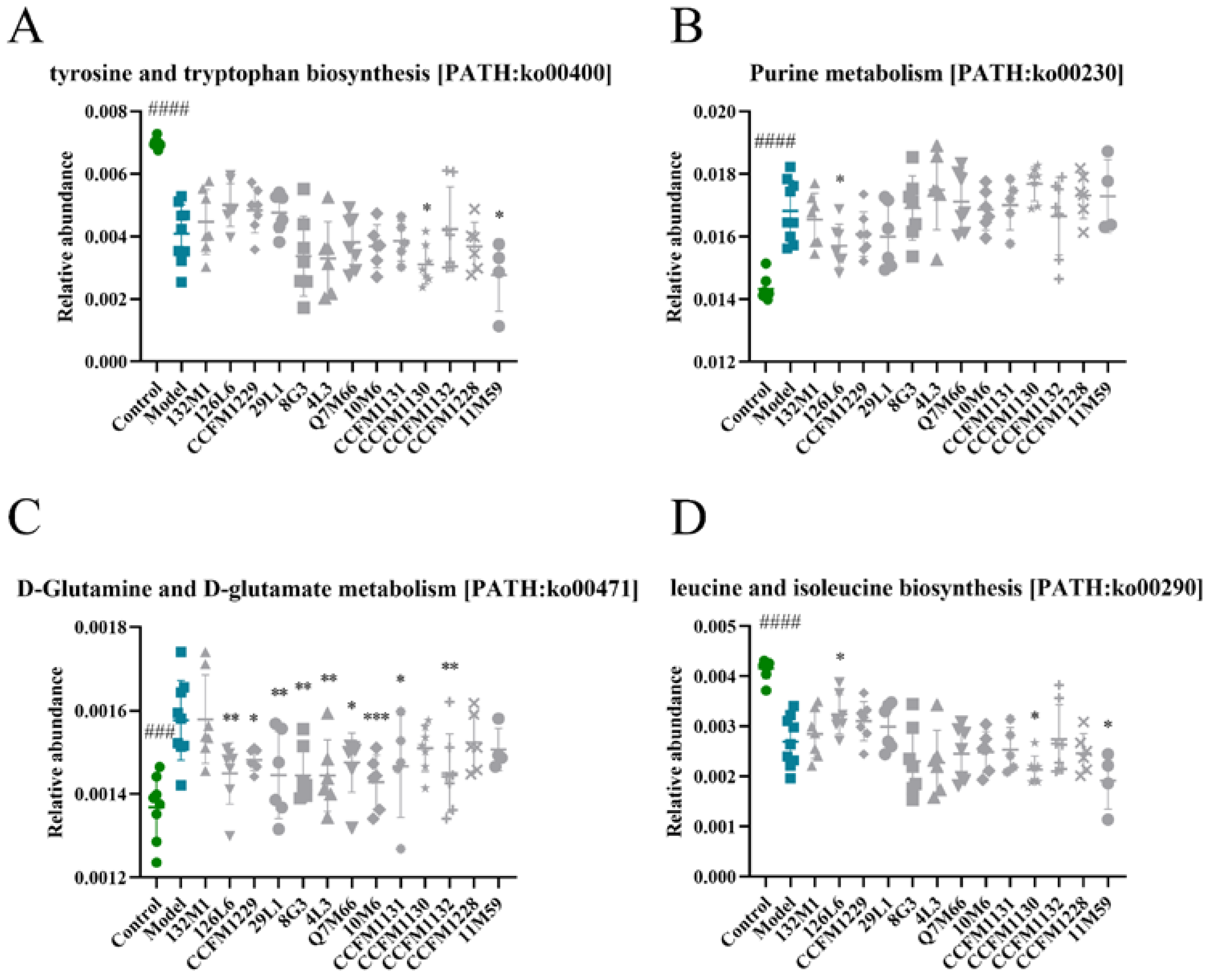
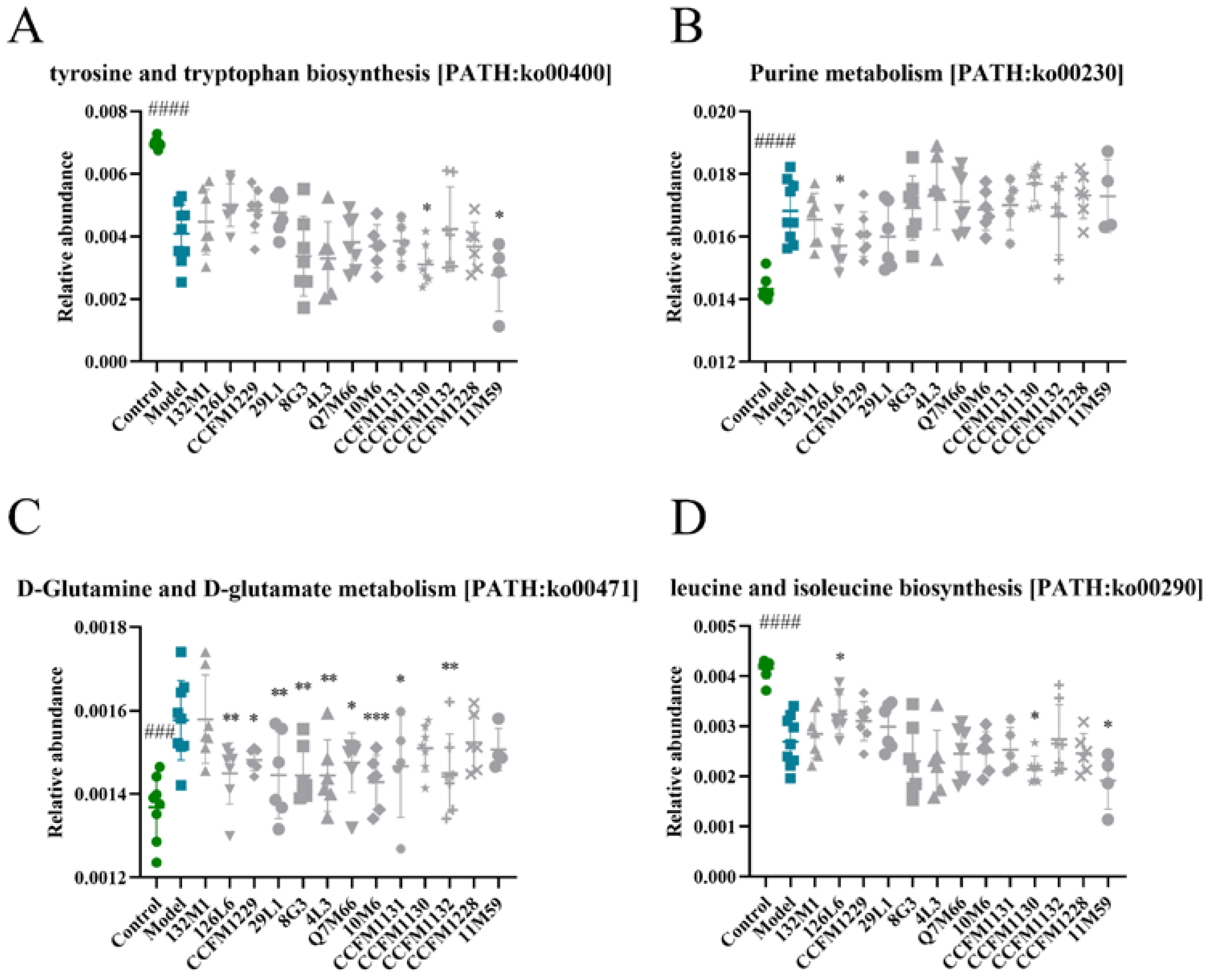
3.5. Effect of Lactobacillus on Purine Metabolism
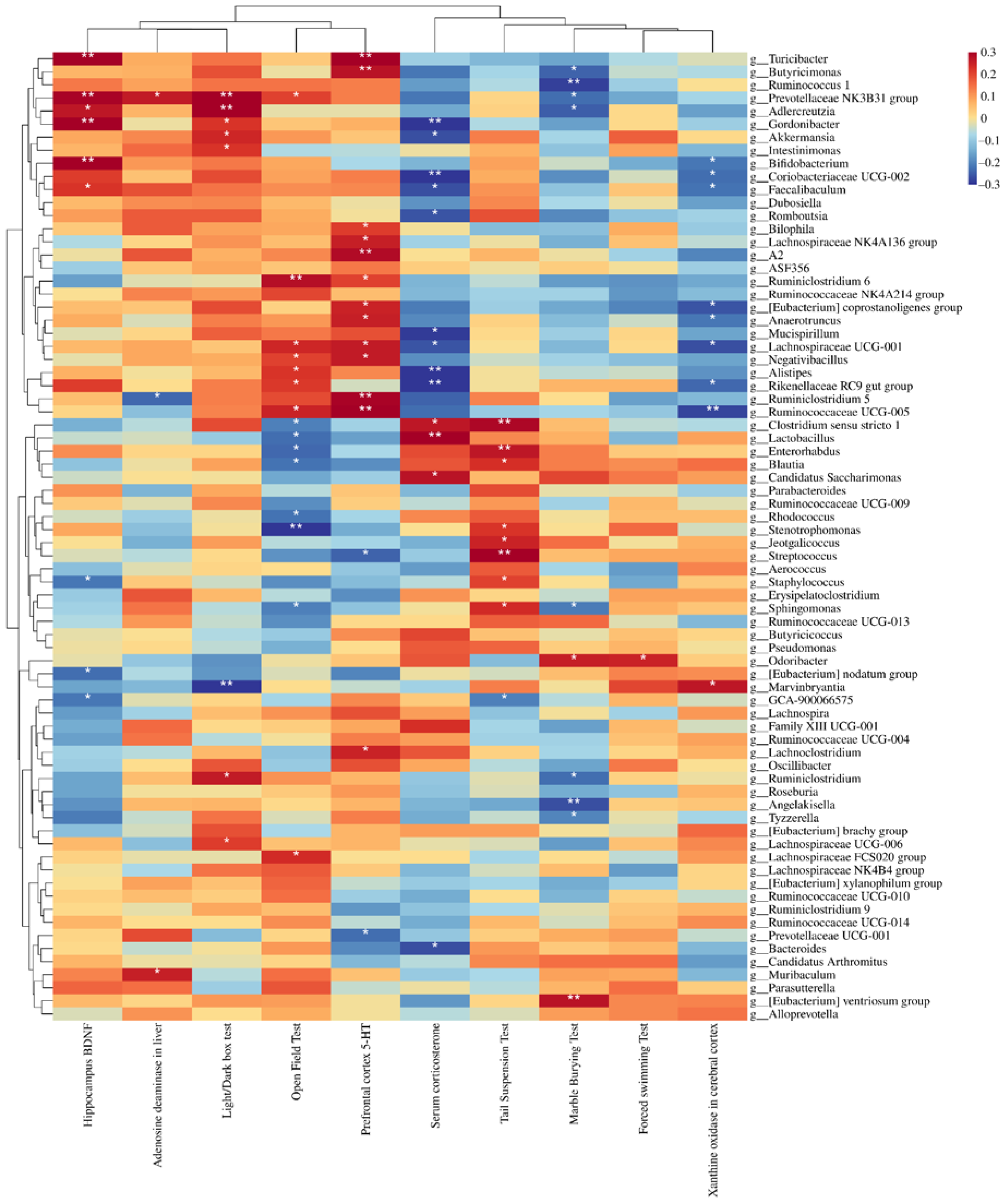
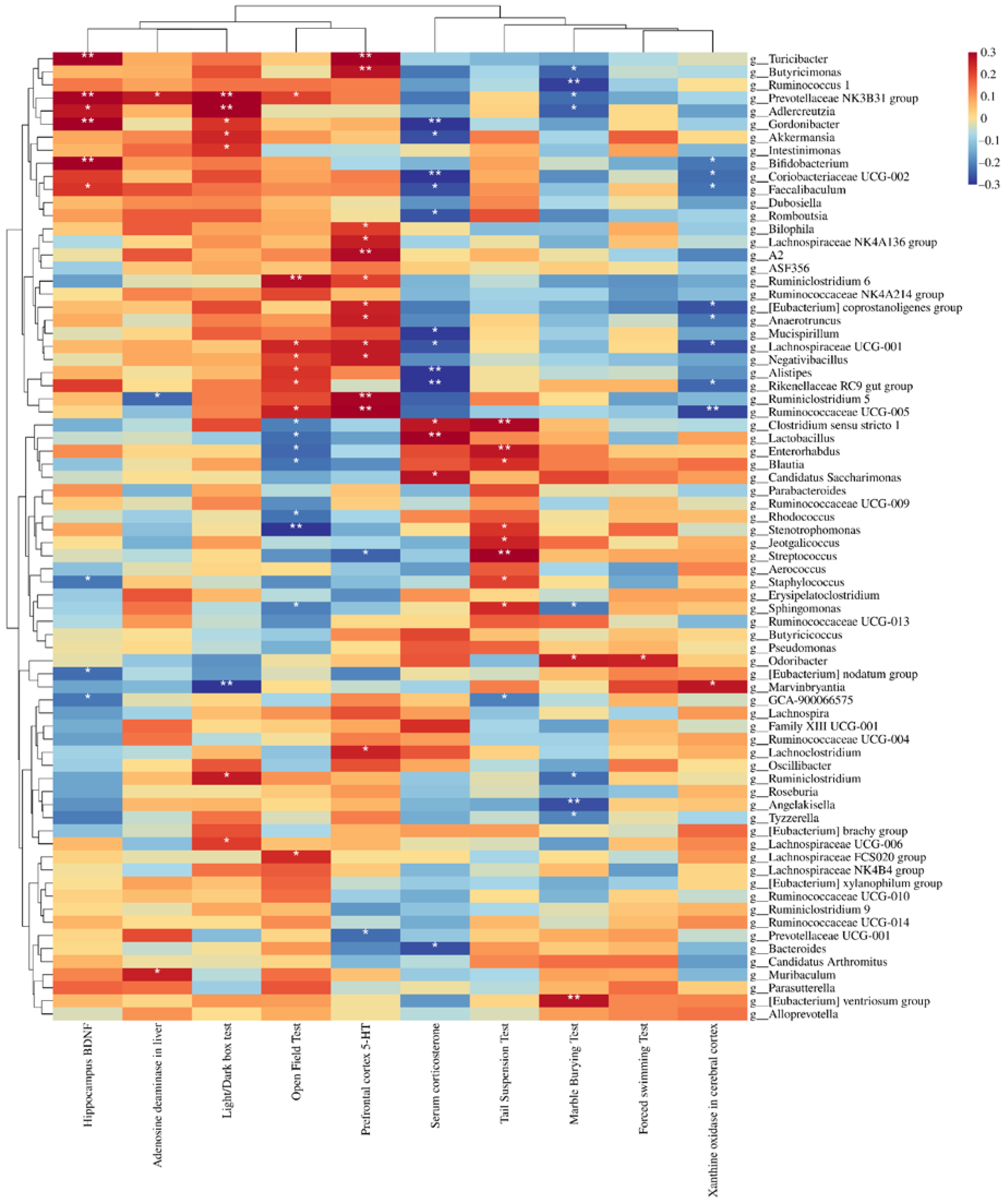
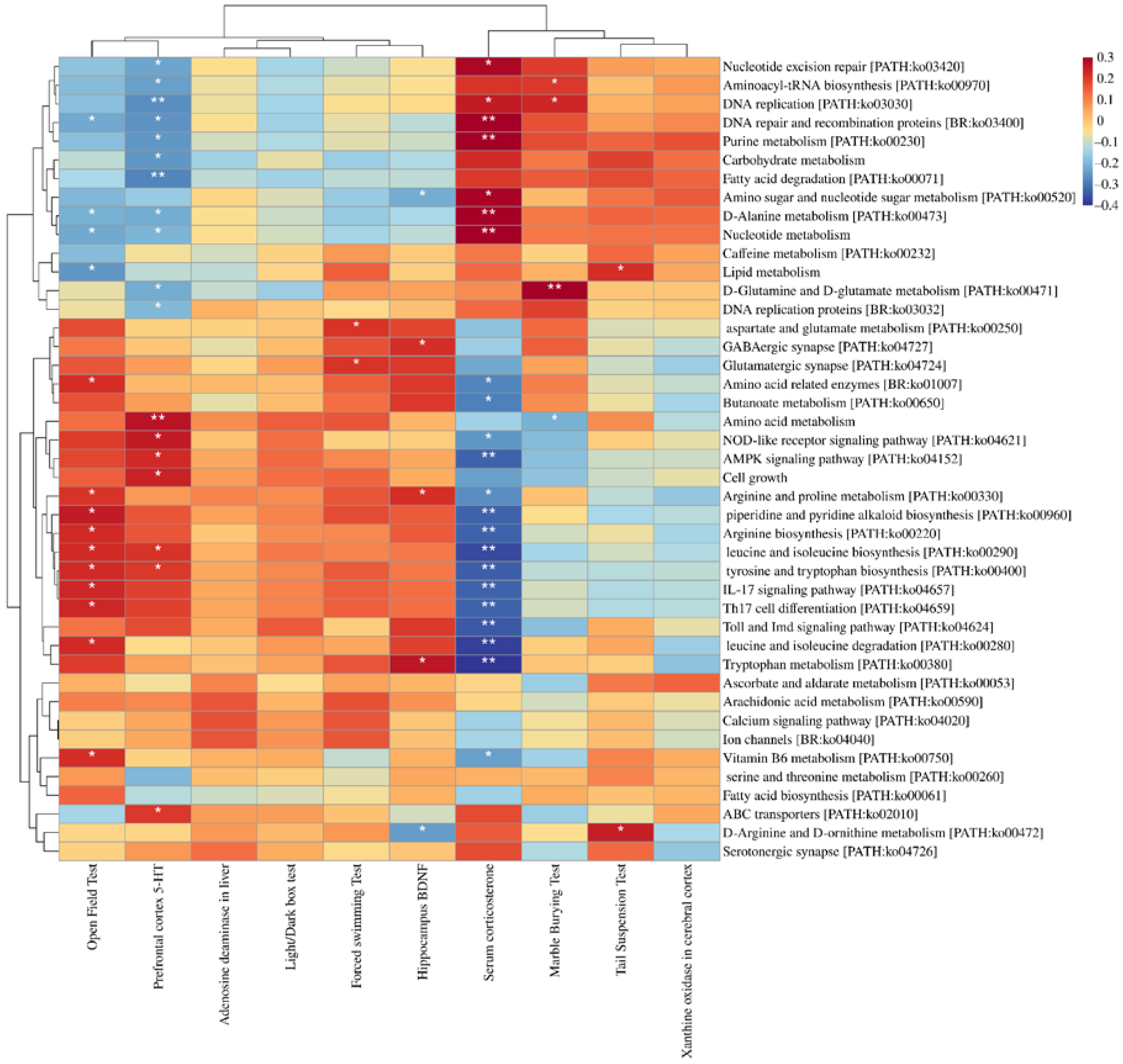
3.6. Depression Indices Were Closely Related to Changes in the Microflora and KEGG Functionality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moussavi, S.; Chatterji, S.; Verdes, E.; Tandon, A.; Patel, V.; Ustun, B. Depression, chronic diseases, and decrements in health: Results from the World Health Surveys. Lancet 2007, 370, 851–858. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Logan, A.C.; Katzman, M. Major depressive disorder: Probiotics may be an adjuvant therapy. Med. Hypotheses 2005, 64, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Bjorkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, K.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol. Motil. 2011, 23, 255-e119. [Google Scholar] [CrossRef]
- Tian, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J. Nutr. Biochem. 2019, 66, 43–51. [Google Scholar] [CrossRef]
- Wei, C.-L.; Wang, S.; Yen, J.-T.; Cheng, Y.-F.; Liao, C.-L.; Hsu, C.-C.; Wu, C.-C.; Tsai, Y.-C. Antidepressant-like activities of live and heat-killed Lactobacillus paracasei PS23 in chronic corticosterone-treated mice and possible mechanisms. Brain Res. 2019, 1711, 202–213. [Google Scholar] [CrossRef]
