
Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
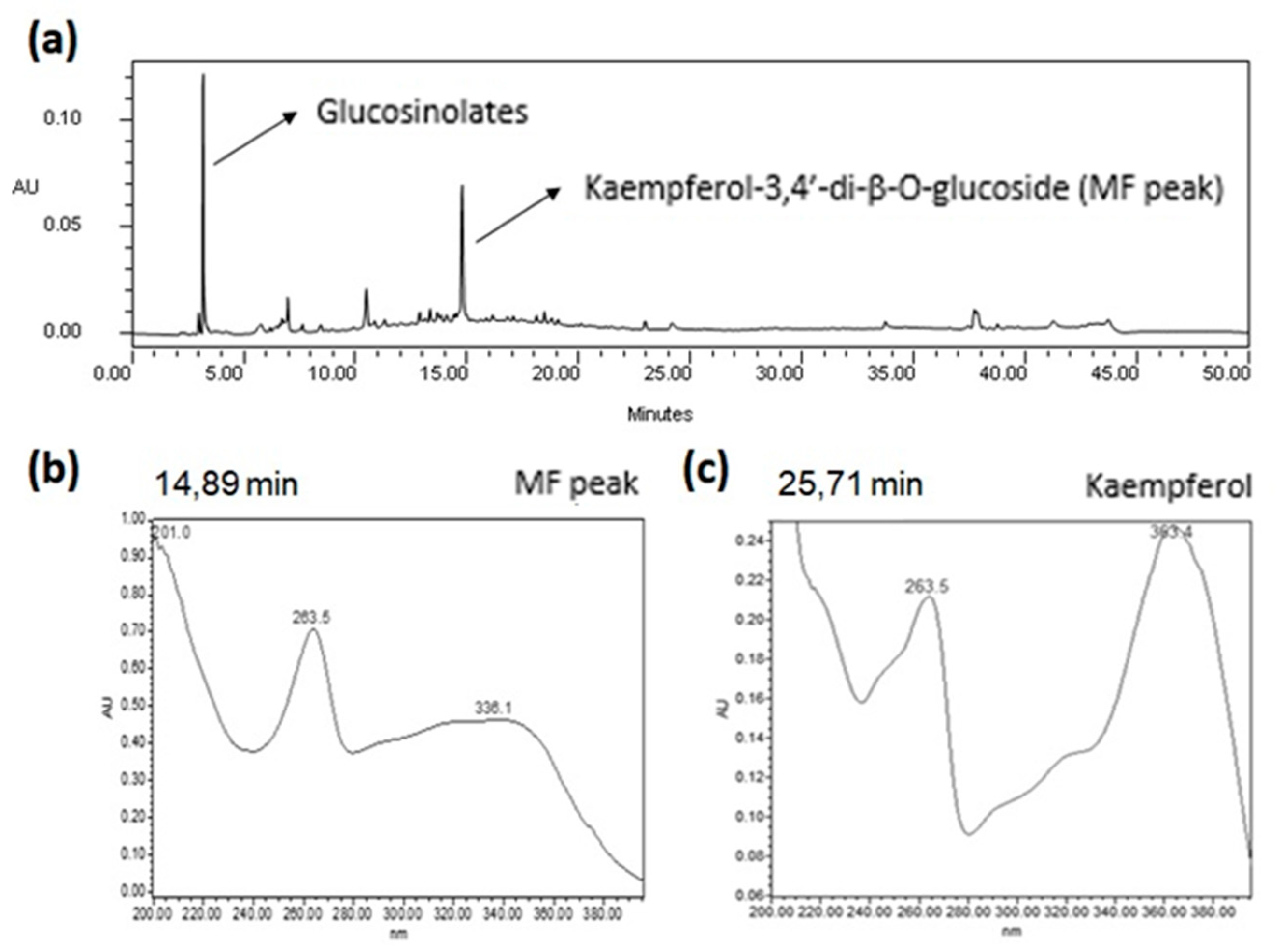
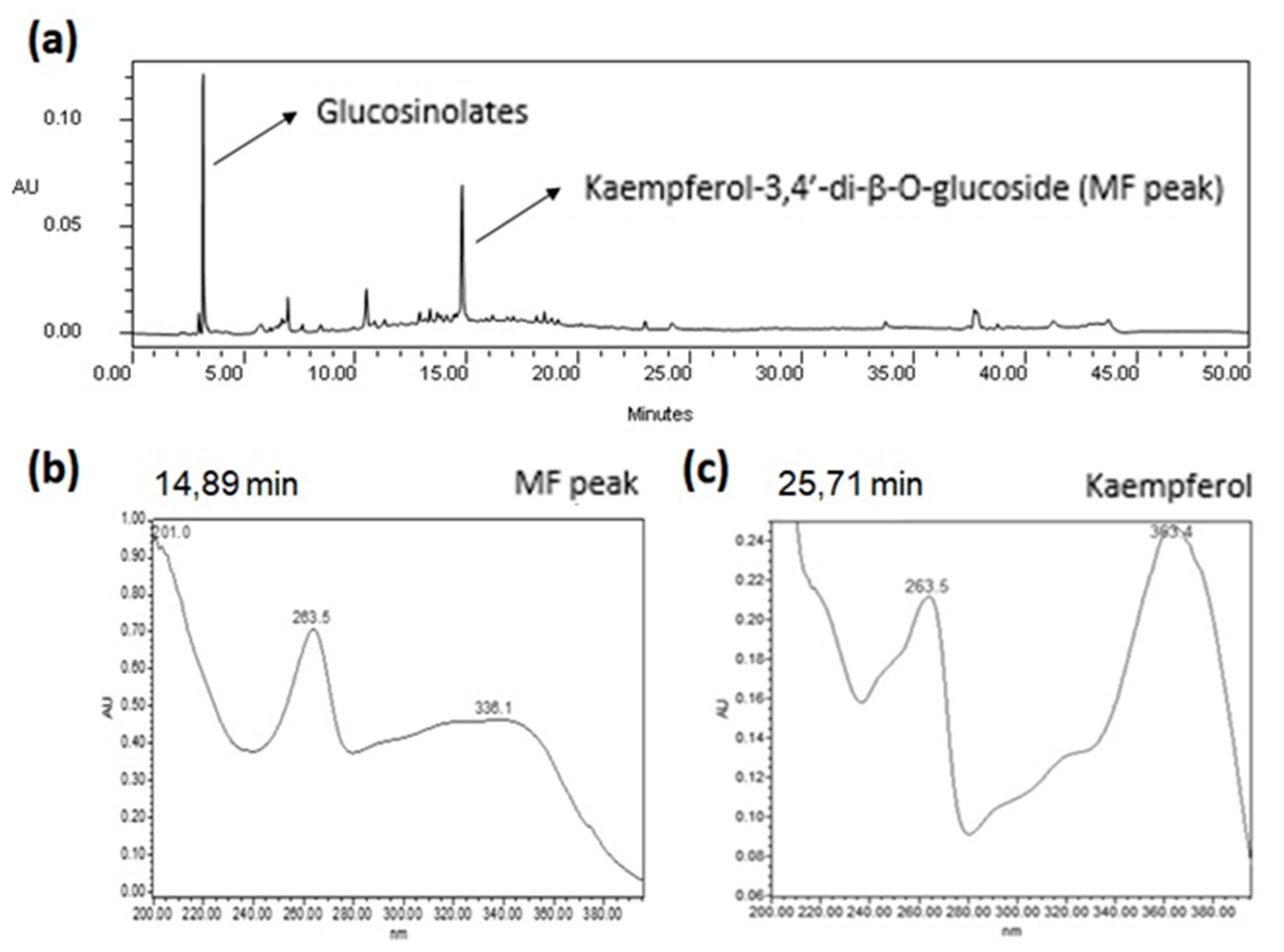
2.1. Qualitative HPLC Analyses of EE from E. sativa Leaves
2.1.1. Qualitative HPLC-DAD Analysis
2.1.2. Qualitative UHPLC-ESI-QTOF Analysis
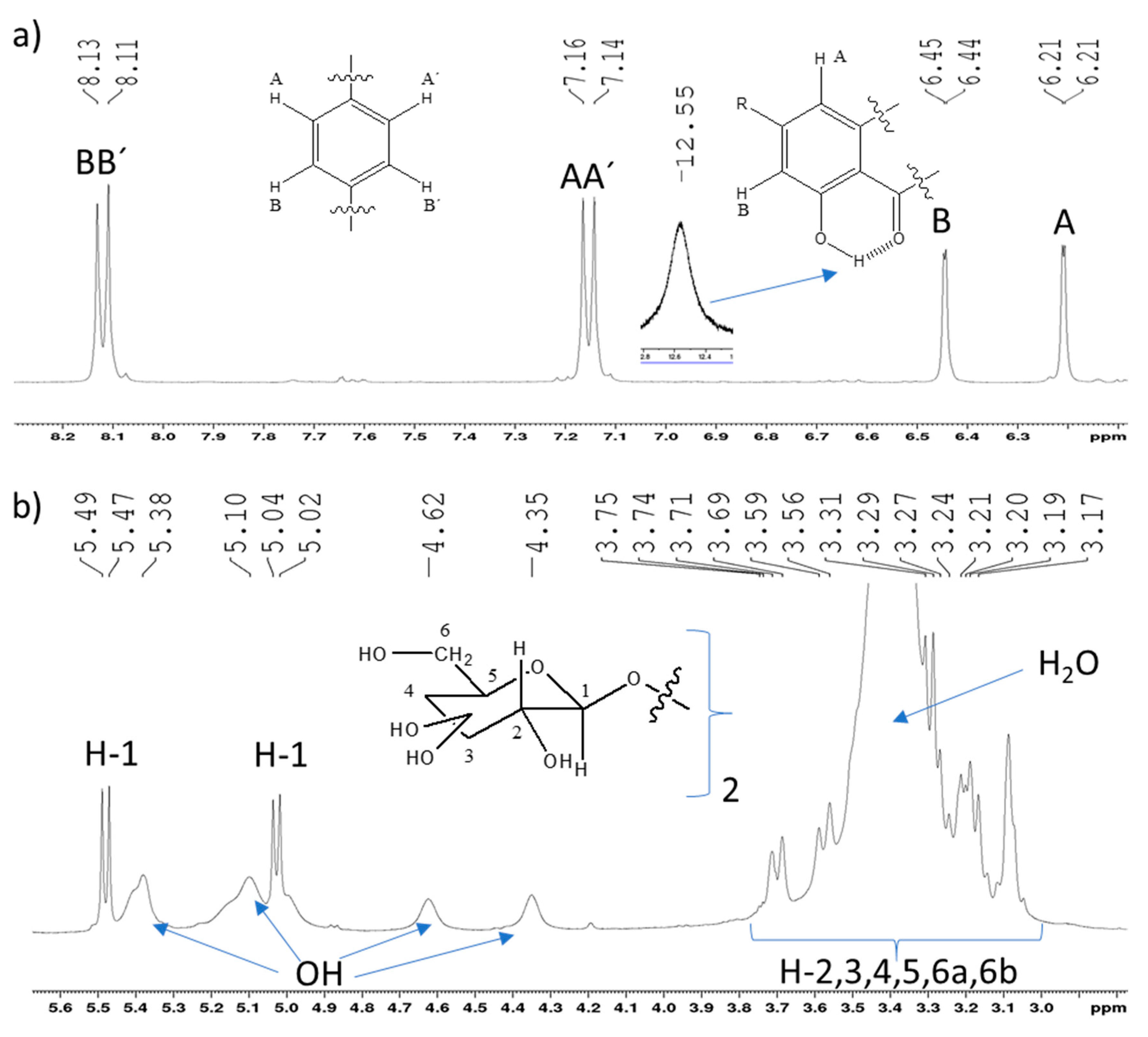
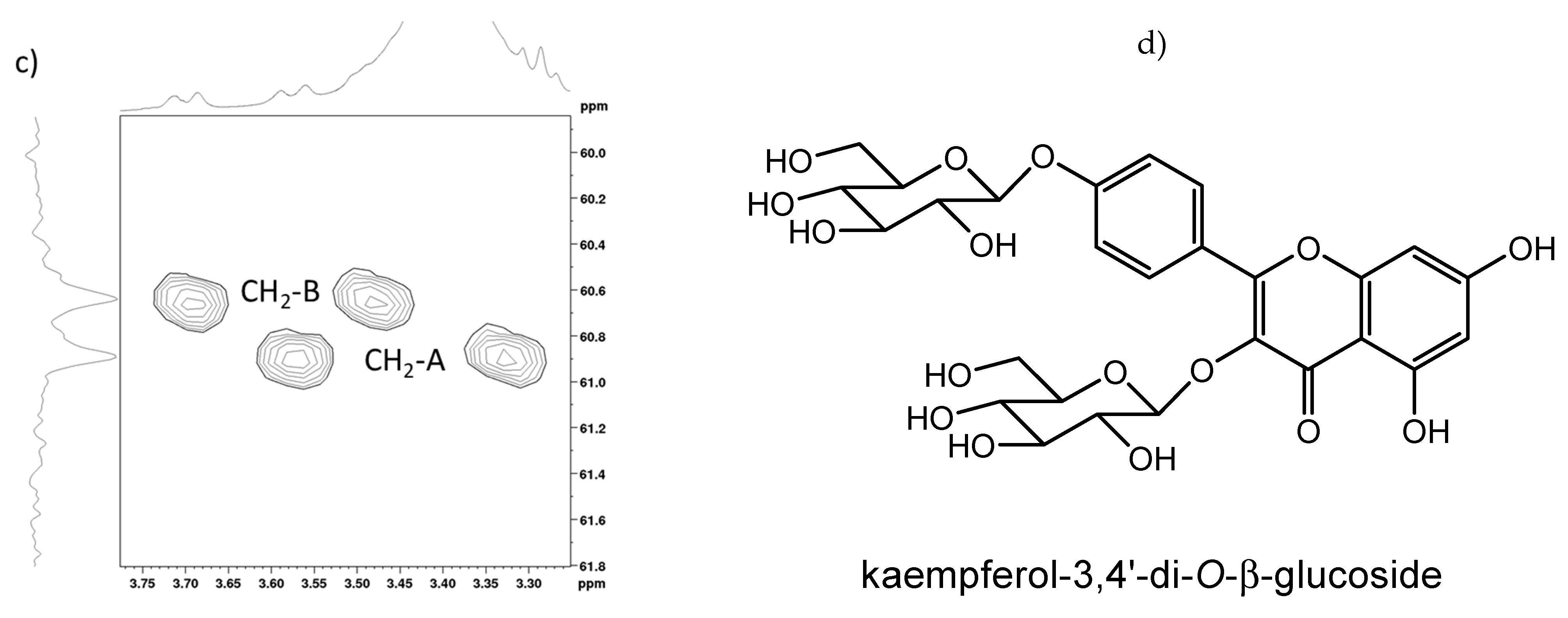
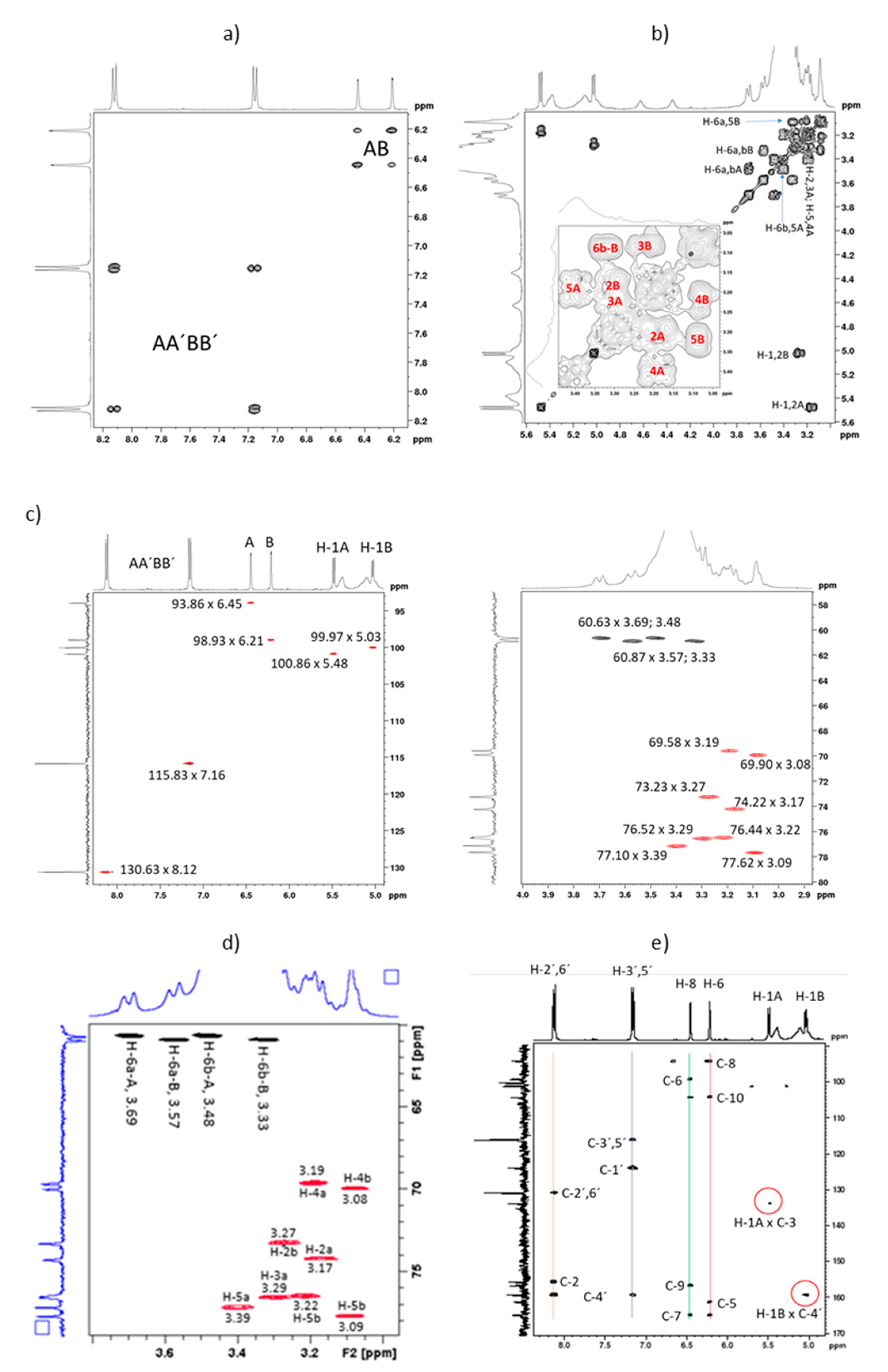
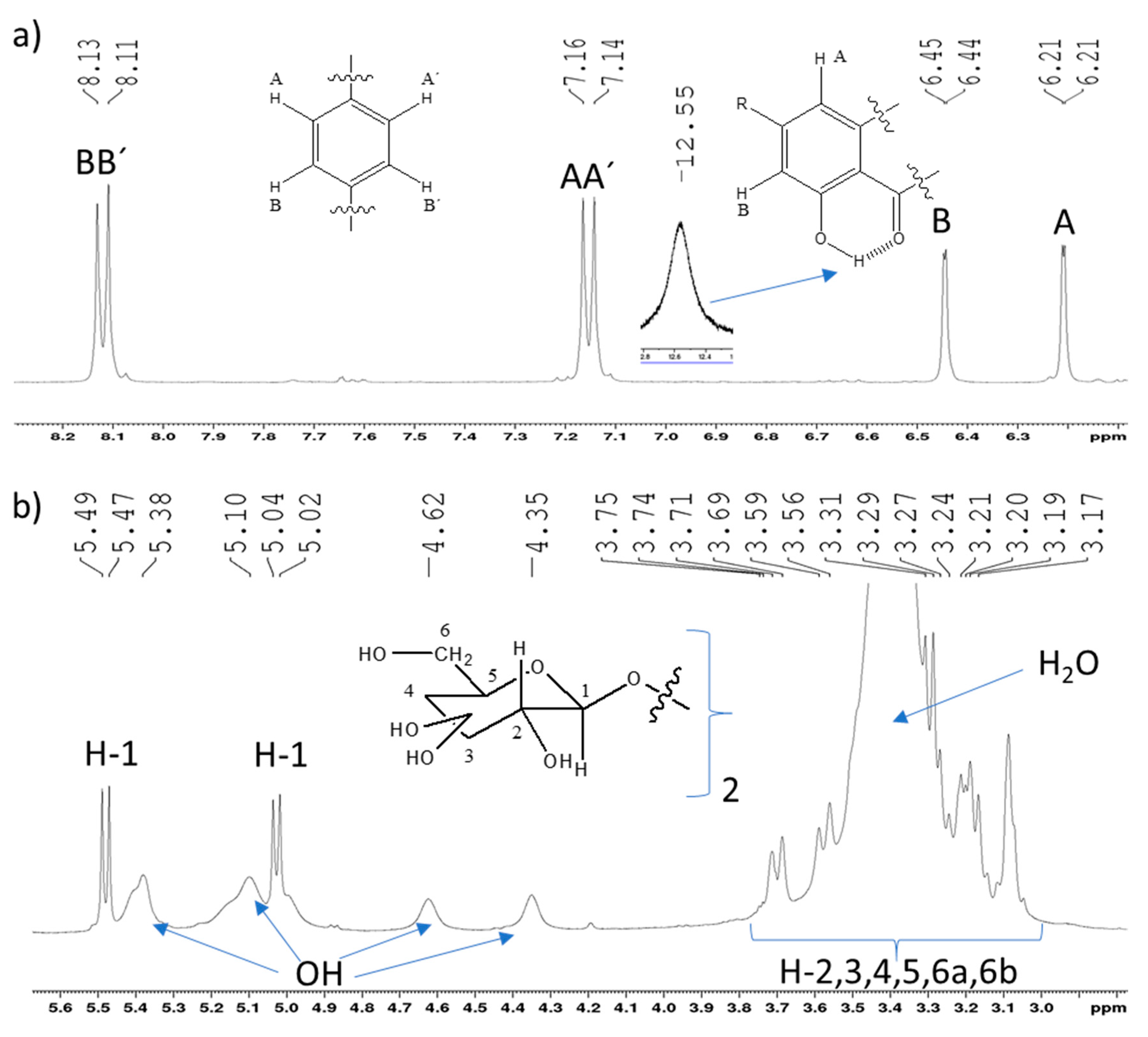
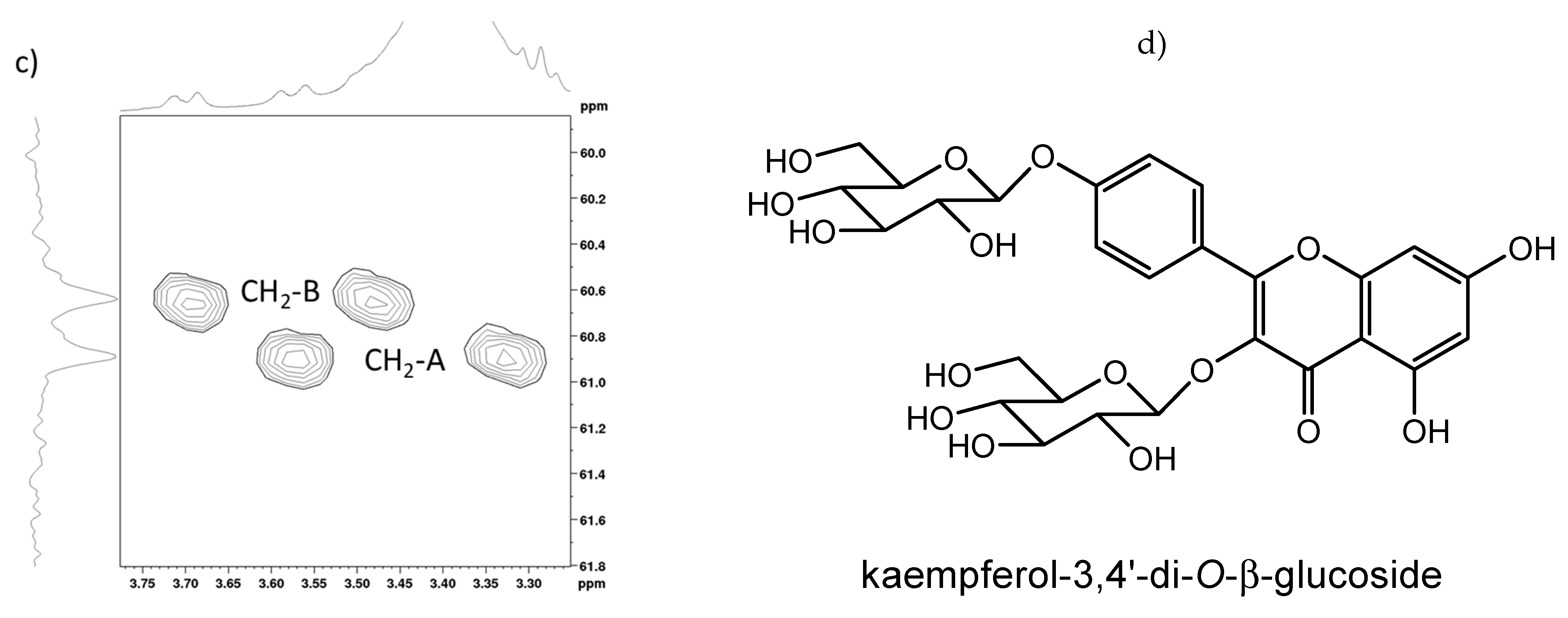
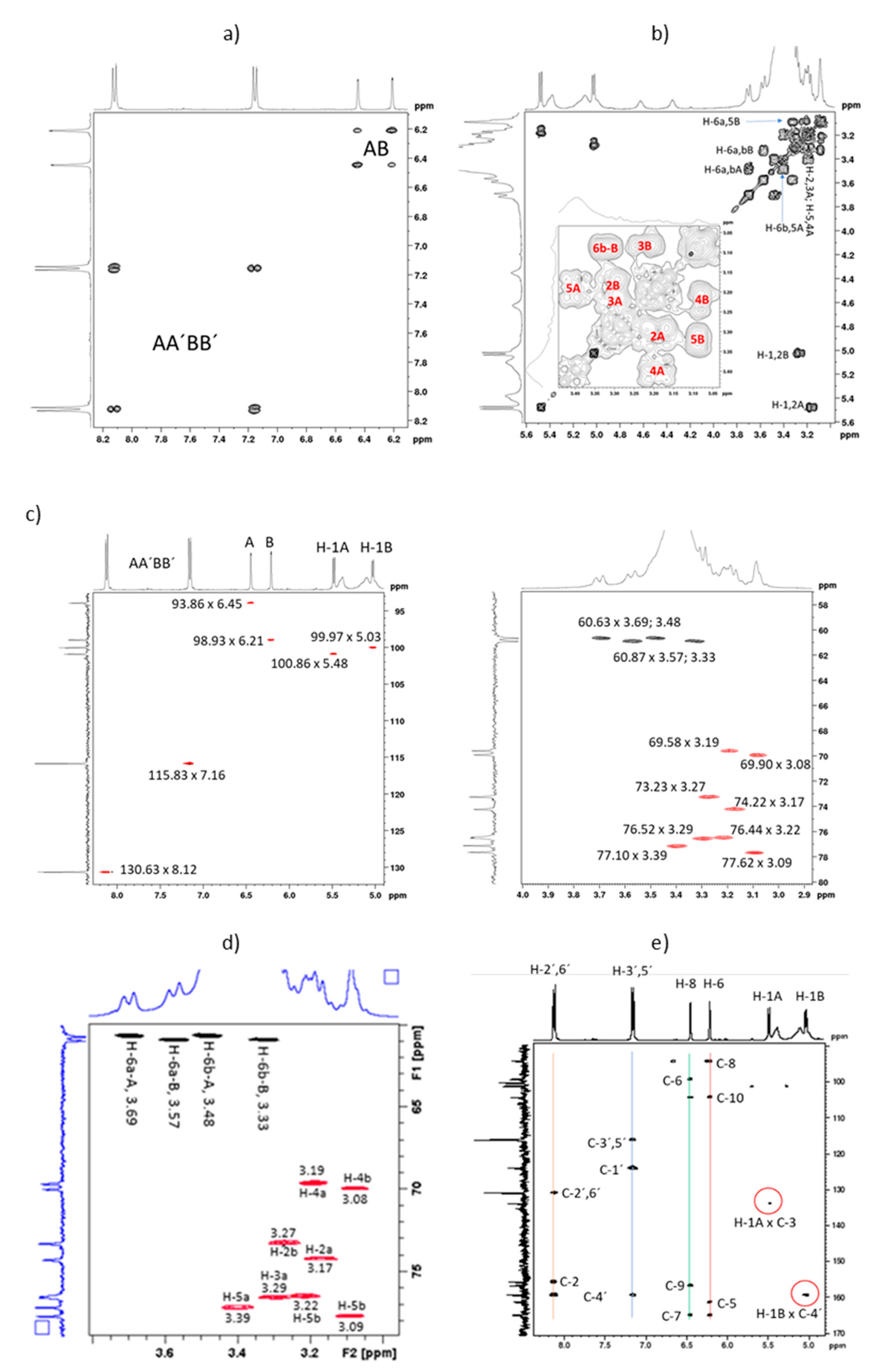
2.2. Full NMR Structural Characterisation of Kaempferol-3,4′-di-O-β-Glucoside
2.3. Quantification of the Major Flavonoid Kaempferol-3,4′-di-O-β-glucoside in the EE of E. sativa Leaves by the Validated HPLC-DAD Method
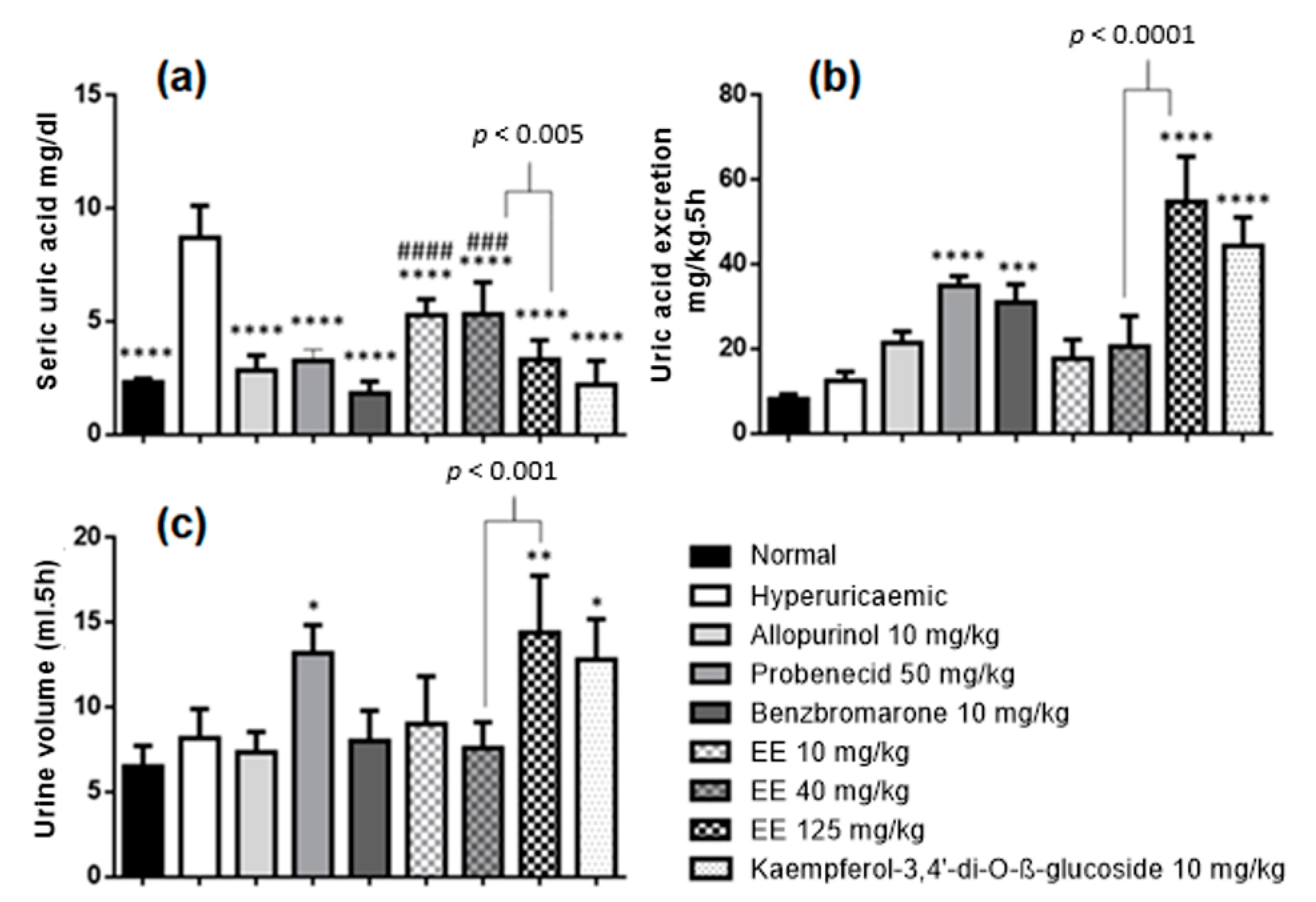
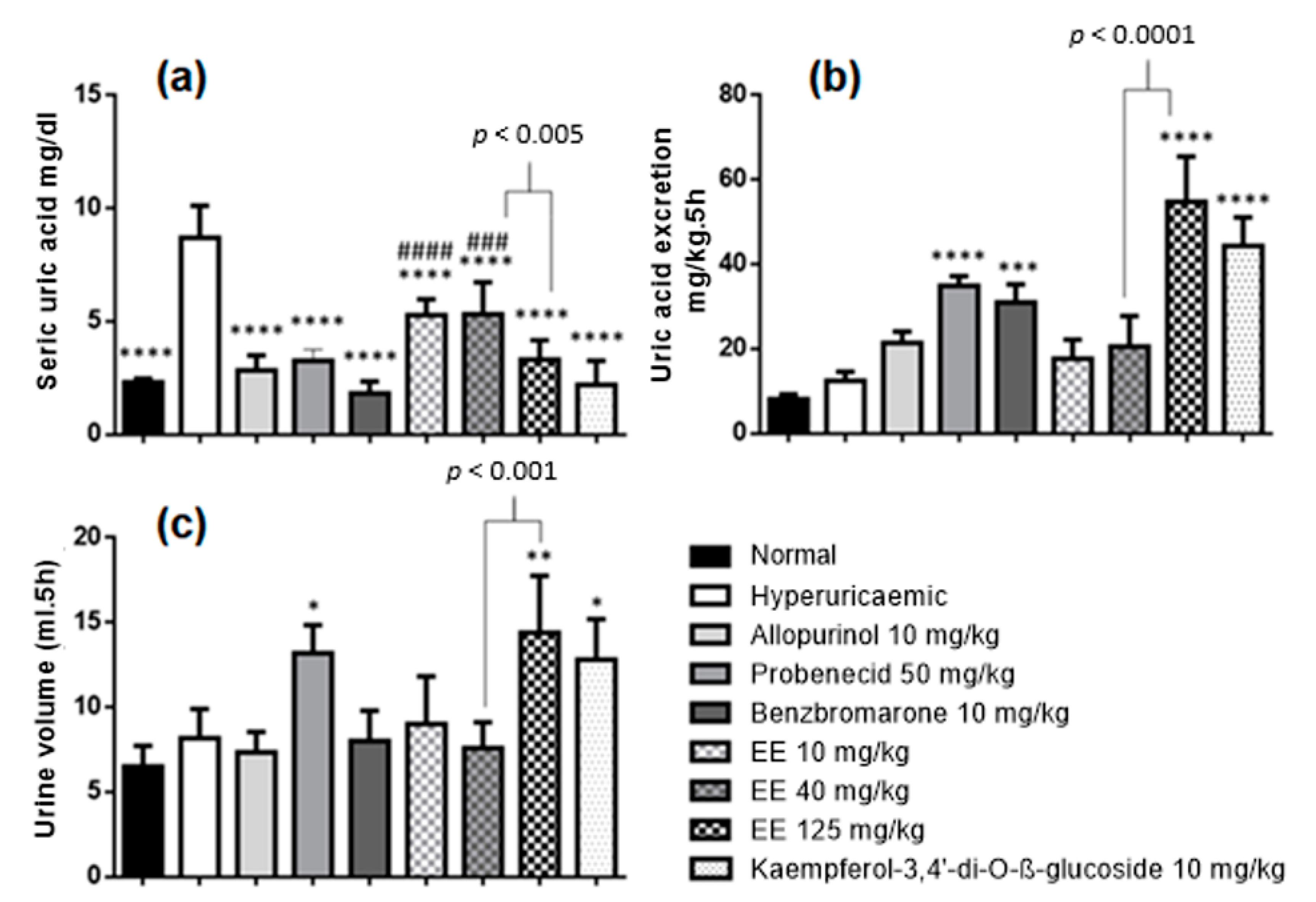
2.4. Effects of Ethanolic Extract of E. sativa Leaves on Hyperuricaemia-Induced Rats
3. Materials and Methods
3.1. Reagents, Chemicals and Analytical Instruments
3.2. Plant Material
3.3. Preparation of the Ethanolic Extract (EE) from E. sativa Leaves
3.4. HPLC Analyses
3.4.1. Qualitative HPLC-DAD Analysis
3.4.2. UHPLC-ESI-QTOF Analysis
3.5. Isolation of the Major Flavonoid in EE
3.6. Validation of a Quantitative Method of the Major Flavonoid in EE by HPLC-DAD
3.7. Quantification of Kaempferol-3,4′-di-O-β-glucoside by HPLC-DAD
3.8. Anti-Hyperuricaemic Assay
3.8.1. Animals
3.8.2. In Vivo Hyperuricaemic Model
3.8.3. Uric Acid Assay
3.8.4. Liver Homogenate Preparation
3.8.5. Total Protein Assay
3.8.6. Hepatic Xanthine Oxidase Activity Assay
