Abstract
Gal-3, also known as galectin-3, a lectin that binds β-galactosides, has gained attention as a novel biomarker and pathophysiological mediator in cardiovascular disease, where it contributes to inflammation, fibrosis, metabolic dysregulation and cardiac remodeling. This narrative review, developed following SANRA (Scale for the Assessment of Narrative Review Articles) guidelines, aims to integrate current clinical and experimental findings on gal-3 into the American Heart Association Life’s Simple 7 (LS7) and Life’s Essential 8 (LE8). By thematically organizing our review across modifiable domains of cardiovascular health, including glucose regulation, lipid metabolism, physical activity, blood pressure, diet, sleep, tobacco use, and body weight (BMI, body mass index), we highlight the role of gal-3 in linking environmental, behavioral and molecular risk factors to cardiometabolic outcomes. Particular attention is given to the oxidative stress, inflammatory–fibrotic axis, and immune activation as mechanistic pathways underlying gal-3-associated cardiovascular damage. We also discuss its relevance to public health and prevention, considering gal-3’s potential for early risk stratification, monitoring lifestyle interventions and personalized prevention strategies. This review bridges molecular mechanisms with clinical and public health relevance, particularly in the context of environmental and lifestyle-related cardiovascular risk.
1. Introduction
Recent perspectives highlight the growing importance of early biomarker identification and long-term lifestyle monitoring in cardiovascular disease (CVD) prevention, particularly arterial hypertension (AH), in relation to environmental exposures and occupational factors [1].
Large-scale population studies, such as the Framingham Heart Study and PREVEND, have established galectin-3 (gal-3) as a biomarker associated with an elevated risk of heart failure (HF), cardiovascular mortality, and all-cause mortality [2,3]. Although the prognostic role of gal-3 is known, there is still a lack of clear integration of this knowledge with current cardiovascular health (CVH) assessment scales—Life’s Simple 7 (LS7) and Life’s Essential 8 (LE8).
Gal-3 performs a range of functions both within the cell and outside of it. It is involved, among other things, in the regulation of cell adhesion, intercellular signaling, immune response, inflammatory processes, fibrosis, and apoptosis [4]. Elevated gal-3 concentrations have been linked to adverse cardiac outcomes and poorer prognosis, supporting its proposed role in cardiovascular risk stratification, assessment of myocardial fibrosis, and monitoring therapeutic response [5].
In this narrative review, the possible role of gal-3 as a marker of CVH, whose definition is included in the LS7 and LE8 scales, is discussed. Furthermore, the structure of gal-3, its key biological functions, and the associated molecular mechanisms are addressed.
1.1. The Place of Galectin-3 in the Guidelines
The role of gal-3 has been included in cardiology guidelines as a complementary biomarker. Since 2010, the U.S. FDA has approved the use of galectin-3 as a complementary prognostic marker in chronic HF, to be interpreted alongside clinical evaluation. The guidelines of the ESC indicate the need for further studies on the clinical utility of gal-3 [6]. Both the European (ESC) and American (ACC/AHA/HFSA) HF guidelines mention gal-3 (alongside sST2) as a so-called ‘new’ prognostic biomarker, albeit with a limited recommendation (class IIb) [7]. This means that its measurement may be considered in certain situations for risk assessment (e.g., risk of hospitalization or death in HF), but routine use is not yet the standard of care. Due to its pleiotropic nature, gal-3 is a promising target for biomedical diagnostic and therapeutic research [8,9,10].
1.2. Ideal Cardiovascular Health
In 2010, the AHA defined ideal CVH through seven parameters encompassing ideal behaviors and health factors. Later, in an educational campaign, the AHA referred to these recommendations as LS7, which included: tobacco smoking, diet, physical activity, body mass index (BMI), total cholesterol, blood pressure (BP), and glycemia. Each baseline CVH measure was classified as ideal, intermediate, or poor. The mentioned healthy diet components are part of the Dietary Approaches to Stop Hypertension [11]. Table 1 presents the AHA LS7 as the ideal CVH definition.

Table 1.
The AHA Life’s Simple 7 scale for adults, based on Lloyd-Jones et al. [11].
In 2022, LS7 was expanded to LE8 by adding sleep health as the eighth element. Sleep was assessed based on the reported average number of hours of sleep per day. The following question was added: “On average, how many hours of sleep do you get per night?” and, when available, data from sleep-monitoring devices were also taken into account. Sleep health was evaluated according to the LE8 scale, where the optimal sleep range for adults (7–< 9 h) was assigned the highest score (100 points). Deviations from this range resulted in a lower score: 90 points for 9–< 10 h, 70 points for 6–< 7 h, 40 points for 5–< 6 h or ≥ 10 h, 20 points for 4–< 5 h, and 0 points for < 4 h of sleep per day [16,17]. LE8 is considered a better predictor of stroke occurrence [18]. LE8 and LS7 parameters are used to assess so-called ideal CVH, and meeting them is associated with a lower risk of cardiovascular events. LS7 and LE8 focus on modifiable risk factors. Meanwhile, gal-3 is not directly included in these scales. It is a biological indicator potentially associated with the consequences of neglecting the aforementioned factors.
2. Materials and Methods
Our narrative review was prepared following the SANRA (Scale for the Assessment of Narrative Review Articles) criteria, ensuring originality and scientific rigor [19]. A literature review was conducted in accordance with the guidelines for preparing narrative reviews (taking into account the SANRA quality criteria). The focus was primarily on the most recent available publications from the last 10–15 years (mainly 2010–2024). The PubMed/MEDLINE, Embase, Google Scholar, and Scopus databases, as well as the Web of Science search engine, were searched.
Primarily original works (clinical studies—both randomized and observational cohort, cross-sectional), meta-analyses, and systematic reviews, as well as society guidelines and recognized narrative reviews concerning gal-3 were included in the review. Studies on human populations were preferred. Focus was directed toward research assessing gal-3 concentrations in relation to cardiovascular risk, the biological importance of gal-3, and its prospective diagnostic and therapeutic value.
Anecdotal reports, single case descriptions (unless they illustrated a unique mechanism), conference abstracts not supported by a full text, and publications lacking scientific merit (e.g., letters to the editor without data) were excluded. Works written in languages other than English or Polish were also excluded. Studies conducted solely on animal or cell models were not considered if they did not significantly contribute to understanding gal-3 in humans.
The search was conducted separately for each subtopic of the review, combining keywords related to gal-3 and the given issue. Language filters (primarily English-language works) and time filters (limited to the last several years) were applied in the databases. A combination of MeSH terms and free-text synonyms for ‘galectin-3’ and each LS7/LE8 domain was used in the databases.
A search was conducted, initially identifying over 3300 records. After screening titles, abstracts, and full texts, 68 articles were selected for narrative synthesis, structured according to the LS7 and LE8 domains. Studies were included if they met the following criteria: (1) original peer-reviewed research; (2) conducted in human populations; (3) reporting circulating or tissue levels of galectin-3; (4) addressing cardiovascular, metabolic, or neurological outcomes related to LS7/LE8 domains; and (5) published in English or Polish. Preclinical studies (in vitro or animal models) were cited selectively only when they provided critical mechanistic insights that were not available from human studies, and their nature was explicitly indicated in the text. This review was designed as a narrative synthesis; therefore, no formal risk of bias assessment was performed.
The reference lists of the identified articles were also reviewed for additional sources (so-called snowballing). As a result of this process, several dozen items were collected, from which the most relevant and highest-quality evidence was selected for discussion in this review.
In reporting the results of the cited primary studies, a simplified approach to statistical significance was applied. p-values are presented as either p < 0.05 or p < 0.001, depending on the level of significance reported in the original sources (except tables). This distinction aims to improve clarity and allows for an approximate assessment of the statistical strength of the findings, without introducing excessive numerical precision. p-values were not interpreted as indicators of effect size or clinical relevance [20,21,22]. Where available, effect estimates (e.g., OR, HR, or mean differences) are also provided along with confidence intervals, which more accurately convey the precision and potential clinical importance of the findings
In accordance with principles of transparency and methodological clarity, this narrative review was prospectively registered on the Open Science Framework under an open-ended registration [23].
3. Properties of Galectin
3.1. Structure and Biological Functions of Galectin-3
Gal-3 belongs to the family of lectins that bind β-galactosides, characterized by a unique structure and diverse biological functions, including the regulation of inflammatory processes, fibrosis, and cell adhesion. Gal-3 is present both inside cells and in the extracellular space, performing functions related to adhesion, cell migration, and the regulation of the immune response [8,10].
Gal-3 possesses a carbohydrate recognition domain of approximately 130 amino acids responsible for binding β-galactosides, along with an unstructured N-terminal domain that facilitates oligomerization [4,5,24]. In solution, galectin-3 typically exists as a monomer but can form pentamers upon interaction with polysaccharides [9]. It is localized broadly within cells, being present in the cytoplasm, nucleus, and extracellular space [5].
In the extracellular environment, gal-3 also participates in angiogenesis and regulates leukocyte activity, influencing the course of the immune response. It can both activate leukocytes and exert anti-inflammatory effects by inducing the apoptosis of T lymphocytes [4,5].
Given these features, gal-3 has been increasingly studied for its diagnostic relevance in cardiovascular diseases and as a target for antifibrotic therapies. Table 2 presents the characteristics and functions of gal-3.
Table 2.
Characteristics and functions of galectin-3.
3.2. The Role in Inflammation and Fibrosis
Galectin-3, secreted by activated macrophages, mediates inflammatory processes and tissue remodeling, particularly by promoting myocardial fibrosis and increasing cardiac stiffness and dysfunction [5]. In other organs, such as the kidneys, liver, and lungs, it also contributes to the progression of fibrosis and inflammation, and its elevated concentrations are associated with the worsening of these processes, for example, in diabetic nephropathy or idiopathic pulmonary fibrosis [7,47]. Gal-3 also plays a significant role in cancer progression by promoting metastasis [48].
In an inflammatory environment, gal-3 acts not only as a macrophage-regulating factor but also as a DAMP/alarmin, signaling the presence of tissue damage or infection. However, its immunomodulatory effects are tissue context-dependent—in some settings, it may support the immune response, while in others, it may exert immunosuppressive effects.
Because of its essential function in tissue remodeling, gal-3 is considered a potential signal of inflammatory and fibrotic activity, notably in heart failure, where elevated blood concentrations may point to active remodeling [5]. Table 3 presents the role of gal-3 in inflammation and fibrosis.
Table 3.
The role of galectin-3 in fibrosis and inflammation.
3.3. Molecular Mechanisms
Galectin-3 exerts its functions through various molecular mechanisms, stemming from its ability to bind sugars as well as interact with proteins. These mechanisms underlie the previously described processes of inflammation and fibrosis, integrating the activation of the mentioned pathways [49,50]. A specific binding pocket in the carbohydrate recognition domain (CRD) recognizes β-galactosides present in glycoproteins and glycolipids. This binding relies on weak interactions (hydrophobic, hydrogen bonds), making it reversible; sugar ligands, such as lactose, can effectively block it. Many functions of gal-3 result from its cooperation or competition with other galectins, such as galectin-1, which influences the regulation of biological responses under various conditions [4,55]. Gal-3 interacts not only with sugars but also with various cytoplasmic and nuclear proteins. For example, through the NWGR motif (Asparagine-Tryptophan-Glycine-Arginine motif) within the CRD, it binds to Bax and Bcl-2, protecting the cell from apoptosis [8,28]. In the cell nucleus, it cooperates with SR proteins (serine/arginine-rich proteins), forming part of spliceosome complexes and regulating pre-mRNA processing. Additionally, gal-3 binds to PI3K, Ras, and FAK kinases, influencing cellular decisions regarding proliferation and migration. Despite lacking a classical signal sequence, galectin-3 is likely secreted via exosomes and microvesicles. In the extracellular environment, it acts in an autocrine and paracrine manner, activating glycan-dependent receptors [8].
The main molecular mechanisms described above are briefly summarized and illustrated in Figure 1. Galectin-3’s CRD binds β-galactosides reversibly [4]. Intracellularly, gal-3 modulates apoptosis via Bax/Bcl-2 interaction and regulates mRNA splicing through SR proteins [8,28]. It also interacts with PI3K, Ras, and FAK kinases, affecting proliferation and migration. Extracellular secretion occurs via exosomes, mediating paracrine signaling through glycan receptor activation [8]. Additionally, gal-3 cooperates or competes with galectin-1, modulating biological responses [4,55].
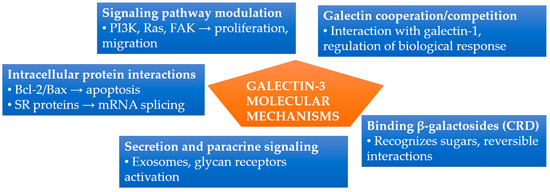
Figure 1.
Summary of molecular mechanisms of galectin-3 activity. Gal-3 functions through carbohydrate binding, intracellular protein interactions, signaling pathway modulation, extracellular secretion, and interactions with other galectins. Abbreviations: CRD, carbohydrate recognition domain; FAK, focal adhesion kinase; PI3K, phosphoinositide 3-kinase; SR proteins, serine/arginine-rich proteins.
Galectin-3 combines in its structure unique features (a single lectin domain with an attached disordered oligomerizing tail), which translate into a wide range of biological functions. Thanks to its ability to recognize sugars and form interaction networks, gal-3 influences cell adhesion, receptor organization on the membrane, and signal transduction. At the same time, through direct binding to intracellular proteins, it regulates key cellular processes such as apoptosis and gene expression. The multifunctionality of gal-3 makes it a crucial player in maintaining homeostasis, and disruptions in its expression or activity contribute to the development of numerous diseases. Therefore, gal-3 is the subject of extensive research regarding its role in the diagnosis and therapy of inflammatory, fibrotic, and neoplastic conditions.
3.4. Galectin-3 as a Biomarker in Cardiology
Galectin-3 contributes to key inflammatory and fibrotic pathways implicated in CVD progression. The most extensive data concern its use in HF, where elevated gal-3 concentrations correlate with increased fibrosis, worse prognosis, and higher mortality, with a hazard ratio of 1.38 for all-cause mortality [7]. Similarly, in acute coronary syndromes, gal-3 acts as a marker of inflammation and cardiac remodeling, significantly elevated in STEMI patients compared to controls (10.6 ng/mL vs. 5.5 ng/mL, respectively) [56]. In atrial fibrillation, gal-3 is recognized as an indicator of atrial fibrosis, associated with increased atrial fibrillation risk (OR = 1.45) [57]. Moreover, gal-3 concentrations correlate with left ventricular hypertrophy in AH, indicating its role in the progression of structural cardiac changes [58]. Finally, in patients with aortic stenosis, elevated gal-3 concentrations correlate with left ventricular dysfunction; however, current data on its prognostic utility in this condition remain inconclusive [59].
Galectin-3 remains under investigation as a prognostic and therapeutic marker, although its routine clinical use requires further validation.
3.5. Galectin-3 Inhibitors
Given the role of gal-3 in promoting cardiac fibrosis and subsequent heart damage, the inhibition of gal-3 has emerged as a potential antifibrotic therapeutic strategy. Preclinical studies evaluating modified citrus pectin (MCP), an oral gal-3 inhibitor, demonstrated promising results, including reduced inflammation, fibrosis, and angiogenesis in various animal models and in vitro studies [60]. Similar beneficial effects were observed in experimental models involving pressure overload or myocardial infarction, indicating possible protective properties against cardiac remodeling and dysfunction [61,62].
