
Marine Bioactive Peptides: Anti-Photoaging Mechanisms and Potential Skin Protective Effects

Abstract

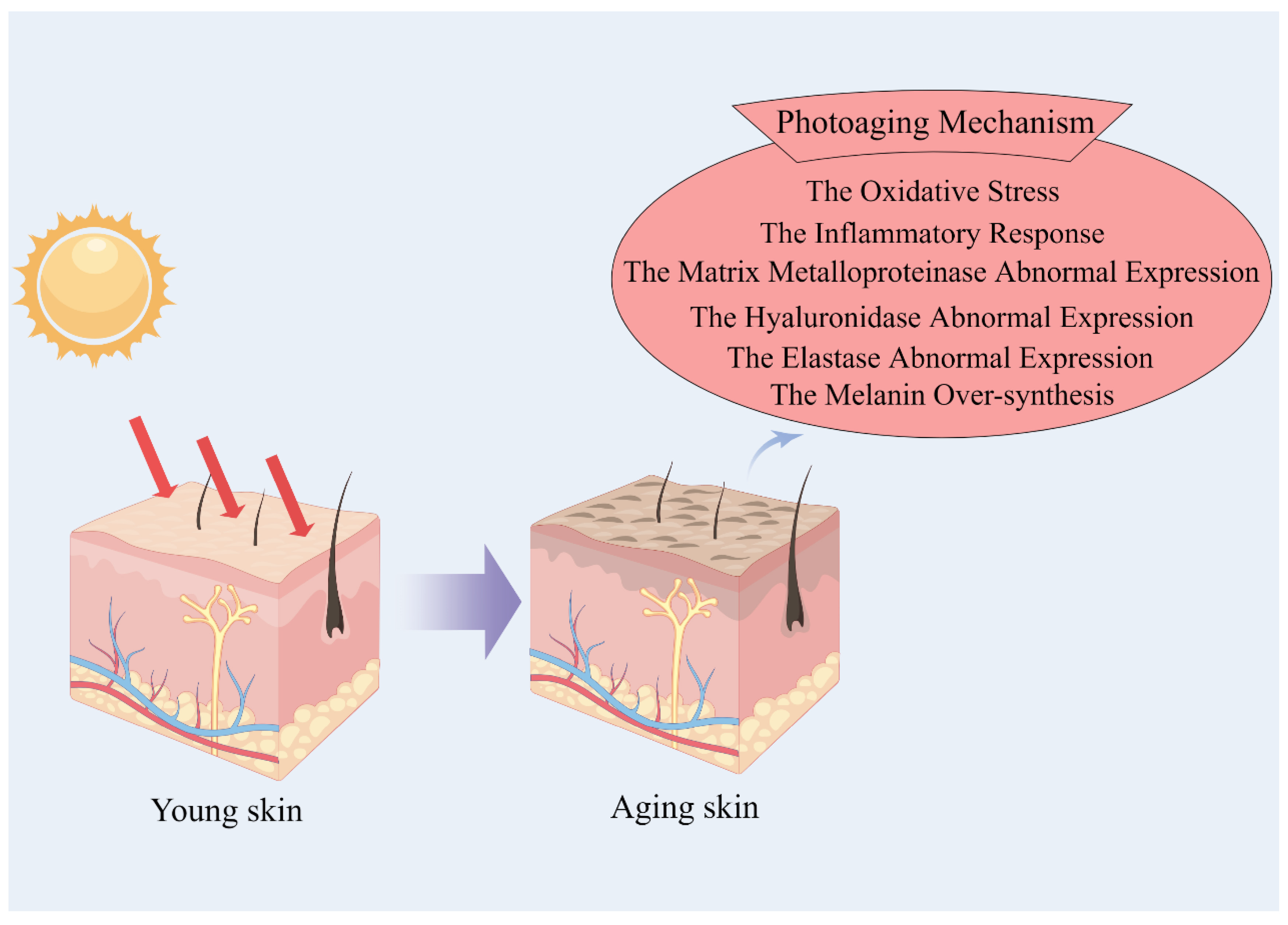
1. Introduction
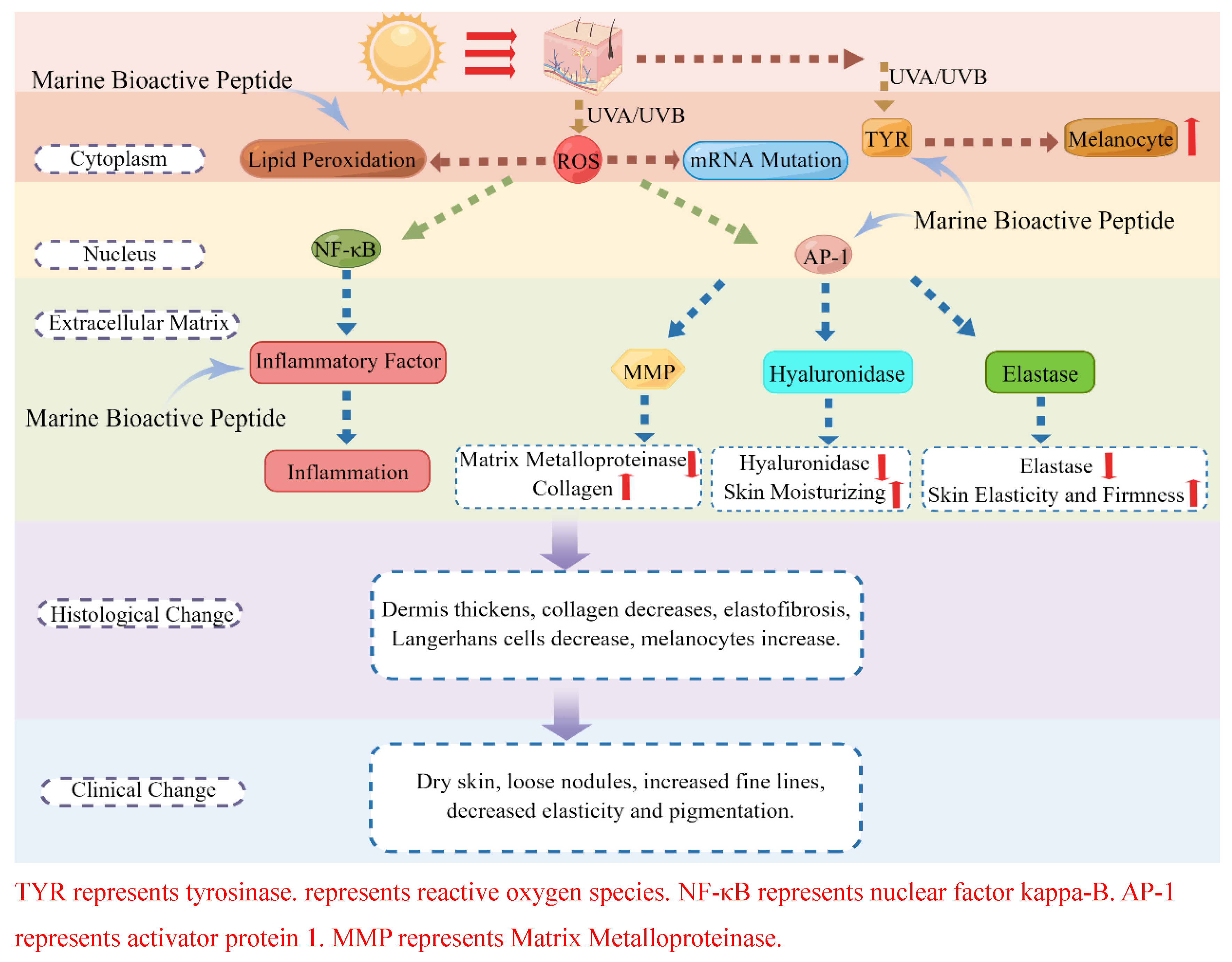
2. Anti-Photoaging Mechanism of Marine Bioactive Peptides
2.1. Peptide Anti-Skin-Photoaging by Inhibiting Oxidative Stress Damage
2.2. Peptide Anti-Skin-Photoaging via Anti-Inflammation
2.3. Peptide Anti-Skin-Photoaging via Inhibition of Matrix Metalloproteinases
2.4. Peptide Anti-Skin-Photoaging via Inhibition of Hyaluronidase
2.5. Peptide Anti-Skin-Photoaging via Inhibition of Elastase
2.6. Peptide Anti-Skin-Photoaging via Inhibition of Melanin Over-Synthesis
3. Skin Protective Effects of Marine Bioactive Peptides
3.1. Peptides Improve Skin via Photoprotective Mechanisms
3.2. Peptides Improve Skin via Anti-Microbial Mechanisms
3.3. Peptides Improve Skin via Skin Repair
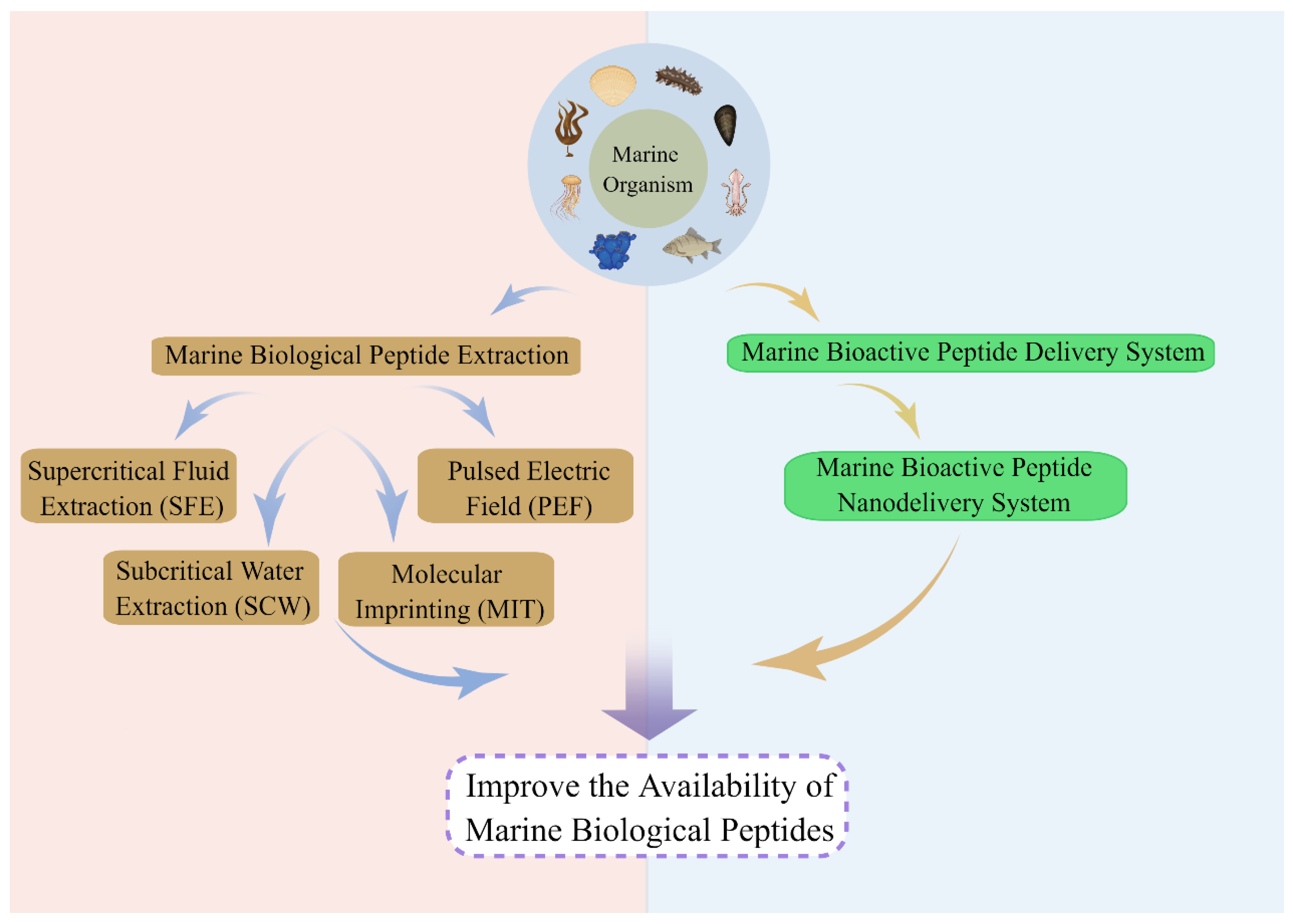
4. Bioavailability of Marine Bioactive Peptides
4.1. Improvement of Bioavailability of Marine Bioactive Peptides via Isolation and Purification
4.2. Improvement of Bioavailability of Marine Bioactive Peptides via Nanodelivery Systems
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vodouhe, M.; Marois, J.; Guay, V.; Leblanc, N.; Weisnagel, S.J.; Bilodeau, J.-F.; Jacques, H. Marginal Impact of Brown Seaweed Ascophyllum nodosum and Fucus vesiculosus Extract on Metabolic and Inflammatory Response in Overweight and Obese Prediabetic Subjects. Mar. Drugs 2022, 20, 174. [Google Scholar] [CrossRef]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, J.; Anil, S.; Kim, S.-K.; Shim, M.S. Marine Fish Proteins and Peptides for Cosmeceuticals: A Review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef]
- Smichi, N.; Parsiegla, G.; Achouri, N.; Zarai, Z.; Abousalham, A.; Fendri, A. Intestinal phospholipase A2 from Sparidae species: Functional properties and cytotoxic potential evaluation. Int. J. Biol. Macromol. 2020, 143, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, Q.; Zhang, B.; Zhao, Y.; Wang, N. Potential Active Marine Peptides as Anti-Aging Drugs or Drug Candidates. Mar. Drugs 2023, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Lv, S.; Ma, Y.; Liu, N.; Zhou, S.; Zhou, D. In vitro antioxidant and anti-aging properties of swim bladder peptides from Atlantic cod (Gadus morhua). Int. J. Food Prop. 2020, 23, 1416–1429. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, X.; Lin, L.; Ding, G.; Yu, F. Immunomodulatory Activity of Low Molecular-Weight Peptides from Nibea japonica in RAW264.7 Cells via NF-kappa B Pathway. Mar. Drugs 2019, 17, 404. [Google Scholar] [CrossRef]
- Song, R.; Wei, R.-B.; Luo, H.-Y.; Wang, D.-F. Isolation and Characterization of an Antibacterial Peptide Fraction from the Pepsin Hydrolysate of Half-Fin Anchovy (Setipinna taty). Molecules 2012, 17, 2980–2991. [Google Scholar] [CrossRef]
- Jensen, I.-J.; Maehre, H.K. Preclinical and Clinical Studies on Antioxidative, Antihypertensive and Cardioprotective Effect of Marine Proteins and Peptides A Review. Mar. Drugs 2016, 14, 211. [Google Scholar] [CrossRef]
- Frau, J.; Flores-Holguin, N.; Glossman-Mitnik, D. Chemical Reactivity Theory and Empirical Bioactivity Scores as Computational Peptidology Alternative Tools for the Study of Two Anticancer Peptides of Marine Origin. Molecules 2019, 24, 1115. [Google Scholar] [CrossRef]
- Khazaei Monfared, Y.; Mahmoudian, M.; Cecone, C.; Caldera, F.; Zakeri-Milani, P.; Matencio, A.; Trotta, F. Stabilization and Anticancer Enhancing Activity of the Peptide Nisin by Cyclodextrin-Based Nanosponges against Colon and Breast Cancer Cells. Polymers 2022, 14, 594. [Google Scholar] [CrossRef]
- Sridhar, K.; Inbaraj, B.S.; Chen, B.-H. Recent developments on production, purification and biological activity of marine peptides. Food Res. Int. 2021, 147, 468. [Google Scholar] [CrossRef]
- Elsheikh, M.A.; Gaafar, P.M.E.; Khattab, M.A.; Helwah, M.K.A.; Noureldin, M.H.; Abbas, H. Dual-effects of caffeinated hyalurosomes as a nano-cosmeceutical gel counteracting UV-induced skin ageing. Int. J. Pharm.-X 2023, 5, 100170. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Zheng, S.; Fang, M.; Kim, M.; Bellere, A.D.; Jeong, J.; Yi, T.-H. Anti-Photoaging Effect of Phaseolus angularis L. Extract on UVB-Exposed HaCaT Keratinocytes and Possibilities as Cosmetic Materials. Molecules 2023, 28, 1407. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Geng, R.; Kang, S.-G.; Huang, K.; Tong, T. Boosting the Photoaged Skin: The Potential Role of Dietary Components. Nutrients 2021, 13, 1691. [Google Scholar] [CrossRef]
- Honeybrook, A.; Bernstein, E. Oral isotretinoin and photoaging: A review. J. Cosmet. Dermatol. 2020, 19, 1548–1554. [Google Scholar] [CrossRef]
- Guan, B.; Wang, F.; Jiang, H.; Zhou, M.; Lin, H. Preparation of Mesoporous Silica Nanosphere-Doped Color-Sensitive Materials and Application in Monitoring the TVB-N of Oysters. Foods 2022, 11, 817. [Google Scholar] [CrossRef]
- Wang, Y.; Glukhov, E.; He, Y.; Liu, Y.; Zhou, L.; Ma, X.; Hu, X.; Hong, P.; Gerwick, W.H.; Zhang, Y. Secondary Metabolite Variation and Bioactivities of Two Marine Aspergillus Strains in Static Co-Culture Investigated by Molecular Network Analysis and Multiple Database Mining Based on LC-PDA-MS/MS. Antibiotics 2022, 11, 513. [Google Scholar] [CrossRef]
- Pallela, R.; Na-Young, Y.; Kim, S.-K. Anti-photoaging and Photoprotective Compounds Derived from Marine Organisms. Mar. Drugs 2010, 8, 1189–1202. [Google Scholar] [CrossRef]
