
α-Glucosidase Inhibitory Activity and Anti-Adipogenic Effect of Compounds from Dendrobium delacourii

 , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
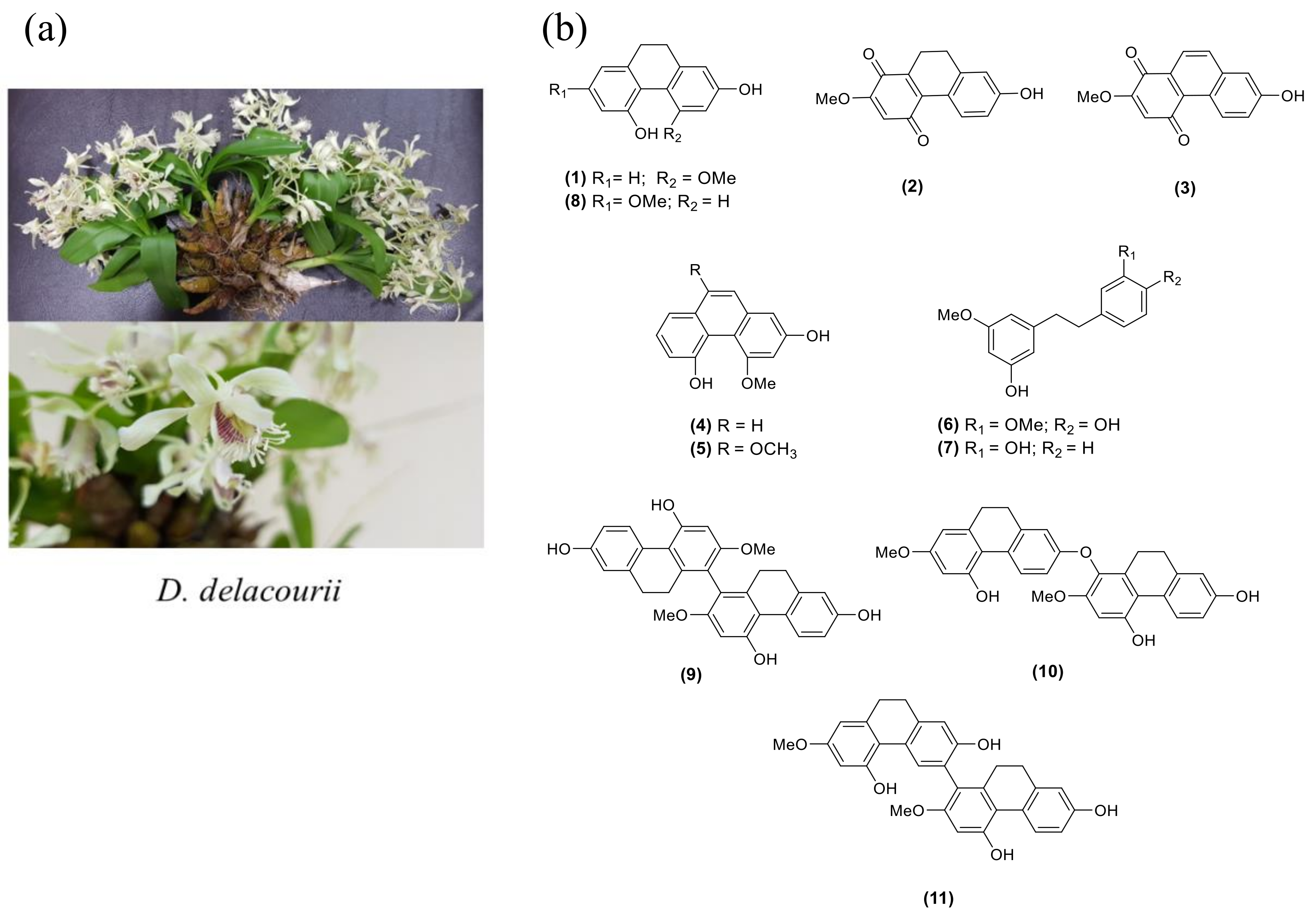
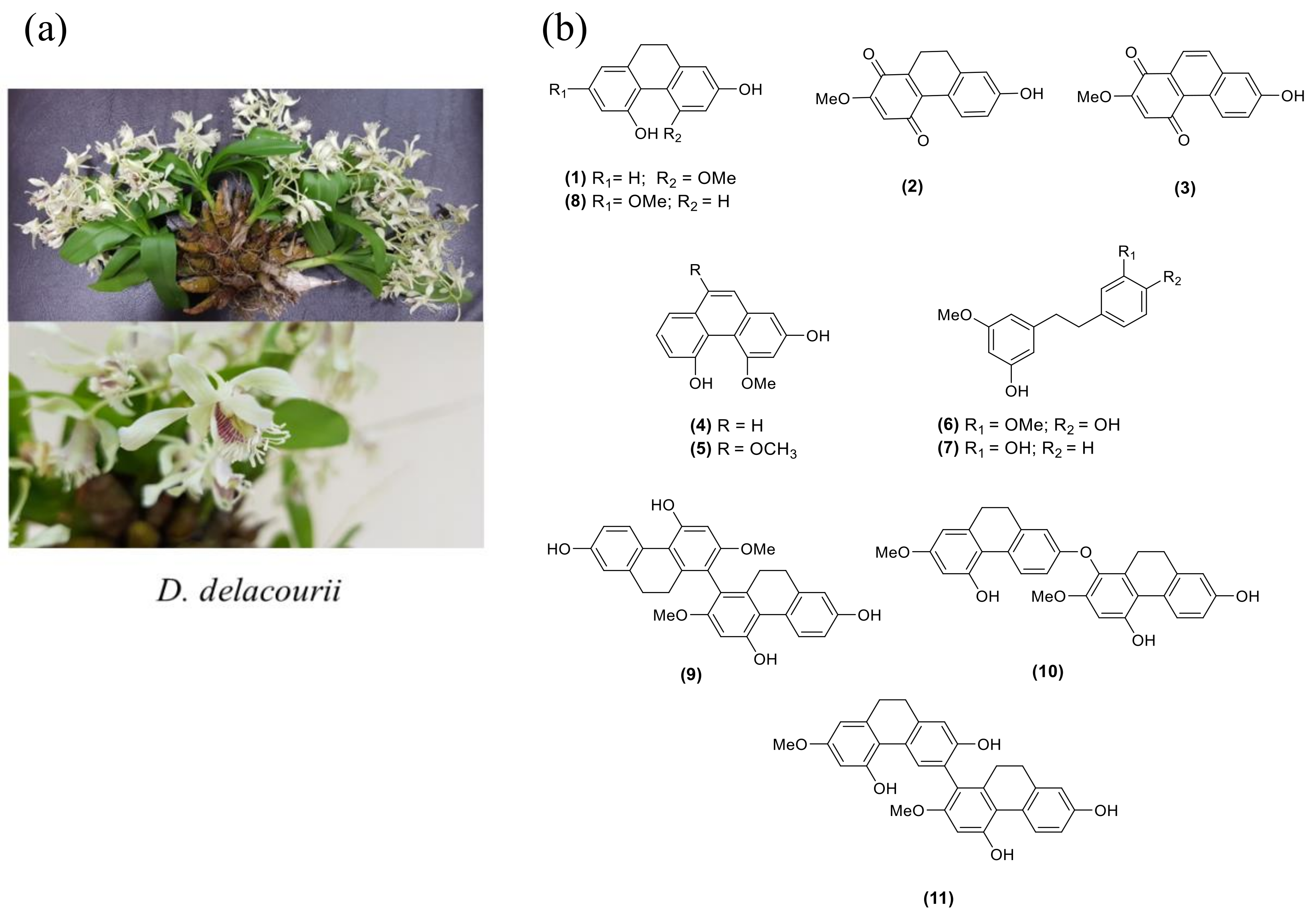
2.1. Structure Determination
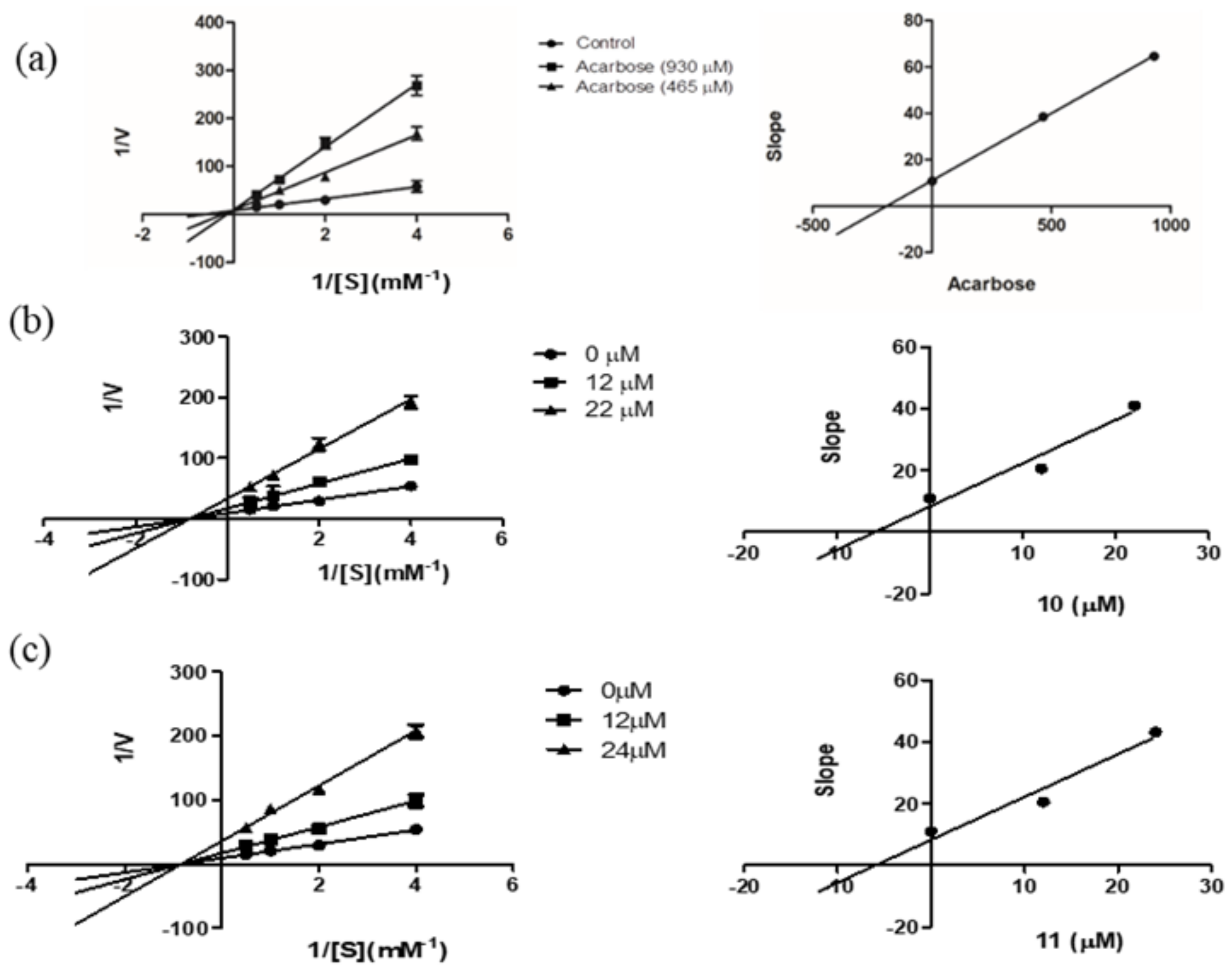
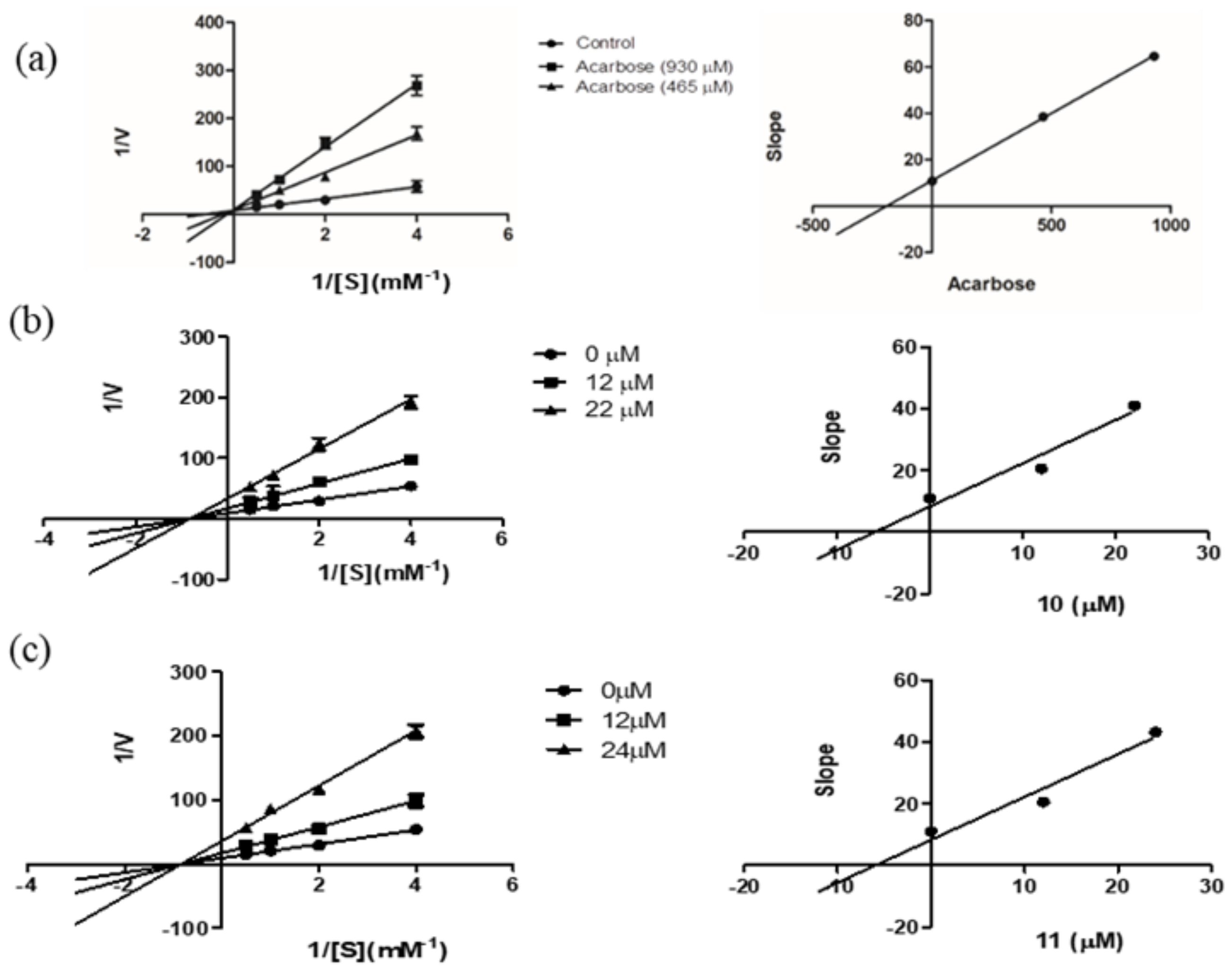
2.2. α-Glucosidase Inhibitory Activity
2.3. Anti-Adipogenic Activity
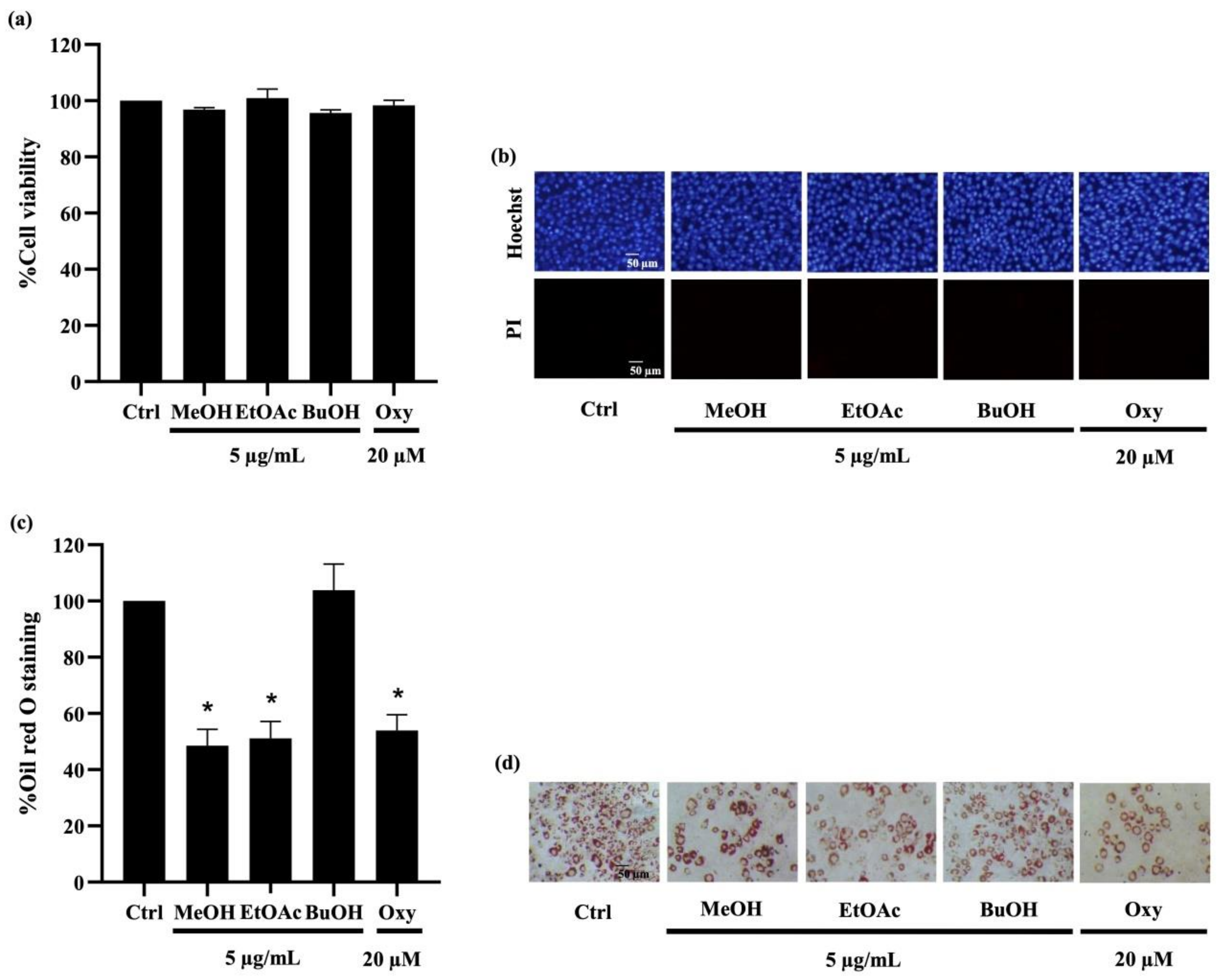
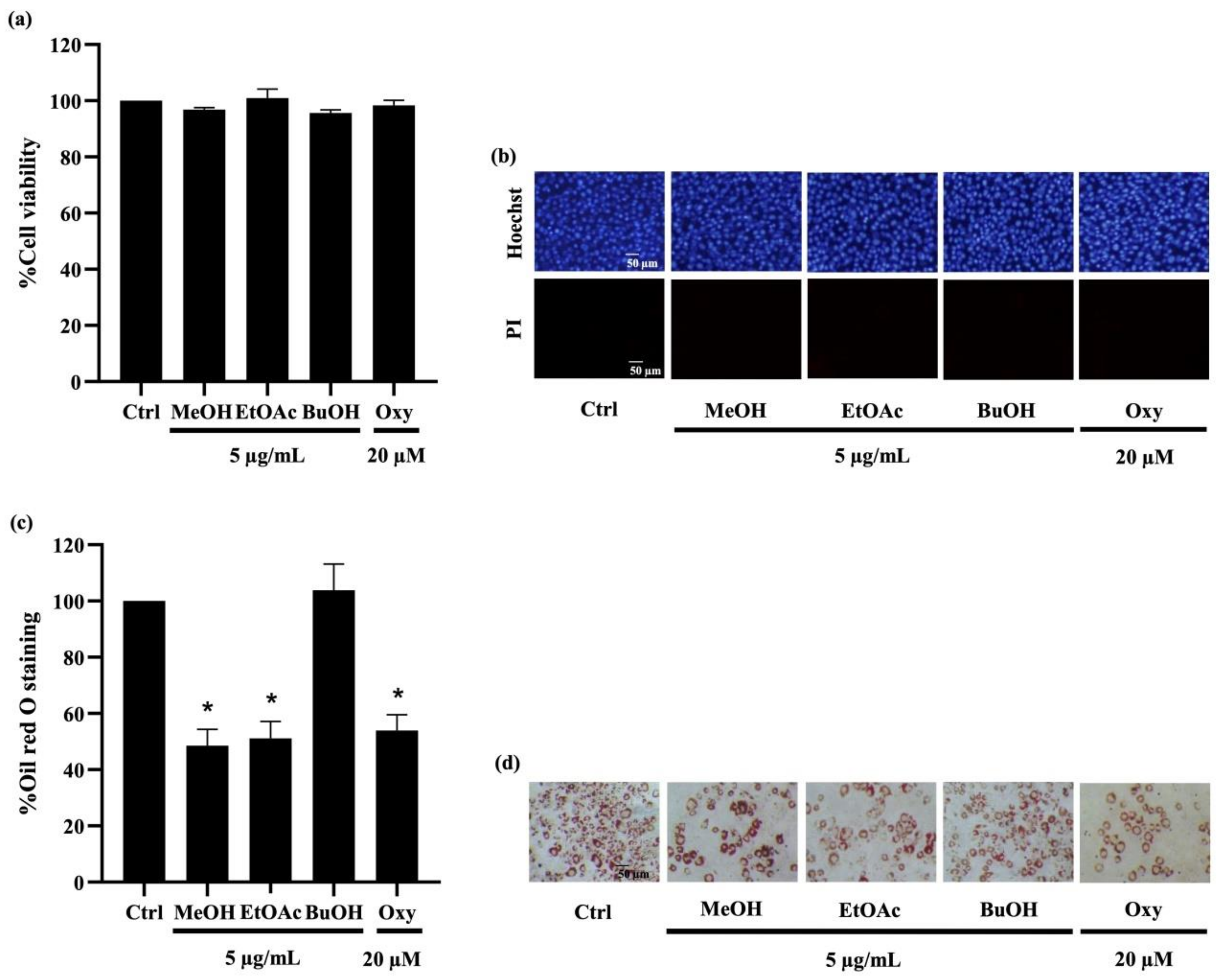
2.3.1. Ethyl Acetate Extracts from D. delacourii Attenuate Lipid Accumulation in Differentiated Adipocytes
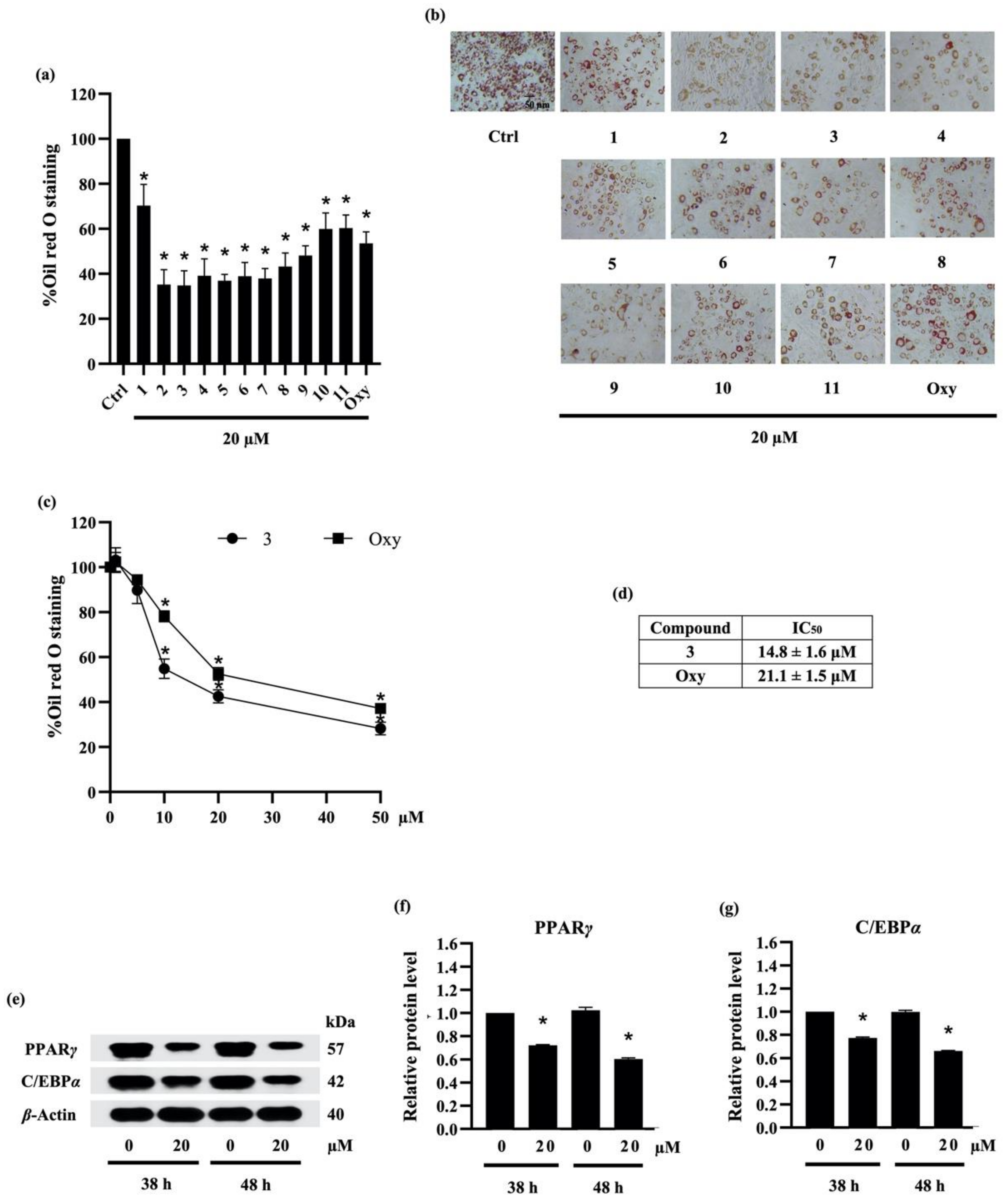
2.3.2. Screening for Anti-Adipogenic Activity of Compounds (1–11)
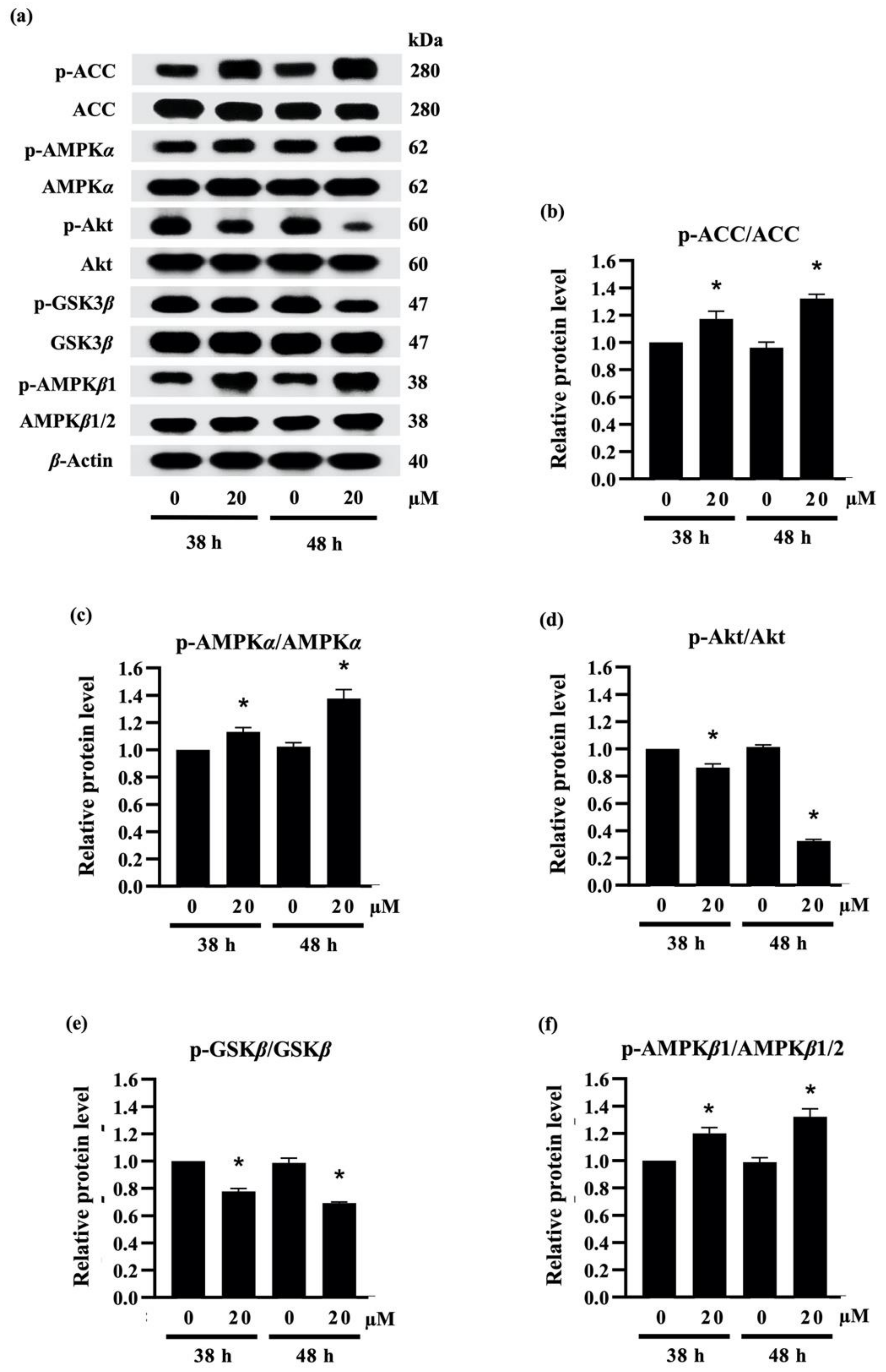
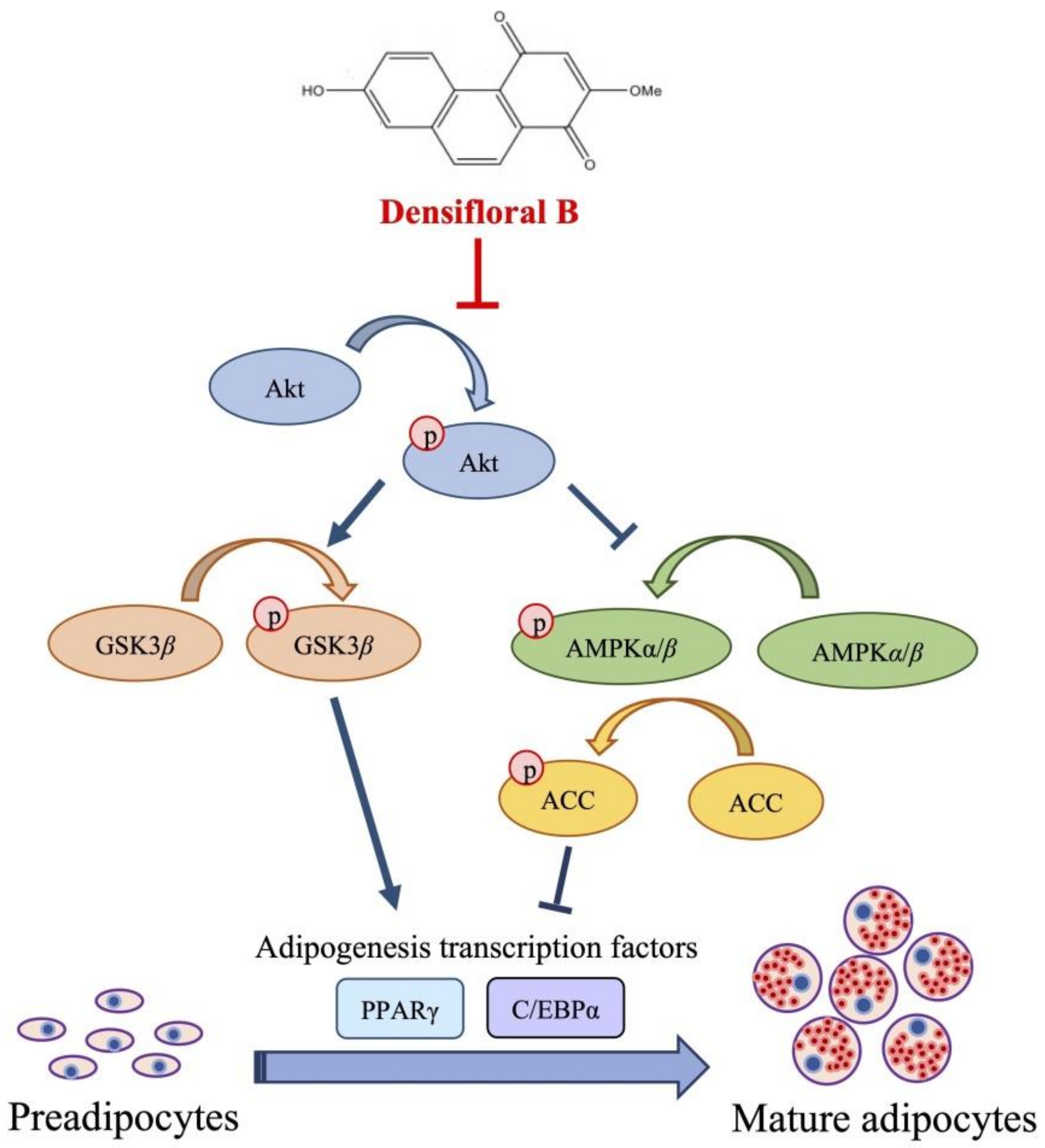
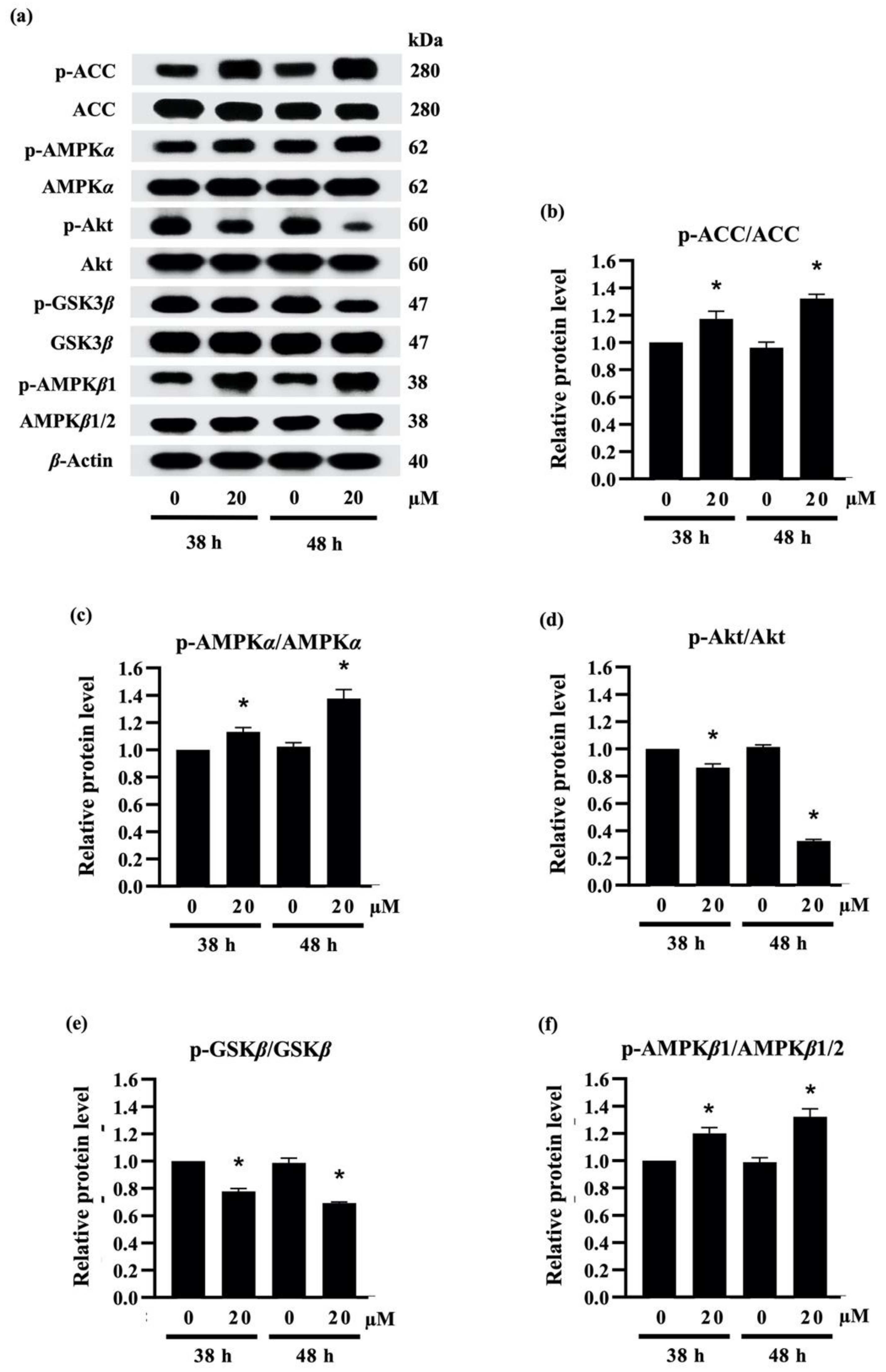
2.3.3. Densifloral B (3) Suppresses Adipocyte Differentiation-Related Proteins
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction and Isolation
3.4. Assay for α-Glucosidase Inhibitory Activity
3.5. Assay for Anti-Adipogenic Activity
3.5.1. Cell Culture and Adipocyte Differentiation
3.5.2. Determination of Cytotoxicity
3.5.3. Quantification of Cellular Lipid Content Using Oil Red O Staining
3.5.4. Western Blot Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- WHO. Definition, Diagnosis and Classification of Diabetes Mellitus and Its Complication (Part 1); World Health Organization: Geneva, Swizerland, 1999; pp. 2–32. [Google Scholar]
- Bischoff, H. Pharmacology of α-glucosidase inhibition. Eur. J. Clin. Investig. 1994, 24, 3–10. [Google Scholar]