- Liu, Y.-W.; Ong, J.S.; Gan, C.Y.; Khoo, B.Y.; Yahaya, S.; Choi, S.B.; Low, W.Y.; Tsai, Y.-C.; Liong, M.T. Lactobacillus fermentum PS150 showed psychotropic properties by altering serotonergic pathway during stress. J. Funct. Foods 2019, 59, 352–361. [Google Scholar] [CrossRef]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsythe, P.; Sudo, N.; Dinan, T.; Taylor, V.H.; Bienenstock, J. Mood and gut feelings. Brain Behav. Immun. 2010, 24, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Wang, W.; Wang, T.; Li, J.; Zhang, S.; Chen, Z.; Qi, X.; Chen, J.; Cheng, K.; Xie, P. CD36 deficiency affects depressive-like behaviors possibly by modifying gut microbiota and the inflammasome pathway in mice. Transl. Psychiatry 2021, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M. Glucocorticoids and Hippocampal Atrophy in Neuropsychiatric Disorders. Arch. Gen. Psychiatry 2000, 57, 925–935. [Google Scholar] [CrossRef]
- Lau, C.G.; Zukin, R.S. NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat. Rev. Neurosci. 2007, 8, 413–426. [Google Scholar] [CrossRef]
- Boda, E. Myelin and oligodendrocyte lineage cell dysfunctions: New players in the etiology and treatment of depression and stress-related disorders. Eur. J. Neurosci. 2021, 53, 281–297. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. Synaptic and behavioral effects of chronic stress are linked to dynamic and sex-specific changes in microglia function and astrocyte dystrophy. Neurobiol. Stress 2021, 14, 15. [Google Scholar] [CrossRef]
