
Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Culture and Water Stress
2.2. Growth Parameters
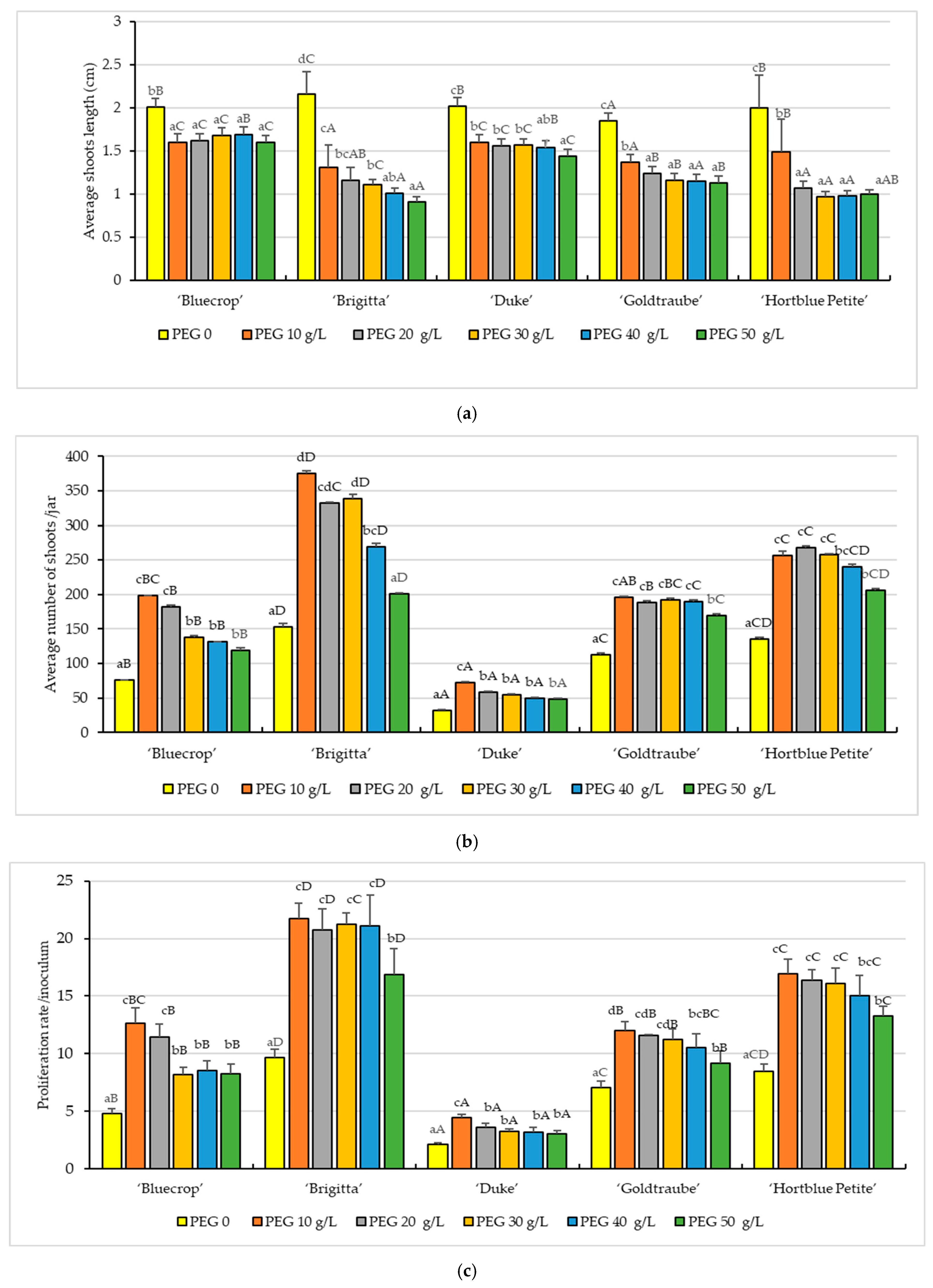
- Shoots length (cm): The shoots from 48 initial explants were measured (three vessels/treatment/variety) (SL).
- Proliferation rate (number of shoots/explants): Three recipients/treatment/variety were measured (PR).
- The average number of shoots/culture jar: The shoots from three recipients/treatment/variety were measured (NS).