However, these positive outcomes have not yet been replicated in clinical trials. The first randomized clinical study assessing MCP did not show a significant reduction in organ fibrosis, although it confirmed the safety and good tolerability of the compound. These findings indicate that achieving a clinically meaningful antifibrotic effect through gal-3 inhibition might require longer treatment durations or possibly combination therapy [58].
Belapectin (GR-MD-02), a galactoarabino-rhamnogalacturonan derived from pectin, is among the most clinically advanced galectin-3 inhibitors. In a phase II clinical trial involving patients with liver fibrosis secondary to NASH, belapectin modestly reduced the incidence of new esophageal varices and slightly lowered portal vein pressure; however, its antifibrotic effect on liver tissue itself remained uncertain [63]. Another galectin-3 inhibitor, GB1107—a small-molecule oral inhibitor targeting the CRD domain—is currently under investigation. A phase I clinical trial (GB1211, ClinicalTrials.gov Identifier: NCT05009680) assessing safety and tolerability in patients with cirrhosis of various etiologies is ongoing, and definitive conclusions about efficacy cannot yet be drawn [64].
In summary, despite promising preclinical data, the clinical utility of galectin-3 inhibitors remains uncertain. Results from ongoing clinical trials will be essential to better define their therapeutic potential.
3.6. Galectin-3 and Myocardial Extracellular Volume
Myocardial extracellular volume (ECV), most commonly measured using magnetic resonance imaging (MRI) with T1 mapping, is an indicator of the degree of fibrosis and remodeling in the heart myocardium. Since gal-3 promotes fibrosis, the relationship between gal-3 concentrations and ECV is of particular interest. Several studies have demonstrated a significant positive correlation between blood concentrations of gal-3 and myocardial ECV.
In patients with a history of myocardial infarction, this relationship was demonstrated by Perea et al. [65]. A study involving 26 patients after STEMI measured biomarkers (gal-3, BNP) and performed MRI during the acute phase, with follow-up MRI at 6 months to assess ECV in the post-infarction myocardium. Gal-3 concentrations on day 7 post-infarction were found to be significantly and positively correlated with ECV values at 6 months (r = 0.428; p < 0.05). By comparison, BNP showed only a trend toward correlation (r = 0.38; p = 0.059). Moreover, gal-3 was an independent predictor of ECV increase in multivariate analysis; together with BNP, it explained about 30% of ECV variability (R2 = 0.34; p < 0.05). As mentioned earlier, exceeding a gal-3 threshold of > 10.15 ng/mL in the acute phase identified patients with significant ECV expansion with high sensitivity (AUC = 0.76). These findings suggest that circulating gal-3 during the peri-infarction period reflects an intensified fibrotic and remodeling process that contributes to increased extracellular matrix in the post-infarction healing phase. Gal-3 may serve as an early marker to identify patients prone to maladaptive cardiac scarring.
In chronic conditions such as heart failure with preserved ejection fraction (HFpEF), the relationship between gal-3 and ECV is also of interest. HFpEF is characterized by diffuse myocardial fibrosis, leading to impaired relaxation. Serial measurements of gal-3 in patients with HFpEF increased alongside fibrosis progression and were associated with worsening diastolic function, as inferred from E/e’ on echocardiography [66,67]. Further studies are needed that integrate precise imaging measurements (such as ECV from MRI) with biomarkers to more fully define the relationship between gal-3 and fibrotic burden in the heart among patients with chronic diseases. In summary, there is a consensus that gal-3 correlates with the degree of myocardial fibrosis as measured by ECV. In acute conditions (infarction, inflammation), elevated gal-3 predicts greater fibrosis during the healing phase [65,68]. In chronic diseases, gal-3 also reflects, at least partially, the fibrotic burden, although its concentration is influenced by multiple factors (e.g., kidney function).
Studies using ECV as the “gold standard” have shown that gal-3 is a useful, though nonspecific, marker of the extracellular matrix. ECV and gal-3 do not compete but rather complement each other: ECV provides direct information about cardiac fibrosis, while gal-3 offers a global view of pro-fibrotic and inflammatory activation in the body. In clinical practice, measuring gal-3 is incomparably simpler and cheaper than ECV imaging, making it suitable as a screening tool; in turn, elevated gal-3 may serve as an indication for advanced imaging to assess the extent of fibrosis.
3.7. Genetic Determinants of Circulating Galectin-3 Concentrations and Ethnic Differences
In recent years, genome-wide association studies have identified genetic variants significantly associated with serum gal-3 concentrations. A key breakthrough has been the identification of two major loci influencing gal-3: variants within the LGALS3 gene (polymorphisms rs4644, rs4652) and the ABO gene, which determine blood groups. Variants in LGALS3 may directly modulate gal-3 expression, a finding confirmed at both the transcriptomic and protein levels. Together, these variants explain up to 29% of the variability in serum gal-3 concentrations observed within the European population [69].
Regarding cardiovascular diseases, special attention has been paid to the potential links between gal-3-associated polymorphisms and risks of HF and atherosclerosis. Mendelian randomization studies, however, suggest that although elevated gal-3 concentrations serve as a marker of poor prognosis in CVDs, their causal role in the development of HF and vascular diseases remains uncertain [70,71].
In a large-scale proteogenomic study [72], using cis-Mendelian randomization analysis, higher gal-3 concentrations positively correlated with an increased risk of HF. Nonetheless, other Mendelian randomization analyses failed to confirm a direct causal link between genetically elevated gal-3 concentrations and HF risk. These findings imply that gal-3 is likely a biomarker reflecting cardiac fibrosis and remodeling rather than an etiological factor.
Importantly, ethnic differences might also influence gal-3 concentrations. A study conducted in an Asian population identified specific regulatory variants within LGALS3, differing in frequency compared to European populations. Notably, variant rs28379289, common in East Asian populations, was associated with increased LGALS3 expression in placental tissues, indicating ethnic variation in gal-3 regulation [73].
From an epidemiological perspective, higher gal-3 concentrations observed among individuals of Asian descent might partially explain differences in cardiovascular disease incidence across populations. However, further studies incorporating both genetic and environmental contexts are required to precisely elucidate the clinical significance of these observations. The simplified diagram presented in Figure 2 summarizes the current knowledge on genetic variants influencing gal-3 concentrations, highlights ethnic differences, and outlines potential clinical implications.
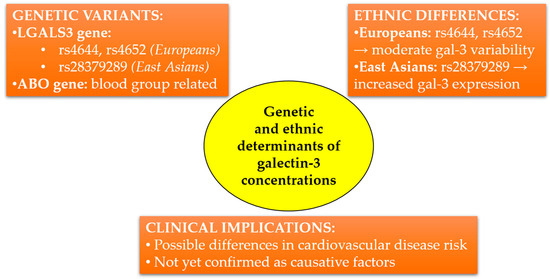
Figure 2.
Genetic and ethnic determinants influencing circulating galectin-3 concentrations. LGALS3 and ABO gene variants differ in frequency between European and East Asian populations, potentially affecting cardiovascular risk. Clinical implications remain to be fully elucidated.
4. Association of Galectin-3 with Individual Components of LS7 and LE8
4.1. Tobacco Smoking
The relationship between tobacco smoking and gal-3 concentrations has been analyzed in numerous studies. Substances contained in cigarette smoke may regulate gal-3 expression and activate related inflammatory and pro-fibrotic mechanisms. However, findings from both clinical and basic research are heterogeneous, indicating the complexity of this association and the need for cautious interpretation of the available data.
The study by de Boer et al. [74], which included a large population (n = 7968), showed a weak but significant negative correlation between tobacco smoking and gal-3 concentrations (β = −0.051, p < 0.001). However, after adjusting for sex, age, and kidney function, this association was no longer significant (β = 0.002, p > 0.05). Smoking may play a limited role as an independent determinant of gal-3 concentration, highlighting the importance of other factors such as renal function or BMI. Nevertheless, the authors did not analyze molecular mechanisms or the effect of smoking cessation on gal-3 concentrations.
Evidence pointing to a direct link between tobacco smoke and the activation of gal-3-related mechanisms was provided by the study by Sharma et al. [75], which observed increased gal-3 expression in AECs exposed to cigarette smoke extract (CSE). Gal-3 mRNA concentrations rose significantly (up to fivefold at 100 µM CSE, p < 0.001) and were associated with enhanced epithelial-to-mesenchymal transition. A key mechanism involved oxidative stress activating the RUNX-2/gal-3 axis. An antioxidant (N-acetylcysteine) and the gal-3 inhibitor GB1107 effectively suppressed these changes, suggesting potential therapeutic strategies.
Similar findings were presented in another study [76], which showed that nicotine directly increases gal-3 expression in breast cancer cells (MCF-7). This expression was dependent on activation of the α9 nicotinic acetylcholine receptor and the STAT3 pathway. Gal-3 expression was significantly higher in breast cancer tissues of smoking patients (twofold increase, p < 0.001). Additionally, gal-3 exerted a protective effect against mitochondrial apoptosis induced by cytostatic agents, indicating a role for nicotine and gal-3 in chemoresistance.
Clinically relevant insights are provided by the study [77], which assessed gal-3 concentrations in patients with chronic obstructive pulmonary disease (COPD). The comparative group consisted of never-smokers and former smokers. Gal-3 concentrations were significantly highest among current smokers, both during exacerbation and recovery phases. Additionally, gal-3 correlated with inflammatory markers such as hsCRP (r = 0.35, p < 0.05) and NT-proBNP (r = 0.32, p < 0.05). Smoking status was the only significant predictor of gal-3 concentrations (β = 0.458, p < 0.05). These findings indicate that gal-3 serves as a smoking-induced inflammatory biomarker, particularly relevant in patients with COPD.
A significant role of gal-3 in the response of EPCs to cigarette smoke was also described in the study [78]. Exposure of the cells to CSE resulted in a marked increase in gal-3 expression (6.7-fold mRNA increase at 8% CSE, p < 0.001) and induction of autophagy. The use of galectin-3 short hairpin RNA (a molecular biology technique used to silence the gene encoding gal-3) inhibited smoke-induced processes such as autophagy and endothelial cell dysfunction. This suggests a key role of gal-3 in the harmful cardiovascular effects of smoking.
Similarly, Zhang et al. [79] demonstrated that gal-3 was essential for the activation of the MUC1-C/EGFR pathway in epithelial cells exposed to cigarette smoke. Smoke exposure led to a threefold increase in gal-3 bound to MUC1-C (p < 0.05). This triggered activation of the pro-inflammatory Src/Jnk pathway, leading to E-cadherin degradation and epithelial-to-mesenchymal transition. Inhibition of MUC1 glycosylation prevented its interaction with gal-3 and suppressed the pathological effects, indicating the potential relevance of this mechanism as a therapeutic target.
The harmful effects of tobacco smoke, confirming the role of gal-3 as a mediator of oxidative stress and inflammation, were demonstrated by Nemmar et al. [80] in a vascular model of exposure to waterpipe smoke. Exposure led to an increase in gal-3 concentrations (p < 0.001) as well as inflammatory and oxidative stress markers (TNF-α, IL-1β, lipid peroxidation). These changes were associated with endothelial dysfunction and DNA damage, indicating a potential mechanism by which smoking affects the cardiovascular system.
Experimental data indicate that galectin-3 plays a significant role in the inflammatory response and oxidative stress induced by nicotine. In a study [81] conducted on rats, daily nicotine exposure (0.5 mg/kg/day) led to a significant increase in galectin-3 concentrations in lung tissue relative to the control group (p < 0.001). This was accompanied by elevated concentrations of pro-inflammatory cytokines: IL-6 and IL-1β (both: p < 0.05), as well as oxidative stress markers—malondialdehyde levels were significantly higher (p < 0.05), while the activity of antioxidant enzymes (GSH-Px, SOD) was reduced. Administration of dexpanthenol significantly lowered gal-3 concentrations and inflammatory markers, restoring them to values close to the control group, suggesting its potential protective effect. Notably, the study did not evaluate the effect of smoking cessation on gal-3 concentrations, which represents a significant limitation in the context of cardiovascular prevention and the reversibility of these changes.
Table 4 presents a summary of studies analyzing the effects of tobacco use (in various forms: cigarette smoke, nicotine, waterpipe smoke) on gal-3 concentrations, the molecular mechanisms regulating its expression, and associated biological effects.
Table 4.
The impact of tobacco smoking and nicotine on galectin-3 concentrations, molecular mechanisms, and biological effects: data from experimental and clinical studies.
In summary, the data indicate a significant, though not fully consistent, association between tobacco smoking and gal-3 concentrations, which may serve as a biomarker of inflammatory responses and oxidative damage induced by tobacco smoke components. However, the mechanisms regulating gal-3 expression require further investigation, particularly regarding the effects of smoking cessation, which have not been adequately addressed in existing studies.
4.2. Diet
Galectin-3 can be significantly modulated by dietary habits. In the LS7 scale, diet is one of the modifiable cardiovascular risk factors, potentially influencing the concentrations of gal-3.
Studies concluded that a high-fat diet (HFD) significantly increases gal-3 expression and activates inflammatory and fibrogenic pathways. It has been shown that consumption of a HFD induces gal-3 expression and activation of the NLRP3 inflammasome via TLR4 [83]. This heightens inflammatory processes in the liver and potentially offers a role in the pathogenesis of non-alcoholic steatohepatitis (NASH). Similar results were presented by Iacobini et al. [84], who, in a knockout mouse model (Lgals3−/−), demonstrated that the lack of gal-3 protected animals from diet-induced NASH. These findings suggest that limiting saturated fat intake may reduce the risk of chronic inflammatory and fibrotic processes mediated by gal-3.
Marín-Royo et al. [85] confirmed that excess saturated fats in the diet induce gal-3 overexpression in the hearts of rats, leading to lipotoxicity, oxidative stress, and mitochondrial dysfunction. The use of MCP, a gal-3 inhibitor, significantly reduced these adverse effects. This suggests potential benefits of diets rich in components that limit gal-3 activity.
The study by Gonçalves et al. [86] in rats fed a hypercaloric diet (rich in fats, simple sugars, and salt) also confirms that even short-term consumption of such a diet significantly increases gal-3 concentrations, resulting in cardiac hypertrophy and myocardial fibrosis. This evidence highlights the necessity of a healthy diet in the prevention of CVDs.
On the other hand, the absence of gal-3 may paradoxically exacerbate the metabolic consequences of a HFD, including increased adipose tissue mass and heightened inflammation [87]. This highlights the important role of gal-3 in maintaining metabolic balance in the context of various dietary habits. A healthy diet, by reducing pro-inflammatory factors associated with HFD, may support the protective function of gal-3 against the metabolic consequences of obesity and insulin resistance.
Wensvoort [88] presents a hypothesis that a Western diet rich in processed red meat may promote vascular remodeling through excessive activation of the elastin receptor complex (ERC) by elastin-derived peptides. Although the author did not conduct direct studies on gal-3, he included it in the model as a potential additional ligand of ERC, suggesting an indirect link to inflammation and vascular remodeling characteristic of metabolic syndrome. In conclusion, he recommends avoiding processed meats and other components that enhance chronic inflammation. A healthy diet should avoid pro-inflammatory components, potentially modulating mechanisms indirectly related to gal-3.
