
Changes and Relationships between Components in the Plant-Soil System and the Dominant Plant Functional Groups in Alpine Kobresia Meadows Due to Overgrazing

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
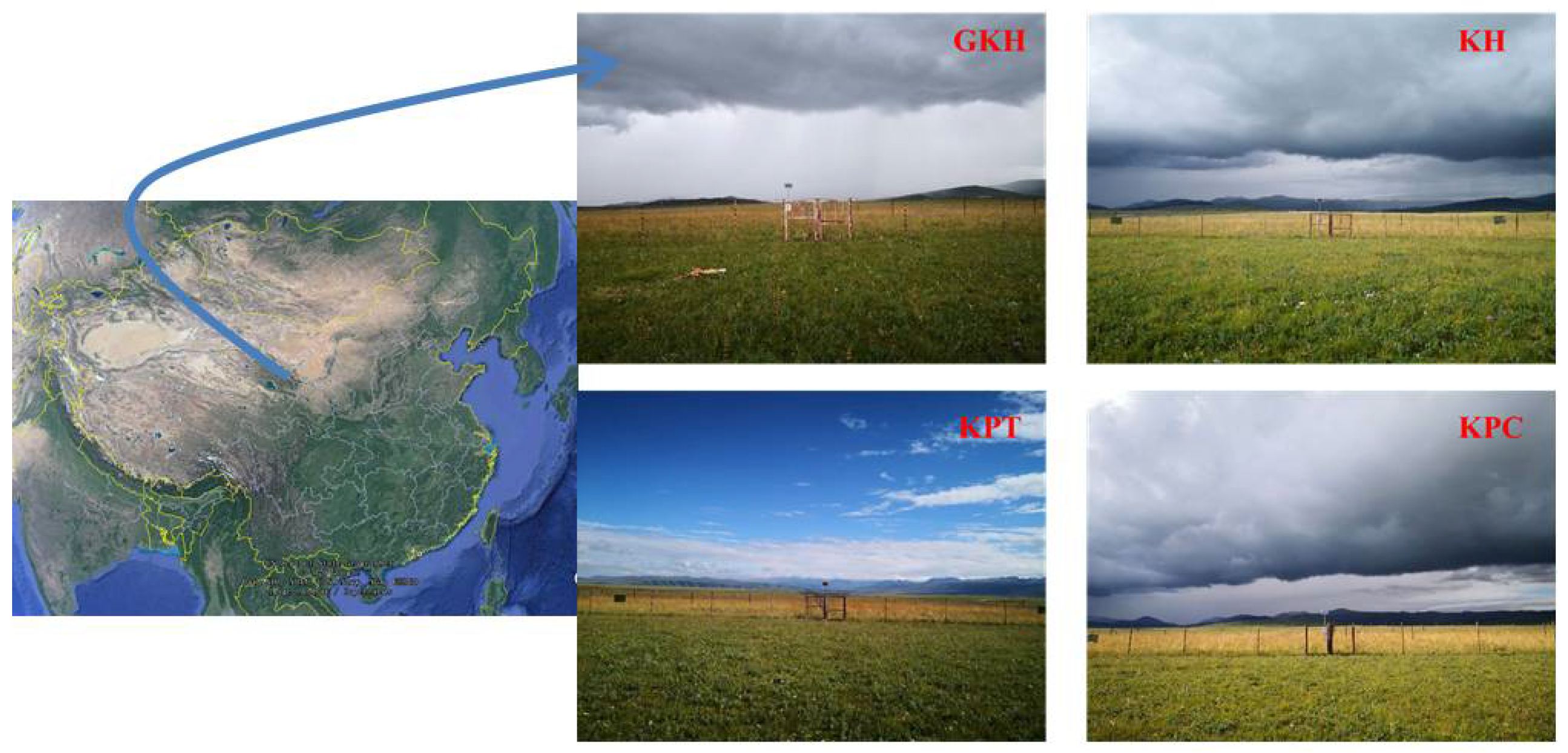
2.1. Study Sites
2.2. Experimental Design
2.3. Soil and Plant Sampling
2.4. Statistical Analyses
3. Results
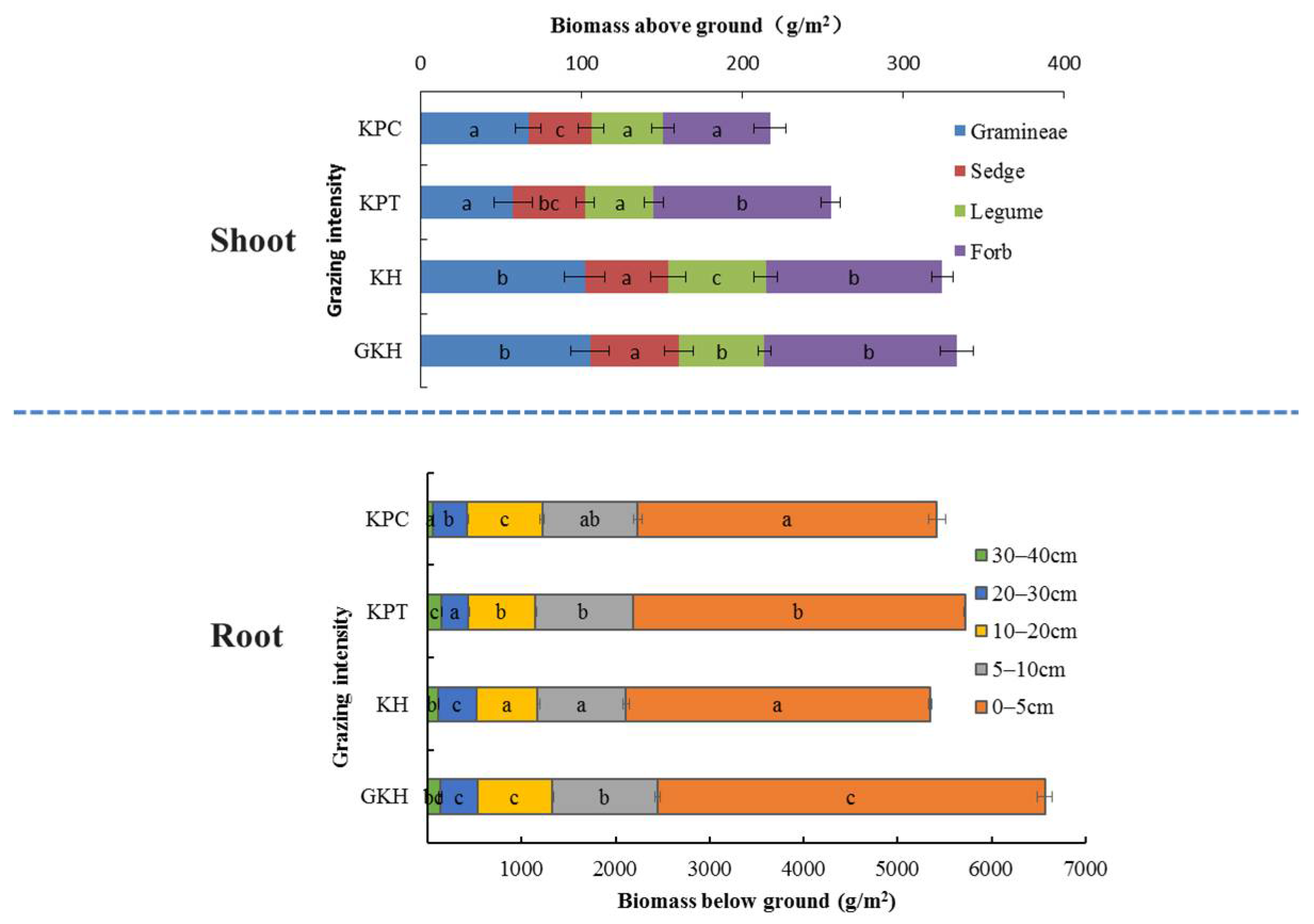
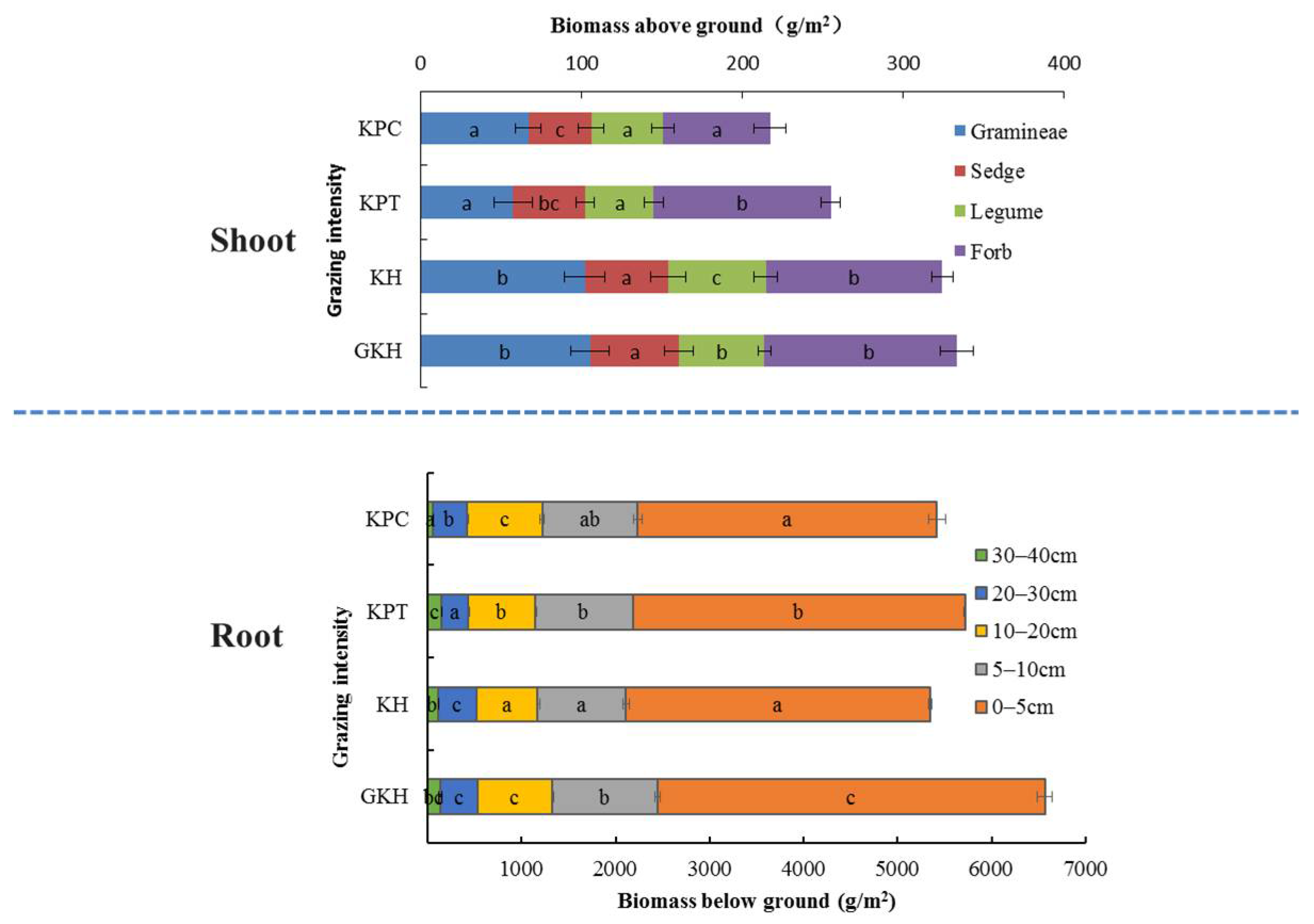
3.1. Plant Community Characteristics
3.1.1. Aboveground Plant Community Characteristics
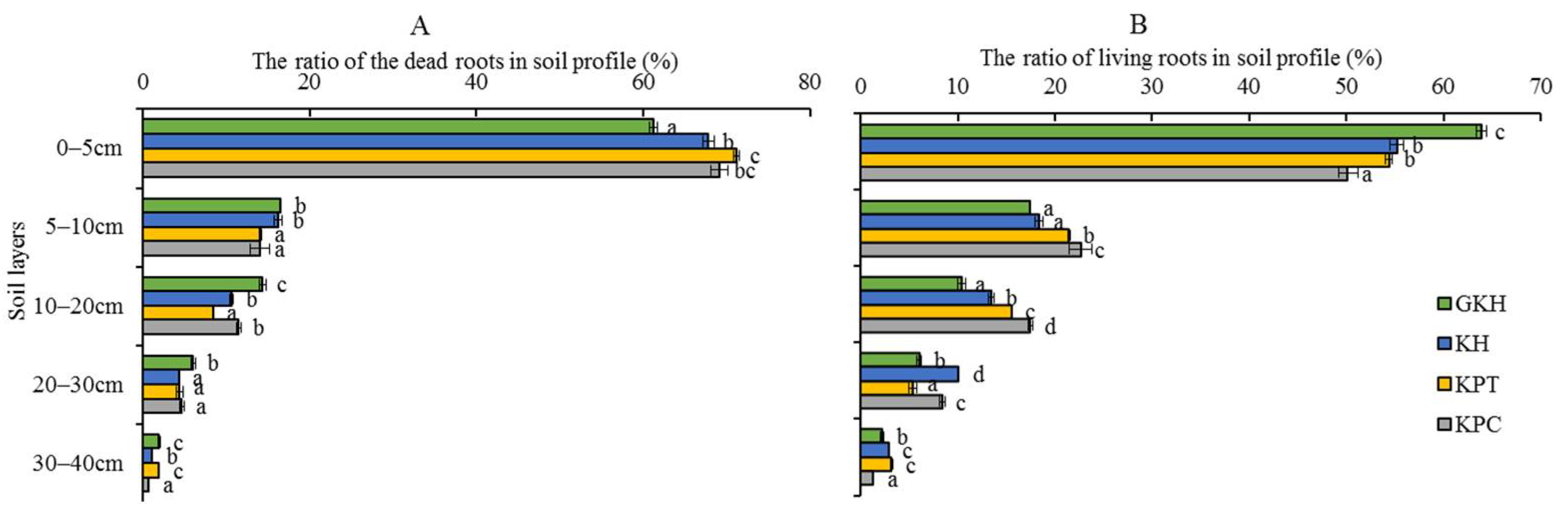
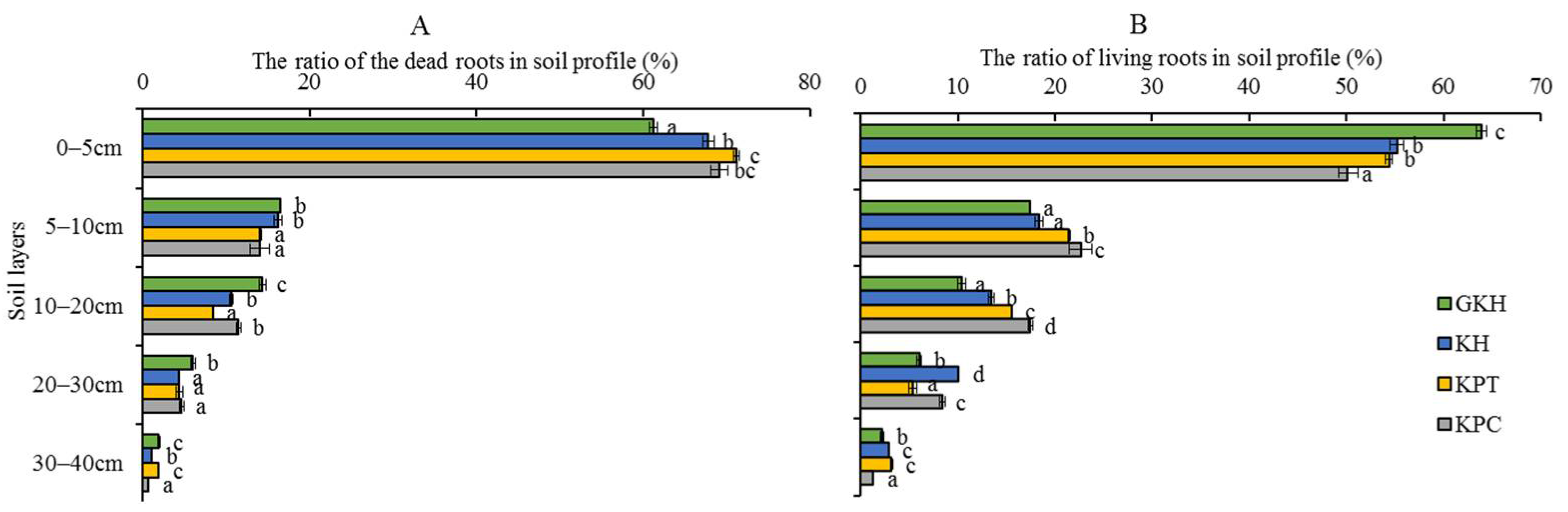
3.1.2. Belowground Plant Community Characteristics
3.2. Concentrations of C and N in the Plant Community
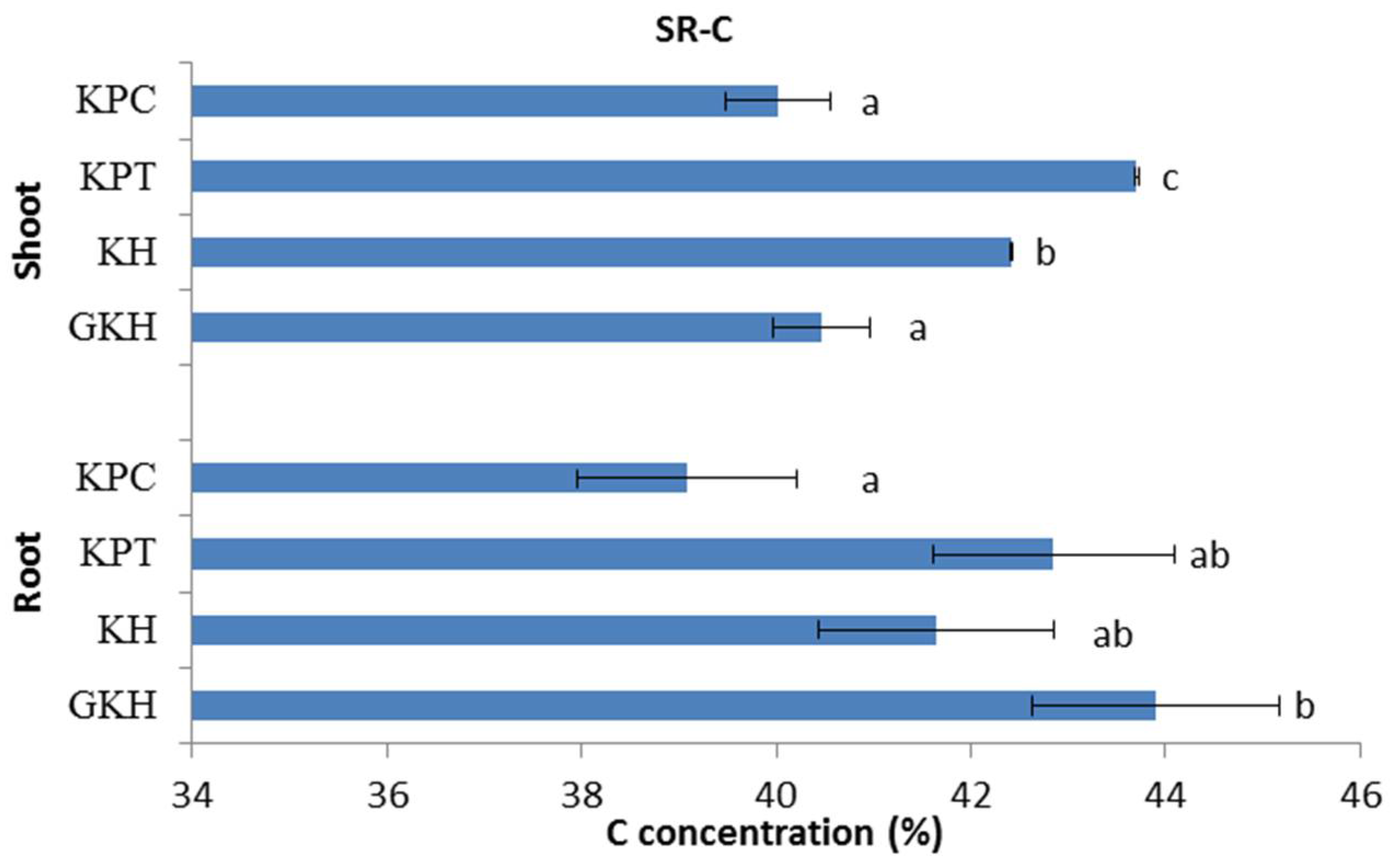
3.2.1. Concentration of C in the Plant Community
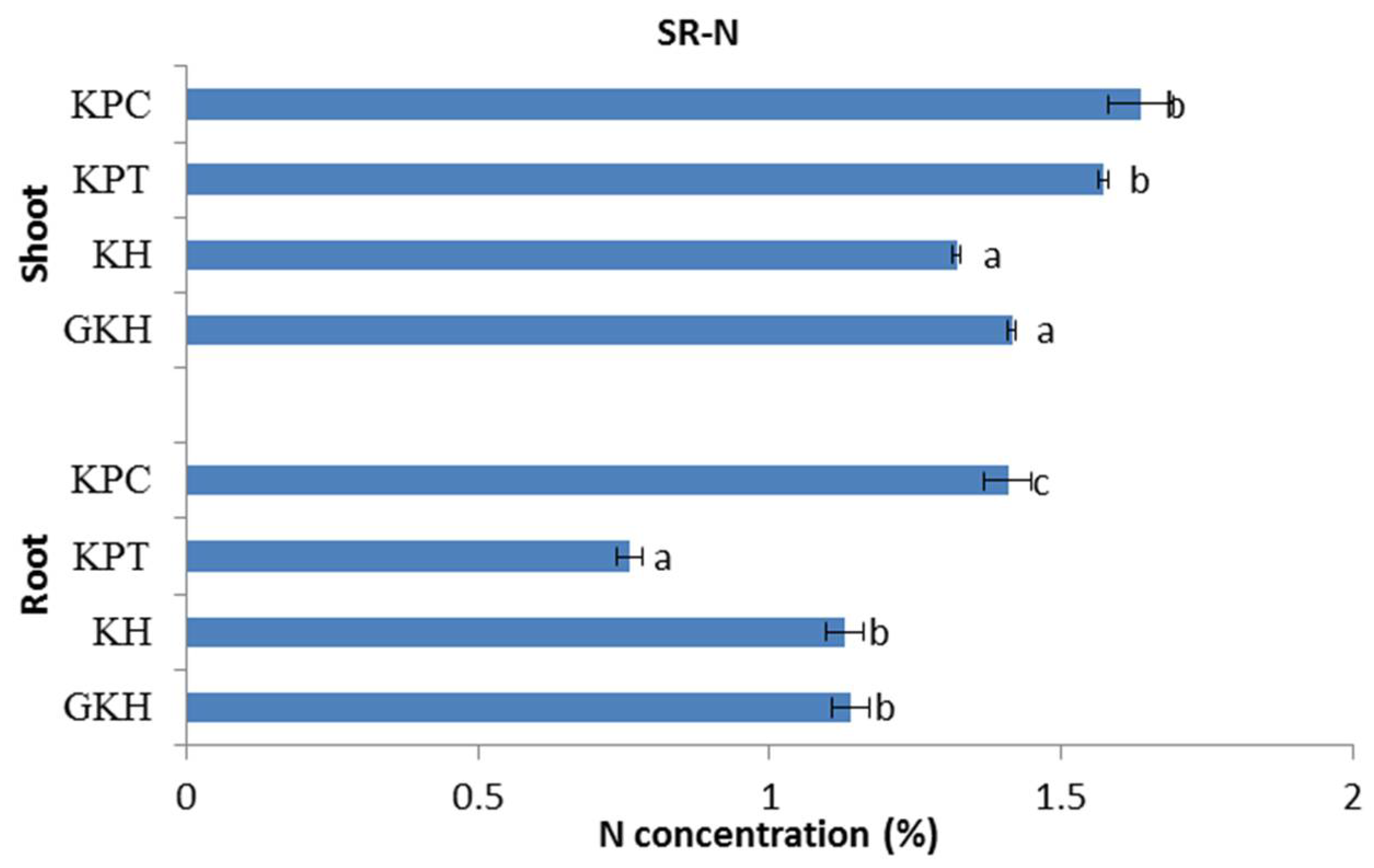
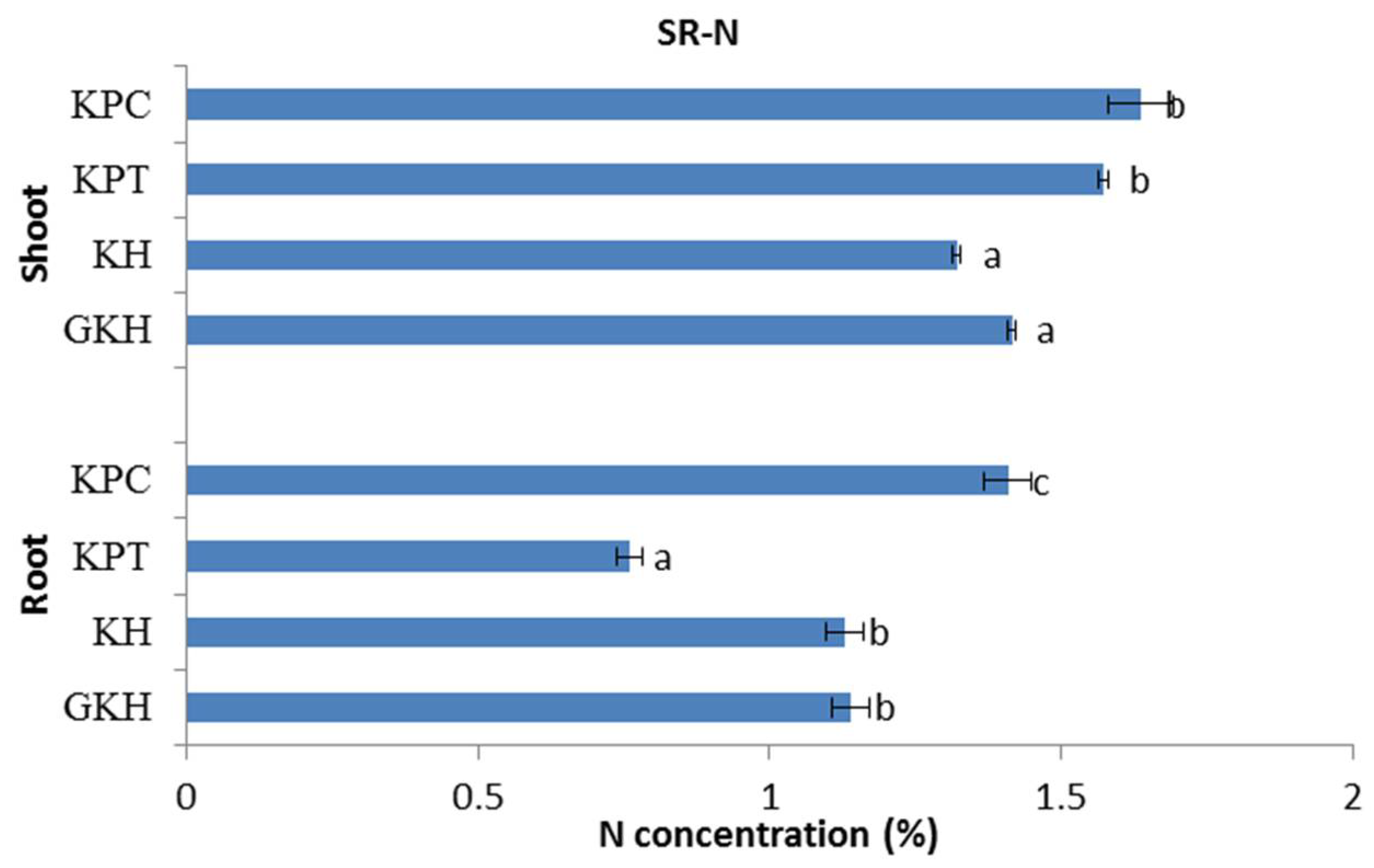
3.2.2. Concentration of N in the Plant Community
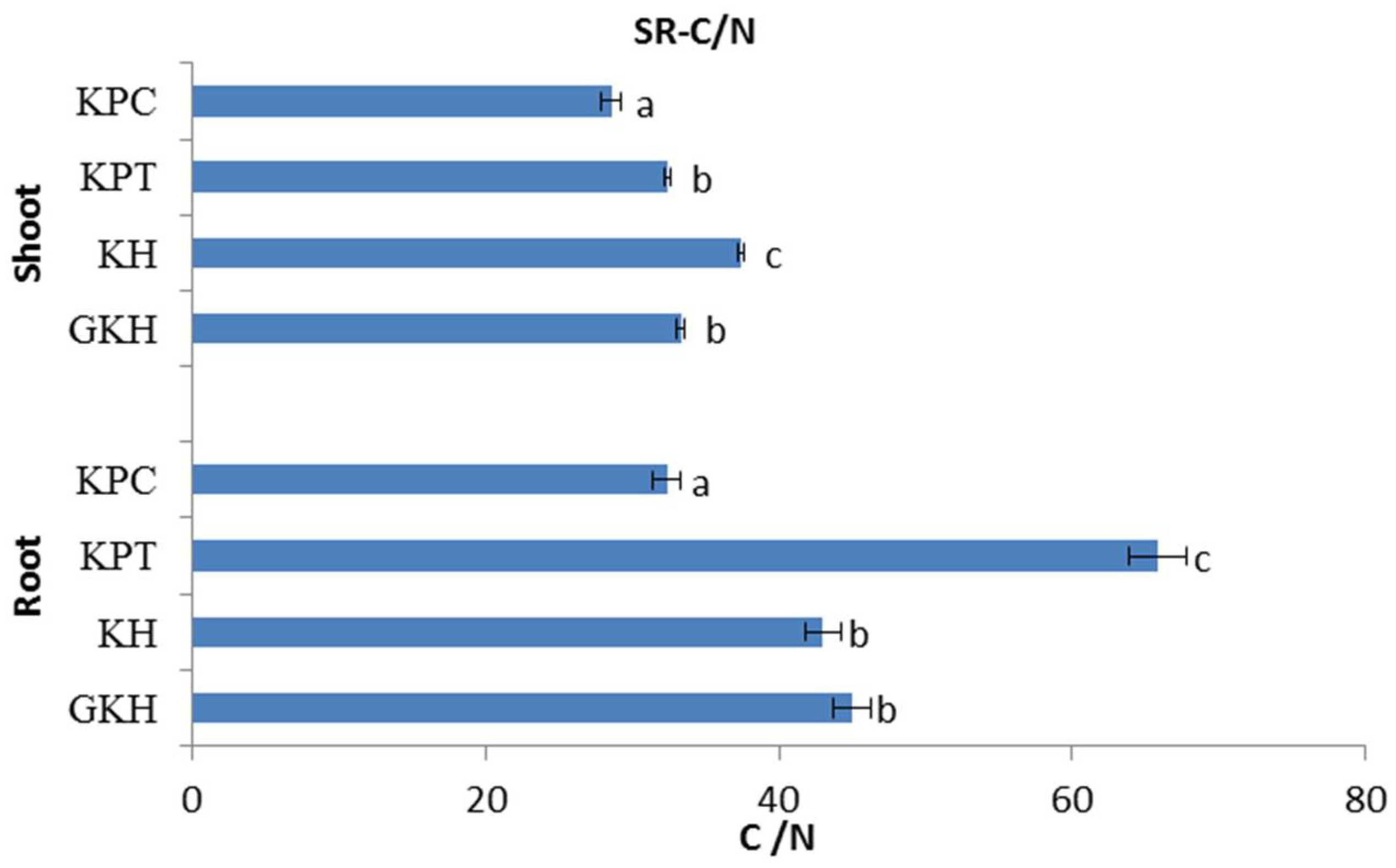
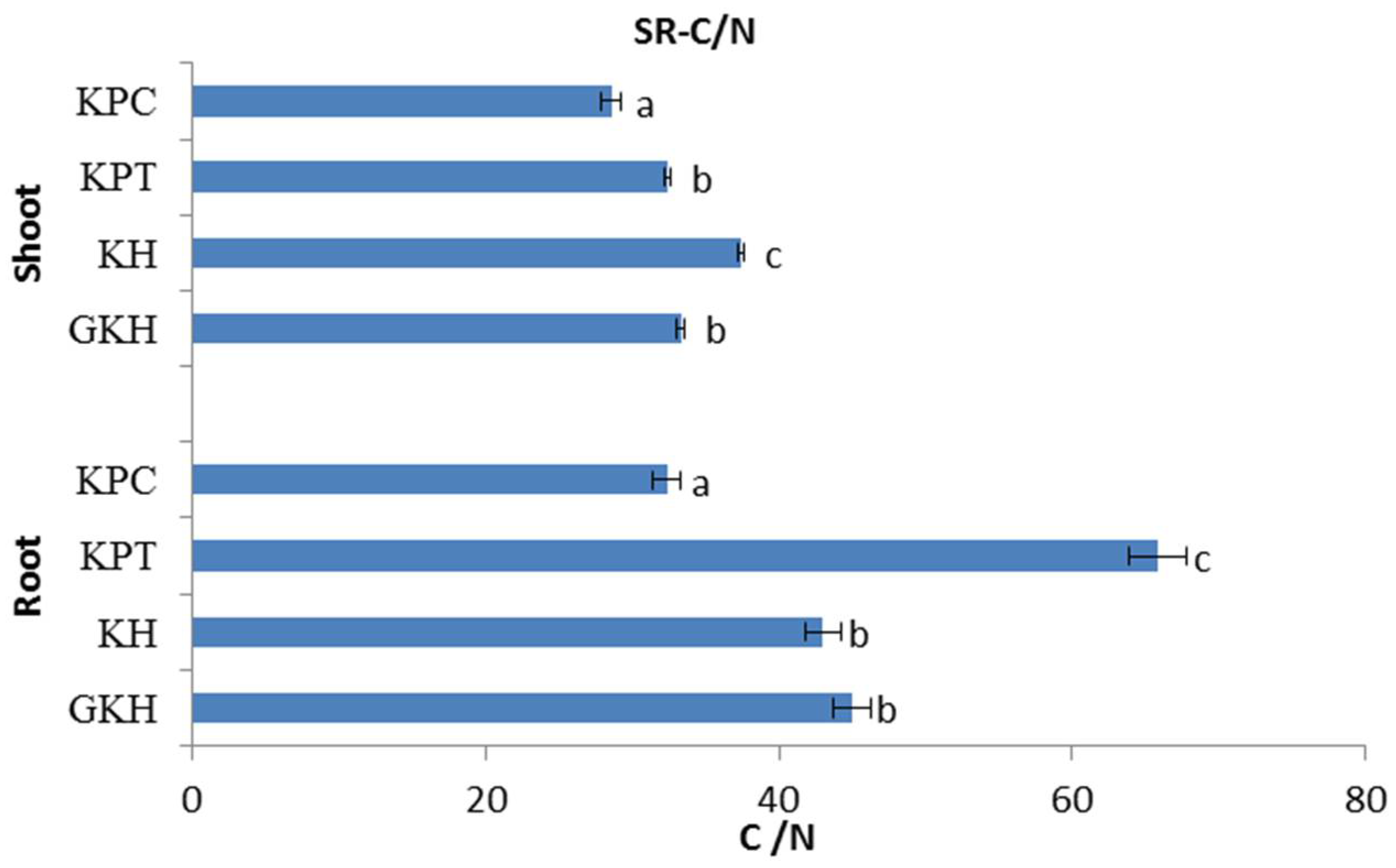
3.2.3. C/N in the Plant Community
3.3. Soil Nutrient Content
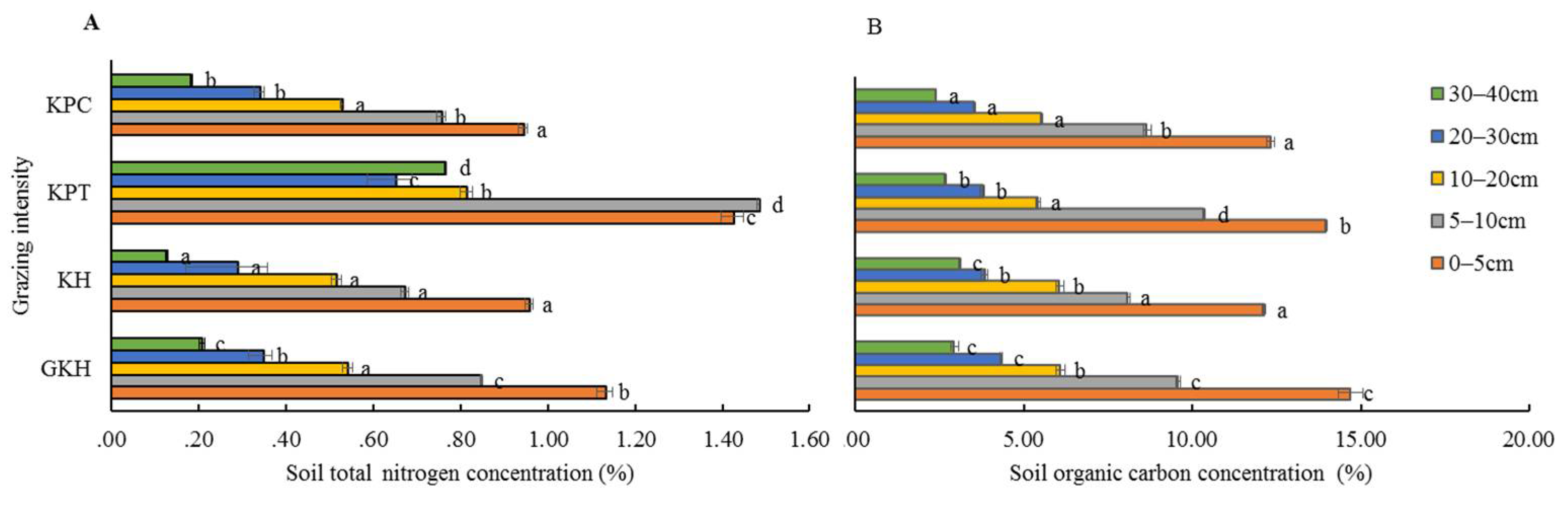
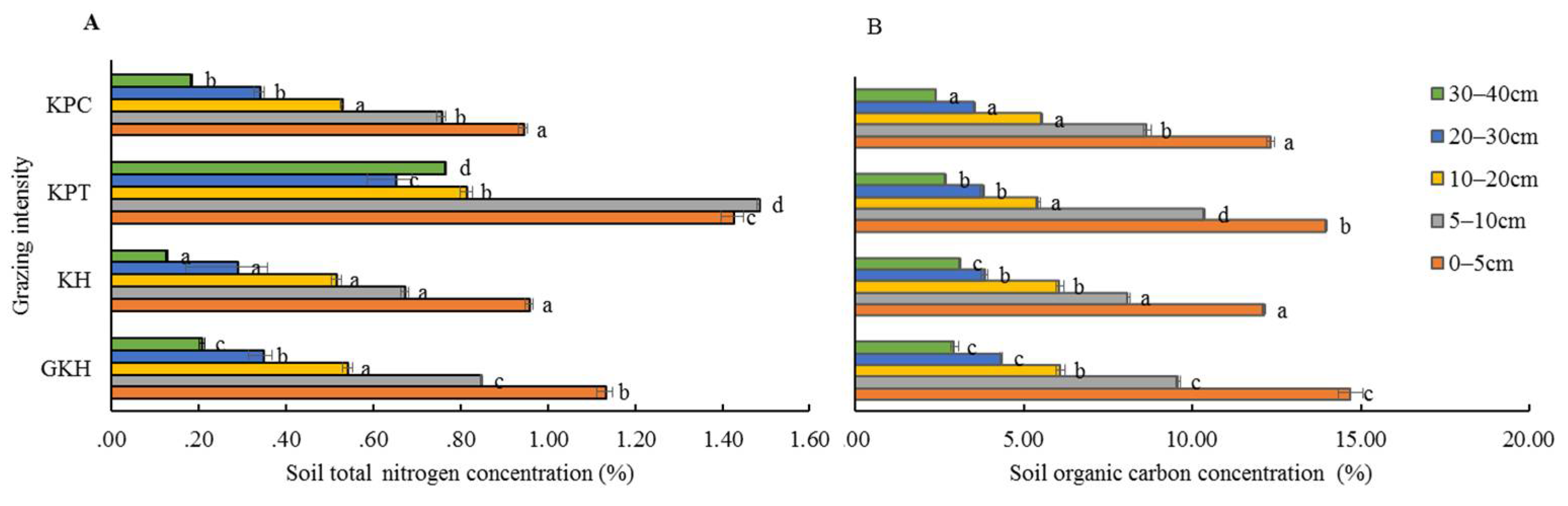
3.3.1. Concentration of Organic C in Soil
3.3.2. Concentration of Total N in Soil
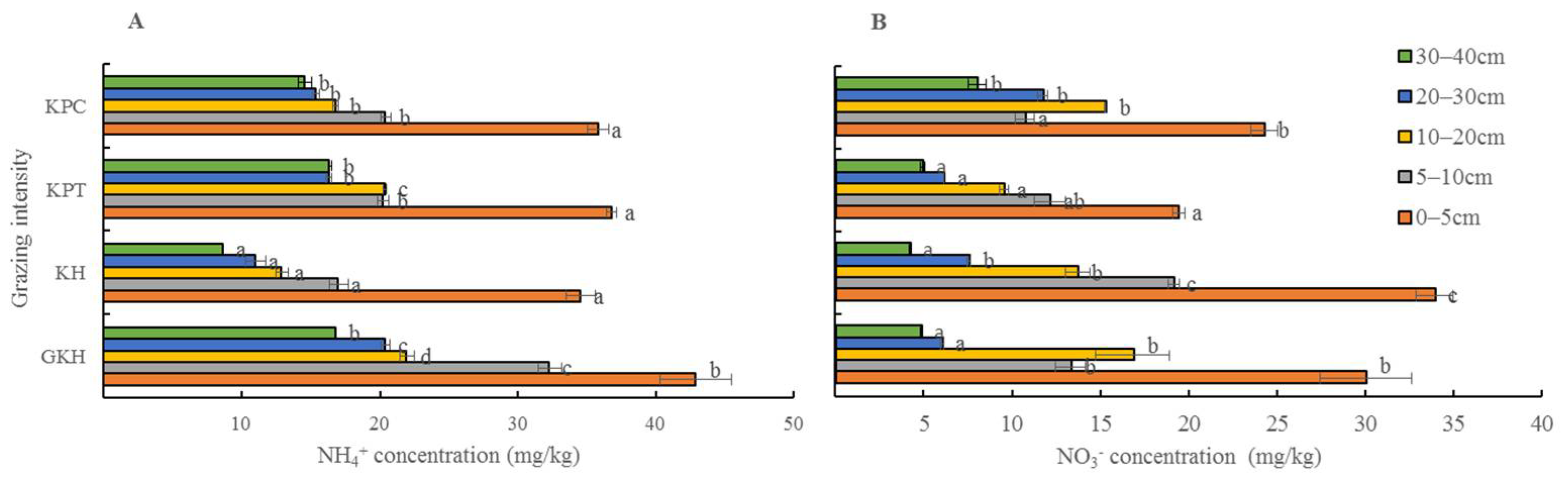
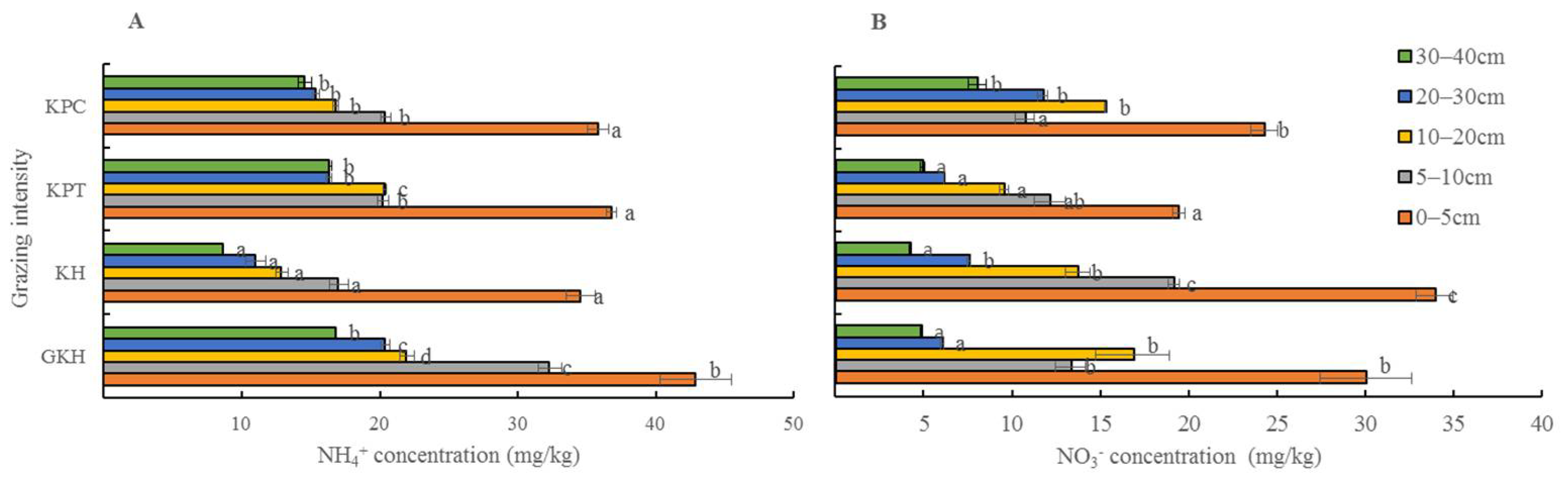
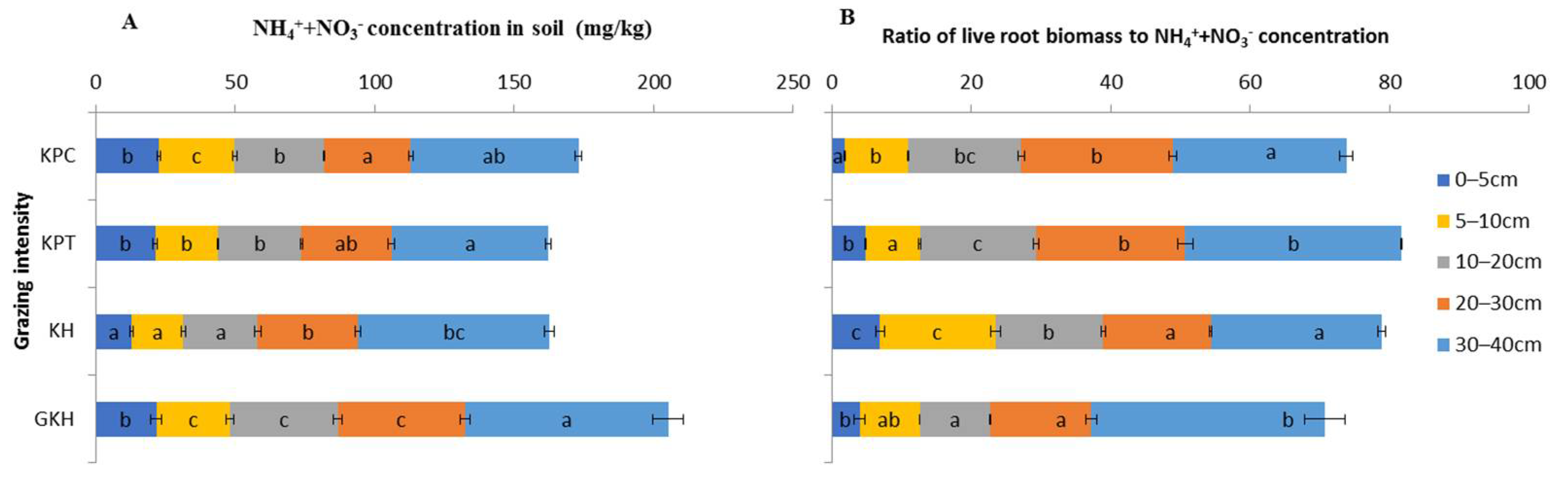
3.3.3. Concentration of Inorganic N in Soil
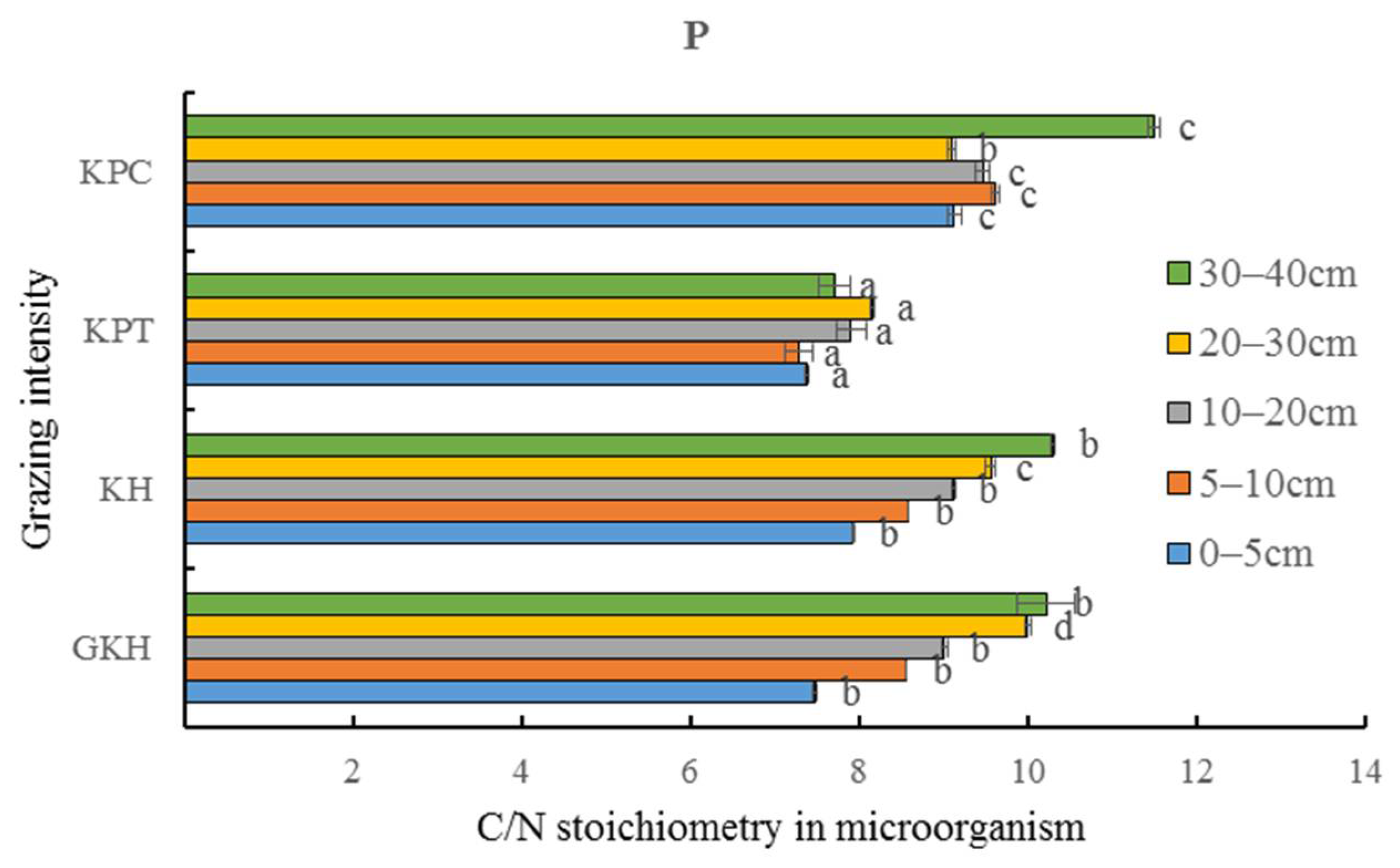
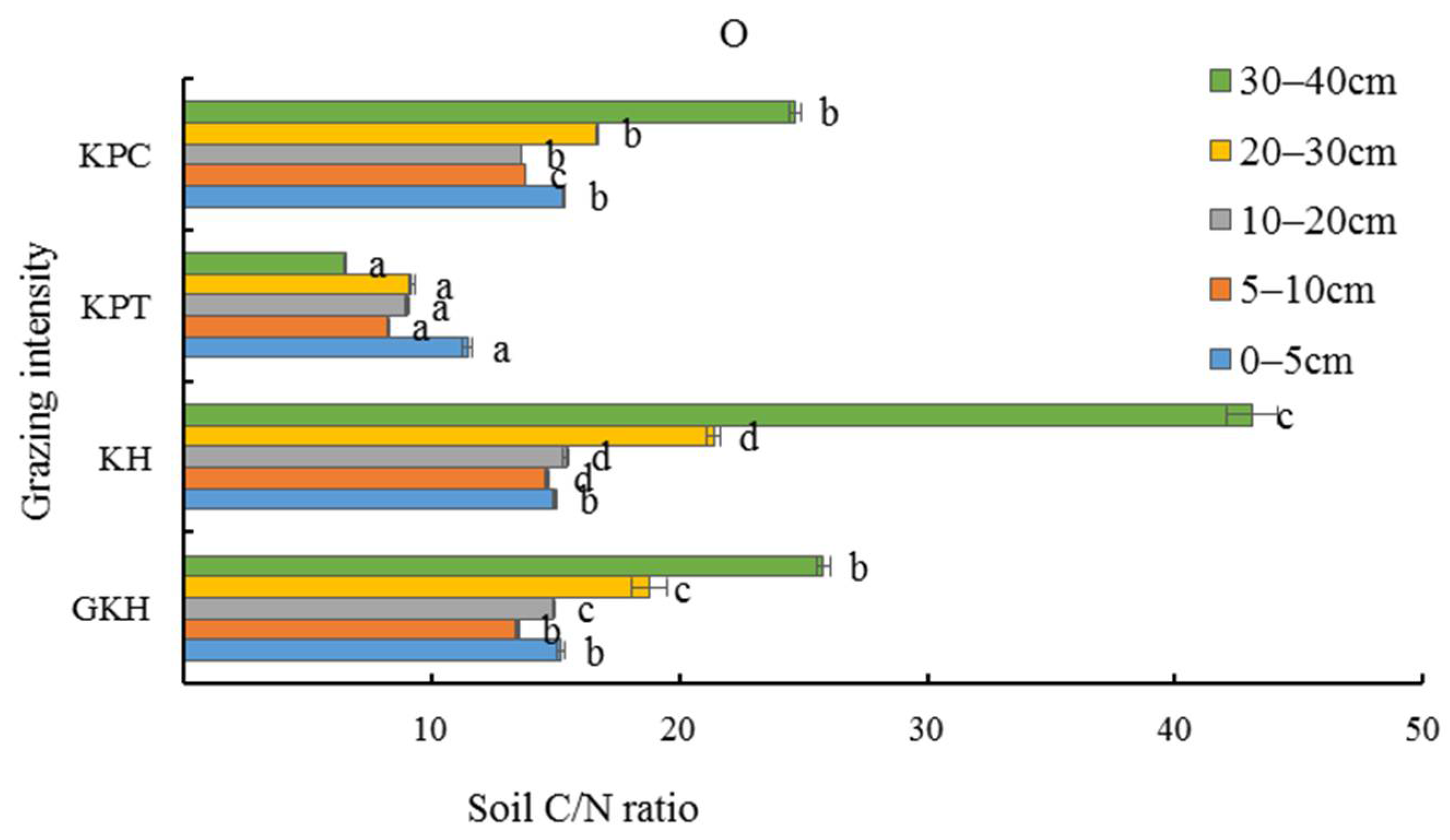
3.3.4. C/N Stoichiometry in Soil
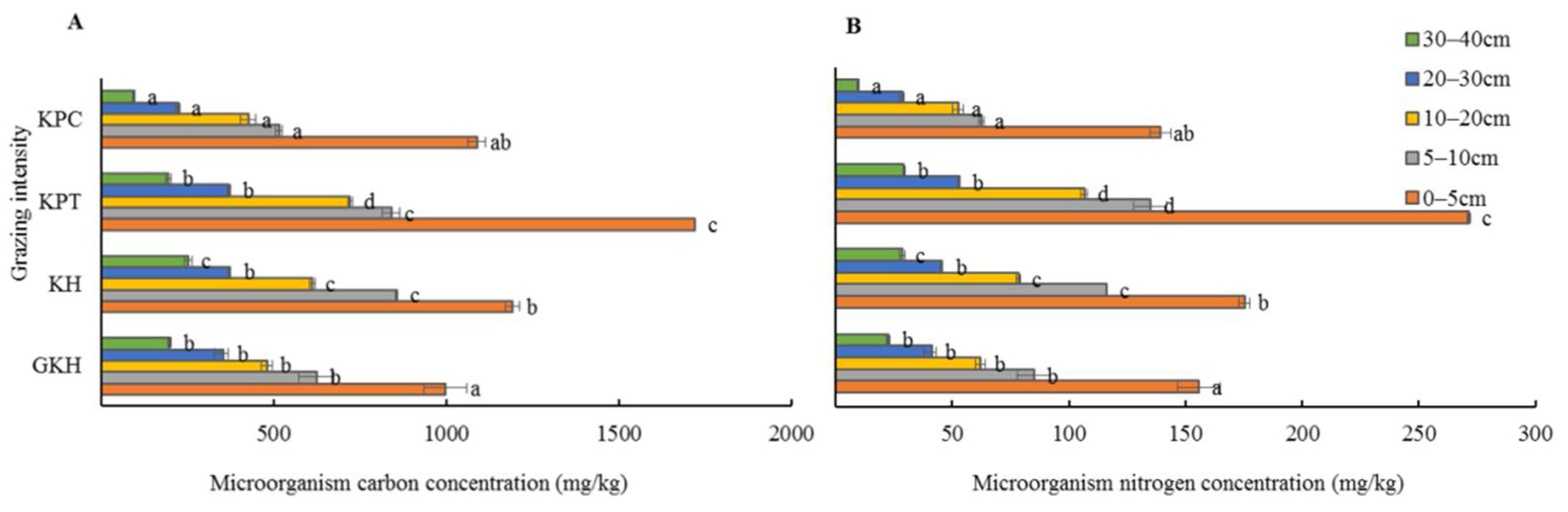
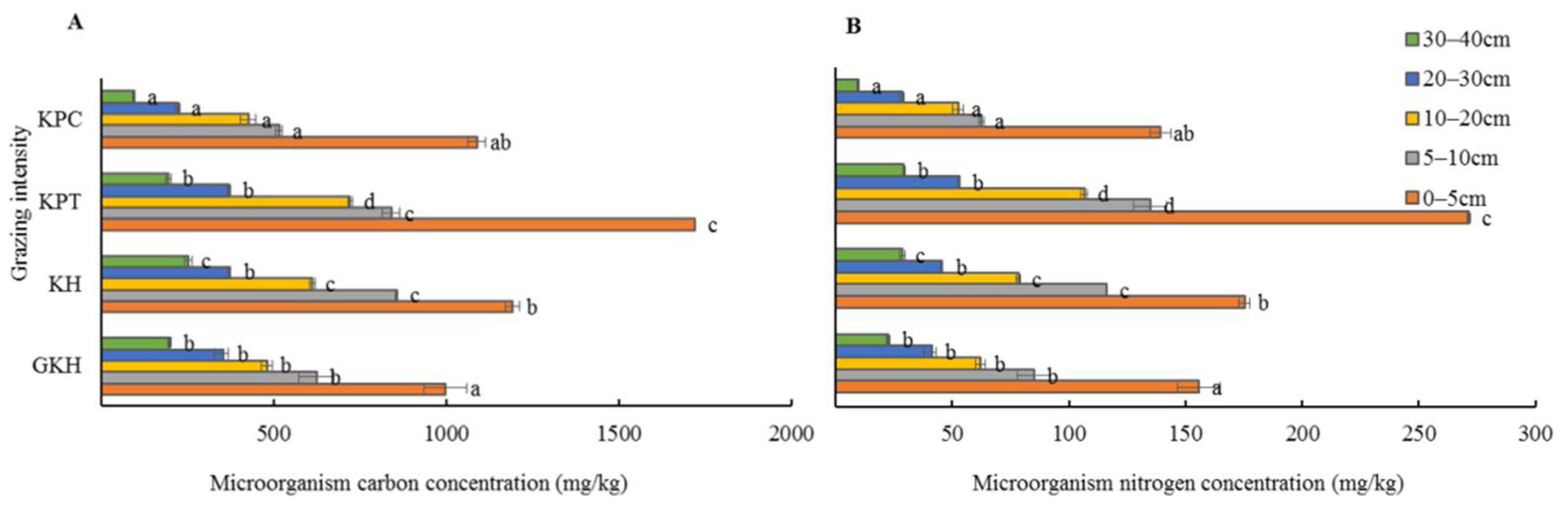
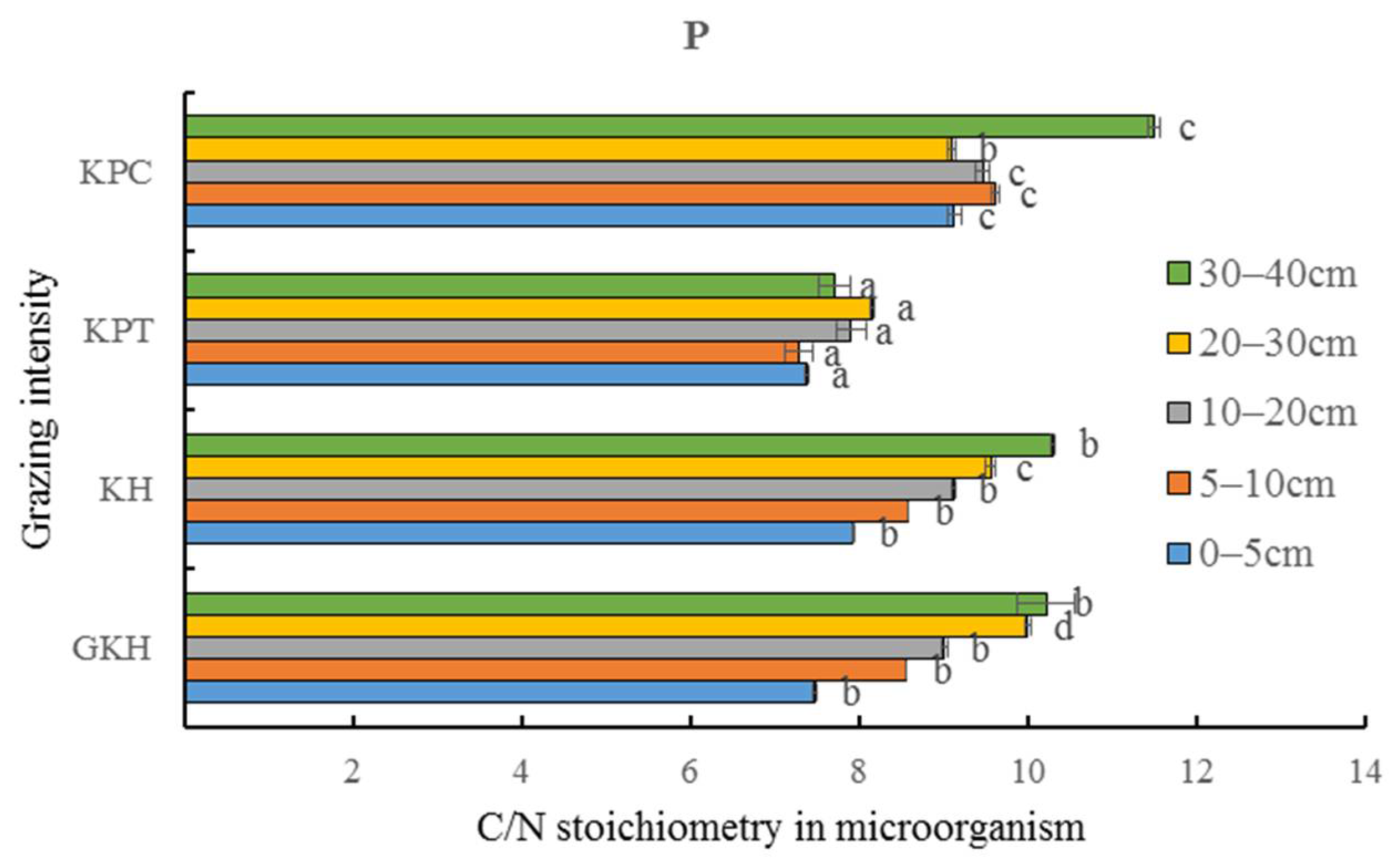
3.4. Quantity of C and N in Soil Microorganisms
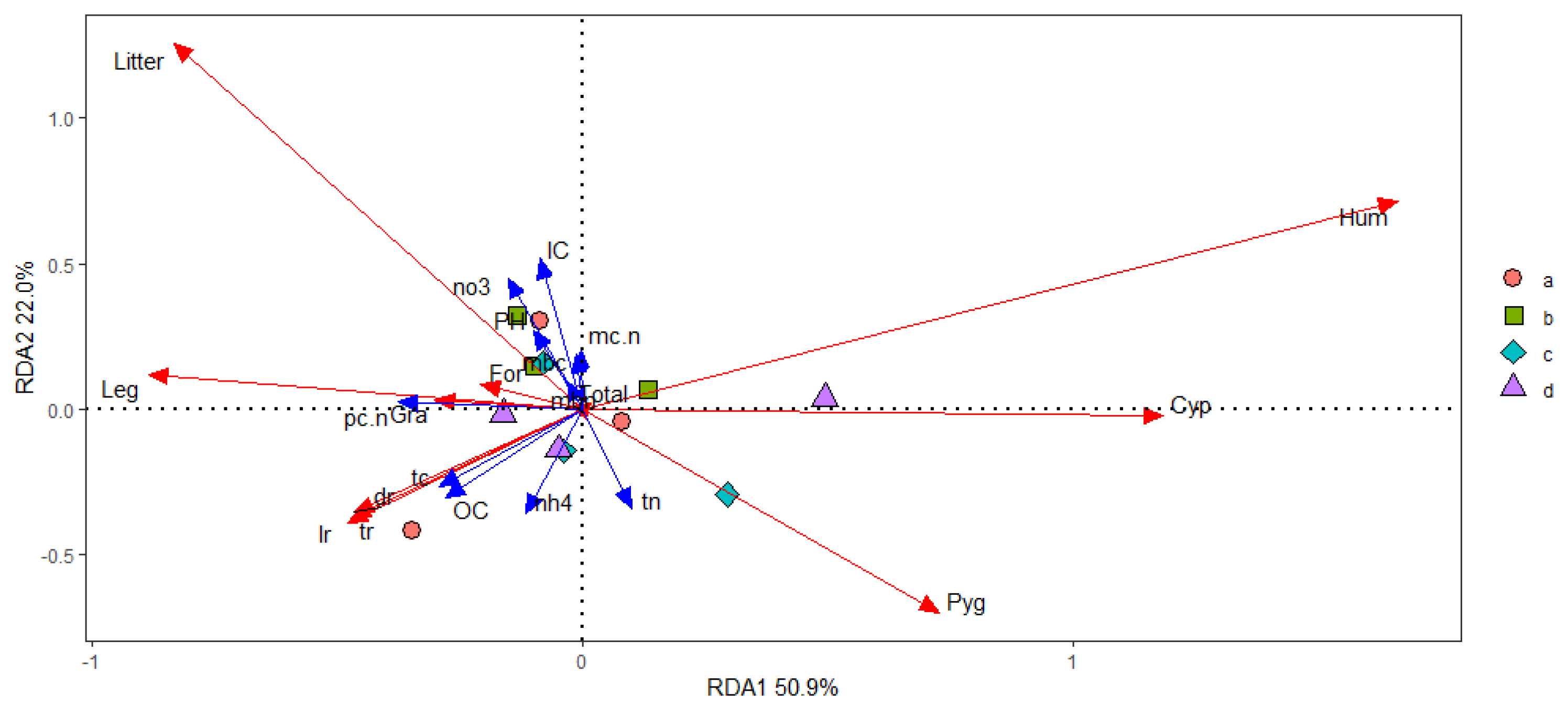
3.5. The Relationships between Plant Community Yield, Soil Nutrients and Soil Microbes Mediated by C and N under Overgrazing
4. Discussion