- Han, J.; Huang, Z.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Li, Y.; Ming, T.; Wang, Z.J.; Su, X. The novel peptides ICRD and LCGEC screened from tuna roe show antioxidative activity via Keap1/Nrf2-ARE pathway regulation and gut microbiota modulation. Food Chem. 2020, 327, 94. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, P.; Xiao, Z.; Chen, M.-F.; Gong, F.; Li, C.; Zhou, C.; Hong, P.; Jung, W.-K.; Qian, Z.-J. Antiphotoaging effect of boiled abalone residual peptide ATPGDEG on UVB-induced keratinocyte HaCaT cells. Food Nutr. Res. 2019, 63, 3508. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Lecci, R.M.; Durante, M.; Meli, F.; Piraino, S. The Bright Side of Gelatinous Blooms: Nutraceutical Value and Antioxidant Properties of Three Mediterranean Jellyfish (Scyphozoa). Mar. Drugs 2015, 13, 4654–4681. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of Collagen and Collagen Hydrolysate from Jellyfish (Rhopilema esculentum) on Mice Skin Photoaging Induced by UV Irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H.; Lu, J.; Zhang, K.; Li, B. Protective effect of gelatin and gelatin hydrolysate from salmon skin on UV irradiation-induced photoaging of mice skin. J. Ocean Univ. China 2016, 15, 711–718. [Google Scholar] [CrossRef]
- Kong, J.; Hu, X.-M.; Cai, W.-W.; Wang, Y.-M.; Chi, C.-F.; Wang, B. Bioactive Peptides from Skipjack Tuna Cardiac Arterial Bulbs (II): Protective Function on UVB-Irradiated HaCaT Cells through Antioxidant and Anti-Apoptotic Mechanisms. Mar. Drugs 2023, 21, 105. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Y.; Zhuang, Y. Antiphotoaging effect and purification of an antioxidant peptide from tilapia (Oreochromis niloticus) gelatin peptides. J. Funct. Foods 2013, 5, 154–162. [Google Scholar] [CrossRef]
- Cai, W.-W.; Hu, X.-M.; Wang, Y.-M.; Chi, C.-F.; Wang, B. Bioactive Peptides from Skipjack Tuna Cardiac Arterial Bulbs: Preparation, Identification, Antioxidant Activity, and Stability against Thermal, pH, and Simulated Gastrointestinal Digestion Treatments. Mar. Drugs 2022, 20, 626. [Google Scholar] [CrossRef]
- Hu, X.-M.; Wang, Y.-M.; Zhao, Y.-Q.; Chi, C.-F.; Wang, B. Antioxidant Peptides from the Protein Hydrolysate of Monkfish (Lophius litulon) Muscle: Purification, Identification, and Cytoprotective Function on HepG2 Cells Damage by H2O2. Mar. Drugs 2020, 18, 153. [Google Scholar] [CrossRef]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Fractionation and identification of antioxidant peptides from an enzymatically hydrolysed Palmaria palmata protein isolate. Food Res. Int. 2017, 100, 416–422. [Google Scholar] [CrossRef]
- Je, J.-Y.; Qian, Z.-J.; Lee, S.-H.; Byun, H.-G.; Kim, S.-K. Purification and Antioxidant Properties of Bigeye Tuna (Thunnus obesus) Dark Muscle Peptide on Free Radical-Mediated Oxidative Systems. J. Med. Food 2008, 11, 629–637. [Google Scholar] [CrossRef]
- Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2012, 42, 1641–1649. [Google Scholar] [CrossRef]
- Lee, W.-S.; Jeon, J.-K.; Byun, H.-G. Characterization of a novel antioxidative peptide from the sand eel Hypoptychus dybowskii. Process Biochem. 2011, 46, 1207–1211. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Qian, Z.-J.; Ryu, B.; Park, J.W.; Kim, S.-K. In vitro antioxidant activity of a peptide isolated from Nile tilapia (Oreochromis niloticus) scale gelatin in free radical-mediated oxidative systems. J. Funct. Foods 2010, 2, 107–117. [Google Scholar] [CrossRef]
- Jiang, H.; Tong, T.; Sun, J.; Xu, Y.; Zhao, Z.; Liao, D. Purification and characterization of antioxidative peptides from round scad (Decapterus maruadsi) muscle protein hydrolysate. Food Chem. 2014, 154, 158–163. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Kim, S.K. Antioxidant properties of a radical-scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate. J. Agric. Food Chem. 2005, 53, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-Y.; Lee, J.-H.; Samarakoon, K.; Kim, J.-S.; Jeon, Y.-J. Purification and determination of two novel antioxidant peptides from flounder fish (Paralichthys olivaceus) using digestive proteases. Food Chem. Toxicol. 2013, 52, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H. Protective effect of gelatin polypeptides from Pacific cod (Gadus macrocephalus) against UV irradiation-induced damages by inhibiting inflammation and improving transforming growth factor-beta/Smad signaling pathway. J. Photochem. Photobiol. B-Biol. 2016, 162, 633–640. [Google Scholar] [CrossRef]
- Chen, B.; Yu, L.; Wu, J.; Qiao, K.; Cui, L.; Qu, H.; Su, Y.; Cai, S.; Liu, Z.; Wang, Q. Effects of Collagen Hydrolysate From Large Hybrid Sturgeon on Mitigating Ultraviolet B-Induced Photodamage. Front. Bioeng. Biotechnol. 2022, 10, 908033. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Lai, R.; Deng, H.; Zhou, F.; Wang, P.; Pang, X.; Huang, G.; Chen, X.; Lin, H.; et al. Protective Effect of the Pearl Extract from Pinctada fucata martensii Dunker on UV-Induced Photoaging in Mice. Chem. Biodivers. 2022, 19, 876. [Google Scholar] [CrossRef]
- Lu, J.; Hou, H.; Fan, Y.; Yang, T.; Li, B. Identification of MMP-1 inhibitory peptides from cod skin gelatin hydrolysates and the inhibition mechanism by MAPK signaling pathway. J. Funct. Foods 2017, 33, 251–260. [Google Scholar] [CrossRef]
- Wei, M.; Qiu, H.; Zhou, J.; Yang, C.; Chen, Y.; You, L. The Anti-Photoaging Activity of Peptides from Pinctada martensii Meat. Mar. Drugs 2022, 20, 770. [Google Scholar] [CrossRef]
- Kim, C.-R.; Kim, Y.-M.; Lee, M.-K.; Kim, I.-H.; Choi, Y.-H.; Nam, T.-J. Pyropia yezoensis peptide promotes collagen synthesis by activating the TGF-beta/Smad signaling pathway in the human dermal fibroblast cell line Hs27. Int. J. Mol. Med. 2017, 39, 31–38. [Google Scholar] [CrossRef]
- Zhang, C.; Lv, J.; Qin, X.; Peng, Z.; Lin, H. Novel Antioxidant Peptides from Crassostrea Hongkongensis Improve Photo-Oxidation in UV-Induced HaCaT Cells. Mar. Drugs 2022, 20, 100. [Google Scholar] [CrossRef]
- Xiao, Z.; Liang, P.; Chen, J.; Chen, M.-F.; Gong, F.; Li, C.; Zhou, C.; Hong, P.; Yang, P.; Qian, Z.-J. A Peptide YGDEY from Tilapia Gelatin Hydrolysates Inhibits UVB-mediated Skin Photoaging by Regulating MMP-1 and MMP-9 Expression in HaCaT Cells. Photochem. Photobiol. 2019, 95, 1424–1432. [Google Scholar] [CrossRef] [PubMed]