Important data on dietary interventions also come from studies on intermittent fasting (IF). A WONDERFUL trial (Weekly ONe-Day WatER-only Fasting InterventionaL trial for low-density lipoprotein cholesterol reduction) presented a significant increase in gal-3 concentrations after 26 weeks of IF [89]. This increase correlated with improvements in metabolic parameters, such as reductions in HOMA-IR, glucose, and insulin levels. Conversely, Lee et al. [90] and Kim et al. [91] demonstrated that IF effectively lowers gal-3 concentrations in mice fed a HFD, alleviating liver fibrosis, inflammation, and adipose tissue inflammation. This suggests that IF may modulate gal-3 concentrations, reducing the metabolic and inflammatory effects of excessive caloric and fat intake.
The result of dietary sodium content on kidney function and fluid–electrolyte balance has also been described in a mouse model with gal-3 deficiency [92]. Although the study did not directly address healthy dietary patterns, its findings suggest that sodium intake control may indirectly influence gal-3-regulated mechanisms, particularly those related to electrolyte balance and BP regulation.
Table 5 presents selected studies on the impact of dietary interventions on gal-3 concentrations and associated inflammatory and metabolic mechanisms. The results indicate that a HFD consistently leads to increased gal-3 concentrations and enhanced inflammatory responses, while intermittent fasting may modulate gal-3 concentrations depending on the physiological context.
Table 5.
The impact of various dietary interventions on galectin-3 concentrations and associated inflammatory pathways: a summary of experimental and clinical studies.
In summary, the available data indicate that a diet rich in saturated fats, simple sugars, and processed red meat significantly increases gal-3 concentrations, exacerbating inflammation, fibrosis, and metabolic disturbances. In contrast, dietary interventions such as IF or diets that limit saturated fat intake can effectively modulate gal-3 concentrations, contributing to improvements in metabolic parameters, reduction of inflammation, and potential cardiovascular protection. However, data are still lacking regarding the effects of healthy dietary components (e.g., antioxidants, PUFAs, fiber) on gal-3 regulation, which represents an important direction for future research.
4.3. Physical Activity
Gal-3 concentrations can be modulated by both acute, regular physical activity and intense physical exertion. Physical activity is a component of both the LS7 and LE8 scales.
It has been shown that acute high-intensity interval training (HIIT) leads to a significant increase in serum gal-3 concentrations (by an average of 39.5%, p < 0.001), in both runners and cyclists [93]. The authors associated these changes with endothelial activation. They suggested that short-term increases in gal-3 may reflect a response to mechanical vascular stress and the initiation of endothelial remodeling processes, potentially even fibrotic changes via endothelial-to-mesenchymal transition. However, the main limitations of this study were its short observation period and small sample size (n = 18), which make it difficult to assess the clinical significance of the findings.
Similar observations were made by Kaleta-Duss et al. [94] among 35 amateur marathon runners, demonstrating a transient increase in gal-3 after the marathon (from an average of 8.53 to 10.65 ng/mL, p < 0.05). Galectin-3 concentrations returned to baseline values two weeks after the run. The increase in gal-3 correlated with an increase in heart-type fatty acid binding protein (r = 0.52, p < 0.05), suggesting the possibility of adaptive changes related to cardiac load. Further evidence for the transient, adaptive nature of gal-3 increase is provided by findings from Vassalle et al. [95], who observed a significant but short-term increase in gal-3 (by an average of 33%, p < 0.001) in half-marathon runners, which normalized within 24 h. The same was shown in the study by Le Goff et al. [96], where the increase in gal-3 was significantly more pronounced in ultramarathon runners (from 9.39 ± 2.94 ng/mL to 19.8 ± 4.39 ng/mL, p < 0.001) than in runners of shorter distances (from 9.23 ± 2.56 ng/mL to 23.9 ± 5.59 ng/mL, p < 0.001). In the reference group, which ran 10 km, the increase in galectin-3 was less marked (from 10.8 ± 2.63 ng/mL to 14.1 ± 3.38 ng/mL). Three hours after exertion, gal-3 concentrations returned to near-baseline values in all groups, suggesting that the biomarker increase after intense exercise is transient. The study by Lewicka-Potocka et al. [97] complements these observations, indicating that intense endurance exercise (marathon) leads to an increase in gal-3 concentrations, which negatively correlate with right ventricular (RV) ejection fraction (r = −0.48, p < 0.05) and with reduced RV radial shortening. Elevated gal-3 concentrations were connected with lower VO2 max (r = −0.47, p < 0.05), suggesting that intense physical effort may induce adaptive RV remodeling associated with a transient elevation in this biomarker.
The issue of the source of gal-3 increase during exercise has also been analyzed in animal models of vascular and cognitive disorders. It has been shown that regular physical activity significantly reduces gal-3 expression in microglia and white matter [98,99], leading to a reduction in neuroinflammation and improvement in cognitive function in mice with vascular dementia. The evidence implies that a potential neuroprotective effect of regular physical exercise through gal-3 modulation, which may also have implications for CVH.
The beneficial results of regular physical activity are also confirmed by the study [100], which demonstrated that an eight-week training program (either combined or HIIT) significantly reduced gal-3, CRP, and fibrinogen concentrations in patients with CAD after COVID-19 (p < 0.05). Meanwhile, the randomized controlled trial by Keyhani et al. [101] indicates the potential for a significant reduction in gal-3 gene expression in postmenopausal women, both following HIIT (a reduction of 94.15%) and with moderate aerobic activity (84.74%), which was also associated with improvement in lipid profile.
In contrast to the studies described above, the findings of Ahmad et al. [102] suggest an association between elevated gal-3 concentrations and reduced physical performance in patients with HF, where elevated gal-3 concentrations were inversely correlated with the results of the short physical performance battery (r2 = 0.089, p < 0.05) and handgrip strength (r2 = 0.078, p < 0.05).
The study by Pacheco et al. [103] indicates that physical activity status in older adults may influence the expression of galectin-3 binding protein (LGALS3BP), although no significant differences in gal-3 concentrations were observed. The evidence implies that while physical activity may have a beneficial effect on the expression of related proteins regulating inflammation, the direct impact of exercise on gal-3 requires further investigation.
Salvagno et al. [104] demonstrated that extreme physical exertion (a 60 km ultramarathon) causes a transient increase in gal-3 concentrations (2.4-fold increase; p < 0.001), which, however, was not correlated with cardiac injury biomarkers (TnI and NT-proBNP). This suggests that gal-3 may reflect an adaptive response of the body to extreme exertion rather than actual myocardial damage.
Table 6 presents a synthetic summary of findings from experimental and clinical studies analyzing the impact of physical activity, both acute and chronic, on galectin-3 concentrations. The table allows comparison of training effects depending on intensity, duration, and participants’ health status, and also points to potential mechanisms underlying changes in gal-3 concentrations.
Table 6.
Impact of different forms of physical activity on galectin-3 concentrations in the context of cardiovascular health: a summary of literature data.
In summary, intense, short-term physical activity leads to a transient increase in gal-3, which likely reflects adaptive changes within the cardiovascular system and endothelium. Regular exercise, particularly aerobic training or HIIT, tends to reduce gal-3 concentrations, suggesting a role in long-term cardiovascular and cognitive protection. However, the long-term effects of repeated gal-3 elevations in individuals engaged in high-volume endurance sports require further investigation. Larger prospective studies are also necessary to evaluate the clinical significance of gal-3 modulation through physical activity in reducing the risk of CVDs.
4.4. BMI
Gal-3 concentration may be significantly modulated by body mass and fat distribution, particularly abdominal obesity.
A study conducted in a large Chinese population demonstrated a significant, though moderate, association between gal-3 concentration and BMI (R = 0.07, p < 0.05). This correlation lost statistical significance after adjusting for gender and age (β = 0.04; 95% CI: −0.02, 0.1; p = 0.15) [108]. A stronger correlation was noted with waist circumference (WC), which remained significant even after multivariate regression analysis (β = 0.12; 95% CI: 0.04, 0.21; p < 0.05). This suggests that visceral obesity is a better predictor of gal-3 concentration than BMI alone. Gal-3 concentration also correlated with other metabolic markers, including triglycerides (β = 0.11; p < 0.001) and hs-CRP (β = 0.08; p > 0.05), which may indicate a role for this biomarker in obesity-related inflammation.
Similar associations were confirmed by Weigert et al. [109], who analyzed gal-3 concentrations in individuals with normal BMI, overweight, and type 2 diabetes mellitus (T2DM). They found significantly elevated gal-3 concentrations in those with overweight and T2DM compared to the control group. Gal-3 concentration showed a positive association with BMI (r = 0.357; p < 0.05). Notably, the study also observed higher gal-3 concentrations in visceral compared to subcutaneous adipose tissue, suggesting a key role of visceral fat as a source of this biomarker’s synthesis.
Important data on the association between gal-3 and obesity were also provided by the study of Suthahar et al. [110], conducted in the large PREVEND cohort (8202 participants aged 28–75 years, with a median follow-up of 11.3 ± 3.1 years). It was shown that gal-3 concentration significantly increased with rising BMI (obesity: 11.7 ± 2.3 vs. normal body weight: 10.2 ± 1.9 ng/mL, p < 0.001). However, unlike other biomarkers such as NT-proBNP or cTnT, gal-3 did not demonstrate significant prognostic value for HF risk in the context of BMI.
The study [111], conducted in a population of 8687 individuals within the ARIC project (Atherosclerosis Risk in Communities Study), provides important clinical evidence on the relationship between high BMI, increased gal-3 concentrations, and HF risk. Individuals with severe obesity (BMI ≥ 35 kg/m²) had more than twice the risk of elevated gal-3 concentration (OR = 2.32; p < 0.001). This relationship was notably strong in the subgroup of women with severe obesity (OR = 3.00), suggesting potential gender differences. Moreover, the coexistence of severe obesity and high gal-3 was connected with more than a fourfold increased risk of HF (HR = 4.19; p < 0.001).
Interesting findings regarding the impact of weight reduction on gal-3 concentrations were presented by Aksit et al. [112], who assessed patients with obesity undergoing bariatric surgery. Initially, gal-3 concentrations were significantly higher in the group with class III obesity (17.6 ± 4.2 vs. 14.1 ± 3.0 ng/mL; p < 0.05) and presented a positive correlation with BMI (r = 0.375; p < 0.001). After surgery, although a downward trend in gal-3 concentration was observed at 3 and 6 months, these changes did not reach statistical significance, suggesting that the observation period was too short to capture meaningful changes in this biomarker.
Fryk et al. [113], analyzing a population from the POEM study (Prospective Investigation of Obesity, Energy and Metabolism, n = 502), reported different findings. In their analysis, gal-3 did not show significant correlations with BMI (β = 0.07; p = 0.095) or with indicators of fat distribution. These findings indicate that gal-3 might have a minor role in adipogenesis regulation itself or may act as a marker of inflammation originating from sources other than adipose tissue.
Table 7 presents a synthetic summary of findings from studies evaluating the association between gal-3 concentration and BMI as well as obesity-related parameters (including visceral fat and WC).
Table 7.
Galectin-3 dependence on BMI and obesity components: a review of selected population-based and clinical studies.
In summary, available data suggest a significant, though variable, association between gal-3 and BMI, with abdominal obesity appearing to be more strongly linked to this biomarker’s concentration than BMI alone. Gal-3 is also associated with inflammatory markers, which may indicate its involvement in the pathophysiology of chronic inflammation accompanying obesity. Further research should focus on elucidating the molecular mechanisms connecting gal-3 with different types of obesity and on evaluating the potential for modulating gal-3 concentration through weight-reduction interventions.
4.5. Cholesterol
The association between gal-3 and lipid metabolism, as well as its potential impact on cholesterol regulation, is the subject of growing interest, particularly in the context of CVDs. This subsection reviews current evidence on the relationship between lipid profile and gal-3 (with particular focus on HDL, LDL, and total cholesterol).
Zeng et al. [114] demonstrated that a high gal-3 concentration combined with low HDL-C significantly strengthens the risk of death and cardiovascular events within one year after ischemic stroke. Notably, despite the lack of a statistical correlation between HDL-C and gal-3 concentrations (r = −0.005, p = 0.776), their synergistic impact on prognosis was evident; the HR for vascular events was 1.92 (95% CI: 1.26–2.94). These findings suggest that gal-3 may function not only as a marker but could also be actively involved in the pathogenesis of atherosclerotic processes, particularly in the presence of lipid abnormalities.
At the experimental level, Tan et al. [115] demonstrated that inhibition of gal-3 expression in a rat model of atherosclerosis resulted in an improved lipid profile (decrease in TG, LDL-C, TC, and elevation in HDL-C), suggesting a potential role of gal-3 in cholesterol metabolism. This mechanism appears to be mediated by the regulation of cholesterol ester hydrolase expression, indicating a possible pathogenic role of gal-3 in lipid accumulation within the vascular wall. A limitation of the study is its animal model nature, which restricts the direct applicability of the results to humans.
In the clinical study by Winter et al. [116] involving participants with premature myocardial infarction, a positive correlation was obtained between gal-3 concentration and non-HDL cholesterol (r = 0.23, p < 0.05) as well as remnant cholesterol (r = 0.21, p < 0.05). This evidence is consistent with the hypothesis that gal-3 may be involved in the pathogenesis of atherogenic dyslipidemia, although no direct correlation was observed with TC or LDL-C. Importantly, in gal-3 knockout animal models, reduced atherosclerotic lesions were observed under a high-fat diet, further supporting its role in atherogenesis.
Analysis of data from the PREVEND cohort [3] indicated that total cholesterol was an independent factor associated with gal-3 concentration in cross-sectional analysis, but it did not significantly affect its variability over time. This may suggest that TC concentration influences the baseline gal-3 concentration but does not determine its dynamic changes, which could be relevant for planning strategies to monitor this biomarker.
It is also worth highlighting the data from Melin et al. [117], who have shown a positive correlation between gal-3 binding protein and TC concentration in patients with type 1 diabetes, particularly in the male subgroup (AOR = 2.3; p < 0.05). This may suggest a link between gal-3 and an unfavorable lipid profile even in younger, metabolically burdened populations.
In summary, the data suggest that galectin-3 may act not only as an inflammatory marker but also as an active participant in the pathophysiology of atherosclerosis, indirectly influencing lipid metabolism—particularly in the context of low HDL-C and elevated non-HDL cholesterol. However, most studies are correlational in nature (cohort studies, animal models), with a limited number of prospective or randomized trials. To date, no definitive evidence has confirmed a direct role of gal-3 on TC concentration, which represents a significant limitation of current knowledge. Translational and interventional research is necessary to evaluate the impact of gal-3 modulation on lipid parameters and long-term cardiovascular risk.
4.6. Blood Pressure
Gal-3 is a key regulator of vascular homeostasis, and its activity involves both inflammatory processes and vascular wall remodeling. Although a direct association between galectin-3 concentration and BP measurements has not been clearly confirmed in clinical studies, numerous experimental data imply that gal-3 may play a role in the pathogenesis of AH through its impact on vascular stiffness, vascular remodeling, and endothelial dysfunction.
Pioneering studies by Nangia-Makker et al. [118] provided some of the first evidence that gal-3 exhibits activity modulating endothelial function and angiogenesis. In both in vivo and in vitro models, it was shown that gal-3 induces morphogenesis of endothelial cells (HUVEC) and the formation of microvessels in Matrigel matrix. Although this mechanism does not directly relate to BP values, it provides key biological foundations for understanding the impact of gal-3 on the microcirculation and potential changes in vascular tone. From the perspective of current data, the angiogenic activity of gal-3 may be relevant to the development of microangiopathy in the course of AH, though this requires further translational research.