- Ali-Sisto, T.; Tolmunen, T.; Toffol, E.; Viinamäki, H.; Mäntyselkä, P.; Valkonen-Korhonen, M.; Honkalampi, K.; Ruusunen, A.; Velagapudi, V.; Lehto, S.M. Purine metabolism is dysregulated in patients with major depressive disorder. Psychoneuroendocrinology 2016, 70, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Kaddurah-Daouk, R.; Yuan, P.; Boyle, S.H.; Matson, W.; Wang, Z.; Zeng, Z.B.; Zhu, H.; Dougherty, G.G.; Yao, J.K.; Chen, G. Cerebrospinal fluid metabolome in mood disorders-remission state has a unique metabolic profile. Sci. Rep. 2012, 2, 667. [Google Scholar] [CrossRef] [PubMed]
- Niklasson, F.; Agren, H.; Hällgren, R. Purine and monoamine metabolites in cerebrospinal fluid: Parallel purinergic and monoaminergic activation in depressive illness? J. Neurol. Neurosurg. Psychiatry 1983, 46, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.Q.; Li, Y.Y.; Wang, H.L.; Mao, X.T.; Guo, J.X.; Wang, F.; Huang, L.J.; Li, Y.N.; Ma, X.Y.; Gao, Z.J.; et al. Stress-Induced Metabolic Disorder in Peripheral CD4(+) T Cells Leads to Anxiety-like Behavior. Cell 2019, 179, 864–879.e819. [Google Scholar] [CrossRef]
- Michel, T.M.; Camara, S.; Tatschner, T.; Frangou, S.; Sheldrick, A.J.; Riederer, P.; Grünblatt, E. Increased xanthine oxidase in the thalamus and putamen in depression. World J. Biol. Psychiatry 2010, 11, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Fatokun, A.A.; Stone, T.W.; Smith, R.A. Hydrogen peroxide mediates damage by xanthine and xanthine oxidase in cerebellar granule neuronal cultures. Neurosci. Lett. 2007, 416, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini-Giampietro, D.E.; Cherici, G.; Alesiani, M.; Carlà, V.; Moroni, F. Excitatory amino acid release from rat hippocampal slices as a consequence of free-radical formation. J. Neurochem. 1988, 51, 1960–1963. [Google Scholar] [CrossRef] [PubMed]
- Colton, C.; Yao, J.; Grossman, Y.; Gilbert, D. The effect of xanthine/xanthine oxidase generated reactive oxygen species on synaptic transmission. Free Radic. Res. Commun. 1991, 14, 385–393. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, H.C.; He, S.B.; Zhang, X.F.; Ling, Y.H.; Li, X.Y.; Zhang, H.; Hou, D.D. The immunoenhancement effects of sea buckthorn pulp oil in cyclophosphamide-induced immunosuppressed mice. Food Funct. 2021, 12, 7954–7963. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Mu, T.Q.; Jia, H.; Yang, Y.; Wu, Z.L. Protective effects of glycine against lipopolysaccharide-induced intestinal apoptosis and inflammation. Amino Acids 2021. [Google Scholar] [CrossRef]
- Yen-Chun, K.; Pei-Sheng, L.; Yu-Lun, K.; Kalyanam, N.; Chi-Tang, H.; Min-Hsiung, P. Dietary pterostilbene and resveratrol modulate the gut microbiota influenced by circadian rhythm dysregulation. Mol. Nutr. Food Res. 2021, 65, 2100434. [Google Scholar] [CrossRef]