4.1. Overgrazing Initially Drove Ecosystem Degradation by Reducing Nutrient Concentrations and Biomass, Triggering a New Ecosystem Succession Process
4.2. Using Ratio of Gramineae to Cyperaceae or K. Humilis to K. pygmaea Could Indirectly Estimate the Ability of Ecosystem Maintain Stabilization
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.X.; Cao, G.M.; Zhou, D.W.; Hu, Q.W.; Zhao, X. The carbon storage and carbon cycle among the atmosphere, soil, vegetation and animal in the Kobresia humilis alpine meadow ecosystem. Acta Ecol. Sin. 2003, 23, 627–634. [Google Scholar]
- Li, F.Y.H.; Jäschke, Y.; Guo, K.; Wesche, K. Grasslands of China. Encycl. World’s Biomes 2020, 3, 773–784. [Google Scholar] [CrossRef]
- Cao, G.M.; Du, Y.G.; Wang, Q.L.; Wang, C.T.; Liang, D.Y. Character of passive active degradation process and its mechanism in Alpine Kobresia meadow. J. Mt. Sci. 2007, 25, 641–648. [Google Scholar]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef] [PubMed]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Lin, L. Response and Adaptation of Plant-Soil System of Alpine Meadows in Different Successional Stages to Grazing Intensity. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2017. [Google Scholar]
- Tadashi, F.; William, G.L. Alternative stable states, trait dispersion and ecological restoration. Oikos 2006, 113, 353–356. [Google Scholar]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspectives in Plant Ecology. Evol. Syst. 2012, 14, 33–47. [Google Scholar]
- Wang, C.Y.; Wei, M.; Wu, B.D.; Wang, S.H.; Jiang, K. Alpine grassland degradation reduced plant species diversity and stability of plant communities in the Northern Tibet Plateau. Acta Oecol. 2019, 98, 25–29. [Google Scholar] [CrossRef]
- Shang, Z.H.; Yang, S.; Wang, Y.; Shi, J.J.; Ding, L.M.; Long, R.J. Soil seed bank and its relation with above-ground vegetation along the degraded gradients of alpine meadow. Ecol. Eng. 2016, 90, 268–277. [Google Scholar] [CrossRef]
