
Beeswax Alcohol Prevents Low-Density Lipoprotein Oxidation and Demonstrates Antioxidant Activities in Zebrafish Embryos and Human Subjects: A Clinical Study

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. DPPH Free Radical Scavenging Activity
2.3. Isolation of LDL and HDL
2.4. Paraoxonase Assay
2.5. Oxidation of LDL, Electrophoresis, and Quantification of LDL-Oxidized Products
2.6. Electron Microscopic Examination of LDL
2.7. Exposure of HDL to Ferrous Ions
2.8. Microinjection of Zebrafish Embryos
2.9. Reactive Oxygen Species (ROS) and Apoptosis in Zebrafish Embryos
2.10. Clinical Study
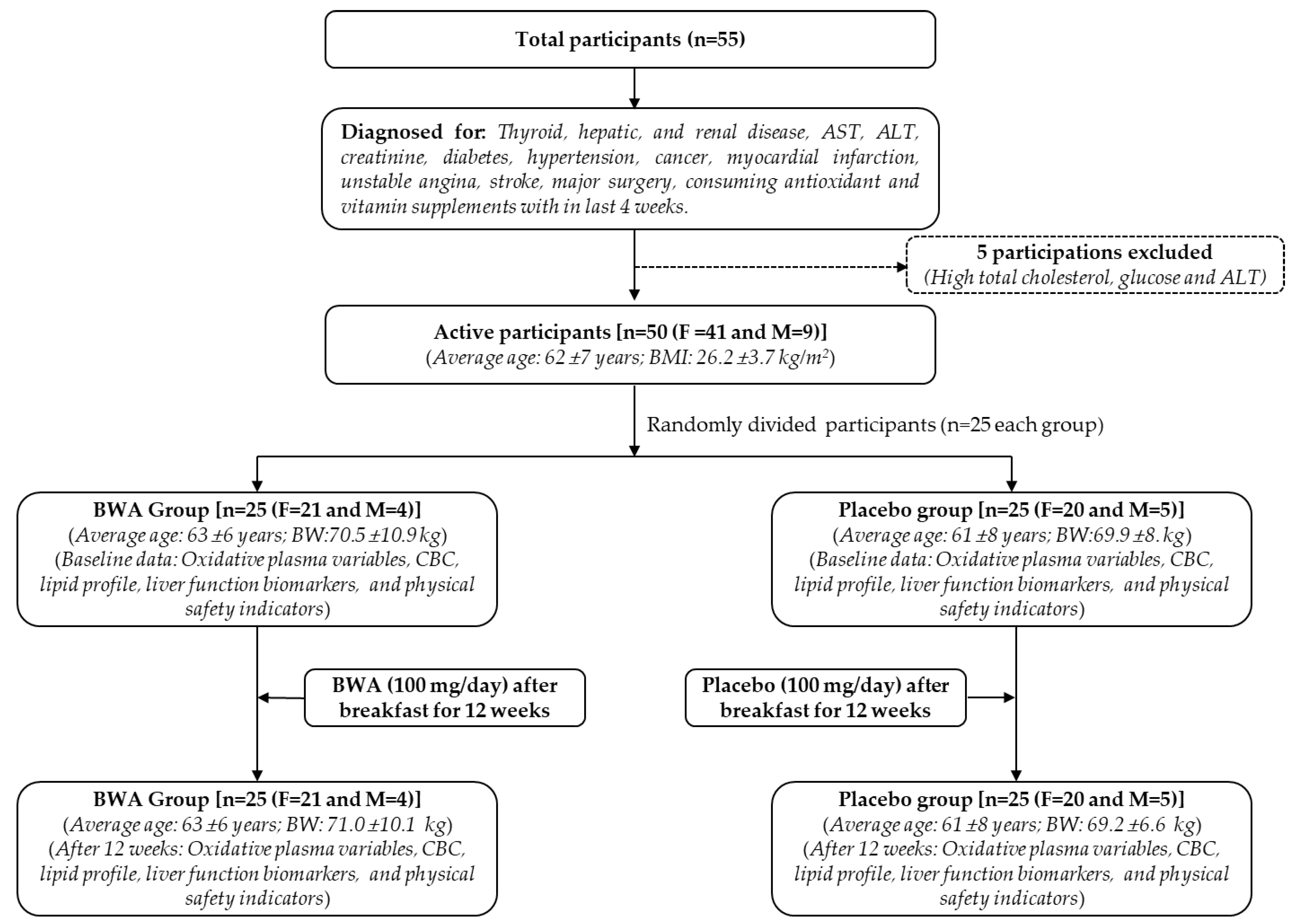
2.10.1. Participants
2.10.2. Study Layout
2.10.3. Anthropometric and Blood Analysis
2.10.4. Oxidative Variables
2.11. Statistical Analysis
3. Results and Discussion
3.1. Antioxidant Activity of BWA
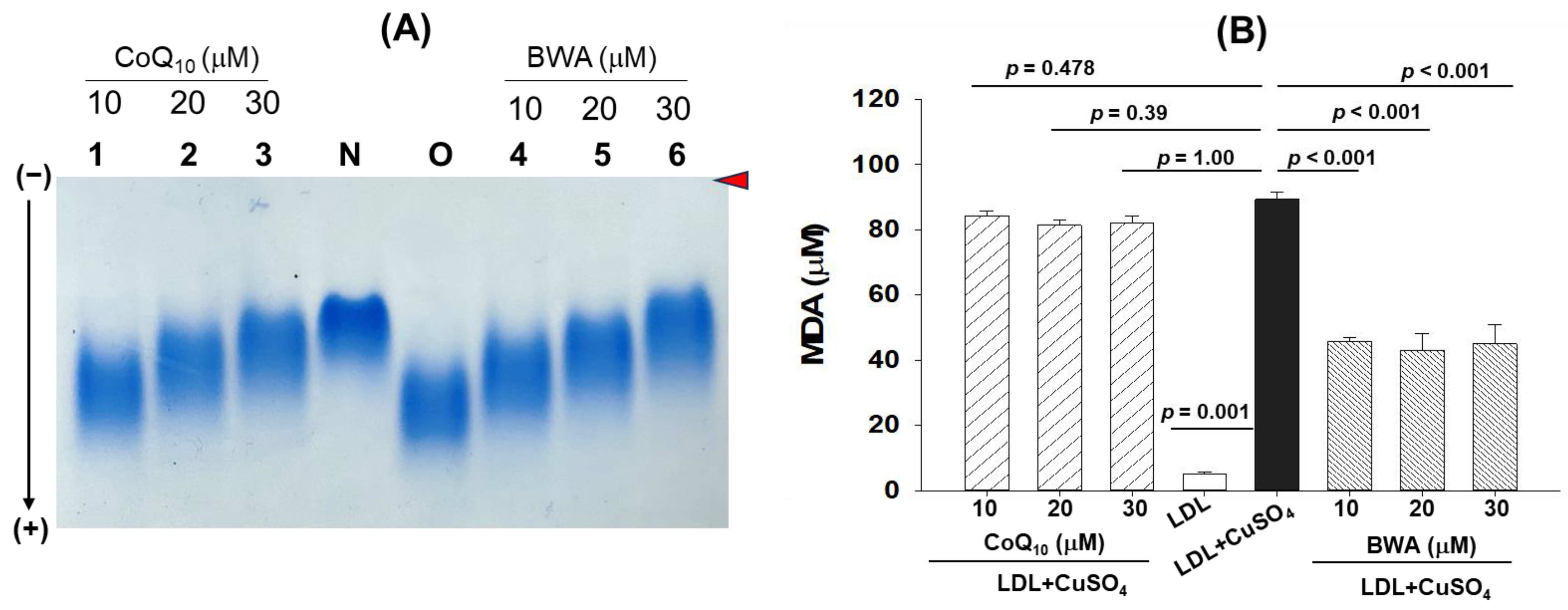
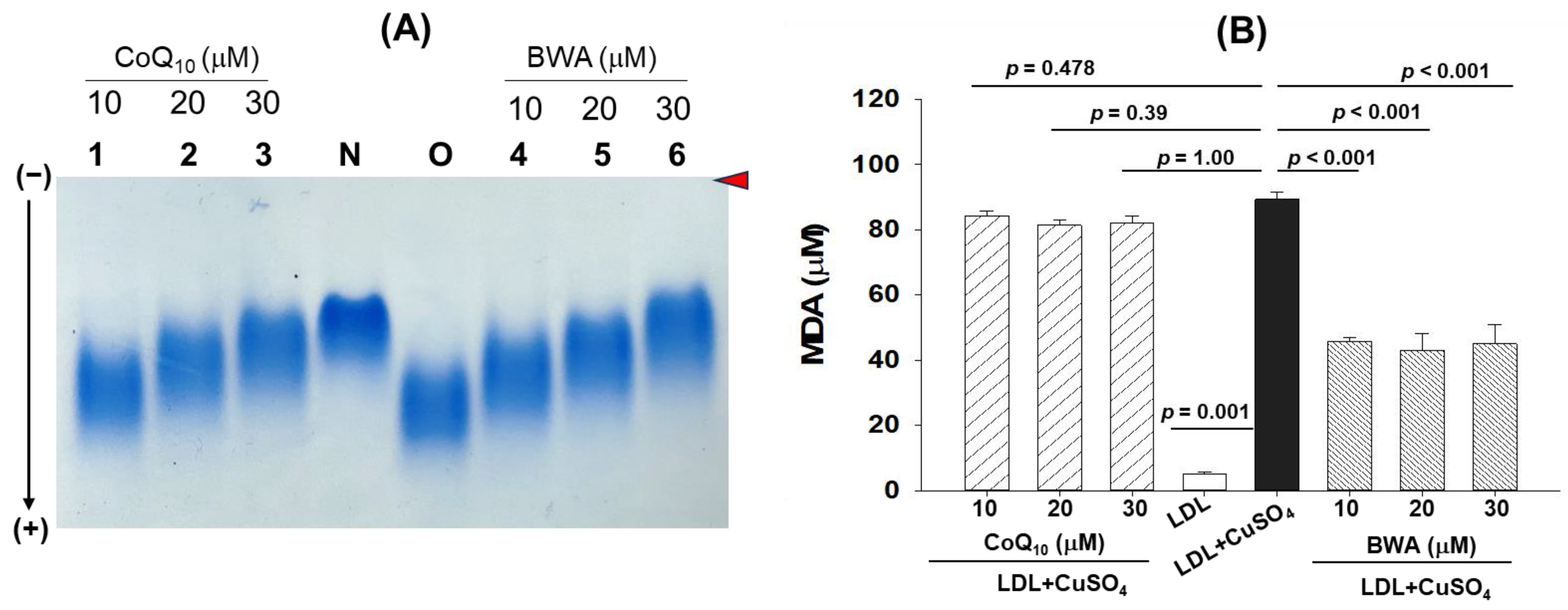
3.2. Inhibition of Cupric Ion (Cu2+)-Mediated LDL Oxidation by BWA