- Peng, X.; Zhang, G.; Liao, Y.; Gong, D. Inhibitory kinetics and mechanism of kaempferol on α-glucosidase. Food Chem. 2016, 190, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Nhiem, N.X.; Kiem, P.V.; Minh, C.V.; Ban, N.K.; Cuong, N.X.; Tung, N.H.; Ha, L.M.; Ha, D.T.; Tai, B.H.; Quang, T.H.; et al. α-Glucosidase inhibition properties of cucurbitane-type triterpene glycosides from the fruits of Momordica charantia. Chem. Pharm. Bull. 2010, 58, 720–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butala, M.A.; Kukkupuni, S.K.; Vishnuprasad, C.N. Ayurvedic anti-diabetic formulation Lodhrasavam inhibits alpha-amylase, alpha-glucosidase and suppresses adipogenic activity in vitro. J. Ayurveda. Integr. Med. 2017, 8, 145–151. [Google Scholar] [CrossRef]
- Abbasi, F.; Brown, B.W.; Lamendola, C.; McLaughlin, T.; Reaven, G.M. Relationship between obesity, insulin resistance, and coronary heart disease risk. J. Am. Coll. Cardiol. 2002, 40, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.Y.; Iris, M.Y.; Li, E.T.; Wang, M. Inhibitory effects of oxyresveratrol and cyanomaclurin on adipogenesis of 3T3-L1 cells. J. Funct. Foods 2015, 15, 207–216. [Google Scholar] [CrossRef]
- Klein, G.; Kim, J.; Himmeldirk, K.; Cao, Y.; Chen, X. Antidiabetes and anti-obesity activity of Lagerstroemia speciosa. Evid. Based Complement. Altern. Med. 2007, 4, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Hwang, J.T.; Park, I.J.; Shin, J.I.; Lee, Y.K.; Lee, S.K.; Baik, H.W.; Park, O.J. Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2005, 338, 694–699. [Google Scholar] [CrossRef]
- Lin, J.; Della-Fera, M.A.; Baile, C.A. Green tea polyphenol epigallocatechin gallate inhibits adipogenesis and induces apoptosis in 3T3-L1 adipocytes. Obes. Res. 2005, 13, 982–990. [Google Scholar] [CrossRef]
