Abstract
CYCLOIDEA (CYC)-like genes belong to the TCP transcription factor family and play important roles associated with flower development. The CYC-like genes in the CYC1, CYC2, and CYC3 clades resulted from gene duplication events. The CYC2 clade includes the largest number of members that are crucial regulators of floral symmetry. To date, studies on CYC-like genes have mainly focused on plants with actinomorphic and zygomorphic flowers, including Fabaceae, Asteraceae, Scrophulariaceae, and Gesneriaceae species and the effects of CYC-like gene duplication events and diverse spatiotemporal expression patterns on flower development. The CYC-like genes generally affect petal morphological characteristics and stamen development, as well as stem and leaf growth, flower differentiation and development, and branching in most angiosperms. As the relevant research scope has expanded, studies have increasingly focused on the molecular mechanisms regulating CYC-like genes with different functions related to flower development and the phylogenetic relationships among these genes. We summarize the status of research on the CYC-like genes in angiosperms, such as the limited research conducted on CYC1 and CYC3 clade members, the necessity to functionally characterize the CYC-like genes in more plant groups, the need for investigation of the regulatory elements upstream of CYC-like genes, and exploration of the phylogenetic relationships and expression of CYC-like genes with new techniques and methods. This review provides theoretical guidance and ideas for future research on CYC-like genes.
1. Introduction
Cubas et al. first proposed the concept of the TCP transcription factor family, which is named according to the first letters of TEOSINTE BRANCHED 1 (TB1) in maize (Zea mays), CYCLOIDEA (CYC) in snapdragon (Antirrhinum majus), and PROLIFERATING CELL FACTOR 1 and 2 (PCF1 and PCF2) in rice (Oryza sativa) [1,2,3,4,5]. Genes encoding proteins with the TCP domain are involved in the regulation of angiosperm growth and development [6,7,8,9]. The TCP family members contain a highly conserved TCP domain, which forms a basic helix–loop–helix (bHLH) structure associated with DNA binding and protein dimerization [10,11]. TB1 is a major regulator of stem and lateral bud growth and the male flower formation of maize, rice, wheat, and other crops [12,13,14], whereas CYC controls the floral dorsal organ characteristics in snapdragon [1,15], and both PCF1 and PCF2 bind to the promoter of PROLIFERATING CELL NUCLEAR ANTIGEN (PCNA), which is crucial for DNA replication and repair, chromatin structure maintenance, chromosome isolation, and the cell cycle in rice [3]. According to their different domains, the members of the TCP family have been divided into the following two categories: TCP-P and TCP-C [16,17,18]. Moreover, TCP-C has been subdivided into the ECE (CYC/TB1) and CINCINNATA (CIN) clades [19,20].
The CYC genes belong to the ECE clade, which is unique to angiosperms [21,22]. In addition to the TCP and R domain sequences, CYC genes encode the glutamate–cysteine–glutamic acid (ECE) motif specific to core eudicots [23,24,25]. Phylogenetic analysis has indicated that CYC genes in angiosperms experienced two major gene duplication events, which led to the formation of the CYC1, CYC2, and CYC3 clades [26,27,28]. In different evolutionary lineages, gene duplication events occurred in each branch at different time points during evolution [29,30,31,32,33,34,35], as shown in Figure 1. A more thorough analysis of the CYC2 subgroup confirmed that they are key regulatory genes for the bilateral symmetry of flowers [36,37,38].
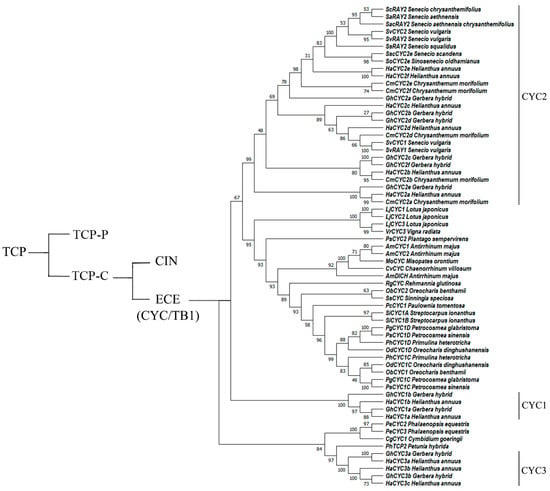
Figure 1.
Phylogenetic tree of selected CYC-like genes in angiosperms. The number beside each node is the bootstrap support value.
Angiosperm flowers can be divided into the following three types according to the presence or absence of a plane of symmetry: (1) radially symmetrical flowers (actinomorphic) with multiple planes of symmetry, (2) bilaterally symmetrical flowers (zygomorphic) with only one plane of symmetry, and (3) asymmetrical flowers with no plane of symmetry [39,40,41,42]. Studies of fossils have revealed that angiosperm flowers were originally actinomorphic and that zygomorphic flowers arose during evolution [43,44,45,46]. Wild-type snapdragon, which has typical zygomorphic flowers, is a model plant for studying floral symmetry [47]. Early investigations on the molecular mechanism regulating snapdragon flower types demonstrated that zygomorphy was determined by CYC, DICHOTOMA (DICH), RADIALIS (RAD), and DIVARICATA (DIV) genes encoding regulators of the dorsoventral characteristics of flowers [1,4]. The functionally redundant CYC and DICH genes belong to the CYC2 clade and originated from gene duplication events associated with angiosperm evolution [48,49,50]. These genes are essential for the development of zygomorphic flowers in angiosperms, especially the CYC genes [45,51,52]. The results of studies on snapdragon compelled researchers to investigate the mechanism by which CYC genes regulate floral symmetry. In addition to snapdragon, the regulatory function of CYC genes has been explored in other insect-pollinated plants with zygomorphic flowers, including Fabaceae, Gesneriaceae, Caprifoliaceae, Scrophulariaceae, and Malpighiaceae species [53,54,55,56,57]. The results of these studies suggest that CYC2 genes are the key genes for regulating the development of zygomorphic flowers.
This review summarizes the status of research and unresolved problems in Fabaceae, Asteraceae, Scrophulariaceae, Gesneriaceae, Orchidaceae, Solanaceae, and other families, while also proposing future directions for studies on CYC-like genes. As the three largest families of angiosperms, Fabaceae, Asteraceae, and Orchidaceae have been the main focus of research on CYC-like genes. In the Fabaceae species, CYC-like genes control floral symmetry, whereas the CYC-like genes in the Asteraceae species modulate changes to the capitulum morphology and regulate elongation of the showy corolla limb of the ray florets. In the Scrophulariaceae and Gesneriaceae species, CYC-like genes mainly affect the morphological characteristics of petals and stamen development. The CYC-like genes in the Orchidaceae species influence floral development or branching. The CYC-like genes in the Solanaceae species mainly affect axillary bud growth and development and stem growth, while also controlling branching, flower differentiation and development, and leaf size. These genes contribute to morphological changes to the dorsoventral floral symmetry in the Caprifoliaceae species. A systematic analysis of the functions and evolution of CYC-like genes may provide researchers and breeders with a theoretical basis for future research.