- White, L.M. Carbohydrate reserves of grass: A review. J. Range Manag. 1973, 26, 13–18. [Google Scholar] [CrossRef]
- Pigliucci, M. How organisms respond to environmental change: From phenotypes to molecules (and vice versa). Trends Ecol. Evol. 1996, 11, 168–173. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Ai, Z.M.; Liang, C.T.; Wang, G.L.; Liu, G.B.; Xue, S. How microbes cope with short-term N addition in a Pinus tabuliformis forest-ecological stoichiometry. Geoderma 2019, 337, 630–640. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Lin, L.; Li, Y.K.; Cui, Y.; Zhang, F.W.; Han, D.R.; Guo, X.W.; Li, J.; Cao, G.M. Relationship between dissolved nitrogen and plant biomass in Qing- Tibet Plateau typical vegetation types. J. Mt. Sci. 2012, 30, 721–727. [Google Scholar]
- Li, Y.; Wang, W.; Liu, Z.; Jiang, S. Grazing gradient versus restoration succession of Leymus chinensis grassland in Inner Mongolia. Restor. Ecol. 2008, 16, 572–583. [Google Scholar] [CrossRef]
- Nunes, J.S.; Araujo, A.S.F.; Nunes, L.A.P.L.; Lima, L.M.; Carneiro, R.F.V.; Salviano, A.A.C.; Tsai, S.M. Impact of land degradation on soil microbial biomass and activity in Northeast Brazil. Pedosphere 2012, 22, 88–95. [Google Scholar] [CrossRef]
- Lin, L.; Cao, G.M.; Li, Y.K.; Zhang, F.W.; Guo, X.W.; Han, D.R. Effects of human activities on organic carbon storage in the Kobresia humilis meadow ecosystem on the Tibetan Plateau. Acta Ecol. Sin. 2010, 30, 4012–4018. [Google Scholar]
- Korhonen, J.F.J.; Pihlatie, M.; Pumpanen, J.; Altonen, H.; Hari, P.; Levul, A.J.; Kieloaho, A.J.; Nikinmaa, E.; Vesala, T.; Ilvesniemi, H. Nitrogen balance of a boreal Scots pine forest. Biogeosciences 2013, 10, 1083–1095. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Ouyang, H.; Richter, A.; Wanek, W.; Cao, G.; Kuzyakov, Y. Spatio-temporal variations determine plant—microbe competition for inorganic nitrogen in an alpine meadow. J. Ecol. 2011, 99, 563–571. [Google Scholar] [CrossRef]