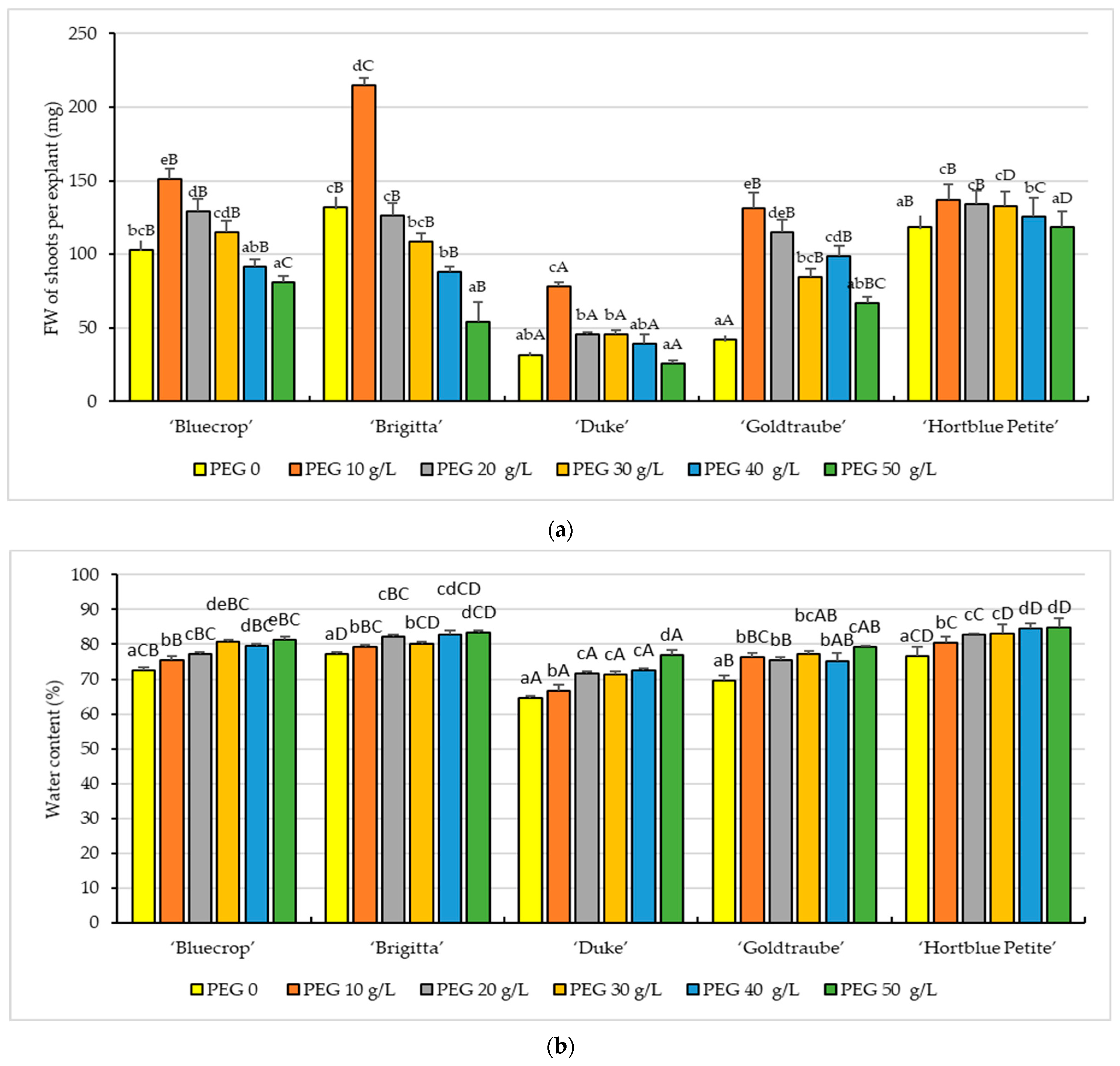
- Fresh weight (FW) of shoots per explant (mg): 30 explants for each treatment/variety were weighed immediately after the material was removed from the in vitro culture medium.
- Dry weight (mg): The material was dried for three days at 45 °C and re-weighted (DW).
- Water content (WC): Based on FW and DW, WC percentage was calculated using the formula [36]:WC (%) = ((Fresh Weight − Dry Weight)/Fresh Weight) ∗ 100
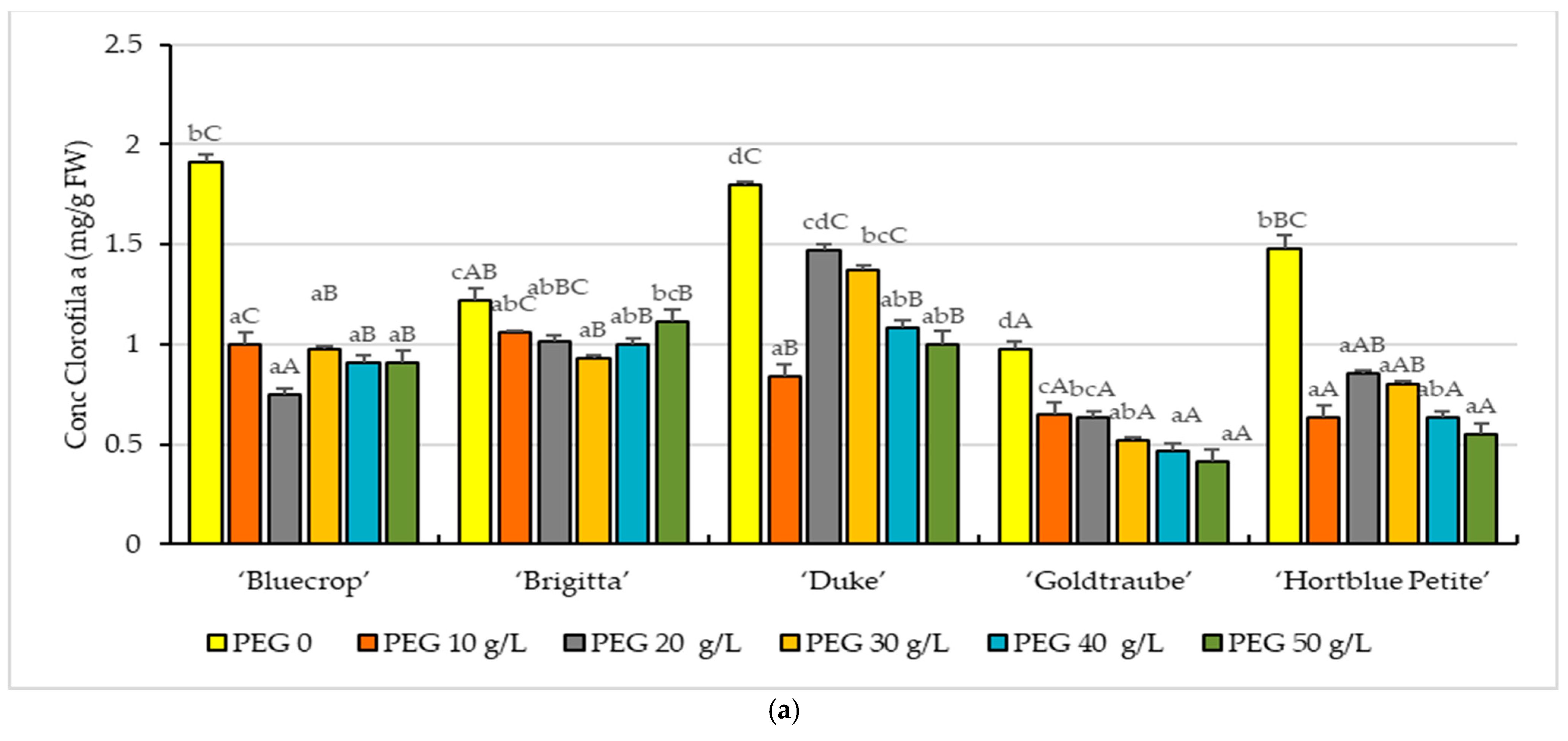
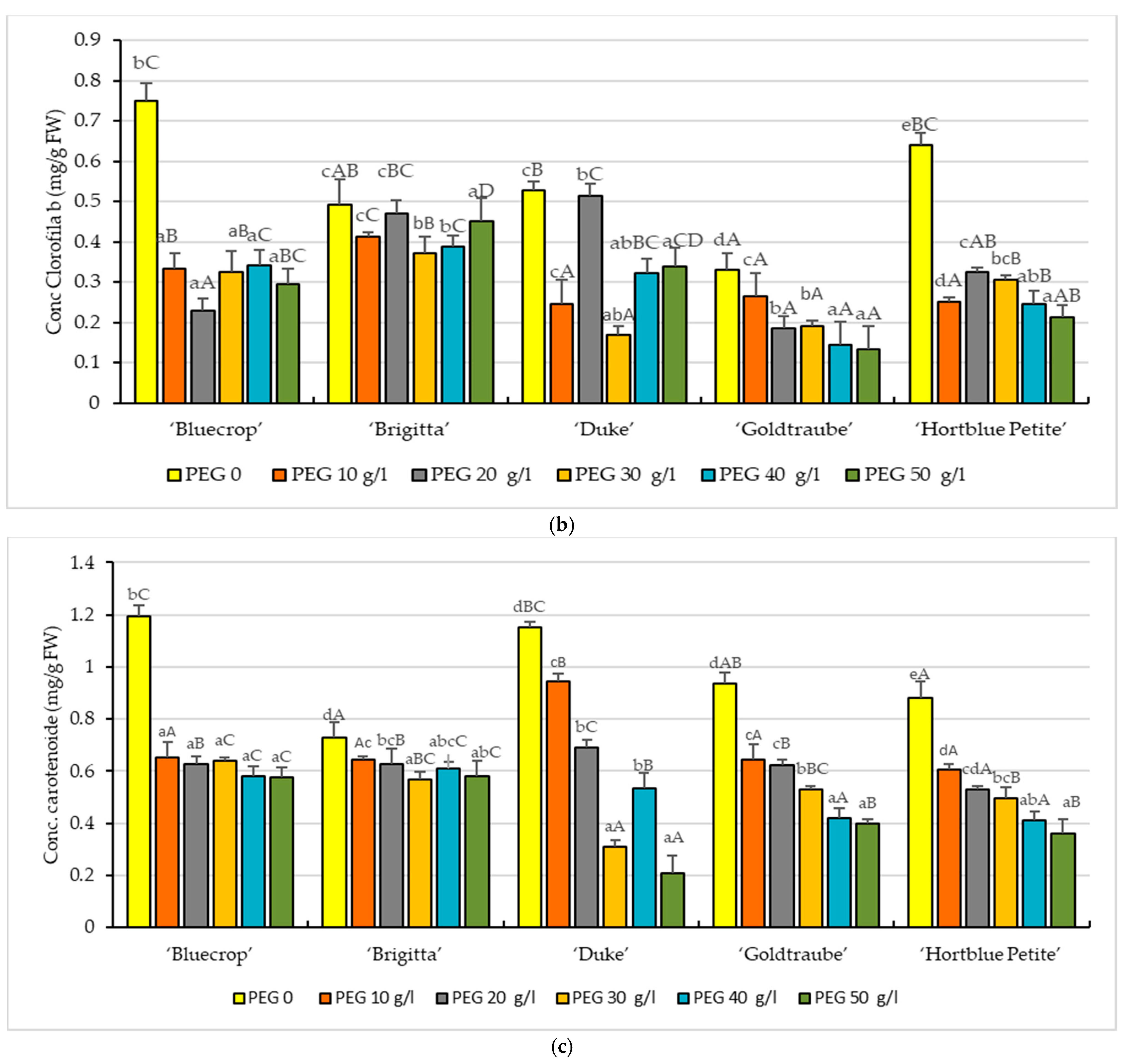
2.3. Photosynthetic Pigments
2.4. Statistical Analysis
3. Results