2. Progress in Research on CYC-like Genes in Fabaceae
The Fabaceae species are distributed worldwide. Because of their diversity in floral symmetry, legumes are suitable for exploring the evolution and underlying mechanism of floral symmetry [58]. Researchers have screened the Fabaceae for homologs of snapdragon CYC genes and then analyzed their functions to clarify the role of CYC-like genes in angiosperm floral development. The differences among the diverse species in terms of the CYC-like genes responsible for floral symmetry revealed a new regulatory system.
The duplication of CYC homologues gave rise to three copies of ECE clade genes in the TCP family in Lotus Japonicus [59]. In L. japonicus, both LjCYC1 and LjCYC2 mediate the development of asymmetrical inflorescences and flowers, and changes in the number of petals and in wing and keel morphology were observed in transgenic plants separately overexpressing LjCYC1 and LjCYC2 [59]. The asymmetrical expression pattern of LjCYC2 is similar to that of the snapdragon CYC gene in the developing flower primordium [59]. However, LjCYC2 is also expressed in the inflorescence primordium of L. japonicus, whereas the CYC gene is expressed only during floral primordium development in snapdragon [4].
Citerne et al. reported that the homologous genes of CYC in legumes can be divided into two major classes, ECE groups I and II, which are the result of an early duplication event [60]. ECE I can be further divided into two subclasses, IA and IB, which originated from duplication near or prior to the divergence of legumes. The LEGCYC genes in Lupinus are homologous to the regulatory gene CYC that controls the floral symmetry and paraxial floral organ characteristics of snapdragon and its related species [61]. Ree et al. suggested based on a molecular evolutionary analysis that positive selection has played a role in the evolution of the LEGCYC1B lineage, which is closely associated with floral morphological changes in Lupinus. Papilionoideae have strongly bilaterally symmetrical flowers, whereas Cadia purpurea flowers show radial symmetry associated with the expression of two CYC homologous genes (LEGCYCs) in the dorsal region of the flower (Figure 2) [58]. In addition, the expression pattern of one gene has expanded from the paraxial to the lateral and posterior regions of the corolla, which may result in reversion to evolutionarily regressive petal characters.

Figure 2.
Flowers of Cadia purpurea and Lupinus nanus [60]. (A) Flower of Cadia purpurea with actinomorphic corolla. (B) Flowers of Lupinus nanus with zygomorphic corolla. Reprinted with permission from Ref. [60]. Copyright © 2023, Oxford University Press.
Wang et al. determined that the expression of three endogenous LjCYC genes is specifically inhibited by different RNAi transgenes [62]. A chimeric RNAi transgene containing LjCYC1- and LjCYC2-specific sequences down-regulated the expression of both endogenous genes. The effect of silencing the three LjCYC genes was mainly confined to the dorsal or lateral part of the petals, implying that the genes are associated with dorsal and lateral activities during the development of zygomorphic flowers [62]. Knockdown of the three LjCYC genes may result in wild-type petals that resemble ventral petals, complete organ internal (IN) asymmetry, and the lack of dorsoventral (DV) pathway-differentiated flowers. This suggests that DV asymmetry during the development of zygomorphic flowers is controlled by LjCYC genes, whereas floral organ IN asymmetry is independently determined by other genetic factors.
The mutation of CYC2 in Lathyrus odoratus causes a change in dorsoventral petal type, resulting in a hooded (hdd) flower mutant with an epidermis and the pigmentation characteristic of a wing petal, and with a concave standard petal, the same as the lobed standard (lst1) mutant in Pisum [63]. Differences in CYC expression and activity may lead to differences in dorsal petal morphology in Fabaceae, and play a role in the negative regulation of petal edge growth in Lathyrus, mainly maintaining the flatness of the dorsal petal [64]. Interestingly, Ojeda et al. found that changes in the timing of LjCYC2 expression during pollination of Lotus by bees and birds may be responsible for changes in flower petal micromorphology and size, whereas changes in the spatial distribution of gene expression had no effect on pollination [65].
Feng et al. determined that the upstream promoter regions of GmCYC genes vary in number and type of hormone response elements in Glycine max [66]. The expression of GmCYC genes is involved in different growth and developmental stages, induced by abscisic acid, brassinosteroids, aminocyclopropane–1–carboxylic acid, salicylic acid, and methyl jasmonate signals [66]. The CYC-like genes may have undergone multiple duplications and losses in different Fabaceae lineages and formed the distinct homologous clades CYC1 and CYC2, but the CYC3 clade was most likely lost [67]. The ancestors of Papilionoideae and Caesalpinioideae probably possessed two CYC1 gene copies, but one of the copies was subsequently lost in Papilionoideae and was retained only in a few species of Caesalpinioideae [67]. The CYC2 gene was replicated more frequently in Papilionoideae than in other legumes [67]. The diversity patterns of CYC1 and CYC2 genes are not associated with floral symmetry in non-papilionoid legumes, but the replication and functional differentiation of CYC2 genes is necessary for floral symmetry in Papilionoideae [67].
The expression pattern of VrCYC3, which is homologous to L. japonicus LjCYC3 and pea PsCYC3, differs from that of VrCYC1 and VrCYC2 in the dorsal, lateral, and ventral petals in mung bean (Vigna radiata) [68]. In addition, VrCYC3, which is localized to the nucleus, can induce transcription [68]. Moreover, it can interact with VrCYC1 and VrCYC2 in yeast cells, but this interaction is weakened by the deletion of two amino acid residues in its R domain [68]. This suggests that LjCYC3/PsCYC3/VrCYC3 play a conserved role in determining the lateral petals shape, and the formation of symmetrical and asymmetrical flowers in Fabaceae.
3. Progress in Research on CYC-like Genes in Asteraceae
Asteraceae is the most highly evolved family of dicotyledonous plants with a complex inflorescence structure, termed a capitulum, that often consists of radially symmetrical disc florets and bilaterally symmetrical ray florets [69,70]. Researchers have cloned snapdragon CYC homologs in Helianthus, Gerbera, Senecio, Chrysanthemum, and other genera, which revealed the considerable abundance of these homologs in Asteraceae, many of which have diverse functions [71,72,73,74,75]. At the single-floret level, the CYC gene in the Asteraceae species uniquely regulates the elongation of the corolla limb of ray florets, which are critical for attracting pollinators [76].
3.1. CYC-like Genes of Helianthus
The radiate sunflower (Helianthus annuus) capitulum consists of bilaterally symmetrical sterile ray florets and radially symmetrical bisexual disc florets. Ten ECE clade members have been identified in the sunflower, and the spatiotemporal expression of these homologous genes varies [77]. The specific temporal expression of the different genes in diverse plant parts, including ray florets, disc florets, leaves, and roots, may maintain the complex sunflower inflorescence structure via coordinated expression.
The sunflower tubular ray flower (turf) mutant has hermaphroditic ray florets with an almost actinomorphic corolla. Fambrini et al. determined that this mutation was caused by the insertion of a TCP motif, a non-autonomous transposable element (TE) from the CYC-like gene HaCYC2c, named Transposable element of turf1 (Tetu1) [78]. The excision of Tetu1 can restore the wild-type phenotype or produce stable mutants, indicating that HaCYC2c is a key regulator of ray floret symmetry. A loss-of-function mutation to HaCYC2c can promote the transition of sterile florets to hermaphroditic florets, which reflects the importance of CYC-like genes for the inhibition of stamen development.