3.8.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gustafsson, D.; Unwin, R. The pathophysiology of hyperuricaemia and its possible relationship to cardiovascular disease, morbidity and mortality. BMC Nephrol. 2013, 14, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.M.D.S.; Coelho, G.B.; de Paula Michel Araújo, M.C.; Saúde-Guimarães, D.A. Lychnophora pinaster ethanolic extract and its chemical constituents ameliorate hyperuricemia and related inflammation. J. Ethnopharmacol. 2019, 242, 112040. [Google Scholar] [CrossRef] [PubMed]
- Amorim, H.C.; Henz, G.P.; Mattos, L.M. Boletim de Pesquisa e Desenvolvimento. Identificação Dos Tipos De Rúcula Comercializados No Varejo do Distrito Federal; Embrapa Hortaliças: Brasília, Brazil, 2007; Volume 34, ISBN 1677-2229. [Google Scholar]
- Hetta, M.H.; Owis, A.I.; Haddad, P.S.; Eid, H.M. The fatty acid-rich fraction of Eruca sativa (rocket salad) leaf extract exerts antidiabetic effects in cultured skeletal muscle, adipocytes and liver cells. Pharm. Biol. 2017, 55, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meligi, N.; Hassan, H.F. Protective effects of Eruca sativa (rocket) on abamectin insecticide toxicity in male albino rats. Environ. Sci. Pollut. Res. 2017, 24, 9702–9712. [Google Scholar] [CrossRef]
- Salma, U.; Khan, T.; Shah, A.J. Antihypertensive effect of the methanolic extract from Eruca sativa Mill., (Brassicaceae) in rats: Muscarinic receptor-linked vasorelaxant and cardiotonic effects. J. Ethnopharmacol. 2018, 224, 409–420. [Google Scholar] [CrossRef]
- Jouad, H.; Haloui, M.; Rhiouani, H.; EL Hilaly, J.; Eddouks, M. Ethnobotanical survey of medicinal plants used for the treatment of diabetes, cardiac and renal diseases in the North centre region of Morocco (Fez–Boulemane). J. Ethnopharmacol. 2001, 77, 175–182. [Google Scholar] [CrossRef]