- Juncan, A.M.; Moisa, D.G.; Santini, A.; Morgovan, C.; Rus, L.-L.; Vonica-Tincu, A.L.; Loghin, F. Advantages of Hyaluronic Acid and Its Combination with Other Bioactive Ingredients in Cosmeceuticals. Molecules 2021, 26, 4429. [Google Scholar] [CrossRef] [PubMed]
- Norzagaray-Valenzuela, C.D.; Valdez-Ortiz, A.; Shelton, L.M.; Jimenez-Edeza, M.; Rivera-Lopez, J.; Valdez-Flores, M.A.; German-Baez, L.J. Residual biomasses and protein hydrolysates of three green microalgae species exhibit antioxidant and anti-aging activity. J. Appl. Phycol. 2017, 29, 189–198. [Google Scholar] [CrossRef]
- Barriga Montalvo, G.E.; Thomaz-Soccol, V.; Vandenberghe, L.P.S.; Carvalho, J.C.; Faulds, C.B.; Bertrand, E.; Prado, M.R.M.; Bonatto, S.J.R.; Soccol, C.R. Arthrospira maxima OF15 biomass cultivation at laboratory and pilot scale from sugarcane vinasse for potential biological new peptides production. Bioresour. Technol. 2019, 273, 103–113. [Google Scholar] [CrossRef]
- Nakchum, L.; Kim, S.M. Preparation of squid skin collagen hydrolysate as an antihyaluronidase, antityrosinase, and antioxidant agent. Prep. Biochem. Biotechnol. 2016, 46, 123–130. [Google Scholar] [CrossRef]
- Hoon, K.; Jeon, B.; Lee, H.-J.; Kyun, C.D. Evaluation of the Skin Moisturizing Efficacy of a Collagen Peptide Isolated from Fish Scales, Using HaCaT Keratinocytes. J. Korean Soc. Food Sci. Nutr. 2020, 49, 454–461. [Google Scholar] [CrossRef]
- Nam, K.A.; You, S.G.; Kim, S.M. Molecular and physical characteristics of squid (Todarodes pacificus) skin collagens and biological properties of their enzymatic hydrolysates. J. Food Sci. 2008, 73, C249–C255. [Google Scholar] [CrossRef]
- Amakye, W.K.; Yang, L.; Yao, M.; Yuan, E.; Ren, R.; Ren, J. Skipjack (Katsuwonus pelamis) elastin hydrolysate-derived peptides attenuate UVA irradiation-induced cell damage in human HaCaT keratinocytes. Food Front. 2021, 2, 184–194. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Liang, C.-H.; Wu, H.-T.; Pang, H.-Y.; Chen, C.; Wang, G.-H.; Chan, L.-P. Antioxidant and anti-inflammatory capacities of collagen peptides from milkfish (Chanos chanos) scales. J. Food Sci. Technol. Mysore 2018, 55, 2310–2317. [Google Scholar] [CrossRef]
- Mechri, S.; Sellem, I.; Bouacem, K.; Jabeur, F.; Laribi-Habchi, H.; Mellouli, L.; Hacene, H.; Bouanane-Darenfed, A.; Jaouadi, B. A biological clean processing approach for the valorization of speckled shrimp Metapenaeus monoceros by-product as a source of bioactive compounds. Environ. Sci. Pollut. Res. 2020, 27, 15842–15855. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.; Cheng, S.; Li, H.; Xu, X.; Wang, Z.; Du, M. Tyrosinase inhibitory effects of the peptides from fish scale with the metal copper ions chelating ability. Food Chem. 2022, 390, 133146. [Google Scholar] [CrossRef] [PubMed]
- Mechri, S.; Sellem, I.; Bouacem, K.; Jabeur, F.; Chamkha, M.; Hacene, H.; Bouanane-Darenfed, A.; Jaouadi, B. Antioxidant and Enzyme Inhibitory Activities of Metapenaeusmonoceros By-Product Hydrolysates Elaborated by Purified Alkaline Proteases. Waste Biomass Valorization 2020, 11, 6741–6755. [Google Scholar] [CrossRef]
- Hu, Z.-Z.; Sha, X.-M.; Zhang, L.; Zha, M.-J.; Tu, Z.-C. From Fish Scale Gelatin to Tyrosinase Inhibitor: A Novel Peptides Screening Approach Application. Front. Nutr. 2022, 9, 853442. [Google Scholar] [CrossRef]
- Wang, J.; Xie, X.; Deng, Y.; Yang, H.; Du, X.; Liu, P.; Du, Y. SOX9 in Keratinocytes Regulates Claudin 2 Transcription during Skin Aging. Contrast Media Mol. Imaging 2022, 2022, 6884308. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Zhang, X.-L. DNA Methyltransferase Inhibitor 5-AZA-DC Regulates TGF beta 1-Mediated Alteration of Neuroglial Cell Functions after Oxidative Stress. Oxid. Med. Cell. Longev. 2022, 2022, 9259465. [Google Scholar] [CrossRef]
- Chu, N.; Yao, G.; Liu, Y.; Cheng, M.; Ikejima, T. Newly synthesized bis-benzimidazole compound 8 induces apoptosis, autophagy and reactive oxygen species generation in HeLa cells. Bioorg. Med. Chem. Lett. 2016, 26, 227–231. [Google Scholar] [CrossRef]
- Jesus, A.; Mota, S.; Torres, A.; Cruz, M.T.; Sousa, E.; Almeida, I.F.; Cidade, H. Antioxidants in Sunscreens: Which and What For? Antioxidants 2023, 12, 138. [Google Scholar] [CrossRef]
- Khan, M.A.; Siddiqui, S.; Ahmad, I.; Singh, R.; Mishra, D.P.; Srivastava, A.N.; Ahmad, R. Phytochemicals from Ajwa dates pulp extract induce apoptosis in human triple-negative breast cancer by inhibiting AKT/mTOR pathway and modulating Bcl-2 family proteins. Sci. Rep. 2021, 11, 10322. [Google Scholar] [CrossRef]
- Zhao, R.; Jiang, X.-X.; Zhao, Q.-L.; Ye, H.-W.; Lin, Y.; Huang, J.; Tang, Y.-P. Immunoenhancing Effects of Cyclina sinensis Pentadecapeptide through Modulation of Signaling Pathways in Mice with Cyclophosphamide-Induced Immunosuppression. Mar. Drugs 2022, 20, 560. [Google Scholar] [CrossRef]
- Lembo, S.; Balato, A.; Di Caprio, R.; Cirillo, T.; Giannini, V.; Gasparri, F.; Monfrecola, G. The Modulatory Effect of Ellagic Acid and Rosmarinic Acid on Ultraviolet-B-Induced Cytokine/Chemokine Gene Expression in Skin Keratinocyte (HaCaT) Cells. Biomed Res. Int. 2014, 2014, 346793. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, X.; Zhang, Q.; Wang, Y.; Wang, S.; Li, Y.; Zhang, Y.; Jing, J.; Qiu, J.; Wang, Z.; et al. Comparative transcriptome profiles of Lindian chicken eyelids identify melanin genes controlling eyelid pigmentation. Br. Poult. Sci. 2019, 60, 15–22. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Wang, S.; Liu, H.; Zhang, D.; Wang, Y.; Ji, H. Swine-Derived Probiotic Lactobacillus plantarum Modulates Porcine Intestinal Endogenous Host Defense Peptide Synthesis Through TLR2/MAPK/AP-1 Signaling Pathway. Front. Immunol. 2019, 10, 02691. [Google Scholar] [CrossRef]