HaCYC2c was mutated in two independent tubular-rayed (tub) mutants, which apparently involved TE insertions, resulting in little or no expression and the formation of radially symmetrical ray florets, which are usually bilaterally symmetrical [79]. If HaCYC2c was inserted into the offspring, ray florets were more likely to replace disc florets at the center of the capitulum, whereas if HaCYC2c expression was inhibited, bilaterally symmetrical ray florets did not develop, and the capitulum comprised only disc florets (Figure 3).

Figure 3.
Capitulum and florets of a wild-type sunflower plant and two mutants [79]. (A,B) Wild-type sunflower with disc and ray florets. (C,D) dbl mutant with disc and ray florets. (E,F) tub mutant with disc and ray florets. The florets from the disc center to the peripheral florets are arranged from left to right. Reprinted with permission from Ref. [79]. Copyright © 2023 Chapman et al.
Thus the turf and tub mutants are characterized by a transition from bilaterally symmetrical to radially symmetrical ray florets because of the insertion of TEs in HaCYC2c [80]. In the dbl or Chrysanthemoides (Chry) mutants, the insertion of HaCYC2c upstream of the coding region results in the ectopic expression of this gene and a transition from radially symmetrical disc florets to bilaterally symmetrical disc florets. The loss-of-function mutation to the CYC-like gene in sunflower turf mutants reportedly results in hermaphroditic tubular-like florets, which replace the normal sterile ray florets and the formation of a capitulum type that is not normally found in sunflower [81].
HaCYC2c was overexpressed after an insertion into the HaNDUA2 promoter region to generate the sunflower long petal mutant (lpm) in which the abnormal elongation of the disc floret corolla and stamen abortion at an early stage of floral organ development was observed [82]. Furthermore, the floret symmetry changed from radial symmetry to bilateral symmetry, thus transforming the disc florets into ray florets. The overexpression of HaCYC2c and its control of HaNDUA2 through transcriptional recognition may be an important regulatory node for floret type and functional differentiation in Helianthus, which was associated with maintaining the balance between the pollinator recruitment ability and the fertility of disc florets [82].
HdCYC2c and HxmCYC2cB belong to the CYC2 subclade in Helianthus [83]. HdCYC2c was differentially expressed in the different floret types of Helianthus decapetalus, with the expression level higher in the ray floret corolla than in the disc floret corolla [83]. In Helianthus × multiflorus, the insertion of TEs in HxmCYC2cB promoted the ectopic expression of HxmCYC2cB throughout the inflorescence, leading to the observed loss of actinomorphic florets and the production of ray florets [83]. Removal of a TE (CTEHM1) and epigenetic regulation of HmCYC2c expression resulted in two capitulum types of Helianthus × multiflorus, Meteor 1 and Meteor 2 [37]. The expression of HmCYC2c in the disc floret of Meteor 2 was significantly higher than that of Meteor 1. The CTEHM1 in HmCYC2c was truncated in Meteor 1, which showed the typical tubular corolla of Helianthus, whereas the remaining presence of CTEHM1 in HmCYC2c of Meteor 2 caused the largest corolla of disc florets to display the characteristics of a ray floret, resulting in an entirely radiate capitula not normally found in Helianthus [37].
Fambrini et al. isolated three CYC2 subclade genes (HrCYC2c, HrCYC2d, and HrCYC2e) associated with the identity of the Helianthus ray–floret (Hr), among which HrCYC2c played an important role in the initiation of the ray floret primordium [38]. The capitula of HrCYC2c-mutant homozygous dominant plants (HrCYC2c/HrCYC2c) and heterozygous dominant plants (HrCYC2c/HrCYC2c-m) initiated ray florets, whereas the recessive homozygous plants (HrCYC2c-m/HrCYC2c-m) did not develop ray florets [38].
3.2. CYC-like Genes of Gerbera Hybrida
The CYC-like homolog GhCYC2a is involved in the differentiation of Gerbera hybrida floret types, and its expression exhibits a gradient along the radial axis of the capitulum [71]. Specifically, GhCYC2a is expressed in the peripheral, bilaterally symmetrical ray florets, but not in the centermost disc florets, which are almost radially symmetrical and have more deeply incised corolla lobes. The overexpression of GhCYC2a results in disc florets acquiring a morphology similar to that of ray florets, whereas the inhibition of GhCYC2a expression leads to the development of limbs that are shorter than those of wild-type ray florets (Figure 4). This provided the first molecular evidence that a CYC-like TCP TF is involved in the definition of the capitulum of the Asteraceae species.
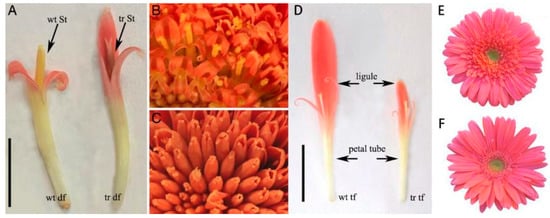
Figure 4.
G. hybrida with GhCYC2a overexpression (A–C) and inhibition (D–F) [71]. (A) Disc florets (df) of wild-type (wt) Gerbera hybrida and transgenic (tr) G. hybrida with obvious phenotypic differences. St, stamen. (B) Pollen presentation on the style of the disc florets of wild-type G. hybrida. (C) Disc florets of transgenic G. hybrida lacking functional stamens. (D) Transitional floret (tf) of wild-type and transgenic G. hybrida. (E) Capitulum of wild-type G. hybrida. (F) Capitulum of genetically modified G. hybrida. Reprinted with permission from Ref. [71]. Copyright © 2023 by The National Academy of Sciences of the USA.
GhCYC2a collaborates with other CYC-like genes to participate in floret differentiation and ultimately determine the complex capitulum structure of G. hybrida [84]. GhCYC2a is specifically expressed in ray florets at an early developmental stage and is only activated in tubular flowers at an advanced developmental stage. GhCYC2b in G. hybrida and HaCYC2d and HaCYC2c in sunflowers belong to the CYC2 clade considered to be a strong candidate as regulators of ray–floret identity [74]. When GhCYC2b expression was inhibited in G. hybrida, the third type of transitional florets were shorter and the ray–floret corolla was five- or eight-lobed and radially symmetrical, which was in accordance with the findings of Broholm [84].
There is a substantial overlap in expression patterns among the CYC2 subclade genes (i.e., GhCYC2a, GhCYC2b, GhCYC2c, and GhCYC2d) in G. hybrida [74]. At the single-floret level, their expression domains in the corolla shifted spatially from the currently known dorsal pattern in bilaterally symmetrical flower species, which may have evolved after the origin of Asteraceae [74]. GhCYC2a, GhCYC2b, and GhCYC2c mediate the positioning in the proximal and distal axes of the capitulum, leading to ray floret differentiation, and also regulate ray–floret corolla growth by affecting cell proliferation until the corolla assumes its final size and shape [74]. In contrast, the expression of GhCYC2d may increase the floret initiation rate during the expansion of the capitulum, while the ectopic expression of GhCYC2d increases the floret density in the capitulum [74]. The upstream regulators of GhCYC2b (i.e., GhCIN1 and GhCIN2) are CINCINNATA-like homologous TCP proteins with unknown expression domains and functions, but are known to delay the development of marginal ray–floret primordia during early ontogeny [85]. In developing ray florets, the class E MADS-box TF GRCD5 activated GhCYC2b expression, whereas the class C MADS-box TF GAGA1 (upstream of GhCYC2b) contributed to stamen development.