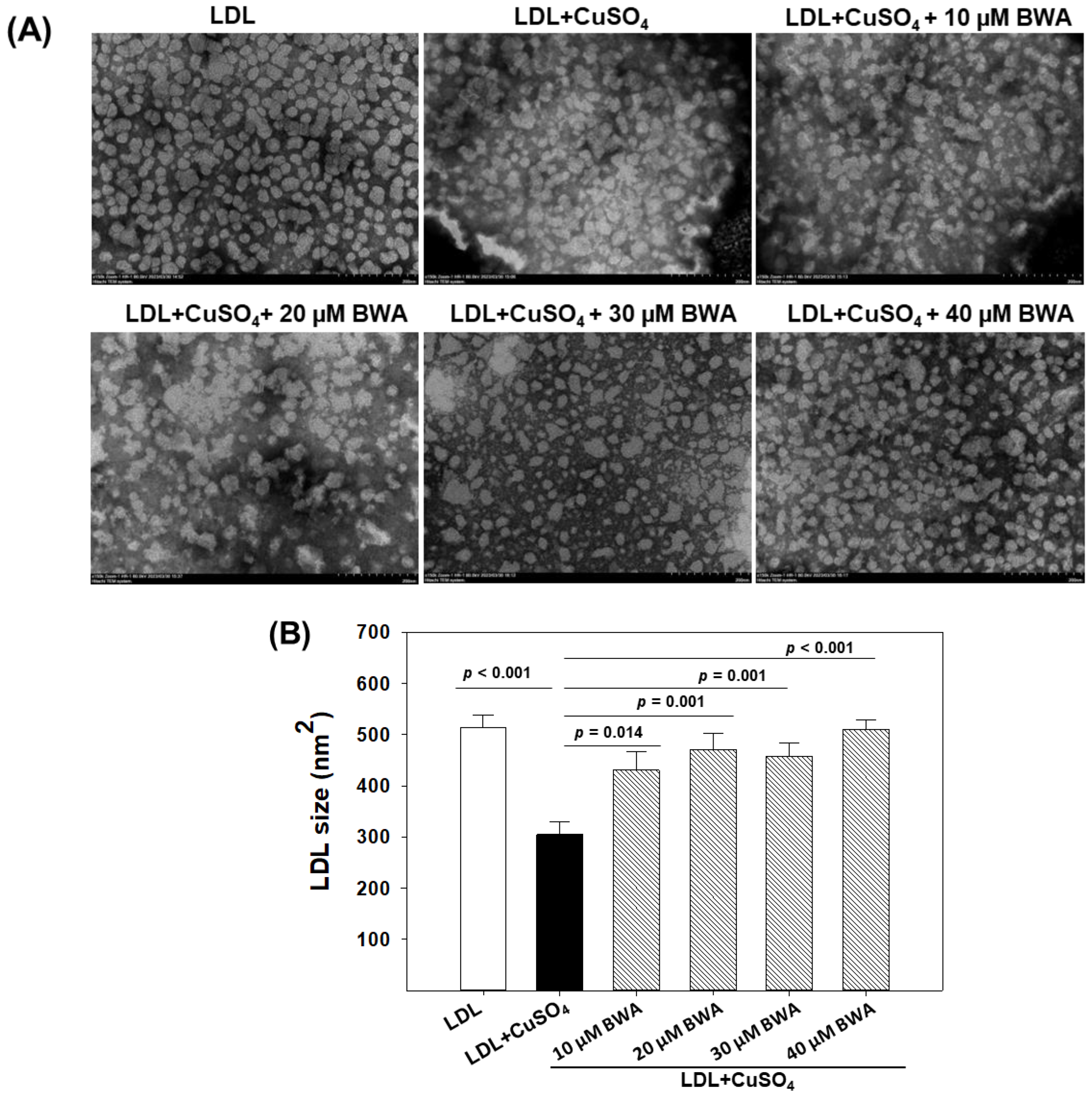
3.3. BWA Restored Cupric Ion (Cu2+)-Mediated Morphological Changes in LDL
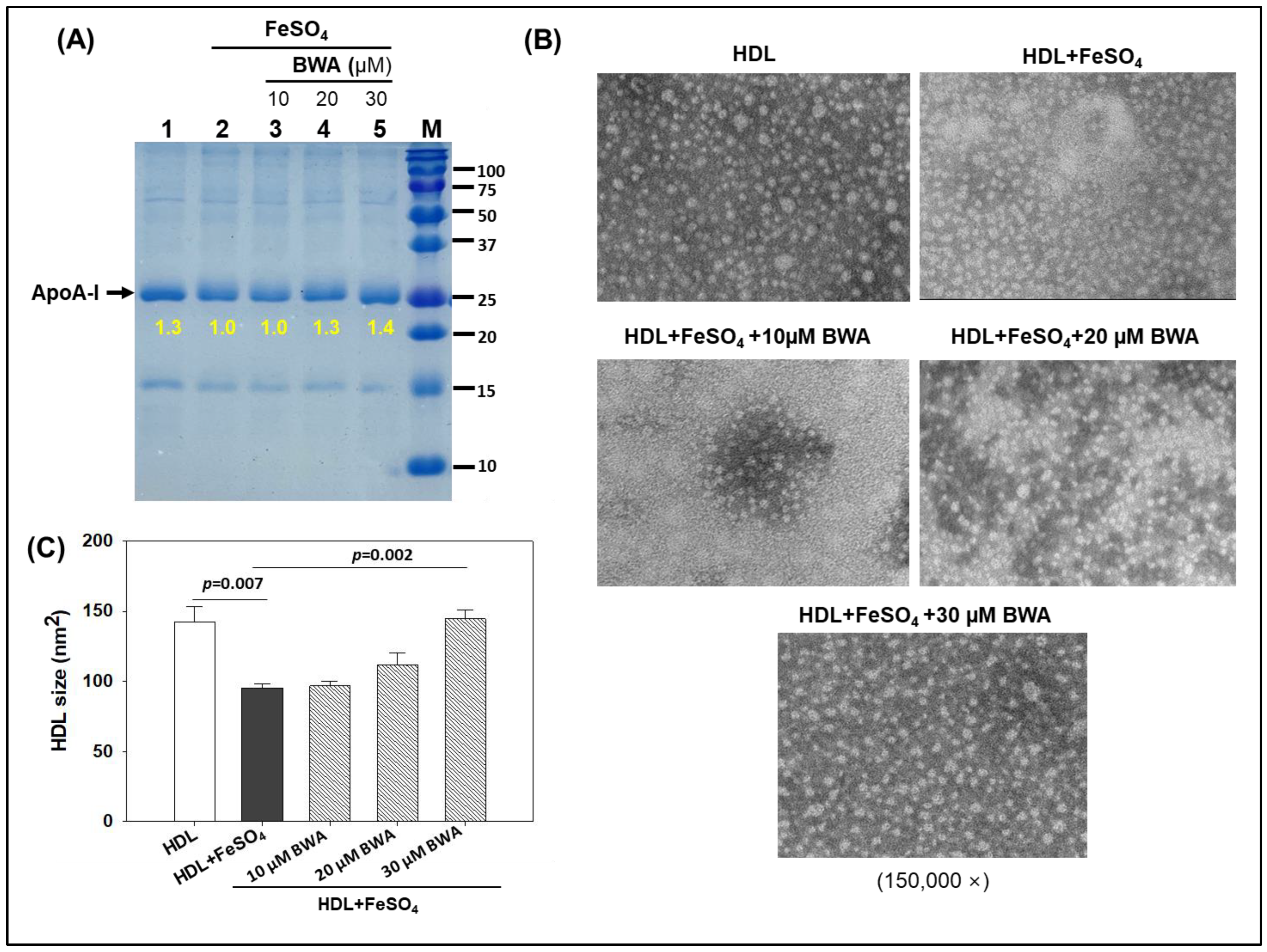
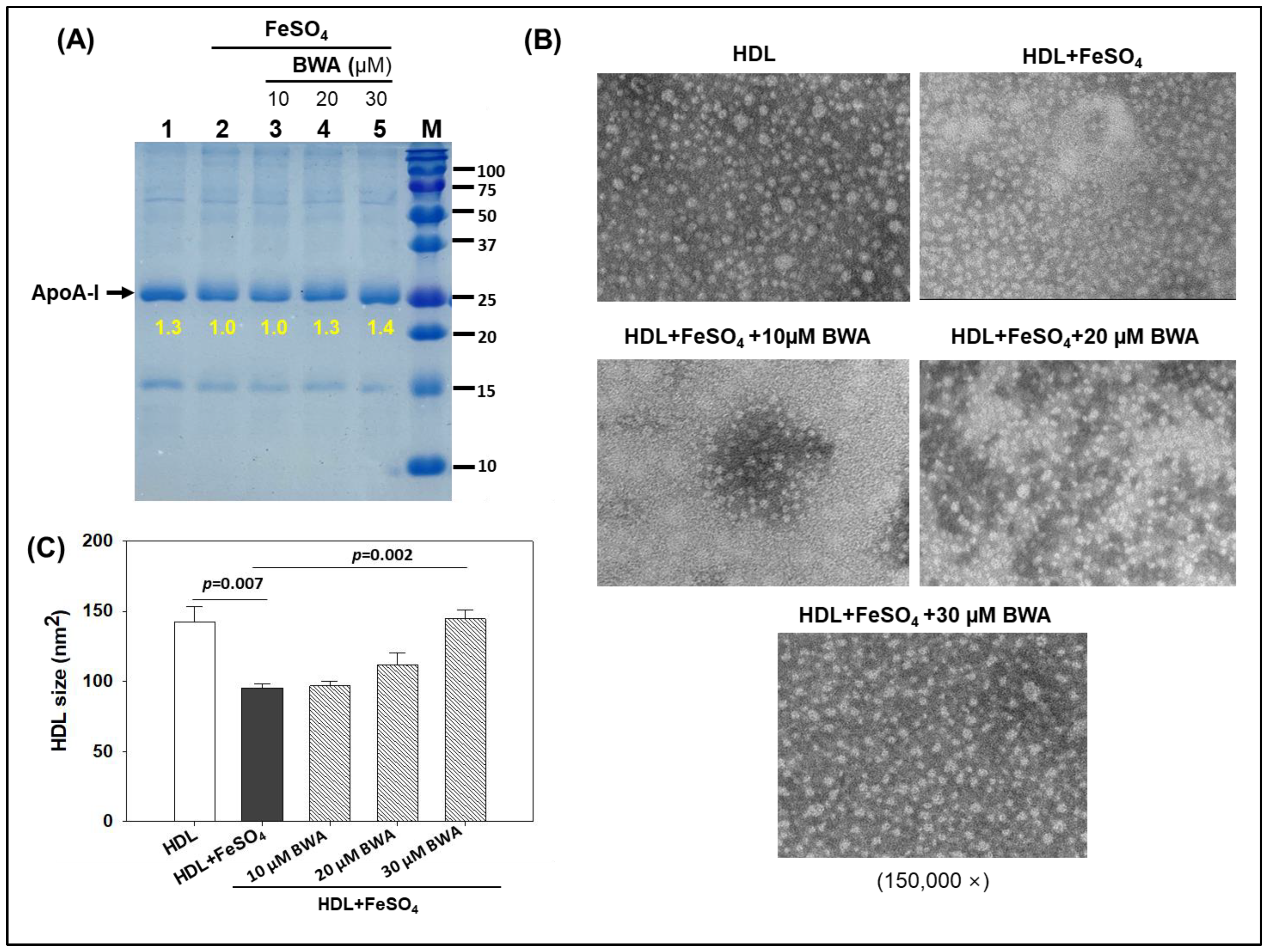
3.4. BWA Prevents HDL Degradation by Ferrous Ions (Fe2+)
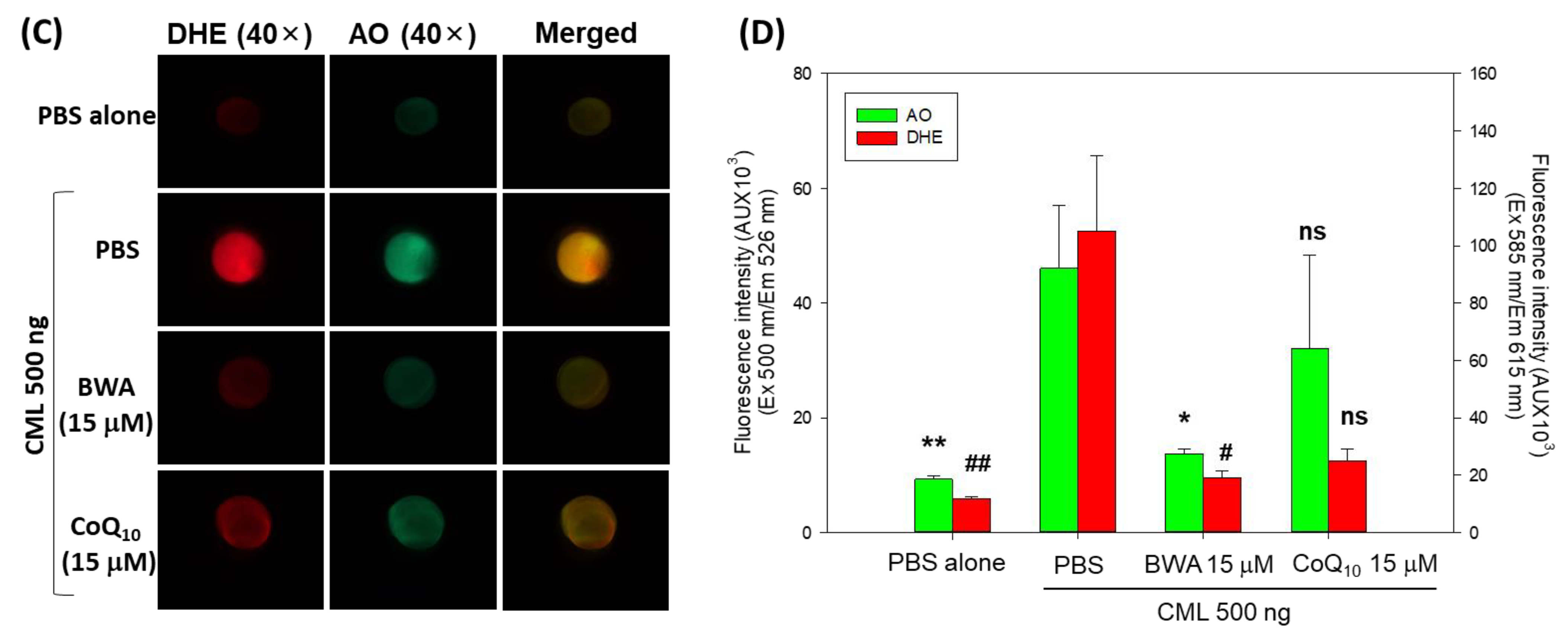
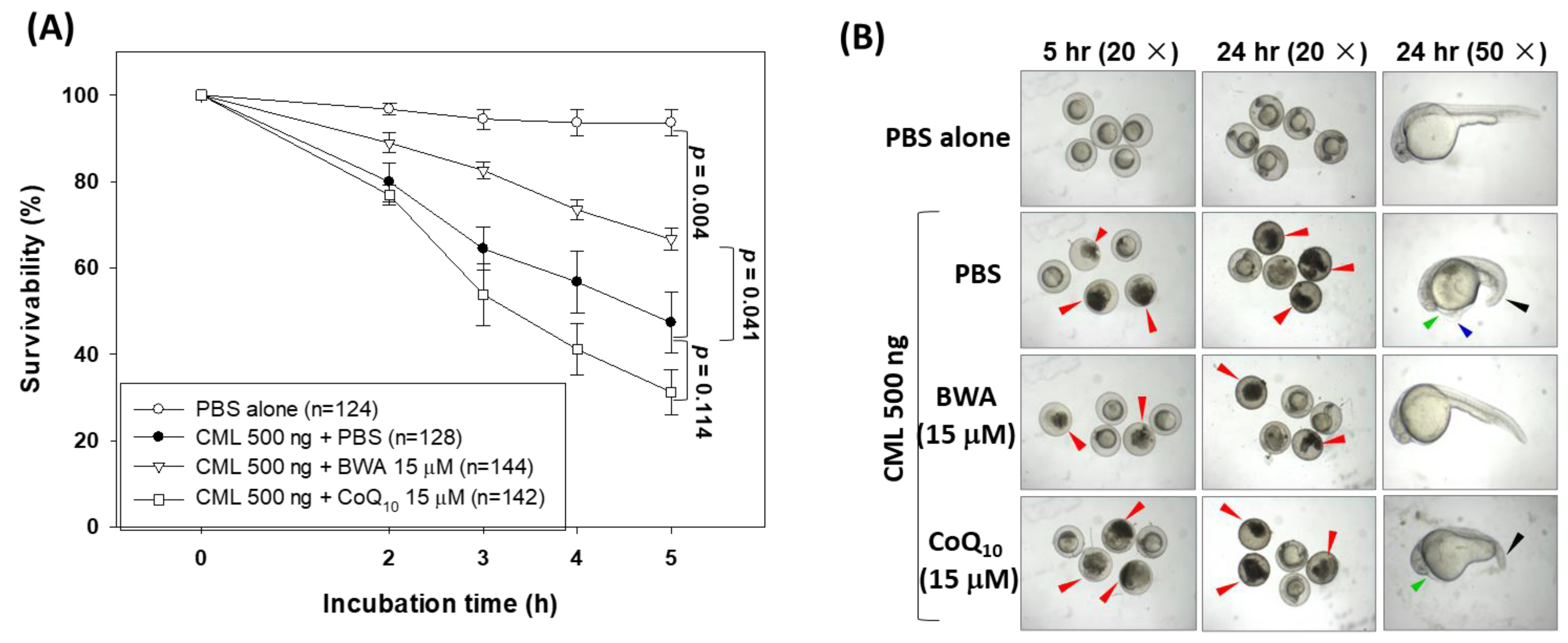
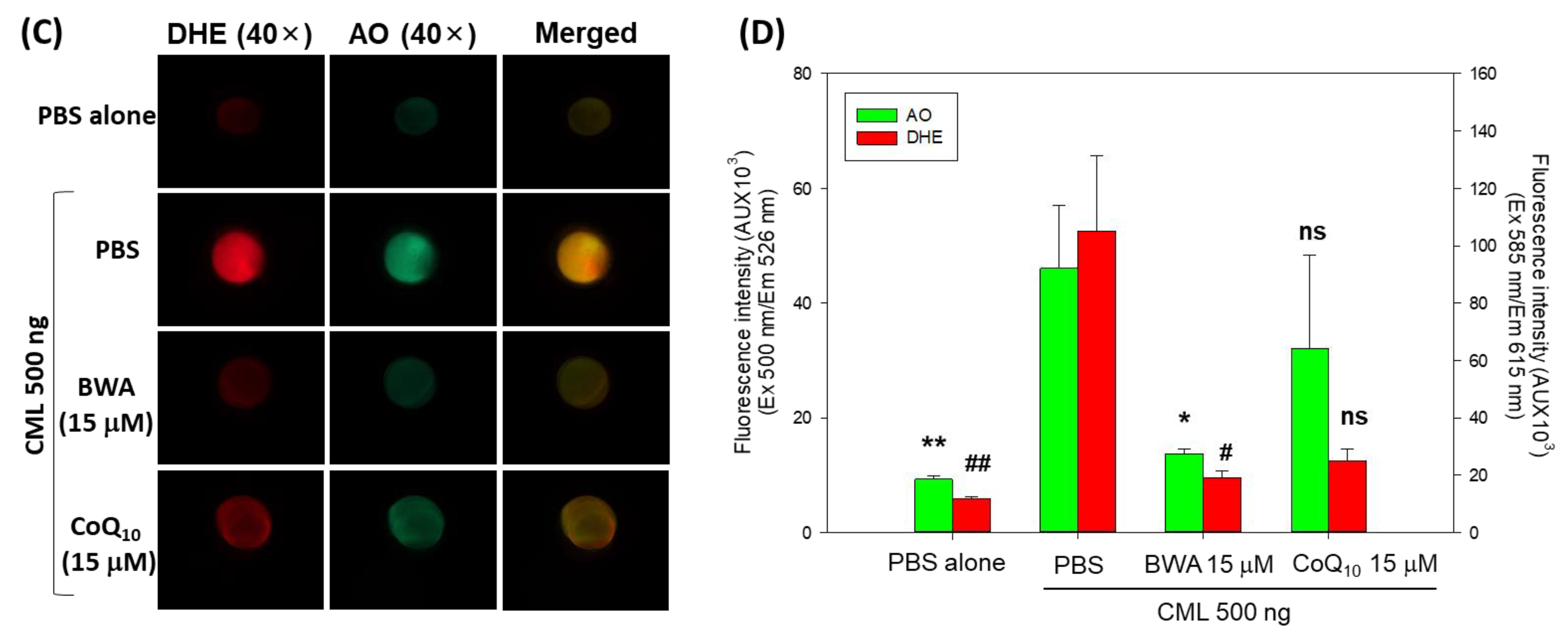
3.5. BWA Enhances the CML-Impaired Embryo Survivability
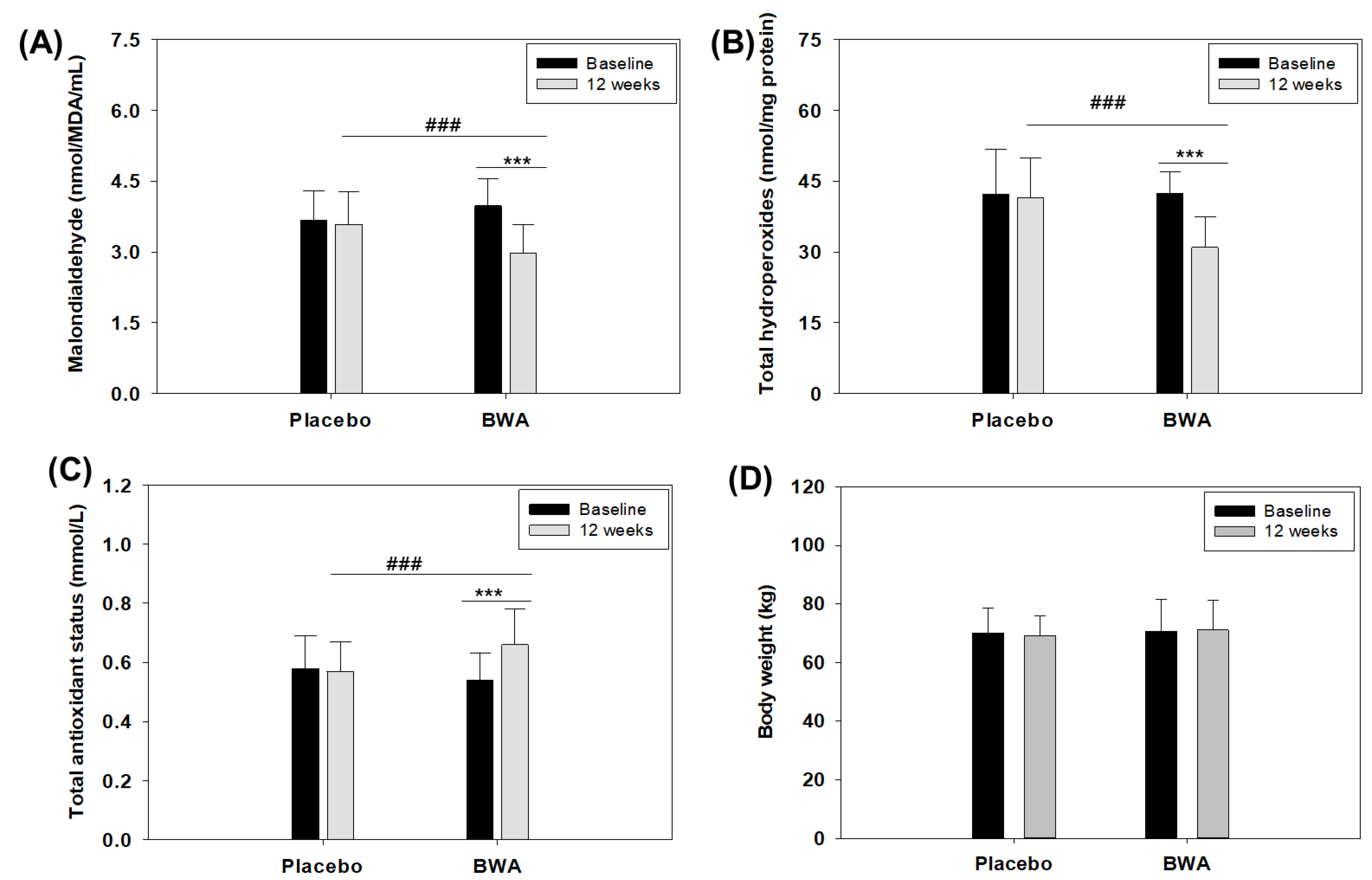
3.6. Consumption of BWA Improved Oxidative Plasma Variables in Human Subjects
3.7. BWA Has No Adverse Effect on Anthropometric Profile, Liver and Kidney Biomarkers, and Lipid Profile

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Baseline (Week 0) | Week 12 |
|---|---|---|
| Malondialdehyde (nmol/MDA/mL) | ||
| Placebo | 3.67 ± 0.63 | 3.59 ± 0.68 |
| BWA | 3.98 ± 0.58 | 2.98 ± 0.61 ***,### |
| Total hydroperoxides (nmol/mg protein) | ||
| Placebo | 42.31 ± 9.37 | 41.57 ± 8.39 |