In one research paper [119] using a mouse model with angiotensin II infusion, it was demonstrated that gal-3 expression in the endothelium significantly increases under the influence of angiotensin II, a change not observed in gal-3 knockout mice (gal-3−/−). Moreover, the absence of gal-3 protected the animals from an increase in systolic BP and endothelial dysfunction, as well as reduced ROS production, NOX2 expression, and inflammatory markers (IL-6, VCAM-1). These conclusions are based on experimental in vivo models and functional vascular analyses (e.g., response to ACh and SNP), which positions the strength of evidence as solid in the context of preclinical research.
Similar mechanisms were confirmed in the study [120], where in a model of induced hypertension, gal-3 was shown to be involved in the activation of the AMPK/TXNIP and PI3K/Akt pathways, resulting in enhanced vascular fibrosis and vascular wall remodeling. The authors also emphasize that gal-3 affects endothelial homeostasis through interactions with the extracellular matrix and modulation of the inflammatory response. Although the study does not provide numerical data on BP values, the described mechanisms are potentially significant in the context of vascular stiffness—a key component of AH pathophysiology.
Further confirmation of the role of gal-3 as an indicator of endothelial dysfunction comes from the study by Hsu et al. [121], involving individuals with chronic kidney disease (CKD). An inverse correlation was described between gal-3 concentration and the vascular reactivity index (r = −0.439, p < 0.001), independent of eGFR and inflammatory markers. These findings support the concept of gal-3 as a marker of endothelial injury, which over time may lead to AH.
In a study involving patients with AH, gal-3 concentration showed a positive correlation with systolic BP values (r = 0.225, p < 0.05); however, this association was not significant in a multivariate model. A much stronger relationship was observed between gal-3 and indicators of left ventricular remodeling, such as left ventricular mass and index, suggesting the potential usefulness of this biomarker in assessing hypertension-related complications [122]. The authors highlight the higher predictive value of gal-3 compared to BNP (AUC: 0.698 vs. 0.53), emphasizing the diagnostic potential of this molecule, though not necessarily as a direct predictor of BP.
Data from the reviews indicate that gal-3 is involved in modulating the characteristics of vascular smooth muscle cells, facilitating their osteogenic transformation and type I collagen synthesis, which leads to medial layer thickening and increased vascular stiffness [123,124]. Although these findings are molecular and review-based, they are consistent with experimental observations and support the hypothesis of a functional role of gal-3 in the pathogenesis of AH.
Clinical studies focusing specifically on the relationship between gal-3 concentration and BP values are scarce and yield inconclusive results. For instance, in the study [125], involving patients with AH and HF, no differences in gal-3 concentration were described between groups, and its relationship with BP values was not assessed. The absence of numerical data and regression analysis prevents drawing conclusions about the importance of gal-3 as a marker of AH in this population.
Table 8 presents a summary of selected molecular and cellular mechanisms involving gal-3 and their associations with key pathophysiological phenomena leading to increased vascular stiffness, endothelial dysfunction, and elevated vascular resistance.
Table 8.
Overview of biological mechanisms involving galectin-3 leading to vascular dysfunction and elevated blood pressure.
In summary, gal-3 may play a key role in the AH pathogenesis by enhancing inflammatory processes, fibrosis, and vascular remodeling (including its impact on endothelial function). However, clinical evidence regarding its direct association with BP values remains inconclusive. Current knowledge suggests that gal-3 acts more as an indirect mediator of vascular injury and endothelial dysfunction rather than as a biomarker of BP itself.
4.7. Glucose Concentration
Gal-3 is increasingly being applied in research on the pathophysiology of T2DM and its cardiovascular complications. Despite a growing number of reports, the relationship between glucose concentration (both fasting blood glucose, FBG, and glycated hemoglobin, HbA1c) and gal-3 concentration remains inconclusive, with study findings suggesting a stronger association with the chronic effects of hyperglycemia rather than with glucose concentration itself.
In the published review [126], significant expression of gal-3 in the heart and its increase under metabolic conditions such as diabetes were highlighted. The authors emphasize its potential role in diabetic cardiomyopathy (DCM); however, the lack of quantitative data prevents an assessment of the strength of the relationship between glucose concentration and gal-3. In another study [127], despite no differences in HbA1c between patients with and without HFpEF, higher baseline gal-3 concentration was an independent predictor of HF development (AUC = 0.876; p < 0.001). It suggests that gal-3 may reflect fibrosis and inflammatory processes independent of glycemia.
The study by Flores-Ramírez et al. [128] demonstrated significantly higher gal-3 concentration in individuals with T2DM, regardless of left ventricular ejection fraction (LVEF). The observations suggest an association between gal-3 and subclinical cardiac dysfunction. Jin et al. [129] described a significant association between gal-3 and HbA1c (r = 0.217; p < 0.05), with no significant associations with FBG or HOMA-IR. This indicates that gal-3 may be more related to chronic rather than acute dysregulation of glycemia.
In the study by Tan et al. [130], a weak but significant correlation between HbA1c and gal-3 (r = 0.06; p < 0.05) was demonstrated, with no association with T2DM duration or BMI. Gal-3 was described as an independent prognostic factor for mortality risk (HR = 1.17; 95% CI: 1.01–1.35; p < 0.05). These observations support its role as an indicator of a chronic inflammatory-fibrotic process, independent of classical metabolic markers.
Data from Kuzan et al. [131] confirmed significant correlations of gal-3 with FBG (r = 0.39; p < 0.001) and HbA1c (r = 0.267; p < 0.05) in a geriatric population, as well as the impact of metformin treatment on lowering gal-3 concentration (p < 0.05). This suggests the possibility of hypoglycemic pharmacotherapy influencing the expression of this protein, which may have therapeutic implications. Similar findings were reported by Weigert et al. [109], who demonstrated a negative correlation between HbA1c and gal-3 (r = −0.323; p < 0.05) and a metformin-induced reduction of gal-3 expression in vitro.
A comprehensive analysis was conducted by Vora et al. [132], documenting a relationship between baseline gal-3 concentration and the development of T2DM over long-term follow-up (OR = 1.20; 95% CI: 1.00–1.45; p < 0.05). Importantly, this relationship was independent of BMI and not associated with HOMA-IR, while gal-3 correlated with C-peptide and inflammatory markers. This suggests that gal-3 may serve as a marker of β-cell injury or dysfunction rather than insulin resistance alone.
Strong correlations of gal-3 with OGTT, FPG, and HOMA-IR parameters were demonstrated by Yilmaz et al. [133]. The diagnostic value of gal-3 was high (AUC = 0.912 for diabetes; AUC = 0.901 for prediabetes), indicating the potential application of this protein as a marker for early detection of glycemic disorders.
Important experimental data are also provided by Fantauzzi et al. [134], in which mice with gal-3 gene deletion exhibited impaired glucose tolerance, insulin resistance, and β-cell dysfunction. These observations support the hypothesis that gal-3 functions not only as a marker but also as an active participant in metabolic processes, including the regulation of insulin sensitivity and insulin secretion.
Table 9 presents a synthetic summary of clinical and experimental studies analyzing the association between gal-3 concentration and carbohydrate metabolism results such as fasting glycemia, HbA1c, insulin resistance, or the risk of developing T2DM.
Table 9.
Relationships between glucose concentration and gal-3 concentration in individuals with type 2 diabetes mellitus and in experimental models.
In summary, available data indicate that gal-3 is associated with T2DM and its complications. However, the relationship with glucose concentration and HbA1c is inconsistent—some studies report correlations with HbA1c [109,131,135], while others do not confirm significant associations with glycemia [127,128]. The role of gal-3 as a mediator of pathogenic processes is supported by both clinical and experimental studies; however, further prospective analyses are needed to assess its diagnostic and prognostic utility in the context of glucose metabolism disorders.
4.8. Sleep Health
Gal-3 is a biomarker associated with sleep disorders, particularly in terms of sleep quality, inflammation, and cardiovascular consequences related to obstructive sleep apnea (OSA). The sleep aspect is included in the LE8 metric, but not in the older LS7.
In a cross-sectional population-based research paper [108], it was demonstrated that impaired sleep quality was significantly associated with elevated gal-3 concentration, with a significant OR of 1.68 (95% CI: 1.05–2.68) in a fully adjusted model that accounted for factors such as age, gender, smoking, alcohol consumption, AH, BMI, WC, FBG, TG, HDL, and hs-CRP. In contrast, nighttime sleep duration (<7 h vs. ≥7 h) and daytime nap duration (≥60 min vs. <60 min) did not show significant correlation with gal-3 concentration, suggesting that sleep quality, rather than sleep duration, may be crucial for this biomarker. In this study, the mechanisms were linked to activation of the hypothalamic–pituitary–adrenal axis, increased sympathetic nervous system activity, and elevated production of pro-inflammatory cytokines (IL-6, TNF-α). Additionally, the authors noted limitations including the lack of objective sleep assessment methods (e.g., polysomnography) and the cross-sectional design, which prevents definitive causal conclusions.
The relationship between gal-3 and OSA was also thoroughly discussed by Andersen et al. [136], who emphasized the role of this biomarker in inflammatory processes and fibrosis, particularly in the context of sleep fragmentation in patients with OSA. Patients with OSA were found to have substantially elevated gal-3 levels, which correlate with both disease severity and atherosclerotic burden in blood vessels. The authors suggest that gal-3 may influence neuroinflammatory mechanisms related to sleep disorders through microglial activation, involving corticotropin-releasing hormone and autophagy dysregulation. From a clinical standpoint, a key observation is that treatment with CPAP reduced gal-3 concentration, indicating the potential for therapeutic intervention to modulate this biomarker and decrease inflammation.
Clinical studies confirm a strong relationship between OSA severity and gal-3 concentration. Hamdi Pusuroglu et al. [137] found that mean gal-3 concentration significantly increased with the severity of OSA (mild: 4.3 ± 1200 ng/mL, moderate: 5.1 ± 1.4 ng/mL, severe: 6.4 ± 1700 ng/mL; p < 0.001). In addition, gal-3 concentration positively correlated with the apnea–hypopnea index (r = 0.337; p < 0.001) and the number of atherosclerotic plaques in coronary arteries (r = 0.417; p < 0.001). In multivariate analysis, gal-3 was an independent predictor of OSA severity (OR = 2.329; 95% CI: 1.558–3.480) and coronary artery atherosclerosis (OR = 2.175; 95% CI: 1.425–3.320). The authors associate this with increased inflammation, hypoxia, and oxidative stress, highlighting the potential of gal-3 as a cardiovascular risk biomarker in individuals with OSA.
Singh et al. [138] expand on these observations by highlighting sex differences in the correlation between gal-3 and OSA. Median gal-3 concentrations were significantly higher in severe and moderate OSA (7.63 ng/mL, n = 471) compared to individuals without or with mild OSA (6.53 ng/mL; p < 0.001). Multivariate analysis showed a significant association, particularly in women (β = 0.14; p < 0.001), whereas in men, these results were not significant (p = 0.86). The mechanism underlying these sex-specific differences remains unclear, although differences in inflammatory responses, microcirculatory function, and the effects of hypoxia and hypothalamic–pituitary–adrenal axis activation have been suggested. However, the authors note limitations due to the cross-sectional design of the study and the relatively small number of men in the analyzed population.
Khalaji et al. [139] and Zong et al. [140] complement these findings by providing context on the neurocognitive consequences of OSA related to gal-3 concentration. The studies suggest that elevated gal-3 concentration in individuals with severe OSA reflects intensified neuroinflammatory processes, oxidative stress, and reduced cognitive function (significantly lower Montreal Cognitive Assessment scores: 22.41 ± 2.56 vs. 27.11 ± 1.32 in the reference group; p < 0.001). After CPAP therapy, a significant decrease in gal-3 concentration was observed (p < 0.001), along with improvements in sleep parameters and cognitive performance (24.86 ± 1.81 in the score; p < 0.05), confirming the role of sleep quality in the regulation of this biomarker.
Table 10 presents a synthetic summary of current literature data regarding the relationship between gal-3 and sleep quality parameters, the severity and presence of OSA, as well as neuroinflammatory and CVH consequences. Both original and review studies are included, indicating the possible application of gal-3 as a biomarker of chronic inflammation, fibrosis, and risk of complications in the context of sleep disorders. A major limitation of most studies is their cross-sectional design and the lack of objective sleep assessment methods, which highlights the need for further prospective research using techniques such as polysomnography.
Table 10.
Sleep health as a biomarker of sleep quality and obstructive sleep apnea: summary of findings from observational and review studies.
In summary, data from available studies indicate that gal-3 is significantly associated with sleep quality, OSA severity, and the inflammatory and neurocognitive consequences of this disorder. However, due to methodological limitations, including the lack of objective sleep assessment tools and the cross-sectional design of the studies, further analyses using precise diagnostic methods are necessary to gain a more comprehensive understanding of this relationship.
5. Limitations and Future Directions
This review is narrative in nature, which involves limited standardization in the process of data selection and analysis. Although the structure follows the SANRA scale, subjectivity in the selection of literature and interpretation of results cannot be excluded. Some of the referenced data come from preclinical or observational studies, which limits the possibility of direct translation into clinical practice. Many findings are based on correlations without clear confirmation of causal relationships. Additionally, despite promising data regarding gal-3 inhibitors, their use in clinical practice remains experimental.
Despite its promising role, gal-3 has several important limitations that restrict its clinical utility. Firstly, gal-3 lacks high specificity: elevated concentrations are not exclusive to HF or fibrosis-related pathologies but are also observed in chronic kidney disease, malignancies, and systemic inflammatory conditions. This non-specificity may confound clinical interpretation. Secondly, a significant inter-assay variability exists among commercially available gal-3 measurement kits, and there is no universally accepted standard for its quantification, which complicates comparisons between studies and clinical implementation. Moreover, even mild renal impairment independently elevates circulating gal-3 concentrations, thus potentially acting as a confounding factor when interpreting biomarker results in patients with concurrent cardiac and renal dysfunction. These limitations must be carefully considered when attempting to translate gal-3 measurement into routine clinical practice.
In the future, attention should be focused on:
- Prospective studies evaluating the predictive value of gal-3 in various risk populations;
- Interventional trials assessing the impact of lifestyle modifications on gal-3 concentration and related clinical outcomes;
- Validation of gal-3 as a marker of therapeutic response in CVDs, particularly in HF and atrial fibrillation;
- Assessment of its role as a marker of environmental and lifestyle effects in the context of LE8/LS7.
The interplay between lifestyle, occupational exposures, and emerging molecular biomarkers, including gal-3, warrants further translational research, as also emphasized in recent proposals for precision diagnostics in AH [1].
From a clinical perspective, in the near future, gal-3 may find application primarily in specialized cardiology clinics, e.g., in assessing the severity of HF with a mixed etiology (where a fibrotic component is suspected, as in HFpEF), or in patients with recurrent hospitalizations to evaluate their long-term prognosis. It may also be useful in the context of screening in at-risk populations, for instance, in patients with long-standing AH or T2DM, where elevated gal-3 concentration could indicate the onset of organ damage and prompt more aggressive risk factor control. However, before this becomes routine, it is necessary to establish clear management protocols for defined gal-3 concentrations (e.g., whether >17 ng/mL is considered high-risk, requiring therapy intensification).