3.3. CYC-like Genes of Senecio
Natural polymorphism of the capitulum in the Senecio species is due to the transfer of a set of regulatory genes containing the RAY locus from the diploid Senecio squalidus to the tetraploid Senecio vulgaris [86]. The RAY locus, which comprises a cluster of CYC-like genes expressed in the periphery of the inflorescence meristem that promote floral asymmetry and lead to increased outcrossing rates, has played a key role in the evolution of radiate capitulum types. The CYC2-like gene RAY3 is initially uniformly expressed in ray florets during capitulum development, but at advanced stages is expressed only in the ventral corolla lobes of ray florets, resulting in the elongation of the ventral corolla limb in S. vulgaris [72]. The diversification of CYC-like genes has led to novel interactions, with SvDIV1B inhibiting RAY3, but potentially activating RAY2. The expression of SvRAY1 may induce lateral cell division during the development of the S. vulgaris ray floret and, as a result, the morphology and arrangement of the ray floret cells change to some extent, thereby affecting the ray floret width [87]. The ray florets of SvRAY1-overexpressing plants were shorter and significantly broader than the wild-type ray florets.
3.4. CYC-like Genes of Chrysanthemum
Researchers have cloned the homologs of snapdragon CYC genes in several Chrysanthemum × morifolium cultivars and analyzed their expression and function [88,89,90,91,92]. Huang et al. identified six CYC2 subclade CmCYC2 genes (i.e., CmCYC2a, CmCYC2b, CmCYC2c, CmCYC2d, CmCYC2e, and CmCYC2f) in the C. × morifolium cultivar ‘Maoxiangyu’, wherein they mainly regulate the development of ray florets [88]. Compared with other CYC homologs, CmCYC2s in chrysanthemum may be similarly expressed or there may be distinct differences in expression patterns. The overexpression of CmCYC2d in wild-type Arabidopsis thaliana and the tcp1 mutant showed that the vegetative growth of the transgenic lines was inhibited, the flowering period was delayed, and the petal size and arrangement were changed, making the originally radially symmetrical petals appear bilaterally symmetrical [88]. Furthermore the CmCYC2 proteins may form homodimers during flower organogenesis and participate in the regulation of ray and disc floret morphogenesis.
Chen et al. observed that the spatial expression patterns of six Asteraceae CYC2-like members are conserved throughout the family, and all of them influence capitulum development [93]. Both CYC2c and CYC2g are important for ray floret formation in Chrysanthemum lavandulifolium, whereas CYC2d inhibited the development of the dorsal corolla lobes and ray–floret stamens. The class A MADS-box genes interacted with CYC2-like genes potentially involved in processes associated with the formation of reproductive organs and the ray–floret corolla, especially corolla differentiation of the disc and ray florets in chrysanthemum [94]. The class B MADS-box gene CDM19 may positively regulate the expression of the CYC2-like genes CmCYC2c and CmCYC2d, thereby modifying the floret symmetry in chrysanthemum [95].
Yang et al. cloned the C. × morifolium gene CmTCP7, which may be involved in the formation of floral buds as well as promote the growth of the corolla of the ray floret and participate in the formation of bilaterally symmetrical ray florets [96]. The CYC-like gene CmCYC2 and WUS-like gene CmWUS, which were highly expressed in floral buds at the time of floral organ differentiation and in reproductive organs at advanced stages of development, coordinately regulate the development of C. × morifolium reproductive organs [89]. Furthermore, CmCYC2 was highly expressed in the corolla of ray florets, which may promote ray floret growth and contribute to the formation of bilaterally symmetrical ray florets.
The expression of CYC2b, CYC2d, CYC2e, and CYC2f was differentially expressed in different types of Chrysanthemum vestitum ray florets, which confirmed the influence of CYC-like genes on floral morphology [90]. Yuan et al. observed that the ectopic expression of CmCYC2 in the Arabidopsis tcp1 mutant altered flower symmetry and flowering time, and the CmCYC2 TF may interact with or bind to the CmCYC2 promoter to regulate floral symmetry development in Chrysanthemum [91]. Liu et al. cloned the CYC2-like gene Cyc2CL from C. × morifolium ‘Pink Carpet’ and revealed for the first time the variable shear pattern of a CYC2-like gene in chrysanthemum [92]. The transcription of Cyc2CL resulted in two mature mRNA sequences (CyC2CL-1 and CyC2CL-2). Both transcripts were present at high levels in ray florets, but at very low levels in disc florets and inhibited the development of petals and stamens in A. thaliana.
The morphogenesis of the marginal florets in Ajania is interrupted shortly after the formation of the floral primordia, possibly because of the lack of expression of the CYC2-like gene ClCYC2g [97]. The decreased expression of ClCYC2g in C. lavandulifolium results in the gradual transformation of ray florets into disc florets (Figure 5). This transition may be associated with changes in pollination strategies under selective pressure. Zhang et al. identified four ClCYC2-like genes (i.e., ClCYC2c, ClCYC2d, ClCYC2e, and ClCYC2f), for which expression levels were significantly higher in ray florets than in disc florets of C. lavandulifolium [98].

Figure 5.
Knockdown of ClCYC2g expression adversely affects the formation of symmetrical marginal florets in the radiate capitulum in chrysanthemum [97]. (A,B) Morphology of the capitulum of wild-type (WT) and transgenic C. lavandulifolium. (C) Ray and disc florets of wild-type C. lavandulifolium. (D) Ray-shaped florets with gradually increasing mutations. Orange and blue arrows indicate abnormal petals and stamens, respectively. (E,F) Expanded marginal florets of the transgenic lines. White and blue arrows indicate the bilabiate corolla with a deeply dentate limb apex and stamens, respectively. Reprinted with permission from Ref. [97]. Copyright © 2023 Society for Experimental Biology and John Wiley & Sons Ltd. (Hoboken, American).
3.5. CYC-like Genes of Other Asteraceae Groups
Bello et al. recovered eight major gene lineages in the highly derived genus Anacyclus (tribe Anthemideae) through phylogenetic reconstruction, comprising two CYC1 genes, four CYC2 genes, and two CYC3 genes [21]. In Anacyclus, three AcCYC2 genes are highly expressed in ray florets, and the expression patterns of four AcCYC2 genes overlap in multiple organs, including the limb of ray florets, anthers, and ovule throughout development. Gene duplication events, as well as the subsequent subfunctionalization and neofunctionalization of SEPALLATA-like genes and CYC-like genes in Asteraceae, have been shown to be conducive to the identification of the floral meristem and the formation of key traits for floral differentiation in this large family [99]. Sun et al. identified five CYC2-like genes in several Gaillardia cultivars with different ray floret types [73]. Analyses of RNA re-sequencing results, quantitative real-time PCR (qRT-PCR) data, and the effects of gene silencing suggested that CYC2c is the main genetic factor affecting the formation of ray florets in Gaillardia.