| BWA | 42.43 ± 4.49 | 30.97 ± 6.56 ***,### |
| Total antioxidant status (mmol/L) | ||
| Placebo | 0.58 ± 0.11 | 0.57 ± 0.10 |
| BWA | 0.54 ± 0.09 | 0.66 ± 0.12 ***,### |
| Pulse (beats/min) | ||
| Placebo | 70.08 ± 3.34 | 70.25 ± 1.07 |
| BWA | 71.36 ± 3.09 | 70.56 ± 1.47 |
| Diastolic blood pressure (mmHg) | ||
| Placebo | 79.20 ± 4.93 | 78.33 ± 3.61 |
| BWA | 78.80 ± 4.40 | 78.40 ± 3.74 |
| Systolic blood pressure (mmHg) | ||
| Placebo | 123.40 ± 11.06 | 125.20 ± 5.41 |
| BWA | 125.60 ± 11.21 | 125.00 ± 6.12 |
| Hemoglobin (g/L) | ||
| Placebo | 12.91 ± 1.31 | 12.98 ± 1.15 |
| BWA | 12.77 ± 0.83 | 12.80 ± 0.85 |
| Hematocrit (%) | ||
| Placebo | 39.31 ± 2.64 | 39.46 ± 2.06 |
| BWA | 38.98 ± 2.06 | 39.50 ± 1.81 |
| Red blood cells count (×1012/L) | ||
| Placebo | 4.23 ± 0.38 | 4.17 ± 0.20 |
| BWA | 4.22 ± 0.33 | 4.26 ± 0.28 |
| White blood cells count (×109/L) | ||
| Placebo | 6.00 ± 1.43 | 6.09 ± 1.15 |
| BWA | 6.38 ± 1.34 | 6.15 ± 0.98 |