In summary, gal-3 is a promising cardio-metabolic indicator that enhances our ability to assess patients beyond traditional risk factors. It serves as a link between lifestyle and risk factors (as captured in LS7/LE8) and actual organ damage; increased gal-3 concentration suggests that long-term exposure to risk factors has already translated into tangible pathological changes (fibrosis, remodeling). The implementation of gal-3 testing in daily practice should go hand-in-hand with staff education regarding result interpretation and the limitations of the test. Its clinical application may, in the future, facilitate the identification of patients who require more intensive preventive or therapeutic interventions. A vision of targeted therapy is also on the horizon. If the development of gal-3 inhibitors proves successful, we may one day be able not only to measure but also to modulate this parameter, slowing the progression of HF in our patients. As of today, gal-3 is an important piece of the puzzle, but it does not replace classical methods; it rather complements the diagnostic and prognostic profile of the patient. Although the path toward full clinical implementation of gal-3 is still ahead of us, this biomarker represents one of the most promising links between precision medicine and primary prevention of chronic diseases. Clinicians should stay updated with new guidelines and research findings, as the role of galectin-3 may evolve in the coming years as new evidence emerges.
6. Conclusions
The complex interplay between gal-3 and individual components of Life’s Essential 8 metrics is summarized in Figure 3. This overview highlights potential mechanisms linking lifestyle, metabolic risk factors, and gal-3-driven cardiovascular pathology.
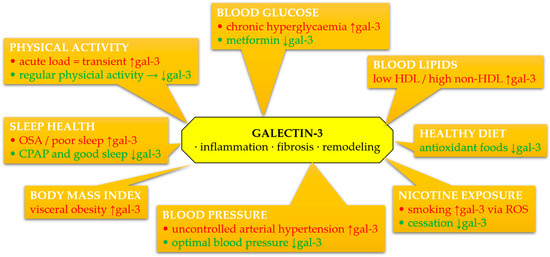
Figure 3.
Schematic summary of associations between circulating galectin-3 (gal-3) and components of Life’s Essential 8 and Life’s Simple 7 metrics. Galectin-3 levels may be modulated by lifestyle and clinical factors, contributing to inflammation, fibrosis, oxidative stress, and extracellular matrix remodeling. Green color denotes favorable effects (decrease in gal-3), red color denotes adverse effects (increase in gal-3). Abbreviations: CPAP, continuous positive airway pressure; gal-3, galectin-3; HDL, high-density lipoprotein; OSA, obstructive sleep apnea; ROS, reactive oxygen species.
Galectin-3 is increasingly recognized as a pivotal mediator in the pathophysiology of cardiovascular diseases, particularly through its involvement in inflammation, fibrosis, and myocardial remodeling. Available data indicate that gal-3 is not only a marker of these processes but may also actively participate in their modulation. Its concentration correlates with various CVH parameters assessed within the LS7 and LE8 frameworks, although it is not directly included in these scales. The integration of gal-3 into the context of primary and secondary prevention requires further validation, but existing evidence suggests the possibility of using this molecule as a complementary biomarker of cardiovascular risk and response to lifestyle interventions.
Author Contributions
Conceptualization, A.M. and P.G.; resources, A.M. and P.P.; writing—original draft preparation, A.M. and P.P.; writing—review and editing, A.M.; supervision, R.P. and P.G.; funding acquisition, P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ACC | American College of Cardiology |
| AECs | Airway epithelial cells |
| AHA | American Heart Association |
| AH | Arterial hypertension |
| AHI | Apnea-hypopnea index |
| ALEs | Advanced lipoxidation end-products |
| AMPK | AMP-activated protein kinase |
| AOR | Adjusted odds ratio |
| ARIC | Atherosclerosis Risk in Communities Study |
| BMI | Body mass index |
| BNP | B-type natriuretic peptide |
| BP | Blood pressure |
| CAD | Coronary artery disease |
| CD68 | Cluster of differentiation 68 |
| CHF | Chronic heart failure |
| CI | Confidence interval |
| CKD | Chronic kidney disease |
| COPD | Chronic obstructive pulmonary disease |
| CPAP | Continuous positive airway pressure |
| CR | Cardiac rehabilitation |
| CSE | Cigarette smoke extract |
| CVD | Cardiovascular disease |
| CVH | Cardiovascular health |
| DAMP | Damage-associated molecular pattern |
| DCM | Diabetic cardiomyopathy |
| DEX | Dexpanthenol |
| ECHO | Echocardiographic |
| ECM | Extracellular matrix |
| ECV | Extracellular volume |
| EGFR | Epidermal growth factor receptor |
| EMT | Epithelial–mesenchymal transition |
| eNOS | Endothelial nitric oxide synthase |
| EPCs | Endothelial progenitor cells |
| ERC | Elastin receptor complex |
| ESC | European Society of Cardiology |
| FAK kinases | Focal adhesion kinase kinases |
| FBG | Fasting blood glucose |
| FDA | Food and Drug Administration |
| FHS | Framingham Heart Study |
| Gal-3 | Galectin-3 |
| GSH-Px | Glutathione peroxidase |
| HbA1C | Hemoglobin A1c (glycated hemoglobin) |
| HCD | Hypercaloric diet |
| HF | Heart failure |
| HFD | High-fat diet |
| HFpEF | Heart failure with preserved ejection fraction |
| HFSA | Heart Failure Society of America |
| HIIT | High-intensity interval training |
| HOMA-IR | Homeostasis Model Assessment of Insulin Resistance |
| HR | Hazard ratio |
| hsCRP | High-sensitivity C-reactive protein |
| HUVECs | Human umbilical vein endothelial cells |
| HVS | Hepatic vein serum |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IF | Intermittent fasting |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| LCN2 | Lipocalin-2 |
| LE8 | Life’s Essential 8 |
| LGALS3BP | Galectin-3 binding protein |
| LLPS | Liquid–liquid phase separation |
| LPC | Lysophosphatidylcholine |
| LPS | Lipopolysaccharide |
| LS7 | Life’s Simple 7 |
| LV | Left ventricle |
| LVEF | Left ventricular ejection fraction |
| LVH | Left ventricular hypertrophy |
| MCP | Modified citrus pectin |
| MDA | Malondialdehyde |
| MIACT | Moderate-intensity aerobic continuous training |
| MRI | Magnetic resonance imaging |
| MUC1-C/EGFR | Mucin 1 C-terminal subunit/Epidermal Growth Factor Receptor |
| NAC | N-acetylcysteine |
| NADPH oxidase 2 | Nicotinamide adenine dinucleotide phosphate oxidase 2 |
| NASH | Nonalcoholic steatohepatitis |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| NWGR | Asparagine-Tryptophan-Glycine-Arginine |
| NT-proBNP | N-terminal pro–B-type natriuretic peptide |
| OGTT | Oral glucose tolerance test |
| OR | Odds ratio |
| OSA | Obstructive sleep apnea |
| oxLDL | Oxidized low-density lipoprotein |
| PI3K | Phosphoinositide 3-kinase |
| POEM | Prospective Investigation of Obesity, Energy and Metabolism |
| PREVEND | Prevention of Renal and Vascular End-stage Disease |
| PUFA | Polyunsaturated fatty acids |
| PVS | Portal vein serum |
| Ref. | References |
| ROS | Reactive oxygen species |
| RUNX-2 | Runt-related transcription factor 2 |
| RV | Right ventricular |
| RVEF | Right ventricular ejection fraction |
| SANRA | Scale for the Assessment of Narrative Review Articles |
| SAT | Subcutaneous adipose tissue |
| SBP | Systolic blood pressure |
| SGLT2i | Sodium-glucose co-transporter 2 inhibitors |
| SIRT1 | Sirtuin 1 |
| SOD | Superoxide dismutase |
| SNP | Sodium nitroprusside |
| SPPB | Short physical performance battery |
| SR proteins | Serine/arginine-rich proteins |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| STEMI | ST-elevation myocardial infarction |
| SVS | Systemic venous serum |
| T2DM | Type 2 diabetes mellitus |
| TC | Total cholesterol |
| TG | Triglycerides |
| TGF-β | Transforming growth factor beta. |
| TLR4 | Toll-like receptor 4 |
| TNF-α | Tumor necrosis factor alpha |
| TOS | Total oxidative status |
| VAT | Visceral adipose tissue |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VCID | Vascular cognitive impairment and dementia |
| VEGFR2 | Vascular endothelial growth factor receptor 2 |
| VO2 | Oxygen uptake |
| VSMCs | Vascular smooth muscle cells |
| WAT | White adipose tissue |
| WC | Waist circumference |
| WONDERFUL | Weekly ONe-Day WatER-only Fasting InterventionaL trial |
| WT | Wild type |
| YAP | Yes-associated protein |
| α9nAChR | α9 nicotinic acetylcholine receptor |
| ↑ | Increase |
| ↓ | Decrease |
References
- Charkiewicz, A.E. A Proposal for Research Involving New Biomarkers of Hypertension, Lifestyle, and Environmental Exposure. Curr. Issues Mol. Biol. 2025, 47, 206. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Bhambhani, V.; Christenson, R.H.; Meijers, W.C.; De Boer, R.A.; Levy, D.; Larson, M.G.; Ho, J.E. Longitudinal Change in Galectin-3 and Incident Cardiovascular Outcomes. J. Am. Coll. Cardiol. 2018, 72, 3246–3254. [Google Scholar] [CrossRef]
- Van Der Velde, A.R.; Meijers, W.C.; Van Den Heuvel, E.R.; Bakker, S.J.; Van Gilst, W.H.; Van Der Harst, P.; Hillege, H.; De Boer, R.A. Determinants of Temporal Changes in Galectin-3 Level in the General Population: Data of PREVEND. Int. J. Cardiol. 2016, 222, 385–390. [Google Scholar] [CrossRef]
- Díaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediat. Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef]
- Suarez, G.; Meyerrose, G. Heart Failure and Galectin 3. Ann. Transl. Med. 2014, 2, 86. [Google Scholar] [CrossRef] [PubMed]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E.; Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA Guideline for the Management of Heart Failure. J. Am. Coll. Cardiol. 2013, 62, e147–e239. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Cai, K.; Xu, C.; Zhan, Q.; Xu, X.; Xu, D.; Zeng, Q. Prognostic Value of Serum Galectin-3 in Chronic Heart Failure: A Meta-Analysis. Front. Cardiovasc. Med. 2022, 9, 783707. [Google Scholar] [CrossRef]
- Funasaka, T.; Raz, A.; Nangia-Makker, P. Nuclear Transport of Galectin-3 and Its Therapeutic Implications. Semin. Cancer Biol. 2014, 27, 30–38. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Qiu, D.-C.; Chang, W.-H.; Yeh, Y.-Q.; Jeng, U.-S.; Liu, F.-T.; Huang, J. The Intrinsically Disordered N-Terminal Domain of Galectin-3 Dynamically Mediates Multisite Self-Association of the Protein through Fuzzy Interactions. J. Biol. Chem. 2017, 292, 17845–17856. [Google Scholar] [CrossRef]
- Sciacchitano, S.; Lavra, L.; Morgante, A.; Ulivieri, A.; Magi, F.; De Francesco, G.; Bellotti, C.; Salehi, L.; Ricci, A. Galectin-3: One Molecule for an Alphabet of Diseases, from A to Z. Int. J. Mol. Sci. 2018, 19, 379. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.M.; Hong, Y.; Labarthe, D.; Mozaffarian, D.; Appel, L.J.; Van Horn, L.; Greenlund, K.; Daniels, S.; Nichol, G.; Tomaselli, G.F.; et al. Defining and Setting National Goals for Cardiovascular Health Promotion and Disease Reduction: The American Heart Association’s Strategic Impact Goal Through 2020 and Beyond. Circulation 2010, 121, 586–613. [Google Scholar] [CrossRef] [PubMed]
- Gaye, B.; Lloyd-Jones, D.M. Primordial Prevention of Cardiovascular Disease: Several Challenges Remain. Int. J. Cardiol. 2019, 274, 379–380. [Google Scholar] [CrossRef]