- Pinent, M.; Blay, M.; Blade, M.C.; Salvado, M.J.; Arola, L.; Ardevol, A. Grape seed-derived procyanidins have an antihyperglycemic effect in streptozotocin-induced diabetic rats and insulinomimetic activity in insulin-sensitive cell lines. Endocrinology 2004, 145, 4985–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daval, M.; Foufelle, F.; Ferré, P. Functions of AMP-activated protein kinase in adipose tissue. J. Physiol. 2006, 574, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Zang, M.; Guo, W. AMPK as a metabolic tumor suppressor: Control of metabolism and cell growth. Future Oncol. 2010, 6, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habinowski, S.A.; Witters, L.A. The effects of AICAR on adipocyte differentiation of 3T3-L1 cells. Biochem. Biophys. Res. Commun. 2001, 286, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Ruderman, N.B. Malonyl-CoA and AMP-activated protein kinase: An expanding partnership. Mol. Cell Biochem. 2003, 253, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lago, F.; Gómez, R.; Gómez-Reino, J.J.; Dieguez, C.; Gualillo, O. Adipokines as novel modulators of lipid metabolism. Trends Biochem. Sci. 2009, 34, 500–510. [Google Scholar] [CrossRef]
- Rizzatti, V.; Boschi, F.; Pedrotti, M.; Zoico, E.; Sbarbati, A.; Zamboni, M. Lipid droplets characterization in adipocyte differentiated 3T3-L1 cells: Size and optical density distribution. Eur. J. Histochem. 2013, 57, e24. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, S.; Cho, E.J. Flavonoids from Acer okamotoanum inhibit adipocyte differentiation and promote lipolysis in the 3T3-L1 cells. Molecules 2020, 25, 1920. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.S.; Park, H.J.; Woo, J.H.; Kim, M.K.; Koh, P.O.; Min, W.; Cho, J.H. Citrus aurantium flavonoids inhibit adipogenesis through the Akt signaling pathway in 3T3-L1 cells. BMC Complement. Altern. Med. 2012, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Yoon, B.K.; Park, H.; Seok, J.W.; Choi, H.; Yu, J.H.; Kim, J.W. Caffeine inhibits adipogenesis through modulation of mitotic clonal expansion and the AKT/GSK3β pathway in 3T3-L1 adipocytes. BMB Rep. 2016, 49, 111. [Google Scholar] [CrossRef] [Green Version]
- Stein, S.A.; Lamos, E.M.; Davis, S.N. A review of the efficacy and safety of oral antidiabetic drugs. Expert. Opin. Drug Saf. 2013, 12, 153–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.G.; Park, C.Y. Anti-obesity drugs: A review about their effects and safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Paudel, M.R.; Bhattarai, H.D.; Pant, B. Traditionally used medicinal Dendrobium: A promising source of active anticancer constituents. In Orchids Phytochemistry, Biology and Horticulture: Fundamentals and Applications; Springer: Cham, Switzerland, 2020; pp. 1–26. [Google Scholar]
- Xu, J.; Han, Q.B.; Li, S.L.; Chen, X.J.; Wang, X.N.; Zhao, Z.Z.; Chen, H.B. Chemistry, bioactivity and quality control of Dendrobium, a commonly used tonic herb in traditional Chinese medicine. Phytochem. Rev. 2013, 12, 341–367. [Google Scholar] [CrossRef]
- Cakova, V.; Bonte, F.; Lobstein, A. Dendrobium: Sources of active ingredients to treat age-related pathologies. Aging Dis. 2017, 8, 827–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Dang, P.P.; Zhao, Z.; Yuan, L.C.; Zhou, Z.H.; Wolf, D.; Luo, Y.B. An assessment of the Chinese medicinal Dendrobium industry: Supply, demand and sustainability. J. Ethnopharmacol. 2019, 229, 81–88. [Google Scholar] [CrossRef]