| Platelets count (×109/L) | ||
| Placebo | 225.70 ± 39.33 | 227.41 ± 35.56 |
| BWA | 213.84 ± 42.42 | 215.36 ± 35.78 |
| Aspartate aminotransferase (U/L) | ||
| Placebo | 24.80 ± 6.26 | 24.75 ± 3.82 |
| BWA | 26.28 ± 5.42 | 25.60 ± 4.23 |
| Alanine aminotransferase (U/L) | ||
| Placebo | 16.88 ± 5.59 | 17.58 ± 4.57 |
| BWA | 17.88 ± 7.13 | 17.32 ± 4.23 |
| Glucose (mmol/L) | ||
| Placebo | 4.36 ± 0.90 | 4.39 ± 0.72 |
| BWA | 4.92 ± 1.05 | 4.61 ± 0.93 |
| Creatinine (μmol/L) | ||
| Placebo | 76.08 ± 16.91 | 77.28 ± 12.10 |
| BWA | 76.12 ± 20.29 | 77.67 ± 14.33 |
| Total cholesterol (mmol/L) | ||
| Placebo | 6.31 ± 1.37 | 6.11 ± 1.10 |
| BWA | 6.34 ± 1.12 | 6.08 ± 0.78 |
| Triglycerides (mmol/L) | ||
| Placebo | 1.95 ± 0.98 | 1.71 ± 0.53 |
| BWA | 1.78 ± 0.74 | 1.68 ± 0.49 |
3.8. Limitations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Saleh, E.A.M.; Al-Dolaimy, F.; Baymakov, S.; Ullah, M.I.; Khlewee, I.H.; Bisht, Y.S.; Alsaalamy, A.H. Oxidative stress affects the beginning of the growth of cancer cells through a variety of routes. Pathol. Res. Prac. 2023, 249, 154664. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M. Free radicals in diseases processes: A compilation of cause and consequence. Free Radic. Res. Commun. 1993, 19, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: From basic research to clinical application. Am. J. Med. 1991, 91, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, Y.; Mishima, M. Redox-based therapeutics for lung diseases. Antioxid. Redox Signal. 2008, 10, 701–704. [Google Scholar] [CrossRef] [PubMed]
- MacNee, W. Oxidative stress and lung inflammation in airways disease. Eur. J. Pharmacol. 2001, 429, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Rye, K.A. High-density lipoproteins, inflammation and oxidative stress. Clin. Sci. 2009, 116, 87–98. [Google Scholar] [CrossRef]
- Brites, F.; Martin, M.; Guillas, I.; Kontush, A. Antioxidative activity of high-density lipoprotein (HDL): Mechanistic insights into potential clinical benefit. BBA Clin. 2017, 8, 66–77. [Google Scholar] [CrossRef]
- Ito, F.; Ito, T. High-density lipoprotein (HDL) triglyceride and oxidized HDL: New lipid biomarkers of lipoprotein-related atherosclerotic cardiovascular disease. Antioxidants 2020, 9, 362. [Google Scholar] [CrossRef]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Nam, K.; Huang, X.; Ahn, D.U. Plant- and animal-based antioxidants’ structure, efficacy, mechanisms, and applications: A review. Antioxidants 2022, 11, 1025. [Google Scholar] [CrossRef] [PubMed]
- Carbajal, D.; Molina, V.; Valdés, S.; Arruzazabala, L.; Más, R. Anti-ulcer activity of higher primary alcohols of beeswax. J. Pharm. Pharmacol. 1995, 47, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Safety Evaluations of Specific Food Additives (Other Than Flavouring Agents). Available online: https://www.inchem.org/documents/jecfa/jecmono/v56je02.pdf (accessed on 16 May 2023).