- Guo, M.; Li, Z.Y. Polysaccharides isolated from Nostoc commune Vaucher inhibit colitis-associated colon tumorigenesis in mice and modulate gut microbiota. Food Funct. 2019, 10, 6873–6881. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.R.; Lv, L.X.; Shi, D.; Ye, J.Z.; Fang, D.Q.; Guo, F.F.; Li, Y.T.; He, X.K.; Li, L.J. Protective Effect of Akkermansia muciniphila against Immune-Mediated Liver Injury in a Mouse Model. Front. Microbiol. 2017, 8, 1804. [Google Scholar] [CrossRef] [PubMed]
- Schrader, T.; Rienhofer, A.; Andreesen, J.R. Selenium-containing xanthine dehydrogenase from Eubacterium barkeri. Eur. J. Biochem. 1999, 264, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Batt, S.M.; Wannemuehler, M.; Dispirito, A.; Beitz, D.C. Effect of feeding of a cholesterol-reducing bacterium, Eubacterium coprostanoligenes, to germ-free mice. Lab. Anim. Sci. 1998, 48, 253–255. [Google Scholar] [PubMed]
- Chen, Y.H.; Bai, J.; Wu, D.; Yu, S.F.; Qiang, X.L.; Bai, H.; Wang, H.N.; Peng, Z.W. Association between fecal microbiota and generalized anxiety disorder: Severity and early treatment response. J. Affect. Disord. 2019, 259, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xiang, X.; Shang, E.; Su, S.; Guo, J.; Qian, D.; Duan, J. Regulatory effect of total phenolic acid from the stems and leaves of Salvia miltiorrhiza Bge on intestinal microflora and short-chain fatty acids in type 2 diabetic nephropathy mice. Acta Pharm. Sin. 2021, 56, 1035–1048. [Google Scholar]