- Chuakul, W. Ethnomedical uses of Thai Orchidaceous plants. Mahidol J. Pharm. Sci. 2002, 29, 41–45. [Google Scholar]
- Fisch, M.H.; Flick, F.H.; Arditti, J. Structure and antifungal activity of hircinol, loroglossol and orchinol. Phytochemistry 1973, 12, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Majumder, P.L.; Sen, R.C. Structure of moscatin—A new phenanthrene derivative from the orchid Dendrobium moscatum. Indian J. Chem. 1987, 26B, 18–20. [Google Scholar]
- Fan, C.; Wang, W.; Wang, Y.; Qin, G.; Zhao, W. Chemical constituents from Dendrobium densiflorum. Phytochemistry 2001, 57, 1255–1258. [Google Scholar] [CrossRef]
- Ono, M.; Ito, Y.; Masuika, C.; Koga, H.; Nohara, T. Antioxidative constituents from Dendrobii Herba (stems of Dendrobium spp.). Food Sci. Technol. Int. 1995, 1, 115–120. [Google Scholar]
- Leong, Y.W.; Kang, C.C.; Harrison, L.J.; Powell, A.D. Phenanthrenes, dihydrophenanthrenes and bibenzyls from the orchid Bulbophyllum vaginatum. Phytochemistry 1997, 44, 157–165. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Yu, H.; Qing, C.; Zhang, Y.; Wang, L.; Liu, Y.; Wang, J. Cytotoxic phenolics from Bulbophyllum odoratissimum. Food Chem. 2008, 107, 169–173. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.; Chen, Y. A new (propylphenyl) bibenzyl derivative from Dendrobium williamsonii. Nat. Prod. Res. 2017, 32, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Wang, J.; Wang, N.L.; Kitanaka, S.; Yao, X.S. 9, 10-Dihydrophenanthrene derivatives from Pholidota yunnanensis