- Chiu, L.-Y.; Wu, N.-L.; Hung, C.-F.; Bai, P.; Dai, Y.-S.; Lin, W.-W. PARP-1 involves in UVB-induced inflammatory response in keratinocytes and skin injury via regulation of ROS-dependent EGFR transactivation and p38 signaling. FASEB J. 2021, 35, e21393. [Google Scholar] [CrossRef]
- Samivel, R.; Nagarajan, R.P.; Subramanian, U.; Khan, A.A.; Masmali, A.; Almubrad, T.; Akhtar, S. Inhibitory Effect of Ursolic Acid on Ultraviolet B Radiation-Induced Oxidative Stress and Proinflammatory Response-Mediated Senescence in Human Skin Dermal Fibroblasts. Oxid. Med. Cell. Longev. 2020, 2020, 1246510. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Jiang, X.; Ma, L.; Li, Z.; Chang, Y.; Wang, Y.; Xue, C. Amino Acid Profiling with Chemometric Analysis as a Feasible Tool for the Discrimination of Marine-Derived Peptide Powders. Foods 2021, 10, 1294. [Google Scholar] [CrossRef]
- Peng, Z.; Gao, J.; Su, W.; Cao, W.; Zhu, G.; Qin, X.; Zhang, C.; Qi, Y. Purification and Identification of Peptides from Oyster (Crassostrea hongkongensis) Protein Enzymatic Hydrolysates and Their Anti-Skin Photoaging Effects on UVB-Irradiated HaCaT Cells. Mar. Drugs 2022, 20, 749. [Google Scholar] [CrossRef]
- Han, S.H.; Ballinger, E.; Choung, S.-Y.; Kwon, J.Y. Anti-Photoaging Effect of Hydrolysates from Pacific Whiting Skin via MAPK/AP-1, NF-κB, TGF-β/Smad, and Nrf-2/HO-1 Signaling Pathway in UVB-Induced Human Dermal Fibroblasts. Mar. Drugs 2022, 20, 308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-G.; Ying, X.-G.; Gao, P.; Wang, C.-L.; Wang, Y.-F.; Yu, X.-W.; Chen, J.; Wang, B.; Luo, H.-Y. Anti-Inflammatory Activity of a Peptide from Skipjack (Katsuwonus pelamis). Mar. Drugs 2019, 17, 582. [Google Scholar] [CrossRef] [PubMed]
- Zotchev, S.B. Marine actinomycetes as an emerging resource for the drug development pipelines. J. Biotechnol. 2012, 158, 168–175. [Google Scholar] [CrossRef]
- Moore, B.S.; Trischman, J.A.; Seng, D.; Kho, D.; Jensen, P.R.; Fenical, W. Salinamides, antiinflammatory depsipeptides from a marine streptomycete. J. Org. Chem. 1999, 64, 1145–1150. [Google Scholar] [CrossRef]
- Schultz, A.W.; Oh, D.-C.; Carney, J.R.; Williamson, R.T.; Udwary, D.W.; Jensen, P.R.; Gould, S.J.; Fenical, W.; Moore, B.S. Biosynthesis and structures of cyclomarins and cyclomarazines, prenylated cyclic peptides of marine actinobacterial origin. J. Am. Chem. Soc. 2008, 130, 4507–4516. [Google Scholar] [CrossRef] [PubMed]
- Eisner, L.; Vambutas, A.; Pathak, S. The Balance of Tissue Inhibitor of Metalloproteinase-1 and Matrix Metalloproteinase-9 in the Autoimmune Inner Ear Disease Patients. J. Interferon Cytokine Res. 2017, 37, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Choi, S.-I.; Jung, T.-D.; Cho, B.-Y.; Lee, J.-H.; Kim, S.-H.; Yoon, S.-A.; Ham, Y.-M.; Yoon, W.-J.; Cho, J.-H.; et al. Anti-Photoaging Effect of Jeju Putgyul (Unripe Citrus) Extracts on Human Dermal Fibroblasts and Ultraviolet B-induced Hairless Mouse Skin. Int. J. Mol. Sci. 2017, 18, 2052. [Google Scholar] [CrossRef]
- Vicentini, F.T.M.C.; He, T.; Shao, Y.; Fonseca, M.J.V.; Verri, W.A., Jr.; Fisher, G.J.; Xu, Y. Quercetin inhibits UV irradiation-induced inflammatory cytokine production in primary human keratinocytes by suppressing NF-kappa B pathway. J. Dermatol. Sci. 2011, 61, 162–168. [Google Scholar] [CrossRef]
- Wang, M.; Charareh, P.; Lei, X.; Zhong, J.L. Autophagy: Multiple Mechanisms to Protect Skin from Ultraviolet Radiation-Driven Photoaging. Oxid. Med. Cell. Longev. 2019, 2019, 8135985. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, S.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. Trehalose against UVB-induced skin photoaging by suppressing MMP expression and enhancing procollagen I synthesis in HaCaT cells. J. Funct. Foods 2020, 74, 104198. [Google Scholar] [CrossRef]
- Bang, J.S.; Choung, S.-Y. Inhibitory effect of oyster hydrolysate on wrinkle formation against UVB irradiation in human dermal fibroblast via MAPK/AP-1 and TGF beta/Smad pathway. J. Photochem. Photobiol. B-Biol. 2020, 209, 111946. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Kim, J.; Karadeniz, F.; Kim, H.R.; Park, S.Y.; Seo, Y.; Kong, C.-S. Santamarine Shows Anti-Photoaging Properties via Inhibition of MAPK/AP-1 and Stimulation of TGF-beta/Smad Signaling in UVA-Irradiated HDFs. Molecules 2021, 26, 3585. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-F.; Ma, K.-H.; Chang, Y.-J.; Lo, L.-C.; Jhap, T.-Y.; Su, Y.-H.; Liu, P.-S.; Chueh, S.-H. Baicalein inhibits matrix metalloproteinase 1 expression via activation of TRPV1-Ca-ERK pathway in ultraviolet B-irradiated human dermal fibroblasts. Exp. Dermatol. 2019, 28, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Kulaberoglu, Y.; Bhushan, B.; Hadi, F.; Chakrabarti, S.; Khaled, W.T.; Rankin, K.S.; Smith, E.S.J.; Frankel, D. The material properties of naked mole-rat hyaluronan. Sci. Rep. 2019, 9, 6632. [Google Scholar] [CrossRef]
- Swathi, N.; Srikanth, K.; Venipriya, S. Does the addition of hyaluronidase improve the quality of peribulbar anesthesia in cataract surgery?—A randomized double blinded study. Saudi J. Ophthalmol. 2018, 32, 204–210. [Google Scholar] [CrossRef]
- Robinson, D.M.; Vega, J.; Palm, M.D.; Bell, M.; Widgerow, A.D.; Giannini, A. Multicenter evaluation of a topical hyaluronic acid serum. J. Cosmet. Dermatol. 2022, 21, 3848–3858. [Google Scholar] [CrossRef]
- Lee, H.P.; Kim, D.S.; Park, S.H.; Shin, C.Y.; Woo, J.J.; Kim, J.W.; An, R.-B.; Lee, C.; Cho, J.Y. Antioxidant Capacity of Potentilla paradoxa Nutt. and Its Beneficial Effects Related to Anti-Aging in HaCaT and B16F10 Cells. Plants 2022, 11, 873. [Google Scholar] [CrossRef]
- Morgan, D.J.; Casulli, J.; Chew, C.; Connolly, E.; Lui, S.; Brand, O.J.; Rahman, R.; Jagger, C.; Hussell, T. Innate Immune Cell Suppression and the Link with Secondary Lung Bacterial Pneumonia. Front. Immunol. 2018, 9, 02943. [Google Scholar] [CrossRef]