- Bloemendaal, M.; Szopinska-Tokov, J.; Belzer, C.; Boverhoff, D.; Papalini, S.; Michels, F.; van Hemert, S.; Vasquez, A.A.; Aarts, E. Probiotics-induced changes in gut microbial composition and its effects on cognitive performance after stress: Exploratory analyses. Transl. Psychiatr. 2021, 11, 300. [Google Scholar] [CrossRef]
- Bo, W.; Qing, K.; Xiu, L.; Jian, Z.; Hao, Z.; Wei, C.; Gang, W. A high-fat diet increases gut microbiota biodiversity and energy expenditure due to nutrient difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef]
- Hu, X.T.; Zhao, Y.; Yang, Y.; Gong, W.X.; Sun, X.M.; Yang, L.; Zhang, Q.; Jin, M.L. Akkermansia muciniphila Improves Host Defense against Influenza Virus Infection. Front. Microbiol. 2021, 11, 586476. [Google Scholar] [CrossRef]
- Huang, C.L.; Chen, J.; Wang, J.J.; Zhou, H.; Lu, Y.Y.; Lou, L.H.; Zheng, J.Y.; Tian, L.; Wang, X.P.; Cao, Z.W.; et al. Dysbiosis of Intestinal Microbiota and Decreased Antimicrobial Peptide Level in Paneth Cells during Hypertriglyceridemia-Related Acute Necrotizing Pancreatitis in Rats. Front. Microbiol. 2017, 8, 776. [Google Scholar] [CrossRef]
- Song, H.Z.; Shen, X.C.; Chu, Q.; Zheng, X.D. Vaccinium bracteatum Thunb. fruit extract reduces high-fat diet-induced obesity with modulation of the gut microbiota in obese mice. J. Food Biochem. 