- Cho, K.-H.; Baek, S.-H.; Nam, H.-S.; Bahuguna, A. Enhancement of antioxidant and anti-glycation properties of Beeswax alcohol in reconstituted high-density lipoprotein: Safeguarding against carboxymethyllysine toxicity in Zebrafish. Antioxidants 2023, 12, 2116. [Google Scholar] [CrossRef] [PubMed]
- Herbert, P.N.; Assmann, G.; Gotto, A.M., Jr.; Fredrickson, D.S. Familial lipoprotein deficiency: Alpha beta lipoproteinemia, hypobetalipoproteinemia, and Tangier disease. In The Metabolic Basis of Inherited Disease; Stanbury, J.B., Wyngaarden, J.B., Fredrickson, D.S., Goldstein, J.L., Brown, M.S., Eds.; McGraw-Hill: New York, NY, USA, 1982; pp. 589–651. [Google Scholar]
- Brunham, L.R.; Kruit, J.K.; Iqbal, J.; Fievet, C.; Timmins, J.M.; Pape, T.D.; Coburn, B.A.; Bissada, N.; Staels, B.; Groen, A.K.; et al. Intestinal ABCA1 directly contributes to HDL biogenesis in vivo. J. Clin. Investig. 2006, 116, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Timmins, J.M.; Lee, J.Y.; Boudyguina, E.; Kluckman, K.D.; Brunham, L.R.; Mulya, A.; Gebre, A.K.; Coutinho, J.M.; Colvin, P.L.; Smith, T.L.; et al. Targeted inactivation of hepatic Abca1 causes profound hypoalphalipoproteinemia and kidney hypercatabolism of apoA-I. J. Clin. Investig. 2005, 115, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Molina, V.; Mas, R.; Carbajal, D. D-002 (beeswax alcohols): Concurrent joint health benefits and gastroprotection. Indian J. Pharm. Sci. 2015, 77, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pérez, Y.; Oyárzabal, A.; Mas, R.; Molina, V.; Jiménez, S. Protective effect of D-002, a mixture of beeswax alcohols, against indomethacin-induced gastric ulcers and mechanism of action. J. Nat. Med. 2013, 67, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.N.; Castaño, S.M.; Ferreiro, R.M.; Clara, M.V.; Hernansez, N.M. Effects of oral administration of D-002 (Beeswax alcohols) on histological and functional outcomes in a Rat model of antigen-induced arthritis: Preliminary Study. Int. J. Pharmacol. Phytochem. Ethnomed. 2016, 5, 60–68. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Cho, K.-H.; Nam, H.-S.; Baek, S.-H.; Kang, D.-J.; Na, H.; Komatsu, T.; Uehara, Y. Beneficial effect of Cuban policosanol on blood pressure and serum lipoproteins accompanied with lowered glycated hemoglobin and enhanced high-density lipoprotein functionalities in a randomized, placebo-controlled, and double-blinded trial with healthy Japanese. Int. J. Mol. Sci. 2023, 24, 5185. [Google Scholar]
- Havel, R.J.; Eder, H.A.; Bragdon, J.H. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Investig. 1955, 34, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H.; Kim, J.-R.; Lee, I.-C.; Kwon, H.-J. Native high-density lipoproteins (HDL) with higher paraoxonase exert a potent antiviral effect against SARS-CoV-2 (COVID-19), while glycated HDL lost the antiviral activity. Antioxidants 2021, 10, 209. [Google Scholar] [CrossRef] [PubMed]
- Blatter Garin, M.C.; Moren, X.; James, R.W. Paraoxonase-1 and serum concentrations of HDL-cholesterol and apoA-I. J. Lipid Res. 2006, 47, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H.; Nam, H.-S.; Kang, D.-J.; Park, M.-H.; Kim, J.-H. Long-term alcohol consumption caused a significant decrease in serum high-density lipoprotein (HDL)-cholesterol and apolipoprotein A-I with the atherogenic changes of HDL in middle-aged Korean women. Int. J. Mol. Sci. 2022, 23, 8623. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Kim, S.-H.; Yadav, D.; Kim, S.-J.; Kim, J.-R.; Cho, K. High consumption of iron exacerbates hyperlipidemia, atherosclerosis, and female sterility in zebrafish via acceleration of glycation and degradation of serum lipoproteins. Nutrients 2017, 9, 690. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Dahm, R. Zebrafish: A Practical Approach, 1st ed.; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of oxidative stress in biological samples using the thiobarbituric acid reactive substances assay. J. Vis. Exp. 2020, 159, e61122. [Google Scholar]
- Nourooz-Zadeh, J. Ferrous ion oxidation in presence of xylenol orange for detection of lipid hydroperoxides in plasma. Methods Enzymol. 1999, 300, 58–62. [Google Scholar]
- Sengupta, S.; Nandi, I.; Bhattacharyya, D.K.; Ghosh, M. Anti-oxidant and anti-bacterial properties of 1-octacosanol isolated from rice bran wax. J. Plant Biochem. Physiol. 2018, 6, 1. [Google Scholar]
- Harrabi, S.; Ferchichi, A.; Bacheli, A.; Fellah, H. Policosanol composition, antioxidant and anti-arthritic activities of milk thistle (Silybium marianum L.) oil at different seed maturity stages. Lipids Health Dis. 2018, 16, 82. [Google Scholar] [CrossRef]
- Guerra, Y.P.; Yera, Á.O.; Despaigne, S.J.; Cuevas, C.V.M.; Ferreiro, R.M. Scavenger effect of D-002 on hydroxyl radicals in the gastric mucosa. Rev. Cuba. Farm. 2012, 46, 87–96. [Google Scholar]
- Shokri, Y.; Variji, A.; Nosrati, M.; Khonakdar-Tarsi, A.; Kianmehr, A.; Kashi, Z.; Bahar, A.; Bagheri, A.; Mahrooz, A. Importance of paraoxonase 1 (PON1) as an antioxidant and antiatherogenic enzyme in the cardiovascular complications of type 2 diabetes: Genotypic and phenotypic evaluation. Diabetes Res. Clin. Pract. 2020, 161, 108067. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H. The Current status of research on high-density lipoproteins (HDL): A paradigm shift from HDL quantity to HDL quality and HDL functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef] [PubMed]
- Zamora, Z.; Molina, V.; Mas, R.; Ravelo, Y.; Perez, Y.; Oyarzabal, A.; Jiménez, S. D-002 treatment attenuates esophagitis in a model of chronic gastroesophageal reflux in rats. IOSR J. Pharm. 2015, 5, 36–40. [Google Scholar]
- Pentikainen, M.O.; Oorni, K.; Ala-Korpela, M.; Kovanen, P.T. Modified LDL-trigger of atherosclerosis and inflammation in the arterial intima. J. Intern. Med. 2000, 247, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M. Review of human studies on oxidative damage and antioxidant protection related to cardiovascular diseases. Free Radic. Res. 2000, 33, 85–97. [Google Scholar]