- Mabry, T.J.; Markham, K.R.; Thomas, M.B. The Systematic Identification of Flavonoids; Springer: Berlin/Heidelberg, Germany, 1970; ISBN 978-3-642-88460-3. [Google Scholar]
- Kiddle, G.; Bennett, R.N.; Botting, N.P.; Davidson, N.E.; Robertson, A.A.B.; Wallsgrove, R.M. High-performance liquid chromatographic separation of natural and synthetic desulphoglucosinolates and their chemical validation by UV, NMR and chemical ionisation-MS methods. Phytochem. Anal. 2001, 12, 226–242. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Botting, N.P.; Eagles, J.; Rosa, E.; Williamson, G. Identification of the major glucosinolate (4-mercaptobutyl glucosinolate) in leaves of Eruca sativa L. (salad rocket). Phytochemistry 2002, 61, 25–30. [Google Scholar] [CrossRef]
- Cataldi, T.R.I.; Rubino, A.; Lelario, F.; Bufo, S.A. Naturally occurring glucosinolates in plant extracts of rocket salad (Eruca sativa L.) identified by liquid chromatography coupled with negative ion electrospray ionization and quadrupole ion-trap mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2374–2388. [Google Scholar] [CrossRef]
- Pasini, F.; Verardo, V.; Caboni, M.F.; D’Antuono, L.F. Determination of glucosinolates and phenolic compounds in rocket salad by HPLC-DAD–MS: Evaluation of Eruca sativa Mill. and Diplotaxis tenuifolia L. genetic resources. Food Chem. 2012, 133, 1025–1033. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Llorach, R.; Gil, M.I.; Ferreres, F. Identification of New Flavonoid Glycosides and Flavonoid Profiles To Characterize Rocket Leafy Salads (Eruca vesicaria and Diplotaxis tenuifolia). J. Agric. Food Chem. 2007, 55, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Cuyckens, F.; Shahat, A.A.; van den Heuvel, H.; Abdel-Shafeek, K.A.; El-Messiry, M.M.; Nasr, M.M.S.