- Lee, J.-O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B.; et al. Antimelanogenesis and skin-protective activities of Panax ginseng calyx ethanol extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef]
- Shukla, S.; Park, J.; Park, J.H.; Lee, J.S.; Kim, M. Development of novel Meju starter culture using plant extracts with reduced Bacillus cereus counts and enhanced functional properties. Sci. Rep. 2017, 7, 11409. [Google Scholar] [CrossRef]
- Han, N.; Kim, J.; Bae, J.H.; Kim, M.; Lee, J.Y.; Lee, Y.-Y.; Kang, M.S.; Han, D.; Park, S.; Kim, H.-J. Effect of Atmospheric-Pressure Plasma on Functional Compounds and Physiological Activities in Peanut Shells. Antioxidants 2022, 11, 2214. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; White, K.L., Jr.; Smith, D.C.; Bowlin, G.L. In vitro evaluations of innate and acquired immune responses to electrospun polydioxanone-elastin blends. Biomaterials 2009, 30, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, D.; Kielmanowicz, M.G.; Vigodman, S.; Hu, Y.P.; Chen, N.; Nkengne, A.; Oddos, T.; Fischer, D.; Seiberg, M.; Lin, C.B. A novel anti-ageing mechanism for retinol: Induction of dermal elastin synthesis and elastin fibre formation. Int. J. Cosmet. Sci. 2011, 33, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Min, S.-G.; Jo, Y.-J. Functionality of porcine skin hydrolysates produced by hydrothermal processing for liposomal delivery system. J. Food Biochem. 2018, 42, 12464. [Google Scholar] [CrossRef]
- Malaspina, P.; Catellani, E.; Burlando, B.; Brignole, D.; Cornara, L.; Bazzicalupo, M.; Candiani, S.; Obino, V.; De Feo, V.; Caputo, L.; et al. Depigmenting potential of lichen extracts evaluated by in vitro and in vivo tests. PeerJ 2020, 8, 9150. [Google Scholar] [CrossRef] [PubMed]
- Roh, E.; Kim, J.-E.; Kwon, J.Y.; Park, J.S.; Bode, A.M.; Dong, Z.; Lee, K.W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, G.; Liao, J.; Tang, M. Five alternative splicing variants of the TYR gene and their different roles in melanogenesis in the Muchuan black-boned chicken. Br. Poult. Sci. 2019, 60, 8–14. [Google Scholar] [CrossRef]
- Li, L.; Tang, Y.; Li, X.; Zhou, T.; Song, Q.; Li, A. Mechanism of skin whitening through San-Bai decoction-induced tyrosinase inhibition and discovery of natural products targeting tyrosinase. Medicine 2023, 102, e33420. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol Treats UVB-Induced Photoaging by Anti-MMP Expression, through Anti-Inflammatory, Antioxidant, and Antiapoptotic Properties, and Treats Photoaging by Upregulating VEGF-B Expression. Oxid. Med. Cell. Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef]
- Gao, X.; Luo, F.; Zhao, H. Cloves Regulate Na+-K+-ATPase to Exert Antioxidant Effect and Inhibit UVB Light-Induced Skin Damage in Mice. Oxid. Med. Cell. Longev. 2021, 2021, 5197919. [Google Scholar] [CrossRef]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Oh, S.-J.; Park, J.-Y.; Won, B.; Oh, Y.-T.; Yang, S.-C.; Shin, O.S. Asterias pectinifera-Derived Collagen Peptides Mixed with Halocynthia roretzi Extracts Exhibit Anti-Photoaging Activities during Exposure to UV Irradiation, and Antibacterial Properties. J. Microbiol. Biotechnol. 2022, 32, 1382–1389. [Google Scholar] [CrossRef]
- Han, S.-B.; Won, B.; Yang, S.-c.; Kim, D.-H. Asterias pectinifera derived collagen peptide-encapsulating elastic nanoliposomes for the cosmetic application. J. Ind. Eng. Chem. 2021, 98, 289–297. [Google Scholar] [CrossRef]
- Fan, J.; Zhuang, Y.; Li, B. Effects of Collagen and Collagen Hydrolysate from Jellyfish Umbrella on Histological and Immunity Changes of Mice Photoaging. Nutrients 2013, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Jiang, J.; Wang, J.; Zhou, Q.; Zhang, X. N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis. Mar. Drugs 2019, 17, 520. [Google Scholar] [CrossRef]
- Chen, C.-L.; Liou, S.-F.; Chen, S.-J.; Shih, M.-F. Protective effects of Chlorella-derived peptide on UVB-induced production of MMP-1 and degradation of procollagen genes in human skin fibroblasts. Regul. Toxicol. Pharmacol. 2011, 60, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, A.S.; Roecken, M.; Ghoreschi, K. Cutaneous immunology: Basics and new concepts. Semin. Immunopathol. 2016, 38, 3–10. [Google Scholar] [CrossRef]
- Wang, L.-F.; Hsu, C.-J.; Miaw, S.-C.; Chiu, H.-C.; Liu, C.-Y.; Yu, H.-S. Cross-priming with an epicutaneously introduced soluble protein antigen generates Tc1 cells. Eur. J. Immunol. 2006, 36, 2904–2911. [Google Scholar] [CrossRef]
- Semreen, M.H.; El-Gamal, M.I.; Abdin, S.; Alkhazraji, H.; Kamal, L.; Hammad, S.; El-Awady, F.; Waleed, D.; Kourbaj, L. Recent updates of marine antimicrobial peptides. Saudi Pharm. J. 2018, 26, 396–409. [Google Scholar] [CrossRef]
- Safronova, V.N.; Panteleev, P.V.; Sukhanov, S.V.; Toropygin, I.Y.; Bolosov, I.A.; Ovchinnikova, T.V. Mechanism of Action and Therapeutic Potential of the β-Hairpin Antimicrobial Peptide Capitellacin from the Marine Polychaeta Capitella teleta. Mar. Drugs 2022, 20, 167. [Google Scholar] [CrossRef]
- Ford, P.W.; Gustafson, K.R.; McKee, T.C.; Shigematsu, N.; Maurizi, L.K.; Pannell, L.K.; Williams, D.E.; de Silva, E.D.; Lassota, P.; Allen, T.M.; et al. Papuamides A-D, HIV-inhibitory and cytotoxic depsipeptides from the sponges Theonella mirabilis and Theonella swinhoei collected in Papua New Guinea. J. Am. Chem. Soc. 1999, 121, 5899–5909. [Google Scholar] [CrossRef]
- Jiao, K.; Gao, J.; Zhou, T.; Yu, J.; Song, H.; Wei, Y.; Gao, X. Isolation and purification of a novel antimicrobial peptide from Porphyra yezoensis. J. Food Biochem. 2019, 43, 12864. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dananjaya, S.H.S.; Chandrarathna, H.P.S.U.; De Zoysa, M.; Whang, I. Octominin: A Novel Synthetic Anticandidal Peptide Derived from Defense Protein of Octopus minor. Mar. Drugs 2020, 18, 56. [Google Scholar] [CrossRef]