4. Progress in Research on CYC-like Genes in Lamiales
Gene duplication, gene family retention, and tissue-specific expression of CYC-like genes are believed to have affected the evolution of corolla symmetry in Lamiales [100,101]. The CYC-like genes were differentially expressed in the higher core clades with high expression levels in adaxial petals, which had been widely replicated in Lamiales (including Lamiaceae, Scrophulariaceae, Gesneriaceae, Oleaceae, Phrymaceae, and many other families) [102,103]. The asymmetrical expression of CYC-like genes was not common but associated with the origin of bilaterally symmetrical corollas [104]. Changes to the cis-regulatory domain and the coding sequence of CYC-like genes may be critical for the symmetrical evolution of both sides of the corolla, with multiple selection mechanisms contributing to gene retention [100].
The expression pattern of CYC2-like genes has gradually evolved, and was widely expressed in the meristem of early-diverging Lamiales with a bilaterally symmetrical corolla, but limited in the meristem of core Lamiales and thus may be related to the origin of corolla bilateral symmetry [105,106]. The repeated loss of bilateral corolla symmetry is relatively frequent in Lamiaceae, which may be caused by different mechanisms and changes in floral symmetry-related genes, such as the loss of the CYC2 clade gene Ml-CYC2A in the genome and the contraction, expansion, or altered expression of Cc-CYC2A [107,108]. Sengupta and Hileman detected the significant enrichment of predicted autoregulatory sites in the 5′-terminal upstream noncoding region of CYC, the upstream regulator of floral zygomorphy in Lamiales. Their results suggest that the correlation between the autoregulation of CYC and the origin of zygomorphic flowers may be associated with zygomorphic flowers independently derived from eudicot lineages [40].
4.1. CYC-like Genes of Scrophulariaceae
Scrophulariaceae inflorescences are typically racemose, spicate, or cymose and often form a panicle [109]. The CYC gene associated with the regulation of floral symmetry was initially isolated from snapdragon, and its homologs in related species were subsequently cloned, including Linaria vulgaris, Mohavea confertiflora, Veronica montana, Gratiola officinalis, and Torenia fournieri [1,110,111,112,113,114]. These genes have diverse functions in Scrophulariaceae, but they primarily affect the morphological characteristics of petals and the development of stamens.
The wild-type snapdragon corolla comprises two dorsal lobes, one ventral lobe, and two lateral lobes. Snapdragon mutants have a semi-abnormal regular flower (semipeloric; CYC mutation) or an abnormal regular flower (peloric; CYC/DICH double mutation), which represent bilaterally symmetrical floral transitions into a radially symmetrical flower (Figure 6) [1,115]. In the classic DICH homozygous mutant, the ventral corolla lobes are more symmetrical than the wild-type ventral lobes and usually separate from each other because of a deep incision between the dorsal lobes [1].
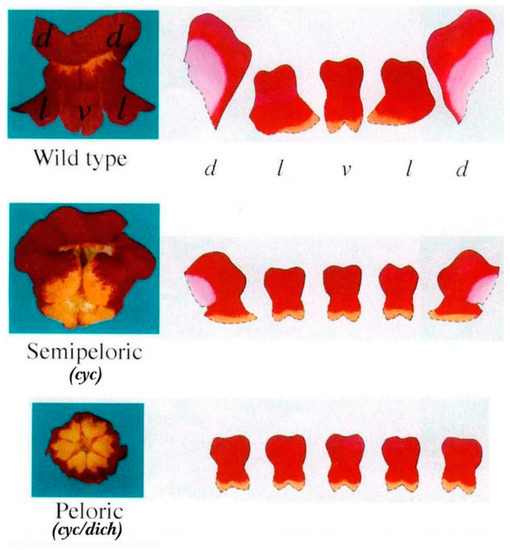
Figure 6.
Flowers of wild-type and mutant snapdragon plants [1]. Photographs of the dorsal corolla lobe (d), lateral corolla lobe (l), and ventral corolla lobe (v) of the wild-type snapdragon flower are presented. The characteristics of the different corolla lobes are shown to the right of each flower. Reprinted with permission from Ref. [1]. Copyright © 2023, Nature Publishing Group.
Corley et al. determined that AmCYC was expressed in the dorsal corolla lobe, in which AmRAD was activated [116]. This ultimately led to the inhibited expression of AmDIV in the ventral and lateral lobes, and the formation of asymmetrical snapdragon flowers. Li et al. revealed that whole-genome duplication (WGD) and tandem replication had contributed to the expansion of the CYC gene family [15]. Both (RAD) and (DIV) controlled floral symmetry downstream of CYC/DICH and interacted with DIV-RAD-INTERACTING FACTOR (DRIF) [15]. The DRIF genes, which had homologous copies similar to CYC/DICH, were also located in the WGD-derived syntenic block [15]. These results further support the view that the key genetic factor regulating the asymmetry of snapdragon flowers was the result of a WGD event.
The peloric flowers (i.e., transitional from bilateral symmetry to radial symmetry) of an L. vulgaris mutant were the result of a spontaneous epigenetic mutation to the CYC allele [110,117]. The mutant harbored a defective LCYC, which is a CYC homolog. LCYC underwent a heritable modification (i.e., extensive methylation and transcriptional silencing) that was co-isolated with the mutant phenotype [117]. The mutant phenotype may be reversed during somatic development, which is associated with the demethylation of LCYC and the restoration of gene expression.
Hileman et al. identified the M. confertiflora CYC and DICH homologs, McCYC1, McCYC2, McDICH1, and McDICH2, of which expression levels increase from the stamen to the outermost floral whorl, which may be due to the change in the expression domain of the regulatory genes in the CYC/DICH pathway [111]. Changes to the McCYC and McDICH expression patterns result in new floral morphological traits, in that the two lateral stamens are aborted and show evidence of the adaxial corolla lobes’ internal symmetry. The expression of CYC-like genes led to delayed growth or degradation of the adaxial floral organs, but it may also be associated with the loss of the adaxial floral organs [1,111].
A conservative floral symmetry gene network exists in V. montana and G. officinalis, in which CYC-like genes evolved after the gene duplication event, although the detailed genetic mechanisms of dorsal and ventral stamen abortion differ [114]. Specifically, VmCYC1, GoCYC1, and GoCYC2 are only expressed in the dorsal region of the floral meristem and in developing flowers, in which expression patterns are independent of stamen abortion patterns, whereas the expression of VmCYC2 and GoCYC3 is mainly detected in vegetative and floral tissues.
A dorsally expressed CYC-like gene and the downstream target genes RAD and DIV are absent in Plantago major but are present in Aragoa abietina [53]. This CYC-like gene is expressed in all parts of the flower in A. abietina, including the dorsal, ventral, and lateral regions, similar to the expression of its homolog in the related species Veronica serpyllifolia. The duplication of CYC-like genes led to the evolution of radially symmetrical A. abietina/P. major flowers, and further disintegration of the symmetrical flower-related gene pathway led to the wind-pollination syndrome of P. major [66]. This model emphasizes the potential importance of gene loss in the evolution of important ecological traits.