- Banach, M.; Burchardt, P.; Chlebus, K.; Dobrowolski, P.; Dudek, D.; Dyrbuś, K.; Gąsior, M.; Jankowski, P.; Jóźwiak, J.; Kłosiewicz-Latoszek, L.; et al. WYTYCZNE PTL/KLRwP/PTK/ PTDL/PTD/PTNT DIAGNOSTYKI I LECZENIA ZABURZEŃ LIPIDOWYCH W POLSCE 2021. Nadciśnienie Tętnicze Prakt. 2021, 7, 113–222. [Google Scholar]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; De Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the Management of Arterial Hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef] [PubMed]
- Araszkiewicz, A.; Bandurska-Stankiewicz, E.; Borys, S.; Budzyński, A.; Cyganek, K.; Cypryk, K.; Czech, A.; Czupryniak, L.; Drzewoski, J.; Dzida, G.; et al. 2021 Guidelines on the Management of Patients with Diabetes. A Position of Diabetes Poland. Clin. Diabetol. 2021, 10, 1–113. [Google Scholar] [CrossRef]
- Patel, R.; Peesay, T.; Krishnan, V.; Wilcox, J.; Wilsbacher, L.; Khan, S.S. Prioritizing the Primary Prevention of Heart Failure: Measuring, Modifying and Monitoring Risk. Prog. Cardiovasc. Dis. 2024, 82, 2–14. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.M.; Allen, N.B.; Anderson, C.A.M.; Black, T.; Brewer, L.C.; Foraker, R.E.; Grandner, M.A.; Lavretsky, H.; Perak, A.M.; Sharma, G.; et al. Life’s Essential 8: Updating and Enhancing the American Heart Association’s Construct of Cardiovascular Health: A Presidential Advisory from the American Heart Association. Circulation 2022, 146, e18–e43. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Tian, P.; Li, L.; Qu, Q. Comparison of the Associations between Life’s Essential 8 and Life’s Simple 7 with Stroke: NHANES 1999–2018. J. Stroke Cerebrovasc. Dis. 2025, 34, 108238. [Google Scholar] [CrossRef]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—A Scale for the Quality Assessment of Narrative Review Articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef]
- Wasserstein, R.L.; Lazar, N.A. The ASA Statement on P-Values: Context, Process, and Purpose. Am. Stat. 2016, 70, 129–133. [Google Scholar] [CrossRef]
- Kyriacou, D.N. The Enduring Evolution of the P Value. JAMA 2016, 315, 1113. [Google Scholar] [CrossRef] [PubMed]
- Harrington, D.; D’Agostino, R.B.; Gatsonis, C.; Hogan, J.W.; Hunter, D.J.; Normand, S.-L.T.; Drazen, J.M.; Hamel, M.B. New Guidelines for Statistical Reporting in the Journal. N. Engl. J. Med. 2019, 381, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Martuszewski, A.; Paluszkiewicz, P.; Poręba, R.; Gać, P. Galectin-3 in Cardiovascular Health: A Narrative Review Based on the Life’s Essential 8 and Life’s Simple 7 Frameworks. 2025. Available online: https://osf.io/xfwr5/resources (accessed on 3 May 2025). [CrossRef]
- Sato, S. Why Does Galectin-3 Have a Unique Intrinsically Disordered Region? Glycoforum 2023, 26, A8. [Google Scholar]
- Liu, F. Intracellular Functions of Galectins. Biochim. Biophys. Acta (BBA) Gen. Subj. 2002, 1572, 263–273. [Google Scholar] [CrossRef]
- Lin, H.-M. Galectin-3 Mediates Genistein-Induced G2/M Arrest and Inhibits Apoptosis. Carcinogenesis 2000, 21, 1941–1945. [Google Scholar] [CrossRef]
- Akahani, S.; Nangia-Makker, P.; Inohara, H.; Kim, H.R.; Raz, A. Galectin-3: A Novel Antiapoptotic Molecule with a Functional BH1 (NWGR) Domain of Bcl-2 Family. Cancer Res. 1997, 57, 5272–5276. [Google Scholar]
- Harazono, Y.; Kho, D.H.; Balan, V.; Nakajima, K.; Zhang, T.; Hogan, V.; Raz, A. Galectin-3 Leads to Attenuation of Apoptosis through Bax Heterodimerization in Human Thyroid Carcinoma Cells. Oncotarget 2014, 5, 9992–10001. [Google Scholar] [CrossRef]
- Yang, R.Y.; Hsu, D.K.; Liu, F.T. Expression of Galectin-3 Modulates T-Cell Growth and Apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 6737–6742. [Google Scholar] [CrossRef]
- Fukumori, T.; Takenaka, Y.; Oka, N.; Yoshii, T.; Hogan, V.; Inohara, H.; Kanayama, H.; Kim, H.-R.C.; Raz, A. Endogenous Galectin-3 Determines the Routing of CD95 Apoptotic Signaling Pathways. Cancer Res. 2004, 64, 3376–3379. [Google Scholar] [CrossRef]
- Liu, L.; Sakai, T.; Sano, N.; Fukui, K. Nucling Mediates Apoptosis by Inhibiting Expression of Galectin-3 through Interference with Nuclear Factor kappaB Signalling. Biochem. J. 2004, 380, 31–41. [Google Scholar] [CrossRef]
- Yu, F.; Finley, R.L.; Raz, A.; Kim, H.-R.C. Galectin-3 Translocates to the Perinuclear Membranes and Inhibits Cytochrome c Release from the Mitochondria. J. Biol. Chem. 2002, 277, 15819–15827. [Google Scholar] [CrossRef] [PubMed]
- Matarrese, P.; Fusco, O.; Tinari, N.; Natoli, C.; Liu, F.T.; Semeraro, M.L.; Malorni, W.; Iacobelli, S. Galectin-3 Overexpression Protects from Apoptosis by Improving Cell Adhesion Properties. Int. J. Cancer 2000, 85, 545–554. [Google Scholar] [CrossRef]
- Califice, S.; Castronovo, V.; Bracke, M.; Van Den Brûle, F. Dual Activities of Galectin-3 in Human Prostate Cancer: Tumor Suppression of Nuclear Galectin-3 vs Tumor Promotion of Cytoplasmic Galectin-3. Oncogene 2004, 23, 7527–7536. [Google Scholar] [CrossRef] [PubMed]
- Califice, S.; Castronovo, V.; Van Den Brûle, F. Galectin-3 and Cancer (Review). Int. J. Oncol. 2004, 25, 983–992. [Google Scholar]
- Moisa, A.; Fritz, P.; Eck, A.; Wehner, H.-D.; Mürdter, T.; Simon, W.; Gabius, H.-J. Growth/Adhesion-Regulatory Tissue Lectin Galectin-3: Stromal Presence but Not Cytoplasmic/Nuclear Expression in Tumor Cells as a Negative Prognostic Factor in Breast Cancer. Anticancer Res. 2007, 27, 2131–2139. [Google Scholar]
- Haudek, K.C.; Spronk, K.J.; Voss, P.G.; Patterson, R.J.; Wang, J.L.; Arnoys, E.J. Dynamics of Galectin-3 in the Nucleus and Cytoplasm. Biochim. Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 181–189. [Google Scholar] [CrossRef]
- Hughes, R. Secretion of the Galectin Family of Mammalian Carbohydrate-Binding Proteins. Biochim. Biophys. Acta (BBA) Gen. Subj. 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Jones, J.L.; Saraswati, S.; Block, A.S.; Lichti, C.F.; Mahadevan, M.; Diekman, A.B. Galectin-3 Is Associated with Prostasomes in Human Semen. Glycoconj. J. 2010, 27, 227–236. [Google Scholar] [CrossRef]
- Nabi, I.R.; Shankar, J.; Dennis, J.W. The Galectin Lattice at a Glance. J. Cell Sci. 2015, 128, 2213–2219. [Google Scholar] [CrossRef]
- Markowska, A.I.; Liu, F.-T.; Panjwani, N. Galectin-3 Is an Important Mediator of VEGF- and bFGF-Mediated Angiogenic Response. J. Exp. Med. 2010, 207, 1981–1993. [Google Scholar] [CrossRef]
- Merlin, J.; Stechly, L.; De Beaucé, S.; Monté, D.; Leteurtre, E.; Van Seuningen, I.; Huet, G.; Pigny, P. Galectin-3 Regulates MUC1 and EGFR Cellular Distribution and EGFR Downstream Pathways in Pancreatic Cancer Cells. Oncogene 2011, 30, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Knights, A.J.; Yik, J.J.; Mat Jusoh, H.; Norton, L.J.; Funnell, A.P.W.; Pearson, R.C.M.; Bell-Anderson, K.S.; Crossley, M.; Quinlan, K.G.R. Krüppel-like Factor 3 (KLF3/BKLF) Is Required for Widespread Repression of the Inflammatory Modulator Galectin-3 (Lgals3). J. Biol. Chem. 2016, 291, 16048–16058. [Google Scholar] [CrossRef] [PubMed]
- Sygitowicz, G.; Maciejak-Jastrzębska, A.; Sitkiewicz, D. The Diagnostic and Therapeutic Potential of Galectin-3 in Cardiovascular Diseases. Biomolecules 2021, 12, 46. [Google Scholar] [CrossRef]
- Kasai, K. The Keyhole of Galectin Is Made Loose, and the Inserted Key Easily Comes Out. Glycoforum 2020, 23, A17. [Google Scholar]
- Miyanishi, N.; Nishi, N.; Abe, H.; Kashio, Y.; Shinonaga, R.; Nakakita, S.; Sumiyoshi, W.; Yamauchi, A.; Nakamura, T.; Hirashima, M.; et al. Carbohydrate-Recognition Domains of Galectin-9 Are Involved in Intermolecular Interaction with Galectin-9 Itself and Other Members of the Galectin Family. Glycobiology 2007, 17, 423–432. [Google Scholar] [CrossRef]
- Tian, L.; Chen, K.; Han, Z. Correlation between Galectin-3 and Adverse Outcomes in Myocardial Infarction Patients: A Meta-Analysis. Cardiol. Res. Pract. 2020, 2020, 7614327. [Google Scholar] [CrossRef]
- Fortuna-Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S.G. Extracellular Galectin-3 in Tumor Progression and Metastasis. Front. Oncol. 2014, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming Growth Factor (TGF)-β Signaling in Cardiac Remodeling. J. Mol. Cell. Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef]
- Loffredo, F.S.; Nikolova, A.P.; Pancoast, J.R.; Lee, R.T. Heart Failure With Preserved Ejection Fraction: Molecular Pathways of the Aging Myocardium. Circ. Res. 2014, 115, 97–107. [Google Scholar] [CrossRef]
- Málek, F. Impact of galectin 3 as myofibrosis marker in clinical cardiology. Vnitr. Lek. 2014, 60, 327–330. [Google Scholar]
- Nangia-Makker, P.; Hogan, V.; Raz, A. Galectin-3 and Cancer Stemness. Glycobiology 2018, 28, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Saccon, F.; Gatto, M.; Ghirardello, A.; Iaccarino, L.; Punzi, L.; Doria, A. Role of Galectin-3 in Autoimmune and Non-Autoimmune Nephropathies. Autoimmun. Rev. 2017, 16, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, X.; Ma, L.; Zhuang, Y.; Wei, Y.; Zhang, L.; Jin, S.; Liang, W.; Shen, X.; Li, C.; et al. Galectin-3 May Serve as a Marker for Poor Prognosis in Colorectal Cancer: A Meta-Analysis. Pathol. Res. Pract. 2019, 215, 152612. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Ilarregui, J.M. Conveying Glycan Information into T-cell Homeostatic Programs: A Challenging Role for Galectin-1 in Inflammatory and Tumor Microenvironments. Immunol. Rev. 2009, 230, 144–159. [Google Scholar] [CrossRef]
- Pruc, M.; Gaca, Z.; Swieczkowski, D.; Kubica, J.; Galwankar, S.; Salak, A.; Szarpak, L. A Systematic Review and Meta-Analysis of the Diagnostic Value of Galectin-3 in Acute Coronary Syndrome. J. Clin. Med. 2024, 13, 4504. [Google Scholar] [CrossRef]
- Gong, M.; Cheung, A.; Wang, Q.; Li, G.; Goudis, C.A.; Bazoukis, G.; Lip, G.Y.H.; Baranchuk, A.; Korantzopoulos, P.; Letsas, K.P.; et al. Galectin-3 and Risk of Atrial Fibrillation: A Systematic Review and Meta-analysis. Clin. Lab. Anal. 2020, 34, e23104. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.S.; Liu, E.; Paniagua, S.M.; Sarma, A.A.; Zampierollo, G.; López, B.; Díez, J.; Wang, T.J.; Ho, J.E. Galectin-3 Inhibition With Modified Citrus Pectin in Hypertension. JACC Basic. Transl. Sci. 2021, 6, 12–21. [Google Scholar] [CrossRef]
- Arangalage, D.; Nguyen, V.; Robert, T.; Melissopoulou, M.; Mathieu, T.; Estellat, C.; Codogno, I.; Huart, V.; Duval, X.; Cimadevilla, C.; et al. 0574: Determinants and Prognostic Value of Galectin-3 in Patients with Aortic Valve Stenosis—The COFRASA-GENERAC Study. Arch. Cardiovasc. Dis. Suppl. 2016, 8, 59. [Google Scholar] [CrossRef]
- Nangia-Makker, P.; Hogan, V.; Honjo, Y.; Baccarini, S.; Tait, L.; Bresalier, R.; Raz, A. Inhibition of Human Cancer Cell Growth and Metastasis in Nude Mice by Oral Intake of Modified Citrus Pectin. JNCI J. Natl. Cancer Inst. 2002, 94, 1854–1862. [Google Scholar] [CrossRef]
- Jiménez-González, S.; Delgado-Valero, B.; Islas, F.; Romero-Miranda, A.; Luaces, M.; Ramchandani, B.; Cuesta-Corral, M.; Montoro-Garrido, A.; Martínez-Martínez, E.; Cachofeiro, V. The Detrimental Role of Galectin-3 and Endoplasmic Reticulum Stress in the Cardiac Consequences of Myocardial Ischemia in the Context of Obesity. FASEB J. 2024, 38, e23818. [Google Scholar] [CrossRef]