and scavenging activity on DPPH free radical. J. Asian Nat. Prod. Res. 2007, 9, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Wang, J.; Wang, N.L.; Kitanaka, S.; Liu, H.W.; Yao, X.S. New stilbenoids from Pholidota yunnanensis and their inhibitory effects on nitric oxide production. Chem. Pharm. Bull. 2006, 54, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Chougale, A.D.; Ghadyale, V.A.; Panaskar, S.N.; Arvindekar, A.U. Alpha glucosidase inhibition by stem extract of Tinospora cordifolia. J. Enzyme Inhib. Med. Chem. 2009, 24, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Ghadyale, V.; Takalikar, S.; Haldavnekar, V.; Arvindekar, A. Effective control of postprandial glucose level through inhibition of intestinal alpha glucosidase by Cymbopogon martinii (Roxb.). Evid. Based Complement. Altern. Med. 2012, 2012, 372909. [Google Scholar] [CrossRef]
- Yang, M.H.; Chin, Y.W.; Chae, H.S.; Yoon, K.D.; Kim, J. Anti-adipogenic constituents from Dioscorea opposita in 3T3-L1 cells. Bio. Pharm. Bull. 2014, 37, 1683–1688. [Google Scholar] [CrossRef]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Rhee, D.K.; Kim, B.O.; Pyo, S. Inhibitory effect of sinigrin on adipocyte differentiation in 3T3-L1 cells: Involvement of AMPK and MAPK pathways. Biomed. Pharmacother. 2018, 102, 670–680. [Google Scholar] [CrossRef]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular mechanisms of adipogenesis: The anti-adipogenic role of AMP-activated protein kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Ding, Y.; Kim, H.S.; Jeon, Y.J.; Lee, S.H. Inhibition of adipogenesis by diphlorethohydroxycarmalol (DPHC) through AMPK activation in adipocytes. Mar. Drugs 