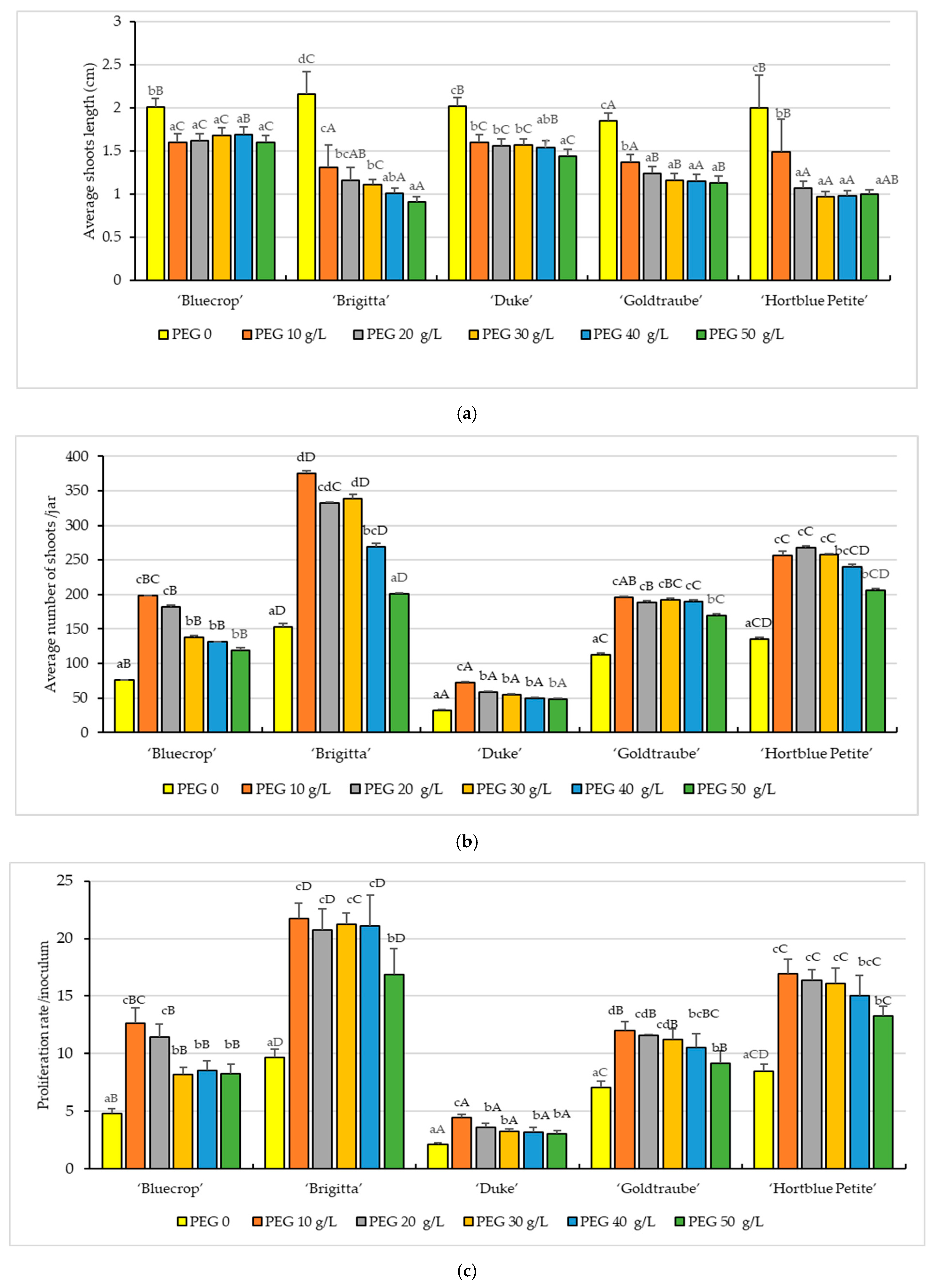
3.1. In Vitro Growth Parameters
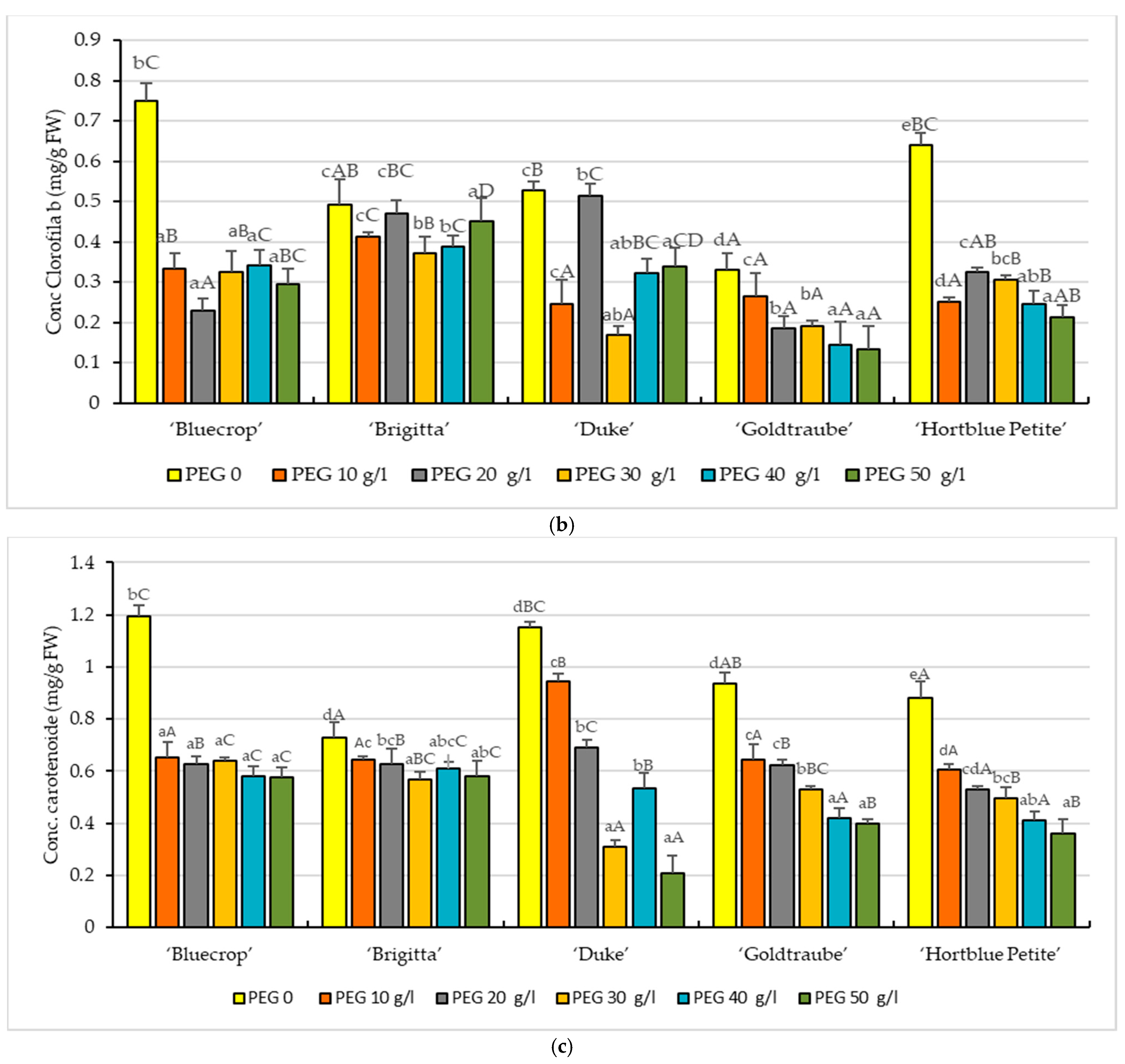
3.2. Photosynthetic Pigments
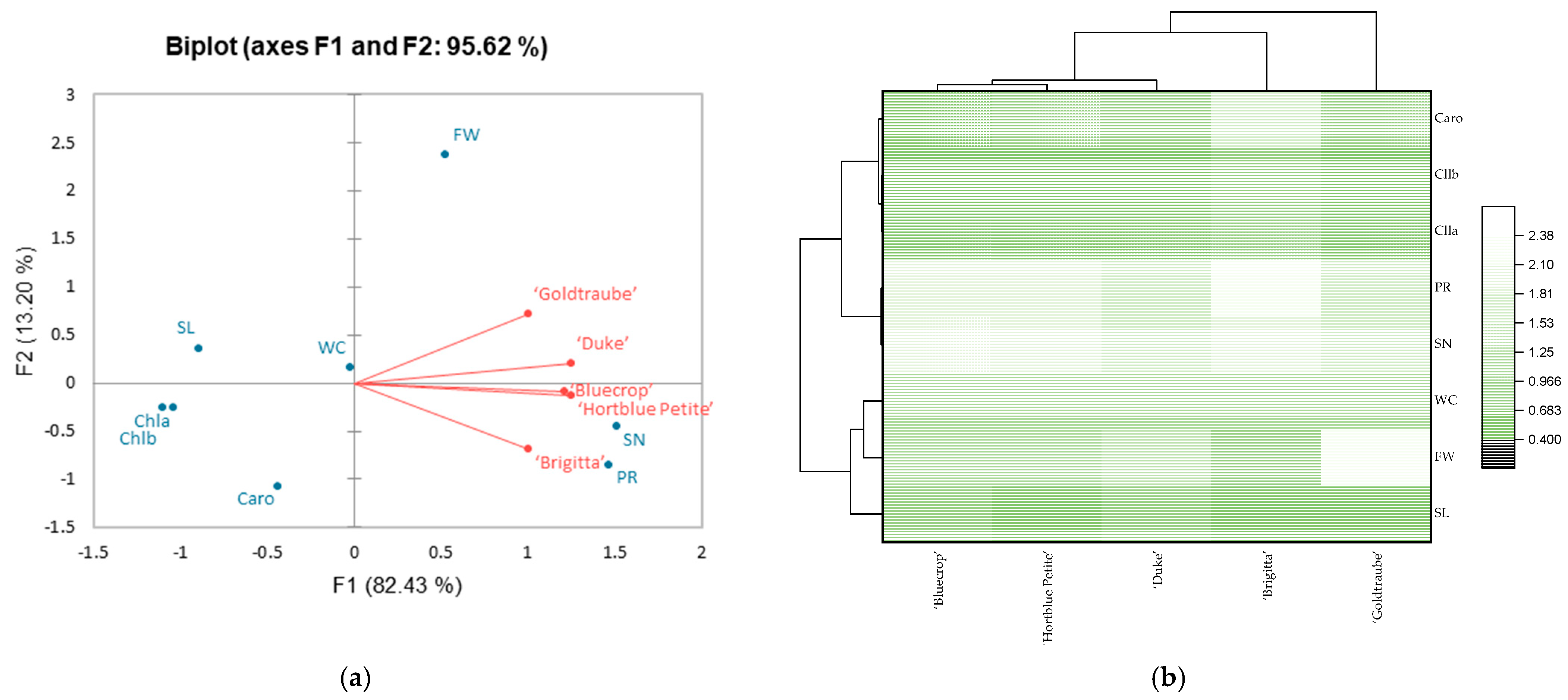
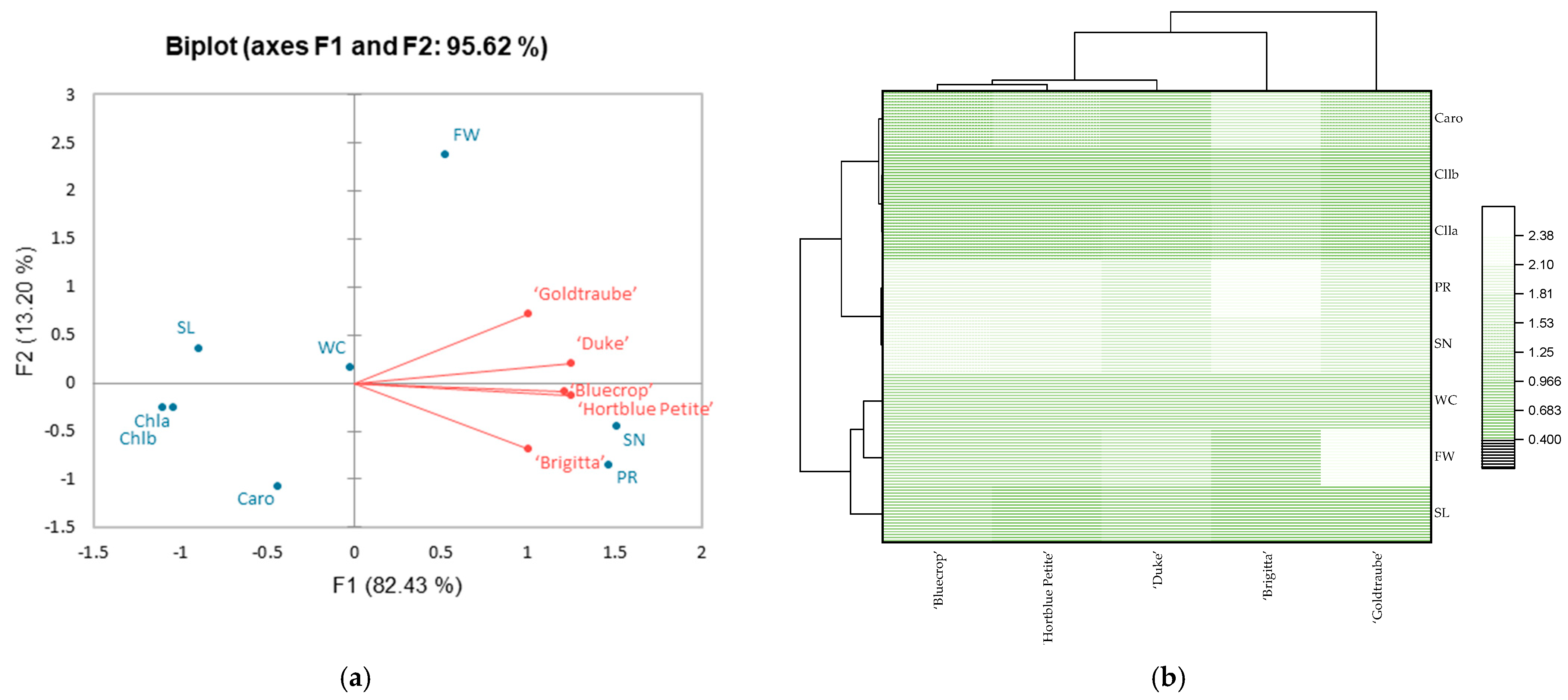
3.3. Principal Component Analysis (PCA)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brauman, K.A.; Siebert, S.; Foley, J.A. Improvements in crop water productivity increase water sustainability and food security—A global analysis. Environ. Res. Lett. 2013, 8, 024030. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar] [CrossRef] [Green Version]