- Kirk, J.A.; Frangogiannis, N.G. Galectin-3 in the Pathogenesis of Heart Failure: A Causative Mediator or Simply a Biomarker? Am. J. Physiol.-Heart Circ. Physiol. 2018, 314, H1256–H1258. [Google Scholar] [CrossRef] [PubMed]
- Kram, M. Galectin-3 Inhibition as a Potential Therapeutic Target in Non-Alcoholic Steatohepatitis Liver Fibrosis. World J. Hepatol. 2023, 15, 201–207. [Google Scholar] [CrossRef]
- Aslanis, V.; Slack, R.J.; MacKinnon, A.C.; McClinton, C.; Tantawi, S.; Gravelle, L.; Nilsson, U.J.; Leffler, H.; Brooks, A.; Khindri, S.K.; et al. Safety and Pharmacokinetics of GB1211, an Oral Galectin-3 Inhibitor: A Single- and Multiple-Dose First-in-Human Study in Healthy Participants. Cancer Chemother. Pharmacol. 2023, 91, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Perea, R.J.; Morales-Ruiz, M.; Ortiz-Perez, J.T.; Bosch, X.; Andreu, D.; Borras, R.; Acosta, J.; Penela, D.; Prat-González, S.; De Caralt, T.M.; et al. Utility of Galectin-3 in Predicting Post-Infarct Remodeling after Acute Myocardial Infarction Based on Extracellular Volume Fraction Mapping. Int. J. Cardiol. 2016, 223, 458–464. [Google Scholar] [CrossRef]
- Severino, P.; D’Amato, A.; Prosperi, S.; Fanisio, F.; Birtolo, L.I.; Costi, B.; Netti, L.; Chimenti, C.; Lavalle, C.; Maestrini, V.; et al. Myocardial Tissue Characterization in Heart Failure with Preserved Ejection Fraction: From Histopathology and Cardiac Magnetic Resonance Findings to Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 7650. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Dong, G.; Liu, J.; Shuang, X.; Liu, C.; Yang, C.; Qing, W.; Qiao, W. Clinical Implications of Plasma Galectin-3 in Heart Failure With Preserved Ejection Fraction: A Meta-Analysis. Front. Cardiovasc. Med. 2022, 9, 854501. [Google Scholar] [CrossRef]
- Radziwolek, L.; Radunski, U.K.; Koopmann, K.; Bohnen, S.; Zeller, T.; Lund, G.; Krull, A.D.; Hauschild, N.; Stehning, C.; Adam, G.; et al. Myocardial Injury and Fibrogenesis: Extracellular Volume Expansion Is Associated with Elevated Galectin-3 Levels in Patients with Myocarditis. J. Cardiovasc. Magn. Reson. 2014, 16, P290. [Google Scholar] [CrossRef]
- De Boer, R.A.; Verweij, N.; Van Veldhuisen, D.J.; Westra, H.-J.; Bakker, S.J.L.; Gansevoort, R.T.; Muller Kobold, A.C.; Van Gilst, W.H.; Franke, L.; Leach, I.M.; et al. A Genome-Wide Association Study of Circulating Galectin-3. PLoS ONE 2012, 7, e47385. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Zhu, J.; Liu, Y.; Zhuang, L.; Zhang, Z.; Zhong, D.; Zhang, W.; Lai, D. Exploring the Causal Effects of Circulating ST2 and Galectin-3 on Heart Failure Risk: A Mendelian Randomization Study. Front. Cardiovasc. Med. 2022, 9, 868749. [Google Scholar] [CrossRef]
- Gou, Y.; Chen, M.; Zhu, Z.; Cui, C. Galectin-3 and Peripheral Artery Disease: A Mendelian Randomization Study. Front. Cardiovasc. Med. 2024, 10, 1279396. [Google Scholar] [CrossRef]
- Henry, A.; Gordillo-Marañón, M.; Finan, C.; Schmidt, A.F.; Ferreira, J.P.; Karra, R.; Sundström, J.; Lind, L.; Ärnlöv, J.; Zannad, F.; et al. Therapeutic Targets for Heart Failure Identified Using Proteomics and Mendelian Randomization. Circulation 2022, 145, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, S.M.; Norwitz, E.R.; Kim, J.H.; Jung, Y.M.; Park, C.-W.; Jun, J.K.; Lee, D.; Jin, Y.; Kim, S.; et al. Placental Expression Quantitative Trait Loci in an East Asian Population. Hum. Genet. Genom. Adv. 2024, 5, 100276. [Google Scholar] [CrossRef]
- De Boer, R.A.; Van Veldhuisen, D.J.; Gansevoort, R.T.; Muller Kobold, A.C.; Van Gilst, W.H.; Hillege, H.L.; Bakker, S.J.L.; Van Der Harst, P. The Fibrosis Marker Galectin-3 and Outcome in the General Population. J. Intern. Med. 2012, 272, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.R.; Agraval, H.; Yadav, U.C.S. Cigarette Smoke Induces Epithelial-to-Mesenchymal Transition, Stemness, and Metastasis in Lung Adenocarcinoma Cells via Upregulated RUNX-2/Galectin-3 Pathway. Life Sci. 2023, 318, 121480. [Google Scholar] [CrossRef] [PubMed]
- Guha, P.; Bandyopadhyaya, G.; Polumuri, S.K.; Chumsri, S.; Gade, P.; Kalvakolanu, D.V.; Ahmed, H. Nicotine Promotes Apoptosis Resistance of Breast Cancer Cells and Enrichment of Side Population Cells with Cancer Stem Cell-like Properties via a Signaling Cascade Involving Galectin-3, A9 Nicotinic Acetylcholine Receptor and STAT3. Breast Cancer Res. Treat. 2014, 145, 5–22. [Google Scholar] [CrossRef]
- Feng, W.; Wu, X.; Li, S.; Zhai, C.; Wang, J.; Shi, W.; Li, M. Association of Serum Galectin-3 with the Acute Exacerbation of Chronic Obstructive Pulmonary Disease. Med. Sci. Monit. 2017, 23, 4612–4618. [Google Scholar] [CrossRef]
- Pei, C.; Wang, X.; Lin, Y.; Fang, L.; Meng, S. Inhibition of Galectin-3 Alleviates Cigarette Smoke Extract-Induced Autophagy and Dysfunction in Endothelial Progenitor Cells. Oxidative Med. Cell. Longev. 2019, 2019, 7252943. [Google Scholar] [CrossRef]
- Zhang, L.; Gallup, M.; Zlock, L.; Chen, Y.T.F.; Finkbeiner, W.E.; McNamara, N.A. Pivotal Role of MUC1 Glycosylation by Cigarette Smoke in Modulating Disruption of Airway Adherens Junctions in Vitro. J. Pathol. 2014, 234, 60–73. [Google Scholar] [CrossRef]
- Milanic, M.; Hren, R.; Stergar, J.; Simoncic, U. WITHDRAWAL Monitoring of Caffeine Consumption Effect on Skin Blood Properties by Diffuse Reflectance Spectroscopy. Physiol. Res. 2025, 74, 161. [Google Scholar] [CrossRef]
- Aslan, M.; Gürel, E.; Üremis, N.; Üremis, M.; Taslidere, E. Anti-Inflammatory and Antioxidative Effects of Dexpanthenol on Nicotine-Induced Lung Injury in Rats. Toxicol. Environ. Health Sci. 2023, 15, 303–313. [Google Scholar] [CrossRef]
- Nemmar, A.; Beegam, S.; Zaaba, N.; Elzaki, O.; Pathan, A.; Ali, B. Waterpipe Smoke Inhalation Induces Lung Injury and Aortic Endothelial Dysfunction in Mice. Physiol. Res. 2023, 72, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cao, Z.; Wang, L.; Li, J.; Cheng, X.; Tang, Y.; Xing, M.; Yao, P. Chronic High-Fat Diet Induces Galectin-3 and TLR4 to Activate NLRP3 Inflammasome in NASH. J. Nutr. Biochem. 2023, 112, 109217. [Google Scholar] [CrossRef]
- Iacobini, C.; Menini, S.; Ricci, C.; Fantauzzi, C.B.; Scipioni, A.; Salvi, L.; Cordone, S.; Delucchi, F.; Serino, M.; Federici, M.; et al. Galectin-3 Ablation Protects Mice from Diet-Induced NASH: A Major Scavenging Role for Galectin-3 in Liver. J. Hepatol. 2011, 54, 975–983. [Google Scholar] [CrossRef]
- Marín-Royo, G.; Gallardo, I.; Martínez-Martínez, E.; Gutiérrez, B.; Jurado-López, R.; López-Andrés, N.; Gutiérrez-Tenorio, J.; Rial, E.; Bartolomé, M.V.; Nieto, M.L.; et al. Inhibition of Galectin-3 Ameliorates the Consequences of Cardiac Lipotoxicity in a Rat Model of Diet-Induced Obesity. Dis. Models Mech. 2018, 11, dmm032086. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, N.; Silva, A.F.; Rodrigues, P.G.; Correia, E.; Moura, C.; Eloy, C.; Roncon-Albuquerque, R.; Falcão-Pires, I.; Leite-Moreira, A.F. Early Cardiac Changes Induced by a Hypercaloric Western-Type Diet in “Subclinical” Obesity. Am. J. Physiol.-Heart Circ. Physiol. 2016, 310, H655–H666. [Google Scholar] [CrossRef] [PubMed]
- Pejnovic, N.N.; Pantic, J.M.; Jovanovic, I.P.; Radosavljevic, G.D.; Milovanovic, M.Z.; Nikolic, I.G.; Zdravkovic, N.S.; Djukic, A.L.; Arsenijevic, N.N.; Lukic, M.L. Galectin-3 Deficiency Accelerates High-Fat Diet–Induced Obesity and Amplifies Inflammation in Adipose Tissue and Pancreatic Islets. Diabetes 2013, 62, 1932–1944. [Google Scholar] [CrossRef]
- Wensvoort, G. Human C-Peptide Is a Ligand of the Elastin-Receptor-Complex and Therewith Central to Human Vascular Remodelling and Disease in Metabolic Syndrome. Med. Hypotheses 2022, 168, 110964. [Google Scholar] [CrossRef]
- Horne, B.D.; Anderson, J.L.; May, H.T.; Le, V.T.; Galenko, O.; Drakos, S.G.; Bair, T.L.; Knowlton, K.U.; Muhlestein, J.B. Intermittent Fasting and Changes in Galectin-3: A Secondary Analysis of a Randomized Controlled Trial of Disease-Free Subjects. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 1538–1548. [Google Scholar] [CrossRef]
- Lee, J.; An, H.S.; Shin, H.J.; Jang, H.M.; Im, C.O.; Jeong, Y.; Eum, K.; Yoon, S.; Lee, S.J.; Jeong, E.A.; et al. Intermittent Fasting Reduces Neuroinflammation and Cognitive Impairment in High-Fat Diet-Fed Mice by Downregulating Lipocalin-2 and Galectin-3. Nutrients 2024, 16, 159. [Google Scholar] [CrossRef]
- Kim, K.E.; Shin, H.J.; Ju, Y.; Jung, Y.; An, H.S.; Lee, S.J.; Jeong, E.A.; Lee, J.; Hwang, G.-S.; Roh, G.S. Intermittent Fasting Attenuates Metabolic-Dysfunction-Associated Steatohepatitis by Enhancing the Hepatic Autophagy–Lysosome Pathway. Nutrients 2023, 15, 4574. [Google Scholar] [CrossRef]
- Bichara, M.; Attmane-Elakeb, A.; Brown, D.; Essig, M.; Karim, Z.; Muffat-Joly, M.; Micheli, L.; Eude-Le Parco, I.; Cluzeaud, F.; Peuchmaur, M.; et al. Exploring the Role of Galectin 3 in Kidney Function: A Genetic Approach. Glycobiology 2006, 16, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Kröpfl, J.M.; Beltrami, F.G.; Gruber, H.; Schmidt-Trucksäss, A.; Dieterle, T.; Spengler, C.M. Circulating Gal-3 and sST2 Are Associated with Acute Exercise-induced Sustained Endothelial Activation: Possible Relevance for Fibrosis Development? Exp. Physiol. 2023, 108, 1259–1267. [Google Scholar] [CrossRef]
- Kaleta-Duss, A.M.; Lewicka-Potocka, Z.; Dąbrowska-Kugacka, A.; Raczak, G.; Lewicka, E. Myocardial Injury and Overload among Amateur Marathoners as Indicated by Changes in Concentrations of Cardiovascular Biomarkers. Int. J. Environ. Res. Public Health 2020, 17, 6191. [Google Scholar] [CrossRef]
- Vassalle, C.; Masotti, S.; Lubrano, V.; Basta, G.; Prontera, C.; Di Cecco, P.; Del Turco, S.; Sabatino, L.; Pingitore, A. Traditional and New Candidate Cardiac Biomarkers Assessed before, Early, and Late after Half Marathon in Trained Subjects. Eur. J. Appl. Physiol. 2018, 118, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, C.; Kaux, J.-F.; Farre Segura, J.; Stojkovic, V.; Ancion, A.; Seidel, L.; Lancellotti, P.; Cavalier, E. Evolution of the Slopes of ST2 and Galectin-3 during Marathon and Ultratrail Running Compared to a Control Group. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 314–321. [Google Scholar] [CrossRef]
- Lewicka-Potocka, Z.; Kaleta-Duss, A.M.; Lewicka, E.; Kubik, M.; Faran, A.; Szymeczko, P.; Gałąska, R.; Raczak, G.; Dąbrowska-Kugacka, A. Post-Marathon Decline in Right Ventricular Radial Motion Component Among Amateur Sportsmen. Front. Physiol. 2022, 12, 811764. [Google Scholar] [CrossRef]
- Trigiani, L.J.; Lacalle-Aurioles, M.; Bourourou, M.; Li, L.; Greenhalgh, A.D.; Zarruk, J.G.; David, S.; Fehlings, M.G.; Hamel, E. Benefits of Physical Exercise on Cognition and Glial White Matter Pathology in a Mouse Model of Vascular Cognitive Impairment and Dementia. Glia 2020, 68, 1925–1940. [Google Scholar] [CrossRef] [PubMed]