-E.; Pieters, L.; Vlietinck, A.J.; Claeys, M. The Application of Liquid Chromatography-Electrospray Ionization Mass Spectrometry and Collision-Induced Dissociation in the Structural Characterization of Acylated Flavonol O-Glycosides from the Seeds of Carrichtera Annua. Eur. J. Mass Spectrom. 2003, 9, 409–420. [Google Scholar] [CrossRef]
- Cavaliere, C.; Foglia, P.; Pastorini, E.; Samperi, R.; Laganà, A. Identification and mass spectrometric characterization of glycosylated flavonoids inTriticum durum plants by high-performance liquid chromatography with tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 3143–3158. [Google Scholar] [CrossRef]
- Bell, L.; Methven, L.; Signore, A.; Concha, M.J.O.; Wagstaff, C. Analysis of seven salad rocket (Eruca sativa) accessions: The relationships between sensory attributes and volatile and non-volatile compounds. Food Chem. 2016, 218, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Mithen, R.F.; Dekker, M.; Verkerk, R.; Rabot, S.; Johnson, I.T. The Nutritional Significance, Biosynthesis and Bioavailability of Glucosinolates in Human Foods. J. Sci. Food Agric. 2000, 80, 967–984. [Google Scholar] [CrossRef]
- Orhan, I.E.; Kartal, M.; Sekeroglu, N.; Esiyok, D.; Şener, B.; Ugur, A.; Süntar, I.; Aslan, S. Variations in fatty acid compositions of the seed oil of Eruca sativa Mill. caused by different sowing periods and nitrogen forms. Pharmacogn. Mag. 2010, 6, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Sánchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A Comparative Study of Flavonoid Compounds, Vitamin C, and Antioxidant Properties of Baby LeafBrassicaceaeSpecies. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Concha, M.J.O.; Wagstaff, C. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC–MS: Highlighting the potential for improving nutritional value of rocket crops. Food Chem. 2014, 172, 852–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Nivorozhkin, A.; Szabó, C. Therapeutic Effects of Xanthine Oxidase Inhibitors: Renaissance Half a Century after the Discovery of Allopurinol. Pharmacol. Rev. 2006, 58, 87–114. [Google Scholar] [CrossRef]
- Perez-Ruiz, F.; Hernandez-Baldizon, S.; Herrero-Beites, A.M.; González-Gay, M.A. Risk factors associated with renal lithiasis during uricosuric treatment of hyperuricemia in patients with Gout. Arthritis Care Res. 2010, 62, 1299–1305. [Google Scholar] [CrossRef]
- Selloum, L.; Reichlb, S.; Müllerb, M.; Sebihi, L.; Arnhold, J. Effects of Flavonols on the Generation of Superoxide Anion Radicals by Xanthine Oxidase and Stimulated Neutrophils. Arch. Biochem. Biophys. 2001, 395, 49–56. [Google Scholar] [CrossRef]
- Haidari, F.; Keshavarz, S.A.; Shahi, M.M.; Mahboob, S.-A.; Rashidi, M.-R. Effects of Parsley (Petroselinum crispum) and its Flavonol Constituents, Kaempferol and Quercetin, on Serum Uric Acid Levels, Biomarkers of Oxidative Stress and Liver Xanthine Oxidoreductase Aactivity in Oxonate-Induced Hyperuricemic Rats. Iran. J. Pharm. Res. 2011, 10, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Wang, Y.; Kong, L.D.; Yang, C.; Zhang, X. Effects of Biota orientalis extract and its flavonoid constituents, quercetin and rutin on serum uric acid levels in oxonate-induced mice and xanthine dehydrogenase and xanthine oxidase activities in mouse liver. J. Ethnopharmacol. 