- Toor, M.D.; Adnan, M.; Javed, M.S.; Habibah, U.; Arshad, A.; Din, M.M.; Ahmad, R. Foliar application of Zn: Best way to mitigate drought stress in plants: A review. Int. J. Appl. Sci. 2020, 6, 16–20. [Google Scholar]
- Jogawat, A.; Yadav, B.; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between phytohormones and secondary metabolites in the drought stress tolerance of crop plants: A review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Cattivelli, L.; Rizza, F.; Badeck, F.W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Marè, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Sivritepe, N.; Erturk, U.; Yerlikaya, C.; Turkan, I.; Bor, M.; Ozdemir, F. Response of the cherry rootstock to water stress induced in vitro. Biol. Plant 2008, 52, 573. [Google Scholar] [CrossRef]
- Bryla, D.R.; Strik, B.C. Effects of cultivar and plant spacing on the seasonal water requirements of highbush blueberry. J. Am. Soc. Hortic. Sci. 2007, 132, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Bryla, D.R. Chapter 9. Crop evapotranspiration and irrigation scheduling in blueberry. In Evapotranspiration—From Measurements to Agricultural and Environmental Applications; Gerosa, G., Ed.; Intech: Rijeka, Croatia, 2011; pp. 167–186. Available online: https://books.google.ro/books?hl=ro&lr=&id=JGmQDwAAQBAJ&oi=fnd&pg=PA167&dq=9.%09Bryla,+D.+R.+Chapter+9 (accessed on 10 December 2021).
- Shireman, A.D.; Bryla, D.R.; Yorgey, B. Irrigation Management Effects on Yield and Fruit Quality of Highbush Blueberry. Acta Hortic. 2009, 810, 649–656. [Google Scholar] [CrossRef]
- Strik, B.C. Blueberry: An expanding world berry crop. Chron. Hortic. 2005, 45, 7–12. [Google Scholar]
- Brazelton, C. World Blueberry Acreage & Production; USHBC: Folsom, CA, USA, 2011; Available online: www.oregonblueberry.com (accessed on 21 November 2021).