Su et al. detected recent replication events of a CYC-like gene in T. fournieri, and functional analysis of two genes that show dorsal-specific expression, TfCYC1 and TfCYC2, suggested the existence of a regulatory module integrating the dorsoventral pattern and asymmetric corolla pigmentation [112]. The ectopic expression of TfCYC2 disrupts the asymmetrical corolla coloring pattern, resulting in a strongly dorsal flower, and the CYC–RAD module coordinates petal shape and corolla pigmentation. When TfCYC2 expression was downregulated, the dorsal petal identity was lost. Diversified CYC genes evolved regulatory loops, and TfCYC2 was directly bound to the regulatory region of the R2R3-MYB gene TfMYB1, resulting in asymmetric expression and ultimately the establishment of asymmetric pigmentation patterns [112]. Integration of the Ty1/Copia-like LTR retrotransposon TORE2 into the exon of TfCYC2, to generate the allele TfCYC2TORE2, inhibited the expression of TfCYC2, which is the main regulatory gene involved in anthocyanin pattern enrichment in T. fournieri [118]. The degree of pigmentation of the dorsal corolla lobe of T. fournieri is negatively correlated with TfCYC2 expression.
4.2. CYC-like Genes of Gesneriaceae
The inflorescences of the Gesneriaceae species are usually double-flowered cymes or monochasia. The flowers are usually bilaterally symmetrical, but some species produce radially symmetrical flowers (e.g., Tengia scopulorum). In recent years, CYC-like genes have been isolated from several members of this family, including Saintpaulia ionantha, Sinningia speciosa, Chirita heterotricha, Primulina heterotricha, and Petrocosmea glabristoma [54,57,119,120]. These genes have the typical functions of CYC-like genes, which affect floral symmetry and stamen abortion.
The successive examination of the CYC-like genes in the C. heterotricha, P. heterotricha, and Petrocosmea species showed that their expression in developing flowers is regulated by various mechanisms [54,119,120]. First, the promoter sequences of the CYC homologs ChCYC1C and ChCYC1D in C. heterotricha were isolated. Subsequent analysis indicated the genes may have evolved automatic regulatory loops to maintain expression during the establishment of bilaterally symmetrical flowers [119]. The RAD-like gene ChRAD may be directly targeted by ChCYC1 as part of a regulatory network. Next, the expression and function of two CYC2 genes (CYC1C and CYC1D) in P. heterotricha were analyzed, which revealed positive self-regulatory and cross-regulatory effects [120]. This mechanism may lead to the independent formation of bilaterally symmetrical flowers, which is associated with plant–insect co-evolution and the adaptive radiation of angiosperms. Finally, changes in the dorsal corolla lobe size of P. glabristoma and Petrocosmea sinensis were determined to be mainly mediated by the expression and differentiation of CYC1C and CYC1D, and the changes in the petal shape were associated with the expression-level changes to the CIN-like TCP gene CIN1 (Figure 7) [54]. Highly redundant homologous genes with the same expression patterns and interspecific differences in expression may be controlled by markedly different regulatory pathways, because natural selection may have resulted in diverse regulatory modifications rather than sequence changes to key developmental genes to generate morphological diversity [54].

Figure 7.
Front and side view and floral diagram of normal and peloric flowers of Petrocosmea sinensis [54]. (A–C) Front and side view and floral diagram of P. sinensis normal flowers, which have a typical bilaterally symmetrical corolla. (D–F) Front and side view and floral diagram of P. sinensis peloric flowers, which have a radially symmetrical corolla. DP, dorsal corolla lobes; DS, dorsal stamens; LS, lateral stamens; LP, lateral corolla lobes; VS, ventral stamens; VP, ventral corolla lobe. Reprinted with permission from Ref. [54]. Copyright © 2023, Oxford University Press.
Hsu et al. reported that in S. speciosa, the dorsal corolla lobes are bent outward, the midvein of the lateral corolla lobes is asymmetrical, and the expansion of the ventral area of the corolla is closely related to the SsCYC genotype [57]. Expression shifts of the CYC-like genes SiCYC and SiCYC1B, which show dorsal-specific expression in the wild-type S. ionantha, led to two completely different reversals of radial symmetry, namely dorsalized actinomorphic (DA) and ventralized actinomorphic (VA) peloria, which may be controlled by upstream trans-acting factors or epigenetic regulation [121]. SiCYC and SiCYC1B were metastasized with an ectopically extended expression on all corolla lobes in DA, whereas their dorsal-specific expression was greatly reduced in VA [121]. The main highly expressed copies of SiCYC were constrained by purification selection, whereas selection of the low-expression helper gene SiCYC1B was relaxed after duplication [121]. Heterologous expression of SiCYC in A. thaliana was characterized by delayed corolla growth owing to limited cell proliferation [121].
CYC-like gene duplication events have occurred at least five times in the evolutionary history of Gesneriaceae [51]. Three copies of CYC-like genes in the actinomorphic Conandron ramondioides were not expressed in the corolla, whereas the zygomorphic species Hemiboea bicornuta and Lysionotus pauciflorus retained a CYC1 copy (i.e., GCYC1C and GCYC1D, respectively) expressed in dorsal corolla lobes [51]. Selective relaxation after the duplication of CYC1 created evolutionary diversification, in which multiple copies retained the effect of random differentiation affecting the dorsal-specific expression of genes associated with floral symmetry changes [51]. The promoter region of CpCYC is a key determinant of its specific expression in the dorsal corolla lobe of Chirita pumila, where the LEAFY element may directly activate and regulate CpCYC to form a bilaterally symmetrical flower [122].
Yang et al. determined that the ortholog of LjCYC1 in S. ionantha is highly expressed in the root, leaf, peduncle, calyx, petal, stamen, and pistil of transgenic S. ionantha plants [123]. Two flower-type variations were observed in T1 transgenic plants. The first was the change in floral symmetry. Specifically, radially symmetrical wild-type flowers were replaced by bilaterally symmetrical flowers or flowers with obvious differences between the dorsal and ventral corolla lobes. The second variation involved floral organ morphology (e.g., a lobe incision towards the base of the corolla, and stamen, pistil, and calyx petalization). Liu et al. functionally characterized the CYC-like flower symmetry-related gene CpCYC in C. pumila [124]. By transforming plants with a RNAi:CpCYC vector, vertically radially symmetrical flowers were obtained, implying that CpCYC determines the establishment of zygomorphy and the horizontal plane of flowers. The insertion of a CpCYC promoter:GUS vector into C. pumila confirmed that the CpCYC promoter was active in dorsal corolla lobes, dorsal/lateral staminodes, and pedicels.
4.3. CYC-like Genes of Phrymaceae
The Phrymaceae species have bilaterally symmetrical bisexual flowers borne in spikes at the top of the stem and in the upper leaf axils. To date, there has been relatively little research on the CYC-like genes in this family. The flowers of the Phrymaceae species Diplacus pictus have distinct dorsal, ventral, and lateral corolla lobes. The expression and function of CYC genes may vary between D. pictus and snapdragon [125]. The CYC-like gene DpCYC is expressed in a narrow part of the upper lip of the dorsal corolla lobe. The novel upturned abaxial corolla lobe of D. pictus may be associated with the localized expression of DpCYC on the upper surface of this structure.