- Gasu, E.N.; Ahor, H.S.; Borquaye, L.S. Peptide Mix from Olivancillaria hiatula Interferes with Cell-to-Cell Communication in Pseudomonas aeruginosa. Biomed Res. Int. 2019, 2019, 5313918. [Google Scholar] [CrossRef] [PubMed]
- Oh, R.; Lee, M.J.; Kim, Y.-O.; Nam, B.-H.; Kong, H.J.; Kim, J.-W.; Park, J.-Y.; Seo, J.-K.; Kim, D.-G. Myticusin-beta, antimicrobial peptide from the marine bivalve, Mytilus coruscus. Fish Shellfish Immunol. 2020, 99, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Sivakamavalli, J.; James, R.A.; Park, K.; Kwak, I.-S.; Vaseeharan, B. Purification of WAP domain-containing antimicrobial peptides from green tiger shrimp Peaneaus semisulcatus. Microb. Pathog. 2020, 140, 103920. [Google Scholar] [CrossRef] [PubMed]
- Sruthy, K.S.; Nair, A.; Antony, S.P.; Puthumana, J.; Singh, I.S.B.; Philip, R. A histone H2A derived antimicrobial peptide, Fi-Histin from the Indian White shrimp, Fenneropenaeus indicus: Molecular and functional characterization. Fish Shellfish Immunol. 2019, 92, 667–679. [Google Scholar] [CrossRef]
- Humisto, A.; Jokela, J.; Teigen, K.; Wahlsten, M.; Permi, P.; Sivonen, K.; Herfindal, L. Characterization of the interaction of the antifungal and cytotoxic cyclic glycolipopeptide hassallidin with sterol-containing lipid membranes. Biochim. Biophys. Acta-Biomembr. 2019, 1861, 1510–1521. [Google Scholar] [CrossRef]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.-E.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Song, R.; Wei, R.; Zhang, B.; Wang, D. Optimization of the Antibacterial Activity of Half-Fin Anchovy (Setipinna taty) Hydrolysates. Food Bioprocess Technol. 2012, 5, 1979–1989. [Google Scholar] [CrossRef]
- Ennaas, N.; Hammami, R.; Beaulieu, L.; Fliss, I. Purification and characterization of four antibacterial peptides from protamex hydrolysate of Atlantic mackerel (Scomber scombrus) by-products. Biochem. Biophys. Res. Commun. 2015, 462, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ennaas, N.; Hammami, R.; Beaulieu, L.; Fliss, I. Production of antibacterial fraction from Atlantic mackerel (Scomber scombrus) and its processing by-products using commercial enzymes. Food Bioprod. Process. 2015, 96, 145–153. [Google Scholar] [CrossRef]
- Tanaka, M.; Koyama, Y.-i.; Nomura, Y. Effects of Collagen Peptide Ingestion on UV-B-Induced Skin Damage. Biosci. Biotechnol. Biochem. 2009, 73, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Ding, Y.; Dai, X.; Li, Y. Oral administration of marine collagen peptides from Chum Salmon skin enhances cutaneous wound healing and angiogenesis in rats. J. Sci. Food Agric. 2011, 91, 2173–2179. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.; Mikhal’chik, E.V.; Suprun, M.V.; Papacharalambous, M.; Truhanov, A.I.; Korkina, L.G. Skin Antiageing and Systemic Redox Effects of Supplementation with Marine Collagen Peptides and Plant-Derived Antioxidants: A Single-Blind Case-Control Clinical Study. Oxid. Med. Cell. Longev. 2016, 2016, 4389410. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Li, B.; Zhang, Z.; Xue, C.; Yu, G.; Wang, J.; Bao, Y.; Bu, L.; Sun, J.; Peng, Z.; et al. Moisture absorption and retention properties, and activity in alleviating skin photodamage of collagen polypeptide from marine fish skin. Food Chem. 2012, 135, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Zhao, X.; Li, B.; Zhang, Z.; Zhuang, Y. Inhibition of melanogenic activity by gelatin and polypeptides from pacific cod skin in b16 melanoma cells. J. Food Biochem. 2011, 35, 1099–1116. [Google Scholar] [CrossRef]
- Oh, J.-Y.; Lee, H.G.; Je, J.-G.; Wang, L.; Kim, H.-S.; Jeon, Y.-J. Evaluation of Cosmeceutical Properties of Fish Skin By-product Hydrolysates Collected during Surimi Manufacturing Process. Korean J. Fish. Aquat. Sci. 2020, 53, 297–307. [Google Scholar] [CrossRef]
- Al-Atif, H. Collagen Supplements for Aging and Wrinkles: A Paradigm Shift in the Fields of Dermatology and Cosmetics. Dermatol. Pract. Concept. 2022, 12, e2022018. [Google Scholar] [CrossRef]
- Pozzolini, M.; Millo, E.; Oliveri, C.; Mirata, S.; Salis, A.; Damonte, G.; Arkel, M.; Scarfi, S. Elicited ROS Scavenging Activity, Photoprotective, and Wound-Healing Properties of Collagen-Derived Peptides from the Marine Sponge Chondrosia reniformis. Mar. Drugs 2018, 16, 465. [Google Scholar] [CrossRef]
- Lin, H.; Zheng, Z.; Yuan, J.; Zhang, C.; Cao, W.; Qin, X. Collagen Peptides Derived from Sipunculus nudus Accelerate Wound Healing. Molecules 2021, 26, 1385. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, M.; Liang, R.; Zhao, M.; Zhang, Z.; Li, Y. Oral administration of marine collagen peptides prepared from chum salmon (Oncorhynchus keta) improves wound healing following cesarean section in rats. Food Nutr. Res. 2015, 59, 26411. [Google Scholar] [CrossRef]
- Leon-Lopez, A.; Morales-Penaloza, A.; Manuel Martinez-Juarez, V.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Alvarez, G. Hydrolyzed Collagen-Sources and Applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed]
- Tsitsipatis, D.; Martindale, J.L.; Ubaida-Mohien, C.; Lyashkov, A.; Yanai, H.; Kashyap, A.; Shin, C.H.; Herman, A.B.; Ji, E.; Yang, J.-H.; et al. Proteomes of primary skin fibroblasts from healthy individuals reveal altered cell responses across the life span. Aging Cell 2022, 21, 13609. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef]
- Burki, T. Skin-whitening creams: Worth the risk? Lancet Diabetes Endocrinol. 2021, 9, 10. [Google Scholar] [CrossRef]
- Boyajian, J.L.; Ghebretatios, M.; Schaly, S.; Islam, P.; Prakash, S. Microbiome and Human Aging: Probiotic and Prebiotic Potentials in Longevity, Skin Health and Cellular Senescence. Nutrients 2021, 13, 4550. [Google Scholar] [CrossRef]
- Mansinhos, I.; Goncalves, S.; Rodriguez-Solana, R.; Ordonez-Diaz, J.L.; Moreno-Rojas, J.M.; Romano, A. Ultrasonic-Assisted Extraction and Natural Deep Eutectic Solvents Combination: A Green Strategy to Improve the Recovery of Phenolic Compounds from Lavandula pedunculata subsp. lusitanica (Chaytor) Franco. Antioxidants 2021, 10, 582. [Google Scholar] [CrossRef]