- Schulten, H.R.; Schnitzer, M. The chemistry of soil organic nitrogen: A review. Biol. Fertil. Soils 1998, 26, 1–15. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Dittmar, S.A.T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-knabner, I.; Lehmann, J.; Manning, D.A.C.; Nannipieri, P.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez, S.; Duarte, C.M.; Sandjensen, K. Patterns in decomposition rates among photosynthetic organisms: The importance of detritus C:N:P concentration. Oecologia 1993, 94, 457–471. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X.L. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Zhao, Y.; Ding, Y.; Hou, X.; Li, F.Y.; Yun, X. Effects of temperature and grazing on soil organic carbon storage in grasslands along the Eurasian steppe eastern transect. PLoS ONE 2017, 12, e0186980. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.L.; Liu, H.Y.; Zhao, F.J.; Xu, C.Y. Ecological stoichiometry of N:P:Si in China’s grasslands. Plant Soil 2014, 380, 165–179. [Google Scholar] [CrossRef]
- Yuan, W.Y.; Ma, X.W. The study in the policy system transition of using and property in pasture area. Product. Res. 2013, 11, 26–28. [Google Scholar]
- Wang, C.; Wang, G.; Wang, Y.; Rashad, R.; Ma, L.; Hu, L.; Luo, Y. Fire alters vegetation and soil microbial community in alpine meadow. Land Degrad. Dev. 2015, 27, 1379–1390. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Parry, L.C.; Vance, E.D.; Wu, J.; Harkness, D.D. Modeling the turnover of organic matter in soil. J. Sci. Food Agric. 1988, 45, 132–133. [Google Scholar]