- Howell, A.B. Update on health benefits of cranberry and blueberry. Acta Hortic. 2009, 810, 779–785. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [Green Version]
- Khayatnezhad, M.; Gholamin, R.; Jamaatie-Somarin, S.H.; Zabihi-Mahmoodabad, R. Effects of peg stress on corn cultivars (Zea mays L.) at germination stage. World Appl. Sci. J. 2010, 11, 504–506. [Google Scholar]
- Meher, P.S.; Reddy, K.A.; Rao, D.M. Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J. Biol. Sci. 2018, 25, 285. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, C.; Celletti, S.; Cristofori, V.; Astolfi, S.; Ruggiero, B.; Rugini, E. Olive (Olea europaea L.) plants trans-genic for tobacco osmotin gene are less sensitive to in vitro-induced drought stress. Acta Physiol. Plant 2017, 39, 229. [Google Scholar] [CrossRef]
- Salma, U.K.; Khatun, F.; Bhuiyan, M.J.H.; Yasmin, S.; Khan, T.H. In vitro screening for drought tolerance of some chickpea varieties in Bangladesh. Progress. Agric. 2016, 27, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Youssef, M.; Mohamed, E.S.A.; El-Sayed, E.S.N.; Abouzaid, E. Establishment of an in vitro Evaluation Method for Drought Tolerance in Guava. Res. J. Appl. Biotechnol. 2016, 5, 32–38. [Google Scholar]
- Foti, C.; Kalampokis, I.F.; Aliferis, K.A.; Pavli, O.I. Metabolic Responses of Two Contrasting Lentil Genotypes to PEG-Induced Drought Stress. Agronomy 2021, 11, 1190. [Google Scholar] [CrossRef]
- Hernández-Pérez, C.A.; Gómez-Merino, F.C.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro screening of sugarcane cultivars (Saccharum spp. hybrids) for tolerance to polyethylene glycol-induced water stress. Agronomy 2021, 11, 598. [Google Scholar] [CrossRef]
- Kovalikova, Z.; Jiroutova, P.; Toman, J.; Dobrovolna, D.; Drbohlavova, L. Physiological Responses of Apple and Cherry In Vitro Culture under Different Levels of Drought Stress. Agronomy 2020, 10, 1689. [Google Scholar] [CrossRef]
- Kim, Y.; Kang, S.S.; Cho, K.S.; Hwang, H.S.; Han, J.; Han, T. Evaluation of the growth and responses of in vitro cultures of several pear rootstocks exposed to simulated drought stress using polyethylene glycol treatments. Acta Hortic. 2010, 909, 219–222. [Google Scholar] [CrossRef]
- Debnath, S.C. Propagation of Vaccinium in vitro: A review. Int. J. Fruit Sci. 2008, 6, 47–71. [Google Scholar] [CrossRef]
- Ružić, D.; Vujović, T.; Libiakova, G.; Cerović, R.; Gajdošova, A. Micropropagation in vitro of highbush blueberry (Vaccinium corymbosum L.). J. Berry Res. 2012, 2, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Vescan, L.A.; Pamfil, D.; Clapa, D.; Fira, A.; Sisea, C.R.; Pop, I.F.; Petricele, I.V.; Ciuzan, O.; Pop, R. Efficient micropropagation protocol for highbush blueberry (Vaccinium corymbosum L.) cv.‘Elliot’. Rom. Biotechnol. Lett. 2012, 17, 6893–6902. [Google Scholar]
- Borsai, O.; Hârța, M.; Pamfil, D.; Clapa, D. The Effect of Cytokinins on Micropropagation Success of Highbush Blueberry (Vaccinium corymbosum L.). Agricultura 2019, 3–4, 93–100. [Google Scholar] [CrossRef]
- Debnath, S.C.; Goyali, J.C. In vitro propagation and variation of Antioxidant properties in micropropagated Vaccinium berry plants—A review. Molecules 2020, 25, 788. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, M.; Kondakova, V. In vitro propagation of Vaccinium corymbosum L. Bulg. J. Agric. Sci. 2021, 27, 323–327. [Google Scholar]
- Cohen, D.; Elliot, D. Micropropagation methods for blueberries and tamarillos. Int. Propag. Soc. Comb. Proc. 1979, 29, 177–179. [Google Scholar]
- Debnath, S.C.; Vyas, P.; Goyali, J.C.; Igamberdiev, A.U. Morphological and molecular analyses in micropropagated berry plants acclimatized under ex vitro condition. Can. J. Plant Sci. 2012, 92, 1065–1073. [Google Scholar] [CrossRef]
- Debnath, S.C. Strategies to propagate Vaccinium nuclear stocks for the Canadian berry industry. Can. J. Plant Sci. 2007, 87, 911–922. [Google Scholar] [CrossRef]
- El-Shiekh, A.; Wildung, D.K.; Luby, J.J.; Sargent, K.L.; Read, P.E. Long-term effects of propagation by tissue culture or softwood single-node cuttings on growth habit, yield, and berry weight of Northblue’blueberry. J. Am. Soc. Hortic. Sci. 1996, 121, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, G.; McCown, B. Commercially feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Intl. Plant Prop. Soc. 1980, 30, 421–427. [Google Scholar]
- Clapa, D.; Bunea, C.; Borsai, O.; Pintea, A.; Hârța, M.; Ştefan, R.; Fira, A. The role of Sequestrene 138 in highbush blueberry (Vaccinium corymbosum L.) micropropagation. HortScience 2018, 53, 1487–1493. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, M.; Siekierzyńska, A.; Jacek, B.; Litwińczuk, W. Differences in response to drought stress among highbush blueberry plants propagated conventionally and by tissue culture. Plant Biosyst. 2021, 155, 172–178. [Google Scholar] [CrossRef]
- Britton, G.; Schiedt, K.; Liaaen-Jensen, S. Isolation and analysis. In Carotenoids; Birkhauser Verlag: Basel, Switzerland, 1995; Volume 1A, pp. 104–107. [Google Scholar]
- Ameglio, T.; Le Roux, X.; Mingeau, M.; Perrier, C. Water relations of highbush blueberry under drought conditions. Acta Hortic. 2000, 537, 273–278. [Google Scholar] [CrossRef]
- Ortega-Farias, S.; Espinoza-Meza, S.; López-Olivari, R.; Araya-Alman, M.; Carrasco-Benavides, M. Effects of different irrigation levels on plant water status, yield, fruit quality, and water productivity in a drip-irrigated blueberry orchard under Mediterranean conditions. Agric. Water Manag. 2021, 249, 106805. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Tsago, Y.; Andargie, M.; Takele, A. In vitro selection of sorghum (Sorghum bicolor (L) Moench) for polyethylene glycol (PEG) induced drought stress. Plant Sci. Today 2014, 1, 62–68. [Google Scholar] [CrossRef]