- Trigiani, L.J.; Royea, J.; Tong, X.; Hamel, E. Comparative Benefits of Simvastatin and Exercise in a Mouse Model of Vascular Cognitive Impairment and Dementia. FASEB J. 2019, 33, 13280–13293. [Google Scholar] [CrossRef]
- Avazpour, S.; Amini, A.; Shirvani, H.; Arabzadeh, E. Exercise Modulation in Inflammation and Metabolic Hormonal Disorders of COVID-19 to Decrease Risk Factors in Coronary Heart Disease. Horm. Mol. Biol. Clin. Investig. 2023, 44, 199–206. [Google Scholar] [CrossRef]
- Keyhani, D.; Tartibian, B.; Dabiri, A.; Teixeira, A.M.B. Effect of High-Intensity Interval Training Versus Moderate-Intensity Aerobic Continuous Training on Galectin-3 Gene Expression in Postmenopausal Women: A Randomized Controlled Trial. J. Aging Phys. Act. 2020, 28, 987–995. [Google Scholar] [CrossRef]
- Ahmad, F.; Karim, A.; Khan, J.; Qaisar, R. Plasma Galectin-3 and H-FABP Correlate with Poor Physical Performance in Patients with Congestive Heart Failure. Exp. Biol. Med. (Maywood) 2023, 248, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, V.B.; Nery, G.; Fernandes, L.L.; Santana, T.V.; Jimenez, M.; Borges, L.; Hatanaka, E.; Braga, P.; Monteiro, F.R.; Amaral, J.B.; et al. Salivary Proteome, Inflammatory, and NETosis Biomarkers in Older Adult Practitioners and Nonpractitioners of Physical Exercise. Oxidative Med. Cell. Longev. 2022, 2022, 3725056. [Google Scholar] [CrossRef]
- Salvagno, G.L.; Schena, F.; Gelati, M.; Danese, E.; Cervellin, G.; Guidi, G.C.; Lippi, G. The Concentration of High-Sensitivity Troponin I, Galectin-3 and NT-proBNP Substantially Increase after a 60-Km Ultramarathon. Clin. Chem. Lab. Med. (CCLM) 2014, 52, 267–272. [Google Scholar] [CrossRef]
- Billebeau, G.; Vodovar, N.; Sadoune, M.; Launay, J.-M.; Beauvais, F.; Cohen-Solal, A. Effects of a Cardiac Rehabilitation Programme on Plasma Cardiac Biomarkers in Patients with Chronic Heart Failure. Eur. J. Prev. Cardiol. 2017, 24, 1127–1135. [Google Scholar] [CrossRef]
- Fernandes-Silva, M.M.; Guimarães, G.V.; Rigaud, V.O.; Lofrano-Alves, M.S.; Castro, R.E.; De Barros Cruz, L.G.; Bocchi, E.A.; Bacal, F. Inflammatory Biomarkers and Effect of Exercise on Functional Capacity in Patients with Heart Failure: Insights from a Randomized Clinical Trial. Eur. J. Prev. Cardiol. 2017, 24, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Hättasch, R.; Spethmann, S.; De Boer, R.A.; Ruifrok, W.P.; Schattke, S.; Wagner, M.; Schroeckh, S.; Durmus, T.; Schimke, I.; Sanad, W.; et al. Galectin-3 Increase in Endurance Athletes. Eur. J. Prev. Cardiol. 2014, 21, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhen, J.; Liu, S.; Ren, L.; Zhao, G.; Liang, J.; Xu, A.; Li, C.; Wu, J.; Cheung, B.M.Y. Association between Sleep Patterns and Galectin-3 in a Chinese Community Population. BMC Public. Health 2024, 24, 1323. [Google Scholar] [CrossRef]
- Weigert, J.; Neumeier, M.; Wanninger, J.; Bauer, S.; Farkas, S.; Scherer, M.N.; Schnitzbauer, A.; Schäffler, A.; Aslanidis, C.; Schölmerich, J.; et al. Serum Galectin-3 Is Elevated in Obesity and Negatively Correlates with Glycosylated Hemoglobin in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2010, 95, 1404–1411. [Google Scholar] [CrossRef]
- Suthahar, N.; Meems, L.M.G.; Groothof, D.; Bakker, S.J.L.; Gansevoort, R.T.; Van Veldhuisen, D.J.; De Boer, R.A. Relationship between Body Mass Index, Cardiovascular Biomarkers and Incident Heart Failure. Eur. J. Heart Fail. 2021, 23, 396–402. [Google Scholar] [CrossRef]
- Florido, R.; Kwak, L.; Echouffo-Tcheugui, J.B.; Zhang, S.; Michos, E.D.; Nambi, V.; Goldberg, R.B.; Hoogeveen, R.C.; Lazo, M.; Gerstenblith, G.; et al. Obesity, Galectin-3, and Incident Heart Failure: The ARIC Study. J. Am. Heart Assoc. 2022, 11, e023238. [Google Scholar] [CrossRef]
- Zeytinli Aksit, M.; Demet Arslan, F.; Karakoyun, I.; Aydin, C.; Turgut, E.; Parildar, H.; Gokbalci, U.; Isbilen Basok, B.; Duman, C.; Emiroglu, M. Galectin-3 Levels and Inflammatory Response in Patients Undergoing Bariatric Surgery. Cytokine 2022, 151, 155793. [Google Scholar] [CrossRef]
- Fryk, E.; Rodrigues Silva, V.R.; Strindberg, L.; Strand, R.; Ahlström, H.; Michaëlsson, K.; Kullberg, J.; Lind, L.; Jansson, P.-A. Metabolic Profiling of Galectin-1 and Galectin-3: A Cross-Sectional, Multi-Omics, Association Study. Int. J. Obes. 2024, 48, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.; Wang, A.; Xu, T.; Zhong, C.; Zheng, X.; Zhu, Z.; Peng, Y.; Peng, H.; Li, Q.; Ju, Z.; et al. Co-Effect of Serum Galectin-3 and High-Density Lipoprotein Cholesterol on the Prognosis of Acute Ischemic Stroke. J. Stroke Cerebrovasc. Dis. 2019, 28, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Tan, J.; Hong, Y. Effect of Inhibition of Galectin-3 Expression on Lipid Accumulation and Atherosclerosis Lesions in a Rat Model of Carotid Atherosclerosis. J. Biol. Regul. Homeost. Agents 2024, 38, 3215–3223. [Google Scholar] [CrossRef]
- Winter, M.; Wiesbauer, F.; Alimohammadi, A.; Blessberger, H.; Pavo, N.; Schillinger, M.; Huber, K.; Wojta, J.; Lang, I.M.; Maurer, G.; et al. Soluble Galectin-3 Is Associated with Premature Myocardial Infarction. Eur. J. Clin. Investig. 2016, 46, 386–391. [Google Scholar] [CrossRef]
- Melin, E.O.; Dereke, J.; Hillman, M. Female Sex, High Soluble CD163, and Low HDL-Cholesterol Were Associated with High Galectin-3 Binding Protein in Type 1 Diabetes. Biol. Sex. Differ. 2019, 10, 51. [Google Scholar] [CrossRef]
- Nangia-Makker, P.; Honjo, Y.; Sarvis, R.; Akahani, S.; Hogan, V.; Pienta, K.J.; Raz, A. Galectin-3 Induces Endothelial Cell Morphogenesis and Angiogenesis. Am. J. Pathol. 2000, 156, 899–909. [Google Scholar] [CrossRef]
- Pang, Z.-D.; Sun, X.; Bai, R.-Y.; Han, M.-Z.; Zhang, Y.-J.; Wu, W.; Zhang, Y.; Lai, B.-C.; Zhang, Y.; Wang, Y.; et al. YAP-Galectin-3 Signaling Mediates Endothelial Dysfunction in Angiotensin II-Induced Hypertension in Mice. Cell. Mol. Life Sci. 2023, 80, 38. [Google Scholar] [CrossRef]
- Hao, W.-R.; Cheng, C.-H.; Liu, J.-C.; Chen, H.-Y.; Chen, J.-J.; Cheng, T.-H. Understanding Galectin-3’s Role in Diastolic Dysfunction: A Contemporary Perspective. Life 2024, 14, 906. [Google Scholar] [CrossRef]
- Hsu, B.-G.; Wang, C.-H.; Lai, Y.-H.; Tsai, J.-P. Serum Galectin-3 Level Is Positively Associated with Endothelial Dysfunction in Patients with Chronic Kidney Disease Stage 3 to 5. Toxins 2021, 13, 532. [Google Scholar] [CrossRef]
- Yao, Y.; Shen, D.; Chen, R.; Ying, C.; Wang, C.; Guo, J.; Zhang, G. Galectin-3 Predicts Left Ventricular Remodeling of Hypertension. J. Clin. Hypertens. 2016, 18, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Blanda, V.; Bracale, U.M.; Di Taranto, M.D.; Fortunato, G. Galectin-3 in Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 9232. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, V.L. Galectins in Endothelial Cell Biology and Angiogenesis: The Basics. Biomolecules 2021, 11, 1386. [Google Scholar] [CrossRef]
- Bielecka-Dabrowa, A.; Gluba-Brzózka, A.; Michalska-Kasiczak, M.; Misztal, M.; Rysz, J.; Banach, M. The Multi-Biomarker Approach for Heart Failure in Patients with Hypertension. Int. J. Mol. Sci. 2015, 16, 10715–10733. [Google Scholar] [CrossRef] [PubMed]
- Ianoș, R.D.; Cozma, A.; Lucaciu, R.L.; Hangan, A.C.; Negrean, V.; Mercea, D.C.; Ciulei, G.; Pop, C.; Procopciuc, L.M. Role of Circulating Biomarkers in Diabetic Cardiomyopathy. Biomedicines 2024, 12, 2153. [Google Scholar] [CrossRef]
- Lebedev, D.A.; Lyasnikova, E.A.; Vasilyeva, E.Y.; Likhonosov, N.P.; Sitnikova, M.Y.; Babenko, A.Y. Association between Markers of Fibrosis and Heart Failure Incidence in Patients with Type 2 Diabetes Mellitus. J. Diabetes Res. 2021, 2021, 9589185. [Google Scholar] [CrossRef]
- Flores-Ramírez, R.; Azpiri-López, J.R.; González-González, J.G.; Ordaz-Farías, A.; González-Carrillo, L.E.; Carrizales-Sepúlveda, E.F.; Vera-Pineda, R. Global Longitudinal Strain as a Biomarker in Diabetic Cardiomyopathy. A Comparative Study with Gal-3 in Patients with Preserved Ejection Fraction. Arch. Cardiol. México 2017, 87, 278–285. [Google Scholar] [CrossRef]
- Jin, Q.; Lou, Y.; Li, T.; Chen, H.; Liu, Q.; He, X. Serum Galectin-3: A Risk Factor for Vascular Complications in Type 2 Diabetes Mellitus. Chin. Med. J. 2013, 126, 2109–2115. [Google Scholar] [CrossRef]
- Tan, K.C.B.; Cheung, C.; Lee, A.C.H.; Lam, J.K.Y.; Wong, Y.; Shiu, S.W.M. Galectin-3 and Risk of Cardiovascular Events and All-cause Mortality in Type 2 Diabetes. Diabetes Metab. Res. 2019, 35, e3093. [Google Scholar] [CrossRef]
- Kuzan, A.; Królewicz, E.; Kustrzeba-Wójcicka, I.; Lindner-Pawłowicz, K.; Sobieszczańska, M. How Diabetes and Other Comorbidities of Elderly Patients and Their Treatment Influence Levels of Glycation Products. Int. J. Environ. Res. Public Health 2022, 19, 7524. [Google Scholar] [CrossRef]
- Vora, A.; De Lemos, J.A.; Ayers, C.; Grodin, J.L.; Lingvay, I. Association of Galectin-3 With Diabetes Mellitus in the Dallas Heart Study. J. Clin. Endocrinol. Metab. 2019, 104, 4449–4458. [Google Scholar] [CrossRef]
- Yilmaz, H.; Cakmak, M.; Inan, O.; Darcin, T.; Akcay, A. Increased Levels of Galectin-3 Were Associated with Prediabetes and Diabetes: New Risk Factor? J. Endocrinol. Investig. 2015, 38, 527–533. [Google Scholar] [CrossRef]
- Blasetti Fantauzzi, C.; Iacobini, C.; Menini, S.; Vitale, M.; Sorice, G.P.; Mezza, T.; Cinti, S.; Giaccari, A.; Pugliese, G. Galectin-3 Gene Deletion Results in Defective Adipose Tissue Maturation and Impaired Insulin Sensitivity and Glucose Homeostasis. Sci. Rep. 2020, 10, 20070. [Google Scholar] [CrossRef] [PubMed]
- Lindman, B.R.; Dávila-Román, V.G.; Mann, D.L.; McNulty, S.; Semigran, M.J.; Lewis, G.D.; De Las Fuentes, L.; Joseph, S.M.; Vader, J.; Hernandez, A.F.; et al. Cardiovascular Phenotype in HFpEF Patients With or Without Diabetes. J. Am. Coll. Cardiol. 2014, 64, 541–549. [Google Scholar] [CrossRef]
- Andersen, M.L.; Moyses-Oliveira, M.; Tufik, S. Unlocking the Role of Galectin-3: Implications for Sleep Disorders and Health. Sleep Med. 2024, 124, 110–114. [Google Scholar] [CrossRef]
- Pusuroglu, H.; Somuncu, U.; Bolat, I.; Akgul, O.; Ornek, V.; Yıldırım, H.A.; Akkaya, E.; Karakurt, H.; Yıldırım, A.; Savaş, A.U. Galectin-3 Is Associated with Coronary Plaque Burden and Obstructive Sleep Apnoea Syndrome Severity. Kardiol. Pol. 2017, 75, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Hanis, C.L.; Redline, S.; Ballantyne, C.M.; Hamzeh, I.; Aguilar, D. Sleep Apnea and Galectin-3: Possible Sex-Specific Relationship. Sleep Breath. 2019, 23, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Khalaji, A.; Amirkhani, N.; Sharifkashani, S.; Behnoush, A.H. Role of Galectin-3 as a Biomarker in Obstructive Sleep Apnea: A Systematic Review and Meta-Analysis. Sleep Breath. 2023, 27, 2273–2282. [Google Scholar] [CrossRef]
- Zong, D.; Liu, X.; Shen, C.; Liu, T.; Ouyang, R. Involvement of Galectin-3 in Neurocognitive Impairment in Obstructive Sleep Apnea via Regulating Inflammation and Oxidative Stress through NLRP3. Sleep Med. 2023, 101, 1–10. [Google Scholar] [CrossRef]
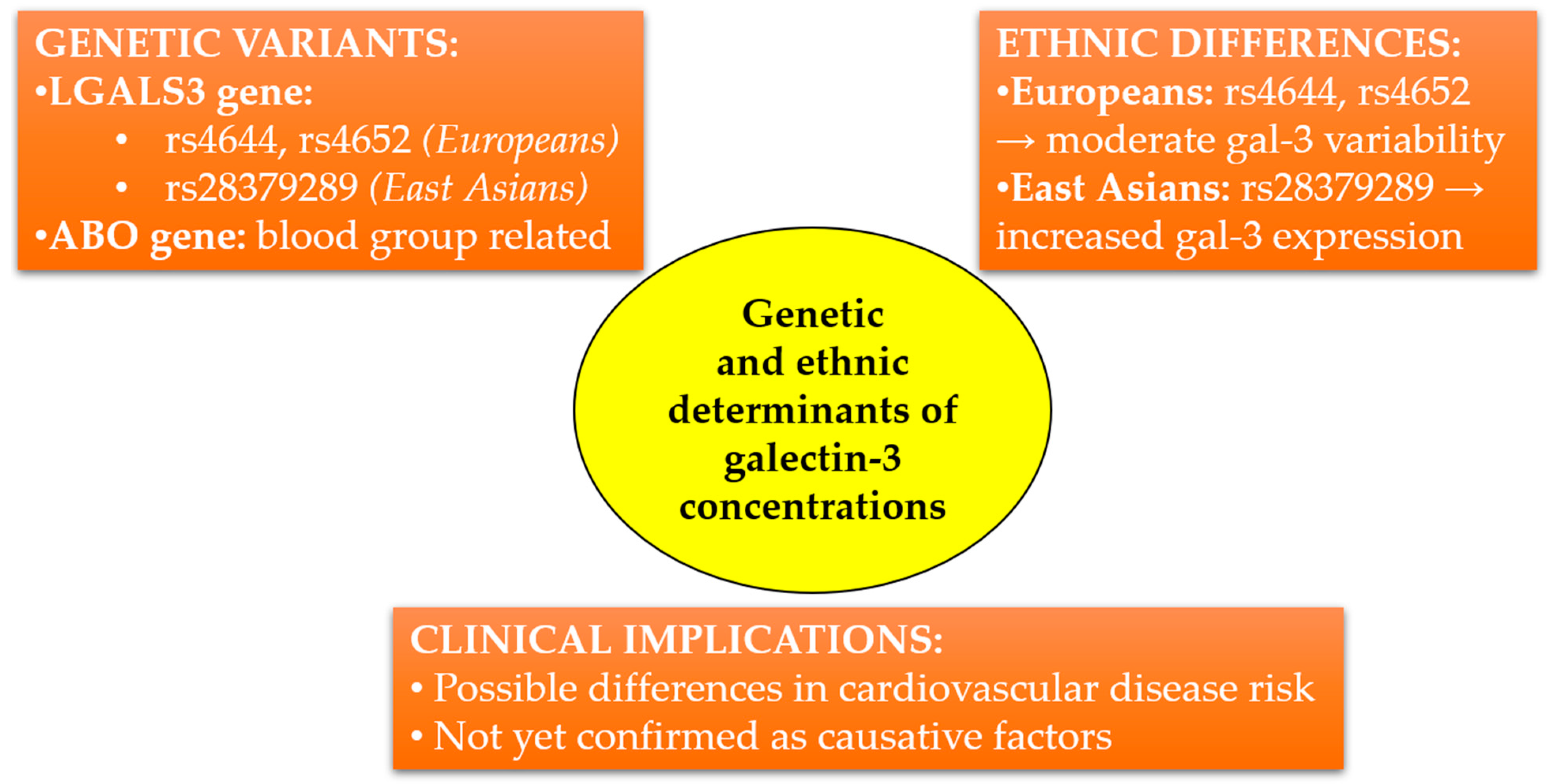
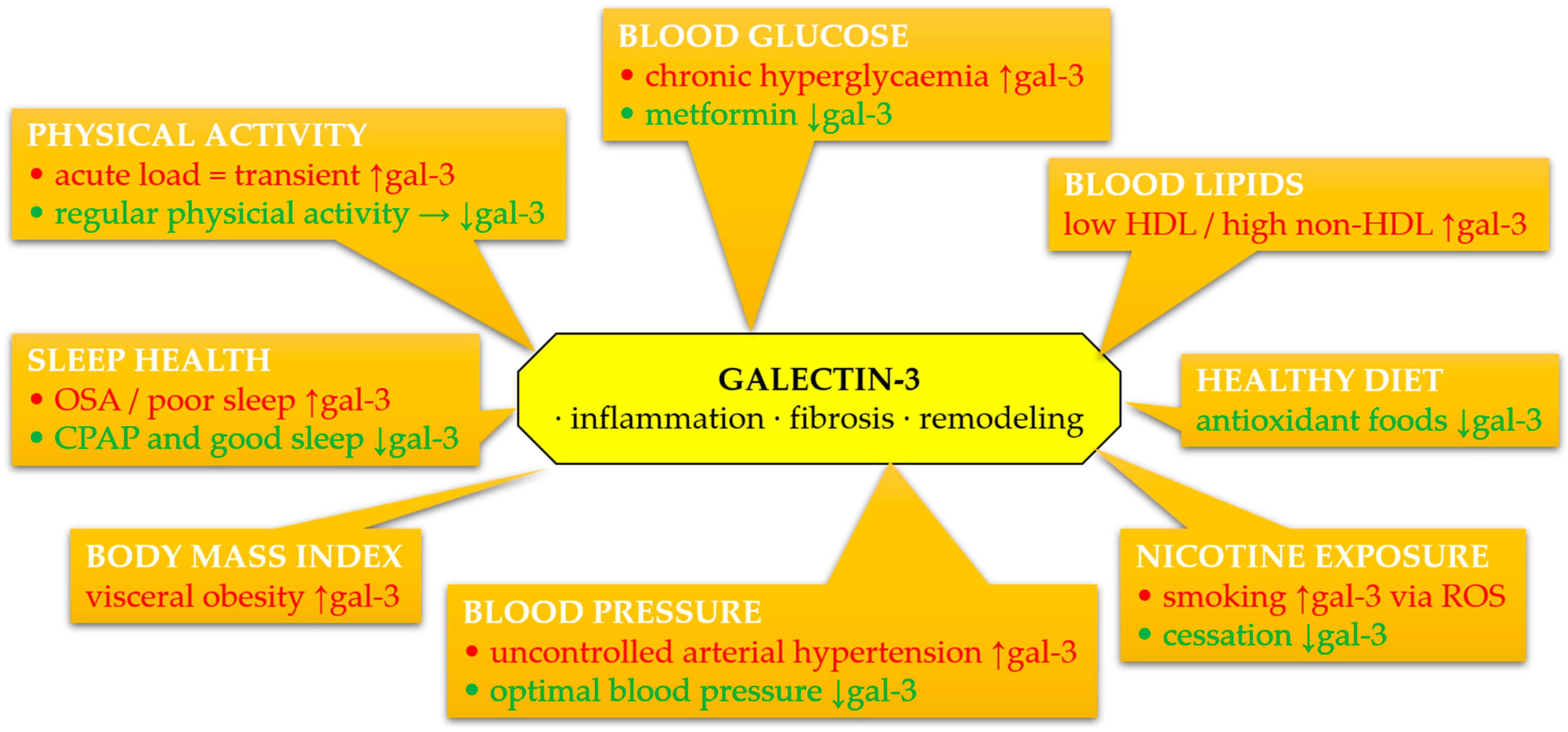
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).