2004, 93, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.P.; Dobson, V.L.; Duthie, S.; Hinselwood, D.C.; Kyle, J.; Collins, A. Bioavailability and efficiency of rutin as an antioxidant: A human supplementation study. Eur. J. Clin. Nutr. 2000, 54, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Bao, Z.; Ma, T.; Lin, S. Hypouricemia effects of corn silk flavonoids in a mouse model of potassium oxonated-induced hyperuricemia. J. Food Biochem. 2021, 45, e13856. [Google Scholar] [CrossRef]
- Kang, D.-H.; Park, S.-K.; Lee, I.-K.; Johnson, R. Uric Acid—Induced C-Reactive Protein Expression: Implication on Cell Proliferation and Nitric Oxide Production of Human Vascular Cells. J. Am. Soc. Nephrol. 2005, 16, 3553–3562. [Google Scholar] [CrossRef] [Green Version]
- Mazzali, M.; Hughes, J.; Kim, Y.-G.; Jefferson, J.A.; Kang, D.-H.; Gordon, K.L.; Lan, H.Y.; Kivlighn, S.; Johnson, R. Elevated Uric Acid Increases Blood Pressure in the Rat by a Novel Crystal-Independent Mechanism. Hypertension 2001, 38, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Hsieh, M.-C.; Chang, S.-J. Metabolic syndrome, diabetes, and hyperuricemia. Curr. Opin. Rheumatol. 2013, 25, 210–216. [Google Scholar] [CrossRef]
- Anzai, N.; Ichida, K.; Jutabha, P.; Kimura, T.; Babu, E.; Jin, C.J.; Srivastava, S.; Kitamura, K.; Hisatome, I.; Endou, H.; et al. Plasma Urate Level Is Directly Regulated by a Voltage-driven Urate Efflux Transporter URATv1 (SLC2A9) in Humans. J. Biol. Chem. 2008, 283, 26834–26838. [Google Scholar] [CrossRef] [Green Version]