5. Progress in Research on CYC-like Genes in Orchidaceae
Orchidaceae is a large family, second in species number only to Asteraceae and the largest family of monocotyledons [126,127]. The bisexual flowers of the members of this family are generally bilaterally symmetrical [128]. The median tepal is typically modified into the labellum, which is often in the abaxial position because the inflorescence droops or the pedicel twists [129]. Many CYC-like genes in Phalaenopsis and Cattleya trianae have been identified and observed to influence floral or inflorescence development [130,131,132].
Lin et al. observed that the ECE clade gene PeCIN8 is highly expressed in the late ovule developmental stage, with overlapping expression on the 16th day after the pollination of Phalaenopsis equestris flowers [130]. Hence PeCIN8 may be crucial for orchid ovule development because of its regulatory effects on cell division. In addition, Liu et al. recently identified 10 ECE clade genes (CgCINs and CgCYCs) from the Cymbidium goeringii genome and observed that flower-specific gene expression may be associated with the regulation of flower development [131].
The CYC-like genes are single, short, low-expressed copies in Orchidoideae [132]. Madrigal et al. performed a phylogenetic analysis of the TCP genes in C. trianae and observed that the TCP genes were resolved into three major clades with multiple gene duplication events detected [132]. Among these genes, the CYC-like genes are single short copies with low expression, and their homogeneous expression in the labial and lateral tepals suggested that they contribute little to bilateral perianth formation.
6. Progress in Research on CYC-like Genes in Solanaceae
The Solanaceae is dominated by species with radially symmetrical flowers, but the early-diverging clades often show bilateral flower symmetry [133]. At present, few studies have explored the influence of CYC-like genes on flower development in the Solanaceae. Petunia hybrida is an annual herb that produces solitary flowers in leaf axils [134]. Its funnel-shaped corolla is white or purple and may be variously striped. There is considerable diversity in the flower types of P. hybrida. Research on Petunia CYC-like genes of the ECE clade showed that they may be mainly associated with the growth and development of axillary buds, while also modulating stem growth and the development of branches, flowers, and leaves (e.g., size) [135,136,137].
Zou et al. isolated the P. hybrida CYC1 clade genes PhTCP3 and PhTCP4, the CYC2 clade gene PhTCP2, and the CYC3 clade gene PhTCP1, which were predominantly associated with axillary bud growth and development [135]. These four genes were highly homologous to the ECE clade genes from tomato (Solanum lycopersicum), gerbera, grape (Vitis vinifera), poplar (Populus), A. thaliana, and other plant species and exhibited tissue-specific expression patterns, and the expression levels in the axillary buds were significantly higher than those in the other tissues analyzed [135]. The overexpression and silencing of PhTCP1 led to decreased and increased branching, respectively, suggestive of the regulatory effects of PhTCP1 on branch formation and development.
Zhang et al. conducted the first genome-wide spatiotemporal expression profile and promoter analysis of the petunia ECE clade PaTCP genes, and reported that these genes may play an important role in the various developmental processes of petunia through multiple hormonal pathways, especially in petal development and the formation of petal size [136]. Sengupta and Hileman observed that the CYC homologous genes positively regulate the RAD homologous genes in tomato, which was similar to previous findings in snapdragon [137].
7. Progress in Research on CYC-like Genes in Other Angiosperms
To date, there have been only a few studies on CYC-like genes in other angiosperm families. These investigations have indicated the genes are primarily expressed in floral organs and affect the establishment of floral symmetry and stamen fertility [138,139,140,141,142]. Thus future research needs to be extended to these less-studied plant families.
7.1. CYC-like Genes of Brassicaceae
The flowers of Brassicaceae are borne in racemes of many small flowers, which are actinomorphic in most species but zygomorphic in a few species [143,144]. Cubas et al. were the first researchers to identify a CYC2 lineage gene, AtTCP1, in A. thaliana that regulates the symmetrical development of floral organs [52]. Arabidopsis flowers are actinomorphic, and AtTCP1 is briefly expressed specifically in the dorsal region of the floral primordium, which suggests that AtTCP1 does not play a key role in floral organ differentiation because there is no flower-specific direct transcriptional autoregulation or other expression pattern that persists until advanced stages of flower differentiation [52].
The corolla of the genus Iberis is zygomorphic, with two small adaxial petals and two large abaxial petals [145]. Busch et al. identified a CYC homolog from Iberis amara, IaTCP1, for which the temporary alteration of expression is important for the control of zygomorphic corolla formation [146]. The timing of IaTCP1 expression differed from that of AtTCP1 and other CYC homologs. Specifically, IaTCP1 expression was lacking early in asymmetric petal development, but the gene was strongly differentially expressed in the corolla during advanced asymmetric petal development. In addition, IaTCP1 activated the expression of many more genes in adaxial petals than it inhibited [147]. These findings suggest that asymmetric corolla formation in I. amara may be caused by the strong expression of the CYC2 clade gene in the small ventral petals and weak expression in the large dorsal petals.
Busch et al. revealed through phylogenetic reconstruction that the zygomorphic genera Iberis, Calepina, and Teesdalia belong to a major Brassicaceae lineage [148]. Zygomorphy is most evident in Iberis, but less so in Calepina and Teesdalia, with an expression-dependent positive correlation between the strength of the difference in CYC2 expression and the degree of zygomorphy [148]. This study suggested that zygomorphy evolved through the heterochronic expression of CYC2, from paraxial expression in the ancestral floral meristem to accumulation of paraxial CYC2 transcripts late in petal development in the Brassicaceae.
7.2. CYC-like Genes of Dipsacales
Howarth and Donoghue identified three major ECE clade CYC-like genes (DipsCYC1, DipsCYC2, and DipsCYC3) in the ancestors of Dipsacales, as well as additional duplication events involving genes in this clade [26]. The DipsCYC1 gene was not involved in subsequent replication events and may not be expressed in flower tissues. In contrast, DipsCYC2 and DipsCYC3 had similar duplication patterns in several clades. In the Caprifoliaceae species Lonicera morrowii, DipsCYC2B was expressed in the four dorsal petals, but not in the ventral petal, whereas DipsCYC3B was expressed in the flowers and petal primordia, with the peak expression level in the ventral petal [26].
Berger et al. compared the expression patterns of six CYC-like genes in the dorsal, lateral, and ventral petals of the inner and outer florets of the capitulum of Knautia macedonica and observed that CYC-like genes duplicated many times were differentially expressed among the petal types and the inner and outer florets [55]. The formation of bilateral symmetry may be regulated by a dorsoventral expression gradient. In addition, dorsoventral morphological specificity was associated with significant changes in ventral petal gene expression.
7.3. CYC-like Genes of Zingiberales
Bartlett and Specht showed that gene replication unique to the Zingiberales, including one replication event involving the ECE clade gene TBL, preceded the diversification of commelinid monocotyledons [142]. The changes in TBL expression were associated with the evolutionary changes in floral symmetry and stamen abortion. In addition, ZinTBL1a was expressed in the posterior (adaxial) stamen lip of Heliconia stricta (Heliconiaceae) and the anterior (abaxial) stamen lip of Costus spicatus (Costaceae) [141]. The ZinTBL2 expression level was highest in the anterior sepals of H. stricta and the posterior fertile stamens of C. spicatus. These findings indicate that the ECE clade genes were repeatedly recruited in the evolutionary process, which accelerated the evolution of bilaterally symmetrical flowers.