- Sparling, G.P.; Shepherd, T.G.; Schipper, L.A. Topsoil characteristics of three contrasting New Zealand soils under four long-term land uses. N. Z. J. Agric. Res. 2000, 43, 569–583. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; Mcintyre, S.; Williams, N.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field- Methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Lin, L.; Cao, G.M.; Zhang, F.W.; Ke, X.; Li, Y.; Xu, X.; Li, Q.; Guo, X.; Fan, B.; Du, D. Spatial and temporal variations in available soil nitrogen—A case study in Kobresia Alpine Meadow in the Qinghai-Tibetan Plateau. J. Geosci. Environ. Prot. 2019, 7, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.M.; Long, R.J.; Zhang, F.W.; Li, Y.K.; Lin, L.; Guo, X.W.; Han, D.R.; Li, J. A method to estimate carbon storage potential in alpine Kobresia meadows on the Qinghai-Tibetan Plateau. Acta Ecol. Sin. 2010, 30, 6591–6597. [Google Scholar]
- Du, Y.G.; Cao, G.M.; Wang, Q.L.; Wang, C.T. Effect of grazing on surface character and soil physical property in alpine meadow. J. Mt. Sci. 2007, 25, 338–343. [Google Scholar]
- Neumann, G.; Bott, S.; Ohler, M.A.; Mock, H.P.; Lippmann, R.; Grosch, R.; Smalla, K. Root exudation and root development of lettuce (Lactuca sativa L. cv. Tizian) as affected by different soils. Front. Microbiol. 2014, 5, 2–6. [Google Scholar]
- Mehdi, C.; Carolyn, F.; Guo, J.W.; Cédric, L.; Meunier, J.S.; Wojciech, U.; Francisco, R.V. An operational framework for the advancement of a molecule-to-biosphere stoichiometry theory. Front. Mar. Sci. 2017, 4, 286–302. [Google Scholar]
- Hertenberger, G.; Zampach, P.; Bachmann, G. Plant species affect the concentration of free sugars and free amino acids in different types of soil. J. Plant Nutr. Soil Sci. 2002, 165, 557–565. [Google Scholar] [CrossRef]
- Baudoin, E.; Benizri, E.; Guckert, A. Impact of artificial root exudates on the bacterial community structure in bulk soil and maize rhizosphere. Soil Biol. Biochem. 2003, 35, 1183–1192. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press Inc.: San Diego, CA, USA, 1995. [Google Scholar]