- Bantia, S.; Parker, C.; Harman, L.; Papac, D.; Hollister, A. OP0106 Effect of BCX4208 add-on therapy to allopurinol 300 mg on plasma hypoxanthine and xanthine concentrations in gout patients. Ann. Rheum. Dis. 2013, 71, 89. [Google Scholar] [CrossRef]
- Hosomi, A.; Nakanishi, T.; Fujita, T.; Tamai, I. Extra-Renal Elimination of Uric Acid via Intestinal Efflux Transporter BCRP/ABCG2. PLoS ONE 2012, 7, e30456. [Google Scholar] [CrossRef] [Green Version]
- Szczurek, P.; Mosiichuk, N.; Woliński, J.; Yatsenko, T.; Grujic, D.; Lozinska, L.; Pieszka, M.; Święch, E.; Pierzynowski, S.G.; Goncharova, K. Oral uricase eliminates blood uric acid in the hyperuricemic pig model. PLoS ONE 2017, 12, e0179195. [Google Scholar] [CrossRef] [Green Version]
- Al-Helaly, L.; Mohammad, F. Uricase Isolated from Seeds of Leek (Allium Ampeloprasum), Celery (Apium Graveolens) and Arugula (Eruca sativa). Iraqi Natl. J. Chem. 2014, 55, 340–356. [Google Scholar]
- Nazif, N.M.; Amira, A.E.H.; Wafaa, A.M.T.; Rasmia, A.H. Chemical Composition and Cytotoxic Activity of Eruca sativa L. Seeds Cultivated in Egypt. Asian J. Chem. 2010, 22, 2407–2416. [Google Scholar]
- Wagner, H.; Bladt, S. Plant Drug Analysis: A Thin Layer Chromatography Atlas, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-642-00574-9. [Google Scholar]
- Brasil RDC N. 166, de 25 de Julho de 2017. Agência Nacional de Vigilância Sanitária. 2017; pp. 1–21. Available online: http://antigo.anvisa.gov.br/documents/10181/2721567/RDC_166_2017_COMP.pdf/d5fb92b3-6c6b-4130-8670-4e3263763401 (accessed on 31 January 2022).