- Costa, C.; Anselmo, H.; Ferro, R.; Matos, A.S.; Casimiro, T.; Aguiar-Ricardo, A. Dry Dosage Forms of Add-Value Bioactive Phenolic Compounds by Supercritical CO2-Assisted Spray-Drying. Molecules 2022, 27, 2001. [Google Scholar] [CrossRef]
- Xing, L.; Wang, Z.; Hao, Y.; Zhang, W. Marine Products As a Promising Resource of Bioactive Peptides: Update of Extraction Strategies and Their Physiological Regulatory Effects. J. Agric. Food Chem. 2022, 70, 3081–3095. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, G.; Han, K.; Sun, D.; Zhou, N.; Song, Z.; Liu, H.; Li, J.; Li, G. Applications of Molecular Imprinting Technology in the Study of Traditional Chinese Medicine. Molecules 2023, 28, 301. [Google Scholar] [CrossRef] [PubMed]
- Barzegar, F.; Kamankesh, M.; Mohammadi, A. Heterocyclic aromatic amines in cooked food: A review on formation, health risk-toxicology and their analytical techniques. Food Chem. 2019, 280, 240–254. [Google Scholar] [CrossRef]
- Wu, H.; Li, J.; Jia, Y.; Xiao, Z.; Li, P.; Xie, Y.; Zhang, A.; Liu, R.; Ren, Z.; Zhao, M.; et al. Essential Oil Extracted from Cymbopogon citronella Leaves by Supercritical Carbon Dioxide: Antioxidant and Antimicrobial Activities. J. Anal. Methods Chem. 2019, 2019, 8192439. [Google Scholar] [CrossRef]
- Marcet, I.; Alvarez, C.; Paredes, B.; Diaz, M. The use of sub-critical water hydrolysis for the recovery of peptides and free amino acids from food processing wastes. Review of sources and main parameters. Waste Manag. 2016, 49, 364–371. [Google Scholar] [CrossRef]
- Melgosa, R.; Trigueros, E.; Teresa Sanz, M.; Cardeira, M.; Rodrigues, L.; Fernandez, N.; Matias, A.A.; Bronze, M.R.; Marques, M.; Paiva, A.; et al. Supercritical CO2 and subcritical water technologies for the production of bioactive extracts from sardine (Sardina pilchardus) waste. J. Supercrit. Fluids 2020, 164, 104943. [Google Scholar] [CrossRef]
- Rivas-Vela, C.I.; Amaya-Llano, S.L.; Castano-Tostado, E.; Castillo-Herrera, G.A. Protein Hydrolysis by Subcritical Water: A New Perspective on Obtaining Bioactive Peptides. Molecules 2021, 26, 6655. [Google Scholar] [CrossRef]
- Gomez, B.; Munekata, P.E.S.; Gavahian, M.; Barba, F.J.; Marti-Quijal, F.J.; Bolumar, T.; Bastianello Campagnol, P.C.; Tomasevic, I.; Lorenzo, J.M. Application of pulsed electric fields in meat and fish processing industries: An overview. Food Res. Int. 2019, 123, 95–105. [Google Scholar] [CrossRef]
- Li, M.; Lin, J.; Chen, J.; Fang, T. pulsed electric field-assisted enzymatic extraction of protein from abalone (haliotis discus hannai ino) viscera. J. Food Process Eng. 2016, 39, 702–710. [Google Scholar] [CrossRef]
- Akaberi, S.; Gusbeth, C.; Silve, A.; Senthilnathan, D.S.; Navarro-Lopez, E.; Molina-Grima, E.; Muller, G.; Frey, W. Effect of pulsed electric field treatment on enzymatic hydrolysis of proteins of Scenedesmus almeriensis. Algal Res.-Biomass Biofuels Bioprod. 2019, 43, 101656. [Google Scholar] [CrossRef]
- Di, D.-L.; Zheng, Y.-Y.; Chen, X.-F.; Huang, X.-Y.; Feng, S.-L. Advance of Application of High Speed Counter-current Chromatography in Separation and Purification of Flavonoids. Chin. J. Anal. Chem. 2011, 39, 269–275. [Google Scholar] [CrossRef]
- Ramezanzade, L.; Hosseini, S.F.; Nikkhah, M. Biopolymer-coated nanoliposomes as carriers of rainbow trout skin-derived antioxidant peptides. Food Chem. 2017, 234, 220–229. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoscale Nutrient Delivery Systems for Food Applications: Improving Bioactive Dispersibility, Stability, and Bioavailability. J. Food Sci. 2015, 80, N1602–N1611. [Google Scholar] [CrossRef]
- Yao, M.; Xiao, H.; McClements, D.J. Delivery of Lipophilic Bioactives: Assembly, Disassembly, and Reassembly of Lipid Nanoparticles. Annu. Rev. Food Sci. Technol. 2014, 5, 53–81. [Google Scholar] [CrossRef]
- Choonpicharn, S.; Tateing, S.; Jaturasitha, S.; Rakariyatham, N.; Suree, N.; Niamsup, H. Identification of bioactive peptide from Oreochromis niloticus skin gelatin. J. Food Sci. Technol.-Mysore 2016, 53, 1222–1229. [Google Scholar] [CrossRef]
- Desai, K.G.H. Chitosan Nanoparticles Prepared by Ionotropic Gelation: An Overview of Recent Advances. Crit. Rev. Ther. Drug Carrier Syst. 2016, 33, 107–158. [Google Scholar] [CrossRef]
- Hosseini, S.F.; Ramezanzade, L.; McClements, D.J. Recent advances in nanoencapsulation of hydrophobic marine bioactives: Bioavailability, safety, and sensory attributes of nano-fortified functional foods. Trends Food Sci. Technol. 2021, 109, 322–339. [Google Scholar] [CrossRef]
- Asserin, J.; Lati, E.; Shioya, T.; Prawitt, J. The effect of oral collagen peptide supplementation on skin moisture and the dermal collagen network: Evidence from an ex vivo model and randomized, placebo-controlled clinical trials. J. Cosmet. Dermatol. 2015, 14, 291–301. [Google Scholar] [CrossRef]
- Maia Campos, P.M.B.G.; Franco, R.S.B.; Kakuda, L.; Cadioli, G.F.; Costa, G.M.D.A.; Bouvret, E. Oral Supplementation with Hydrolyzed Fish Cartilage Improves the Morphological and Structural Characteristics of the Skin: A Double-Blind, Placebo-Controlled Clinical Study. Molecules 2021, 26, 4880. [Google Scholar] [CrossRef] [PubMed]
- Finn Canada. Available online: https://www.finncanada.com/ (accessed on 29 September 2022).
- Kenney & Ross Limited. Kenney and Ross. Available online: https://www.kenneyandross.com/index.php?option=com_content&view=featured&Itemid=101 (accessed on 11 November 2022).
- Nuwen. Marine Active. Available online: https://www.nuwen.com/en/gamme/ingredients-en/marine-actives-ingredients-en/ (accessed on 18 November 2022).
- OneOcean. Collagen Boosting Powerhouses. Available online: https://oneoceanbeauty.com/collections/collagen (accessed on 15 November 2022).