- Cao, B.; Cao, Z.D.; Wang, L.M.; Sun, B.P.; Wang, D.C.; Zhang, X.B.; Liu, J.F. Research progress on soil-fixing in plant root system. Appl. Technol. Soil Water Conserv. 2009, 26–28. [Google Scholar]
- Li, Y.K.; Lin, L.; Zhang, F.W.; Liang, D.Y.; Wang, X.; Cao, G.M. Kobresia pygmaea community-disclimax of alpine meadow zonal vegetation in the pressure of grazing. J. Mt. Sci. 2010, 28, 257–265. [Google Scholar]
- Niu, Y.J. Study the Reason of Soil Crack Formation and Its Ecological Impact Assessment in Alpine Meadow. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2017. [Google Scholar]
- Mulder, C.; Elser, J.J. Soil acidity, ecological stoichiometry and allometric scaling in grassland food webs. Glob. Chang. Biol. 2009, 15, 2730–2738. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.S.; Andrew, J.A.; Finzi, A.C.; Matamala, R.; Schlesinger, D. Effects of free-air CO2 enrichment (FACE) on belowground process in a Pinus taeda forest. Ecol. Appl. 2000, 10, 437–448. [Google Scholar]
- Xu, P. Grassland Survey and Planning Science; Chinese Agricultural Press: Beijing, China, 1999. [Google Scholar]
- Li, X.L.; Perry, G.L.W.; Brierley, G.; Sun, H.Q.; Li, C.H.; Lu, G.X. Quantitative assessment of degradation classifications for degraded alpine meadows (heitutan), Sanjiangyuan, western China. Land Degrad. Dev. 2014, 25, 417–427. [Google Scholar] [CrossRef]
- Yu, Q.; Chen, Q.S.; Elser, J.J.; Cease, A.; He, N.P.; Wu, H.H.; Zhang, G.M.; Wu, J.G.; Bai, Y.F.; Han, X.G. Linking stoichiometric homeostasis with ecosystem structure, functioning, and stability. Ecol. Lett. 2010, 13, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Field, C.B.; Jackson, R.B. Does nitrogen constrain carbon cycling or does carbon input stimulate nitrogen cycling? Ecology 2006, 87, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y. Review: Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil Sci. 2002, 165, 382–396. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Succession Process Stage | Gramineae + Kobresia humilis Community | K. humilis Community | Thickening Mattic Epipedon in K. pygmaea Community | Cracking Mattic Epipedon in K. pygmaea Community | |
|---|---|---|---|---|---|
| Item | |||||
| Code | GKH | KH | KPT | KPC | |
| Location | 37°39.023′ N, 101° E 10.638′, 3230 m | 37°40.155′ N, 101° E 10.021′, 3241 m | 37°40.054′ N, 101° E 10.620′, 3239 m | 37°42.089′ N, 101° E 15.928′, 3278 m | |
| Average grazing rate (sheep per ha) | 3.75 ± 0.75 | 7.25 ± 0.25 | 8.50 ± 0.35 | 11.75 ± 0.95 | |
| Type of herbivore(s) | Sheep | Sheep | Sheep and yak | Sheep | |
| Ground surface feature | No cracks | No cracks | Coverage of crack area < 8% | Coverage of crack area 10–15% | |
| Mattic epipedon feature | Thickness < 5 cm | Thickness 5–6 cm | Thickness 7–8 cm | Thickness 8–9 cm | |
| Plant Succession Process Stage | Gramineae + Kobresia humilis Community | K. humilis Community | Thickening Mattic Epipedon in K. pygmaea Community | Cracking Mattic Epipedon in K. pygmaea Community | |
|---|---|---|---|---|---|
| Item | |||||
| Code | GKH | KH | KPT | KPC | |
| Dominant species in plant community | Stipa spp. Festuca spp. Helictotrichon tibeticum | K. humilis | K. pygmaea | K. pygmaea | |
| Subdominant species in plant community | K. humilis K. pygmaea Gentiana straminea | Stipa spp. Festuca spp. K. pygmaea | K. humilis Thermopsis lanceolata Oxytropis spp. | K. humilis Thermopsis lanceolata Oxytropis spp. | |
| Companion species | Saussurea pulchra Thermopsis lanceolata | Potentilla nivea Oxytropis spp. | Stipa spp. Festuca spp. | Stipa spp. Festuca spp. | |
| Total coverage | 85.7 ± 2.7% | 91.1 ± 0.6% | 71.8 ± 1.6% | 82.2 ± 0.6% | |