- Validation of Analytical Procedures Text and Methodoloy Q2 (R1). ICH Harmonised Tripartite Guideline. In Proceedings of the International Conference on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use, 1995. Available online: https://www.ich.org/ (accessed on 31 January 2022).
- Brasil Manual de Garantia da Qualidade Analítica, 1st ed.; Ministério da Agricultura, Pecuária e Abastecimento: Brasilia, Brazil, 2011; pp. 40–42.
- National Institutes of Health, Public Health Service. Guide for the Care and Use of Laboratory Animals; NIH Publication: Bethesda, MD, USA, 1985; pp. 23–86.
- Murugaiyah, V.; Chan, K.-L. Mechanisms of antihyperuricemic effect of Phyllanthus niruri and its lignan constituents. J. Ethnopharmacol. 2009, 124, 233–239. [Google Scholar] [CrossRef]
- Ferrari, F.C.; Lima, R.D.C.L.; Filha, Z.S.F.; Barros, C.H.; Araújo, M.C.D.P.M.; Saúde-Guimarães, D.A. Effects of Pimenta pseudocaryophyllus extracts on gout: Anti-inflammatory activity and anti-hyperuricemic effect through xantine oxidase and uricosuric action. J. Ethnopharmacol. 2016, 180, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, A.C.F.P.F.; Coelho, G.B.; Araújo, M.C.D.P.M.; Saúde-Guimarães, D.A. In vivo anti-hyperuricemic activity of sesquiterpene lactones from Lychnophora species. Rev. Bras. Farm. 2019, 29, 241–245. [Google Scholar] [CrossRef]
- Bradford, M.A. Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hall, I.H.; Scoville, J.P.; Reynolds, D.J.; Simlot, R.; Duncan, P. Substituted cyclic imides as potential anti-gout agents. Life Sci. 1990, 46, 1923–1927. [Google Scholar] [CrossRef]
- De Almeida, A.; Elian, S.; Nobre, J. Modifications and Alternatives to the Tests of Levene and Brown & Forsythe for Equality of Variances and Means. Rev. Colomb. Estad. 2008, 31, 241–260. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Compound | RT (min) | Experimental Mass Data ESI+ | Reference |
|---|---|---|---|
| Glucosativin glucosinolate | 1.0 | 328.1384; 166.0861 | [10] |
| Leucine | 1.4 | 182.0808; 268.1037; 132.1019 | Fiocruz |
| Glucoraphanin glucosinolate | 1.8 | 438.0554; 196.0457; 358.0987 | [11,12] |
| Tryptophan | 4.2 | 188.0702, 146.0601; 205.0968 | Fiocruz |
| Kaempferol-3,4′-di-O-β-glucoside | 12.6 | 611.1598; 449.1070; 287.0544 | [13] |
| Angustione | 14.5 | 197.1168; 179.1062 | Fiocruz |
| Kaempferol-3-O-β-glucoside | 15.1 | 449.1068; 287.0544 | [13] |
| Kaempferol-3-O-(2-sinapoyl-β-glucoside) -4′-O-glucoside | 16.2 | 817.2178; 817,2166; 655,1641; 369,1166; 207,0646 | [14] |
| Erucamide | 46.4 | 338,3423 | Fiocruz |
| Peak | d C-13 | C-n | d H HSQC | d H HMBC | H-n (J/Hz) |
|---|---|---|---|---|---|
| 1 | 177.49 | C-4 | --- | --- | --- |
| 2 | 164.81 | C-7 | --- | 6.21; 6.45 | --- |
| 3 | 161.20 | C-4′ | --- | 5.03; 8.12 | --- |
| 4 | 159.24 | C-9 | --- | 6.45 | --- |
| 5 | 156.55 | C-5 | --- | 6.21 | --- |
| 6 | 155.53 | C-2 | --- | 8.12 | --- |
| 7 | 133.72 | C-3 | --- | 5.48 | --- |
| 8 | 130.63 | C-2′,6′ | 8.12 | 8.12 | H-2′,6′ |
| 9 | 123.76 | C-1′ | --- | 7.16 | --- |
| 10 | 115.83 | C-3′,5′ | 7.16 | 7.16 | H-3′,5′ |
| 11 | 103.96 | C-10 | --- | 6.21; 6.45 | --- |
| 12 | 100.86 | C-1A | 5.48 | nd | H-1A (7.50) |
| 13 | 99.97 | C-1B | 5.03 | nd | H-1B (7.25) |
| 14 | 98.93 | C-6 | 6.21 | 6.45 | H-6 (1.73) |
| 15 | 93.85 | C-8 | 6.45 | 6.21 | H-8 (1.73) |
| 16 | 77.62 | C-5B | 3.09 | nd | H-5B |
| 17 | 77.10 | C-5A | 3.39 | nd | H-5A |
| 18 | 76.52 | C-3A | 3.29 | nd | H-3A |
| 19 | 76.44 | C-3B | 3.22 | nd | H-3B |
| 20 | 74.21 | C-2A | 3.17 | nd | H-2A |
| 21 | 73.23 | C-2B | 3.27 | nd | H-2B |
| 22 | 69.91 | C-4B | 3.08 | nd | H-4B |
| 23 | 69.58 | C-4A | 3.19 | nd | H-4A |
| 24 | 60.87 | C-6B | 3.57; 3.33 | nd | H-6a,b-B |
| 25 | 60.63 | C-6A | 3.69; 3.48 | nd | H-6a,b-A |
| Day | Linearity (n = 3) | Concentration Level | RSD % (n = 3) | Accuracy % (n = 3) |
|---|---|---|---|---|
| 1 | Y = 3.364e + 0.007X − 44233 R2 = 0.9973 | Lowest | 1.0 | 102.7–99.0 |
| Medium | 3.5 | 100.6–94.5 | ||
| Highest | 1.7 | 98.5–96.9 | ||
| 2 | Y = 3.422e + 0.007X − 53351 R2 = 0.9946 | Lowest | 5.4 | 101.0–92.7 |
| Medium | 6.9 | 104.0–93.5 | ||
| Highest | 2.4 | 99.4–96.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, A.F.; de Souza, J.; Dophine, D.D.; de Souza Filho, J.D.; Saúde-Guimarães, D.A. Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia. Molecules 2022, 27, 1506. https://doi.org/10.3390/molecules27051506
Teixeira AF, de Souza J, Dophine DD, de Souza Filho JD, Saúde-Guimarães DA. Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia. Molecules. 2022; 27(5):1506. https://doi.org/10.3390/molecules27051506
Chicago/Turabian StyleTeixeira, Arthur Ferrari, Jacqueline de Souza, Douglas Daniel Dophine, José Dias de Souza Filho, and Dênia Antunes Saúde-Guimarães. 2022. "Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia" Molecules 27, no. 5: 1506. https://doi.org/10.3390/molecules27051506
APA StyleTeixeira, A. F., de Souza, J., Dophine, D. D., de Souza Filho, J. D., & Saúde-Guimarães, D. A. (2022). Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia. Molecules, 27(5), 1506. https://doi.org/10.3390/molecules27051506