- Aviram, M.; Rosenblat, M. Paraoxonases 1, 2, and 3, oxidative stress, and macrophage foam cell formation during atherosclerosis development. Free Radic. Biol. Med. 2004, 37, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Molina, V.; Valdés, S.; Carbajal, D.; Arruzazabala, L.; Menéndez, R.; Mas, R. Antioxidant effects of D-002 on gastric mucosa of rats with injury induced experimentally. J. Med. Food 2001, 4, 79–84. [Google Scholar] [CrossRef]
- Menéndez, R.; Más, R.; Amor, A.M.; Pérez, Y.; González, R.M.; Fernández, J.; Molina, V.; Jiménez, S. Antioxidant effects of D002 on the in vitro susceptibility of whole plasma in healthy volunteers. Arch. Med. Res. 2001, 32, 436–441. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Martinez, I.; Sourlas, A.; Bouza, K.V.; Campos, F.N.; Torres, V.; Montan, P.D.; Guzman, E. High-density lipoprotein (HDL) functionality and its relevance to atherosclerotic cardiovascular disease. Drugs Context 2018, 7, 212525. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-E.; Nam, H.-S.; Kang, D.-J.; Na, H.-J. Anti-inflammatory activity of CIGB-258 against acute toxicity of carboxymethyllysine in paralyzed Zebrafish via enhancement of high-density lipoproteins stability and functionality. Int. J. Mol. Sci. 2022, 23, 10130. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Martínez, P.; Fernández-Martínez, J.; Ramírez-Casas, Y.; Guerra-Librero, A.; Rodríguez-Santana, C.; Escames, G.; Acuña-Castroviejo, D. The Zebrafish, an outstanding model for biomedical research in the field of melatonin and human diseases. Int. J. Mol. Sci. 2022, 23, 7438. [Google Scholar] [CrossRef] [PubMed]
- Ruchika; Sharma, A.; Saneja, A. Zebrafish as a powerful alternative model organism for preclinical investigation of nanomedicines. Drug Discov. Today 2022, 27, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Hsu, Y.W.; Pan, T.M. Suppression of dimerumic acid on hepatic fibrosis caused from carboxymethyl-lysine (CML) by attenuating oxidative stress depends on Nrf2 activation in hepatic stellate cells (HSCs). Food Chem. Toxicol. 2013, 62, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, G.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Madhusoodanan, U.K.; Sharanabasappa, M.; Ghosh, S.; Jacob, J. Measurement of plasma hydroperoxide concentration by FOX-1 assay in conjunction with triphenylphosphine. Clin. Chim. Acta 2003, 337, 147–152. [Google Scholar] [CrossRef]
- Santas, J.; Guardiola, F.; Rafecas, M.; Bou, R. Determination of total plasma hydroperoxides using a diphenyl-1-pyrenylphosphine fluorescent probe. Anal. Biochem. 2013, 434, 172–177. [Google Scholar] [CrossRef]
- López, E.; Illnai, J.; Molina, V.; Oyarzábal, A.; Fernández, L.; Pérez, Y.; Mas, R.; Mesa, M.; Fernandez, J.; Mendoza, S. Effects of D-002 (beeswax alcohols) on lipid peroxidation in middle-aged and older subjects. Lat. Am. J. Pharm. 2008, 27, 695–703. [Google Scholar]
- Illnait, J.; Rodriguez, I.; Molina, V.; Mendoza, S.; Mas, R.; Fernandez, L.; Oyarzabal, A.; Perez, Y.; Mesa, M.; Fernández, J. Effects of D-002 (beeswax alcohols) on gastrointestinal symptoms and oxidative markers in middle-aged and older subjects. Lat. Am. J. Pharm. 2013, 32, 166–174. [Google Scholar]
- Carbajal, D.; Molina, V.; Noa, M.; Valdes, S.; Arruzazabala, M.L.; Aguiar, M.; Mas, R. Effects of D-002 on gastric mucus composition in ethanol induced ulcer. Pharmacol. Res. 2000, 42, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Carbajal, D.; Molina, V.; Valdés, S.; Arruzazabala, M.L.; Rodeiro, I.; Mas, R.; Magraner, J. Possible cytoprotective mechanism in rats of D-002 an anti-ulcerogenic product isolated from beeswax. J. Pharm. Pharmacol. 1996, 48, 858–860. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Mackness, M.I.; Durrington, P.N.; Arrol, S.; Evans, A.E.; McMaster, D.; Ferrières, J.; Ruidavets, J.B.; Williams, N.R.; Howard, A.N. Paraoxonase activity in two healthy populations with differing rates of coronary heart disease. Eur. J. Clin. Investig. 2000, 30, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.; Illnait, J.; Molina, V.; Oyarzabal, A.; Fernandez, L.; Mesa, M.; Más, R.; Mendoza, S.; Gámez, R.; Jiménez, S. Comparison of the antioxidant effects of D-002 (beeswax alcohols) and grape seed extract (GSE) on plasma oxidative variables in healthy subjects. Lat. Am. J. Pharm. 2010, 29, 255–262. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.-H.; Baek, S.-H.; Nam, H.-S.; Bahuguna, A.; López-González, L.E.; Rodríguez-Cortina, I.; Illnait-Ferrer, J.; Fernández-Travieso, J.C.; Molina-Cuevas, V.; Pérez-Guerra, Y.; et al. Beeswax Alcohol Prevents Low-Density Lipoprotein Oxidation and Demonstrates Antioxidant Activities in Zebrafish Embryos and Human Subjects: A Clinical Study. Curr. Issues Mol. Biol. 2024, 46, 409-429. https://doi.org/10.3390/cimb46010026
Cho K-H, Baek S-H, Nam H-S, Bahuguna A, López-González LE, Rodríguez-Cortina I, Illnait-Ferrer J, Fernández-Travieso JC, Molina-Cuevas V, Pérez-Guerra Y, et al. Beeswax Alcohol Prevents Low-Density Lipoprotein Oxidation and Demonstrates Antioxidant Activities in Zebrafish Embryos and Human Subjects: A Clinical Study. Current Issues in Molecular Biology. 2024; 46(1):409-429. https://doi.org/10.3390/cimb46010026
Chicago/Turabian StyleCho, Kyung-Hyun, Seung-Hee Baek, Hyo-Seon Nam, Ashutosh Bahuguna, Luis Ernesto López-González, Iván Rodríguez-Cortina, José Illnait-Ferrer, Julio César Fernández-Travieso, Vivian Molina-Cuevas, Yohani Pérez-Guerra, and et al. 2024. "Beeswax Alcohol Prevents Low-Density Lipoprotein Oxidation and Demonstrates Antioxidant Activities in Zebrafish Embryos and Human Subjects: A Clinical Study" Current Issues in Molecular Biology 46, no. 1: 409-429. https://doi.org/10.3390/cimb46010026
APA StyleCho, K.-H., Baek, S.-H., Nam, H.-S., Bahuguna, A., López-González, L. E., Rodríguez-Cortina, I., Illnait-Ferrer, J., Fernández-Travieso, J. C., Molina-Cuevas, V., Pérez-Guerra, Y., Oyarzabal Yera, A., & Mendoza-Castaño, S. (2024). Beeswax Alcohol Prevents Low-Density Lipoprotein Oxidation and Demonstrates Antioxidant Activities in Zebrafish Embryos and Human Subjects: A Clinical Study. Current Issues in Molecular Biology, 46(1), 409-429. https://doi.org/10.3390/cimb46010026