2021, 45, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Fujita, Y.; Ren, Q.; Ma, M.; Dong, C.; Hashimoto, K. Bifidobacterium in the gut microbiota confer resilience to chronic social defeat stress in mice. Sci. Rep. 2017, 7, 45942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemi, A.; Noorbala, A.A.; Azam, K.; Eskandari, M.H.; Djafarian, K. Effect of probiotic and prebiotic vs. placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin. Nutr. 2019, 38, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.W.; Xia, Y.J.; Liu, X.F.; Wang, G.Q.; Xiong, Z.Q.; Ai, L.Z. Antrodin A from mycelium of Antrodia camphorata alleviates acute alcoholic liver injury and modulates intestinal flora dysbiosis in mice. J. Ethnopharmacol. 2020, 254, 112681. [Google Scholar] [CrossRef] [PubMed]
- Tung, T.H.; Tung, Y.T.; Lin, I.H.; Shih, C.K.; Nguyen, N.T.K.; Shabrina, A.; Huang, S.Y. Fish Oil, but Not Olive Oil, Ameliorates Depressive-Like Behavior and Gut Microbiota Dysbiosis in Rats under Chronic Mild Stress. Biomolecules 2019, 9, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′ - > 3′) | PrimerBank ID |
|---|---|---|
| Mbp | F-5′-GGCGGTGACAGACTCCAAG-3′ | 51333a1 |
| R-5′-GAAGCTCGTCGGACTCTGAG-3′ | ||
| Grin1 | F-5′-TCCCAACGACCACTTCACTC-3′ | 294997255c3 |
| R-5′-AGTAGATGGACATTCGGGTAGTC-3′ | ||
| Grin2a | F-5′-ACGTGACAGAACGCGAACTT-3′ | 41680704c1 |
| R-5′-TCAGTGCGGTTCATCAATAACG-3′ | ||
| Grin2b | F-5′-GCCATGAACGAGACTGACCC-3′ | 6680099a1 |
| R-5′-GCTTCCTGGTCCGTGTCATC-3′ | ||