- Ostrealia. Available online: https://www.ostrealia.fr/en/categorie-produit/cosmetics/ (accessed on 11 November 2022).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Enzyme Used | Peptides (Amino Acid Sequence) | Mechanism of Action | In Vivo or In Vitro | Reference |
|---|---|---|---|---|---|
| Tuna eggs | - | Ile-Cys-Arg-Asp and Leu-Cys-Gly-Glu-Cys | Inhibition of DPPH radicals and activation of SOD and GSH-Px | in vivo | [21] |
| Boiled abalone by-products | - | Ala-Thr-Pro-Gly-Asp-Glu-Gly | Inhibition of ROS radicals | in vitro | [22] |
| Jellyfish collagen | Pepsin | - | Activation of total antioxidant activity | in vitro | [23] |
| Rhopilema esculentum | Pepsin | - | Activation of SOD, CAT, and GSH-Px | in vivo | [24] |
| Salmon skin | - | - | Activation of SOD, CAT, and GSH-Px | in vivo | [25] |
| Katsuwonus pelamis | - | TCP3, TCP6, and TCP9 | Activation of SOD, CAT, and GSH-Px | in vitro | [26] |
| Tilapia gelatin | - | Leu-Ser-Gly-Tyr-Gly-Pro | Scavenging free radicals | in vitro | [27] |
| Katsuwonus pelamis | - | - | Scavenging free radicals | in vitro | [28] |
| Monkfish | Trypsin | Glu-Trp-Pro-Ala-Gln, Phe-Leu-His-Arg-Pro, and Leu-Met-Gly-Gln-Trp | Inhibition of DPPH radicals and hydroxyl radicals; activation of SOD, CAT, and GSH-Px | in vitro | [29] |
| Macroalga P. palmata | Corolase PP | Ser-Asp-Ile-Thr-Arg-Pro-Gly-Gly-Asn-Met | Activation of oxygen radical absorption capacity (ORAC) and iron reduction antioxidant capacity (FRAP) | in vitro | [30] |
| Thunnus obesus | Alcalase, α-chymotrypsin, neutrase, papain, pepsin, and trypsin | H-Leu-Asn-Leu-Pro-Thr-Ala-Val-Tyr-Met-Val-Thr-OH | Inhibition of DPPH, hydroxyl, superoxide, and alkyl radicals | in vitro | [31] |
| Magalaspis cordyla | Pepsin/trypsin, and α-chymotrypsin | Asn-His-Arg-Tyr-Asp-Arg | Inhibition of DPPH and hydroxyl radicals | in vitro | [32] |
| Otolithes ruber | pepsin/trypsin and α-chymotrypsin | Gly-Asn-Arg-Gly-Phe-Ala-Cys-Arg-His-Ala | Inhibition of DPPH and hydroxyl radicals | in vitro | [32] |
| Hypoptychus dybowskii | Alcalase, neutrase, α-chymotrypsin, papain, pepsin, and trypsin | Ile–Val–Gly–Gly–Phe–Pro–His–Tyr–Leu | Inhibition of DPPH radicals | in vitro | [33] |
| Oreochromis niloticus | Alcalase, pronase E, pepsin, and trypsin | Asp-Pro-Ala-Leu-Ala-Thr-Glu-Pro-Asp-Pro-Met-Pro-Phe | Inhibition of DPPH, hydroxyl, and superoxide radicals | in vitro | [34] |
| Decapterus maruadsi | Alcalase, neutral protease, papain, pepsin, and trypsin | His-Asp-His-Pro-Val-Cys and His-Glu-Lys-Val-Cys | Inhibition of DPPH and hydroxyl radicals | in vitro | [35] |
| Johnius belengerii | Trypsin, R-chymotrypsin, and pepsin | His-Gly-Pro-Leu-Gly-Pro-Leu | Inhibition of DPPH radicals | in vitro | [36] |
| Paralichthys olivaceus | Papain, pepsin, trypsin, neutrase, alcalase, kojizyme, protamex, and α-chymotrypsin | Val-Cys-Ser-Val and Cys-Ala-Ala-Pro | Inhibition of DPPH radicals | in vitro | [37] |
| Source | Enzyme Used | Peptides (Amino Acid Sequence) | Microorganisms | Reference |
|---|---|---|---|---|
| Capitella teleta | - | - | E. coli BL21 | [110] |
| Porphyra yezoensis | Pepsin | Thr-Pro-Asp-Ser-Glu-Ala-Leu | Staphylococcus aureus | [112] |
| Octopus minor | - | Gly-Trp-Leu-Ile-Arg-Gly-Ala-Ile-His-Ala-Gly-Lys-Ala-Ile-His-Gly-Leu-Ile-His-Arg-Arg-Arg-His | Candida albicans | [113] |
| Olivancillaria hiatula | - | - | Pseudomonas aeruginosa | [114] |
| Mytilus coruscus | - | - | Gram-positive bacteria —Bacillus, Bacillus subtilis, Clostridium perfringens, Staphylococcus aureus, Streptococcus, Streptococcus mutans; Gram-negative bacteria—Escherichia coli, Pseudomonas aeruginosa, Vibrio alginolyticus | [115] |
| Green tiger shrimp (Peaneaus semisulcatus) | - | - | Staphylococcus aureus | [116] |
| Hypoptychus dybowskii | - | Ser-Arg-Ser-Ser-Arg-Ala-Gly-Leu-Gln-Phe-Pro-Val-Gly-Arg-Ile-His-Arg-Leu-Leu-Arg-Lys | Staphylococcus aureus and Escherichia coli | [117] |
| Cyanobacteria | - | - | Candida albicans | [118] |
| Source | Functional Product | Processing Method | Cosmeceutical Function | Reference |
|---|---|---|---|---|
| Salmon skin | Collagen peptides | Water, protease | Wound healing | [124] |
| Fish scales | Collagen peptides | Hot water, enzymatic | Improving skin elasticity | [125] |
| Codfish skin | Collagen polypeptides | Water, pepsin, and alkaline protease | Moisturizer, antioxidant | [126] |
| Pacific whiting skin | Hydrolysate gelatin | Hot water | Anti-photoaging, delayed skin wrinkling | [71] |
| Pacific cod skin | Gelatin and polypeptides | Hot water extraction, pepsin, and alkaline protease hydrolysis | Melanogenesis inhibition | [127] |
| Olive flounder and Alaska pollock skins | Fish skin hydrolysates | Enzymatic hydrolysis (pepsin, alcalase, protemax) | Minimizing ROS levels, enhancing the viability of UVB-irradiated HaCat cells and human dermal fibroblasts | [128] |
| Scales of Tilapia zillii | Polypeptides | Pepsin | Increasing skin hydration and decreasing epidermal hyperplasia | [123] |
| Company | Country | By-Product Resource | Bioactive Compounds | Cosmeceutical Function | Reference |
|---|---|---|---|---|---|
| Rousselot | France | Fisk skin and bone | Collagen peptides | Skin moisturization, enhanced skin collagen density | [157] |
| Celergen Inc | Switzerland | Fish skin | Collagen hydrolysate | Enhanced skill elasticity | [125] |
| Abyss | France | Fish skin | Collagen hydrolysate | Reduced appearance of wrinkles | [158] |
| Finn Canada | Canada | Salmon skin | Collagen | Improved skin condition; treatment of various skin problems, such as wrinkles, spots, dryness, dullness, and acne | [159] |
| Kenney and Ross Limited | Canada | Fish skin | Collagen | Stimulates healthy skin, nails, and hair | [160] |
| Nuwen | France | Fish skin | Collagen hydrolysate | Skin moisturization | [161] |
| One Ocean | United States | Fish skin | Collagen | Skin moisturization, anti-wrinkle | [162] |
| Osteralia | France | Mother-of-pearl | Oyster shell | Anti-aging, skin nourishment | [163] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhuang, H.; Wu, S.; Mao, C.; Dai, Y.; Yan, H. Marine Bioactive Peptides: Anti-Photoaging Mechanisms and Potential Skin Protective Effects. Curr. Issues Mol. Biol. 2024, 46, 990-1009. https://doi.org/10.3390/cimb46020063
Zhang X, Zhuang H, Wu S, Mao C, Dai Y, Yan H. Marine Bioactive Peptides: Anti-Photoaging Mechanisms and Potential Skin Protective Effects. Current Issues in Molecular Biology. 2024; 46(2):990-1009. https://doi.org/10.3390/cimb46020063
Chicago/Turabian StyleZhang, Xiaoliang, Hong Zhuang, Sijia Wu, Chen Mao, Yaxi Dai, and Haiyang Yan. 2024. "Marine Bioactive Peptides: Anti-Photoaging Mechanisms and Potential Skin Protective Effects" Current Issues in Molecular Biology 46, no. 2: 990-1009. https://doi.org/10.3390/cimb46020063
APA StyleZhang, X., Zhuang, H., Wu, S., Mao, C., Dai, Y., & Yan, H. (2024). Marine Bioactive Peptides: Anti-Photoaging Mechanisms and Potential Skin Protective Effects. Current Issues in Molecular Biology, 46(2), 990-1009. https://doi.org/10.3390/cimb46020063