- Abdulmalik, M.M.; Usman, I.S.; Usman, A.; Mohammed, M.S.; Sani, L.A. In vitro screening of groundnut (Arachis hypogaea L.) varieties for drought tolerance using polyethylene glycol (PEG 6000). FUDMA J. Sci. (FJS) 2018, 2, 251–255. [Google Scholar]
- Mengesha, B.; Mekbib, F.; Abraha, E. In Vitro Screening of Cactus [Opuntia ficus-indicia (L.) Mill] Genotypes for Drought Tolerance. Am. J. Plant Sci. 2016, 7, 1741–1758. [Google Scholar] [CrossRef] [Green Version]
- Magangana, T.P.; Stander, M.A.; Masondo, N.A.; Makunga, N.P. Steviol glycoside content and essential oil profiles of Stevia rebaudiana Bertoni in response to NaCl and polyethylene glycol as inducers of salinity and drought stress in vitro. PCTOC 2021, 145, 1–18. [Google Scholar] [CrossRef]
- Martínez-Santos, E.; Cruz-Cruz, C.A.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro response of vanilla (Vanilla planifolia Jacks. ex Andrews) to PEG-induced osmotic stress. Sci. Rep. 2021, 11, 22611. [Google Scholar] [CrossRef] [PubMed]
- Tafreshi, S.A.H.; Aghaie, P.; Momayez, H.R.; Hejaziyan, S.A. Response of in vitro-regenerated Myrtus communis L. shoots to PEG-induced water stress. Biocatal. Agric. Biotechnol. 2021, 34, 102033. [Google Scholar] [CrossRef]
- Razavizadeh, R.; Farahzadianpoor, F.; Adabavazeh, F. Komatsu, S. Physiological and morphological analyses of Thymus vulgaris L. in vitro cultures under polyethylene glycol (PEG)-induced osmotic stress. Vitr. Cell. Dev. Biol. Plant 2019, 55, 342–357. [Google Scholar] [CrossRef]
- Clapa, D.; Harta, M. Effects of PEG 6000 stress on strawberry (Fragaria x ananassa duch.) in vitro propagation. Sci. Pap. Ser. B Hortic. 2021, LXV, 58–63. [Google Scholar]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. C. R. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Yang, Y. PEG 6000-stimulated drought stress improves the attributes of in vitro growth, steviol glycosides production, and antioxidant activities in Stevia rebaudiana Bertoni. Plants 2020, 9, 1552. [Google Scholar] [CrossRef]
- Karimi, S.; Yadollahi, A.; Nazari-Moghadam, R.; Imani, A.; Arzani, K. In vitro screening of almond (Prunus dulcis (Mill.)) genotypes for drought tolerance. J. Biol. Environ. Sci. 2012, 6, 263–270. [Google Scholar]
- Gao, H.; Xu, D.; Zhang, H.; Cheng, X.; Yang, Q. Effects of culture medium composition and PEG on hyperhydricity in Dendrobium officinale. Vitr. Cell. Dev. Biol. Plant 2020, 56, 143–149. [Google Scholar] [CrossRef]
- Aazami, M.A.; Torabi, M.; Jalili, E. In vitro response of promising tomato genotypes for tolerance to osmotic stress. Afr. J. Biotechnol. 2010, 9, 4014–4017. [Google Scholar]
- Zhong, Y.P.; Li, Z.; Bai, D.F.; Qi, X.J.; Chen, J.Y.; Wei, C.G.; Lin, M.-M.; Fang, J.B. In vitro variation of drought tolerance in five actinidia species. J. Am. Soc. Hortic. Sci. 2018, 143, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Mastuti, R.; Rosyidah, M. In Vitro Environmental Stresses for Enhancing Withanolides Production in Physalis angulata L. Environ. Earth Sci. 2019, 239, 012011. [Google Scholar] [CrossRef]
- Simsek, O. Effect of drought stress in in vitro and drought related gene expression in Carrizo citrange. Fresenius Environ. Bull. 2018, 7, 9167–9171. [Google Scholar]
- Shereen, A.; Khanzada, M.A.; Baloch, M.A.W.; Shirazi, M.U.; Khan, M.A.; Arif, M. Effects of peg induced water stress on growth and physiological responses of rice genotypes at seedling stage. Pak. J. Bot. 2019, 51, 2013–2021. [Google Scholar] [CrossRef]
- Sen, A.; Alikamanoglu, S. Antioxidant enzyme activities, malondialdehyde, and total phenolic content of PEG-induced hyperhydric leaves in sugar beet tissue culture. Vitr. Cell. Dev. Biol. Plant 2013, 49, 396–404. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Afshari, F.; Nakhaei, F.; Mosavi, S.; Seghatoleslami, M. Evaluating the role of nutri-priming in improving PEG-induced drought stress tolerance of stevia (Stevia rebuadiana Bertoni). Iran. J. Plant Physiol. 2020, 11, 3509–3522. [Google Scholar] [CrossRef]
- El Din, A.F.Z.; Ibrahim, M.F.; Farag, R.; Abd El-Gawad, H.G.; El-Banhawy, A.; Alaraidh, I.A.; Rashad, Y.M.; Lashin, I.; El-Yazied, A.A.; Elkelish, A.; et al. Influence of Polyethylene Glycol on Leaf Anatomy, Stomatal Behavior, Water Loss, and Some Physiological Traits of Date Palm Plantlets Grown In Vitro and Ex Vitro. Plants 2020, 9, 1440. [Google Scholar] [CrossRef]
- Chiu, Y.C.; Chen, B.J.; Su, Y.S.; Huang, W.D.; Chen, C.C. A Leaf Disc Assay for Evaluating the Response of Tea (Camellia sinensis) to PEG-Induced Osmotic Stress and Protective Effects of Azoxystrobin against Drought. Plants 2021, 10, 546. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnar, S.; Clapa, D.; Mitre, V. Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol. Agronomy 2022, 12, 732. https://doi.org/10.3390/agronomy12030732
Molnar S, Clapa D, Mitre V. Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol. Agronomy. 2022; 12(3):732. https://doi.org/10.3390/agronomy12030732
Chicago/Turabian StyleMolnar, Sabin, Doina Clapa, and Viorel Mitre. 2022. "Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol" Agronomy 12, no. 3: 732. https://doi.org/10.3390/agronomy12030732
APA StyleMolnar, S., Clapa, D., & Mitre, V. (2022). Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol. Agronomy, 12(3), 732. https://doi.org/10.3390/agronomy12030732