2019, 17, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Yuan, G.; Zhang, J.; Guo, X. Liraglutide reduces lipogenetic signals in visceral adipose of db/db mice with AMPK activation and Akt suppression. Drug Des. Devel. Ther. 2015, 9, 1177–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yao, Z. Chronic over-nutrition and dysregulation of GSK3 in diseases. Nutr. Metab. 2016, 13, 49. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jope, R.S.; Bijur, G.N. Mood stabilizers, glycogen synthase kinase-3β and cell survival. Mol. Psychiatry 2002, 7, S35–S45. [Google Scholar] [CrossRef] [Green Version]
- Chatsumpun, N.; Sritularak, B.; Likhitwitayawuid, K. New biflavonoids with α-glucosidase and pancreatic lipase inhibitory activities from Boesenbergia rotunda. Molecules 2017, 22, 1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, M.; Fujimori, K. Antiadipogenic effect of dietary apigenin through activation of AMPK in 3T3-L1 cells. J. Agric. Food Chem. 2011, 59, 13346–13352. [Google Scholar] [CrossRef]
- Borah, A.K.; Kuri, P.R.; Singh, A.; Saha, S. Anti-adipogenic effect of Terminalia chebula fruit aqueous extract in 3T3-L1 preadipocytes. Pharmacogn. Mag. 2019, 15, 197–204. [Google Scholar]
- He, F. BCA (bicinchoninic acid) protein assay. Bio-protocol 2011, 1, e44. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) |
|---|---|
| Hircinol (1) | NA |
| Ephemeranthoquinone (2) | NA |
| Densifloral B (3) | NA |
| Moscatin (4) | 390.1 ± 9.8 |
| 4,9-Dimethoxy-2,5-phenanthrenediol (5) | NA |
| Gigantol (6) | 191.3 ± 6.8 |
| Batatasin III (7) | NA |
| Lusianthridin (8) | 195.4 ± 9.6 |
| 4,4′,7,7′-Tetrahydroxy-2,2′-dimethoxy-9,9′,10,10′ -tetrahydro-1,1′-biphenanthrene (9) | 18.4 ± 3.4 |