| Gfap | F-5′-GGGGCAAAAGCACCAAAGAAG-3′ | 32480796a1 |
| R-5′-GGGACAACTTGTATTGTGAGCC-3′ | ||
| CD36 | F-5′-ATGGGCTGTGATCGGAACTG-3′ | 227116348c1 |
| R-5′-GTCTTCCCAATAAGCATGTCTCC-3′ | ||
| Gapdh | F-5′-TCCTGCACCACCAACTGCT-3′ | -- |
| R-5′-GTCAGATCCACGACGGACACA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Tian, P.; Zhu, H.; Zou, R.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Lactobacillus paracasei CCFM1229 and Lactobacillus rhamnosus CCFM1228 Alleviated Depression- and Anxiety-Related Symptoms of Chronic Stress-Induced Depression in Mice by Regulating Xanthine Oxidase Activity in the Brain. Nutrients 2022, 14, 1294. https://doi.org/10.3390/nu14061294
Xu M, Tian P, Zhu H, Zou R, Zhao J, Zhang H, Wang G, Chen W. Lactobacillus paracasei CCFM1229 and Lactobacillus rhamnosus CCFM1228 Alleviated Depression- and Anxiety-Related Symptoms of Chronic Stress-Induced Depression in Mice by Regulating Xanthine Oxidase Activity in the Brain. Nutrients. 2022; 14(6):1294. https://doi.org/10.3390/nu14061294
Chicago/Turabian StyleXu, Mengshu, Peijun Tian, Huiyue Zhu, Renying Zou, Jianxin Zhao, Hao Zhang, Gang Wang, and Wei Chen. 2022. "Lactobacillus paracasei CCFM1229 and Lactobacillus rhamnosus CCFM1228 Alleviated Depression- and Anxiety-Related Symptoms of Chronic Stress-Induced Depression in Mice by Regulating Xanthine Oxidase Activity in the Brain" Nutrients 14, no. 6: 1294. https://doi.org/10.3390/nu14061294