| Gramineae coverage | 36.4 ± 2.8% | 41.1 ± 0.2% | 20.3 ± 1.3% | 5.6 ± 1.4% | |
| Cyperaceae coverage | 37.8 ± 0.2% | 32.1 ± 0.8% | 32.2 ± 1.4% | 16.9 ± 0.5% | |
| Gramineae biomass ratio | 27.7 ± 3.2% | 26.8 ± 0.4% | 18.0 ± 0.2% | 22.8 ± 0.2% | |
| Cyperaceae biomass ratio | 19.8 ± 0.9% | 13.5 ± 0.1% | 13.9 ± 0.2% | 7.9 ± 0.3% | |
| Soil Layer | Variation Coefficient of Micro-C/N (%) | Variation Coefficient of Soil C/N (%) |
|---|---|---|
| 0–5 cm | 2.6 | 3.4 |
| 5–10 cm | 3.0 | 6.1 |
| 10–20 cm | 2.0 | 5.8 |
| 20–30 cm | 2.3 | 8.4 |
| 30–40 cm | 4.3 | 15.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; Cao, G.; Xu, X.; Li, C.; Fan, B.; Li, B.; Lan, Y.; Si, M.; Dai, L. Changes and Relationships between Components in the Plant-Soil System and the Dominant Plant Functional Groups in Alpine Kobresia Meadows Due to Overgrazing. Diversity 2022, 14, 183. https://doi.org/10.3390/d14030183
Lin L, Cao G, Xu X, Li C, Fan B, Li B, Lan Y, Si M, Dai L. Changes and Relationships between Components in the Plant-Soil System and the Dominant Plant Functional Groups in Alpine Kobresia Meadows Due to Overgrazing. Diversity. 2022; 14(3):183. https://doi.org/10.3390/d14030183
Chicago/Turabian StyleLin, Li, Guangmin Cao, Xingliang Xu, Chunli Li, Bo Fan, Bencuo Li, Yuting Lan, Mengke Si, and Licong Dai. 2022. "Changes and Relationships between Components in the Plant-Soil System and the Dominant Plant Functional Groups in Alpine Kobresia Meadows Due to Overgrazing" Diversity 14, no. 3: 183. https://doi.org/10.3390/d14030183
APA StyleLin, L., Cao, G., Xu, X., Li, C., Fan, B., Li, B., Lan, Y., Si, M., & Dai, L. (2022). Changes and Relationships between Components in the Plant-Soil System and the Dominant Plant Functional Groups in Alpine Kobresia Meadows Due to Overgrazing. Diversity, 14(3), 183. https://doi.org/10.3390/d14030183