| Phoyunnanin E (10) | 8.9 ± 0.8 |
| Phoyunnanin C (11) | 12.6 ± 0.9 |
| Acarbose | 514.4 ± 9.2 |
| Inhibitors | Dose (μM) | Vmax ∆OD/min | Km (mM) | Ki (μM) |
|---|---|---|---|---|
| None | - | 0.10 | 1.22 | |
| 10 | 22 | 0.024 | 1.22 | 5.89 |
| 12 | 0.049 | 1.21 | ||
| 11 | 24 | 0.023 | 1.21 | 5.97 |
| 12 | 0.049 | 1.21 | ||
| Acarbose | 930 | 0.11 | 6.47 | 190.57 |
| 465 | 0.10 | 4.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thant, M.T.; Khine, H.E.E.; Nealiga, J.Q.L.; Chatsumpun, N.; Chaotham, C.; Sritularak, B.; Likhitwitayawuid, K. α-Glucosidase Inhibitory Activity and Anti-Adipogenic Effect of Compounds from Dendrobium delacourii. Molecules 2022, 27, 1156. https://doi.org/10.3390/molecules27041156
Thant MT, Khine HEE, Nealiga JQL, Chatsumpun N, Chaotham C, Sritularak B, Likhitwitayawuid K. α-Glucosidase Inhibitory Activity and Anti-Adipogenic Effect of Compounds from Dendrobium delacourii. Molecules. 2022; 27(4):1156. https://doi.org/10.3390/molecules27041156
Chicago/Turabian StyleThant, May Thazin, Hnin Ei Ei Khine, Justin Quiel Lasam Nealiga, Nutputsorn Chatsumpun, Chatchai Chaotham, Boonchoo Sritularak, and Kittisak Likhitwitayawuid. 2022. "α-Glucosidase Inhibitory Activity and Anti-Adipogenic Effect of Compounds from Dendrobium delacourii" Molecules 27, no. 4: 1156. https://doi.org/10.3390/molecules27041156
APA StyleThant, M. T., Khine, H. E. E., Nealiga, J. Q. L., Chatsumpun, N., Chaotham, C., Sritularak, B., & Likhitwitayawuid, K. (2022). α-Glucosidase Inhibitory Activity and Anti-Adipogenic Effect of Compounds from Dendrobium delacourii. Molecules, 27(4), 1156. https://doi.org/10.3390/molecules27041156