Canna indica (Cannaceae) has noticeably asymmetrical flowers, in which the stamens develop into petal-like staminodes or are aborted (semi-fertile stamens) at an early developmental stage associated with three ECE clade homologs (CiTBL1a, CiTBL1b-1, and CiTBL1b-2) (Figure 8) [142]. The overexpression of CiTBL genes in A. thaliana resulted in dwarfism, the production of small petals and relatively few stamens, as well as mature flowers with altered symmetry, which provided evidence of the involvement of ECE clade homologs in the development of asymmetrical C. indica flowers.
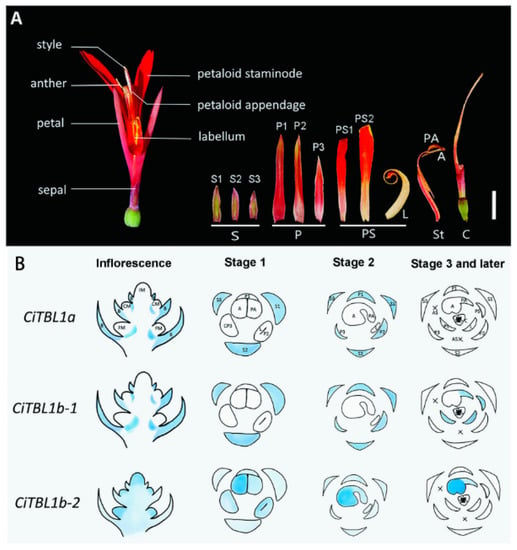
Figure 8.
Morphology of Canna indica flowers and expression of CiTBL genes [142] (Yu et al., 2020). (A) Flower morphology. A: anthers; C: carpel; L: labellum; P: petals; PA: petaloid appendage; PS: petaloid staminodes; S: sepals; St: staminodes. (B) Expression of CiTBL1a, CiTBL1b-1, and CiTBL1b-2 in young inflorescences and flowers at different developmental stages. Gene expression sites are indicated in blue, with the intensity of the coloration reflecting the expression level. AS: abortive staminodes; B: primary bracts; CM: meristem of monochasium; CP: common primordium of the petal and stamen; FM: floral meristem; IM: inflorescence meristem. Reprinted with permission from Ref. [142]. Copyright © 2023 Frontiers Media S.A.
7.4. CYC-like Genes of Ranunculales
Ranunculaceae underwent an evolutionary transition from actinomorphy to zygomorphy, with the accumulation of as many as four copies of the CYC-like gene RanaCyL in zygomorphic species [149,150]. The RanaCyL homologous genes are expressed early in flower bud development, and the expression duration varies with species and gene class. In actinomorphic species, at most one RanaCyL paralog is expressed late in flower development, whereas in zygomorphic species, all paralogs are expressed, constituting a species-specific recognition code for the perianth [149].
Fumariaceae and Papaveraceae show morphological diversity in flower symmetry and inflorescence structure, which may be related to the duplication and functional diversity of CYC-like genes [151]. Damerval et al. reported that the homologous genes of CYC in Papaveraceae, PAPACYL1, and PAPACYL2, which are ECE clade members, were expressed during the flower development of all three species studied and were specifically expressed in the outer petals of the two species with asymmetrical flowers [152].
The CYC-like CYL genes of the Papaveraceae species Eschscholzia california and Cysticapnos vesicaria have highly diverse expression patterns and functions [153]. The silencing of EscaCYL1 enhances the control of bud branching, whereas PapaCYL genes promote germination and growth of stamens. In addition, CyveCYL genes are involved in the regulation of floral symmetry and perianth development of Cysticapnos by regulating B-class floral–organ identity genes to determine sepal and petal characteristics.
7.5. CYC-like Genes of other Families
An ECE clade gene, CcCYC, is not expressed in the radially symmetrical perianth of Tradescantia pallida, but is expressed asymmetrically in the bilaterally symmetrical perianth of Commelina communis and Commelina dianthifolia [138]. These observations were related to genes that were recruited in parallel through the independent evolution of flower bilateral symmetry in the early stage of floral development. The Actinodium cunninghamii (Myrtaceae) capitulum consists of a pseudanthium, with a ray flower that is not a single flower but a branch with a short bud that flowers occasionally; its proximal branch is also similar to the ray flower [139]. The changes in the expression of CYC-like genes in the pseudanthium modulated ray flower structures and branching patterns. This gene expression pattern is similar to that observed in the distantly related Asteraceae species, indicating that flowering plants seem to have recruited CYC-like genes at least twice in their evolutionary history for the development of heterotypic inflorescences.
Horn et al. showed that the ECE clade gene CYCL is present in basal angiosperms and Magnoliaceae species [140]. In Aristolochia, CYCL was involved in the differentiation of the perianth and the mushroom pseudo-structure, but did not participate in the process mediating the formation of zygomorphic flowers. Only when the TCP domain of the Aristolochia CYCL gene was replaced by the CYC2 domain could the functionally similar gene be obtained. The differentiation and evolution of the ECE lineage led to significant changes in the coding region and the cis-regulatory elements, which ultimately established CYC2 as a key gene regulating floral zygomorphy in dicotyledons. Pabon-Mora et al. reported that ECE clade genes may also be involved in cell division in leaves, pistils, and ovules [154]. Specifically, CYC-like genes maintain differential expansion of the perianth by promoting cell division in the distal and ventral extremities during middle and late flower development in Aristolochia fimbriata.
Zhang and co-workers confirmed that the expression of the CYC2-like genes CYC2A and CYC2B was associated with the floral symmetry of Malpighiaceae, and that relaxation of their conserved expression and expansion to a wider floral area (including the dorsal stamen) were related to the development of dorsoventral heteranthery in Hiptage benghalensis and contributed to the elaborated androecium, which is essential for adaptation to the new pollination strategy [56,155]. Berger et al. compared the corolla shape of Fedia graciliflora expressing the wild-type or knocked-out CYC2-like gene FgCYC2A using canonical variable analysis, and observed that gene knockout resulted in significant changes in flower shape, which affected the position of the dorsal lobe relative to the lateral lobe and led to more radially symmetrical flowers [156].
Radially symmetric Rhododendron taxifolium and bilaterally symmetric Rhododendron beyerinckianum have four and five CYC-like genes from shared tandem duplications, respectively [28]. The CYC-like genes are expressed in the longer dorsal petals and stamens, and are highly expressed in the pistil of R. beyerinckianum, whereas in R. taxifolium the orthologs are either ubiquitously expressed, have been lost from the genome, or are weakly expressed [28]. As the main regulatory factor for the growth of differentiated organs in Rhododendron, CYC-like genes did not regulate the expression of RAD-like genes, which revealed a certain deviation from the typical floral symmetry-related gene regulatory network of asterids [28].
Three CYC-like genes (CamCYC1, CamCYC2, and CamCYC3) in Campanulaceae have undergone dynamic changes in replication and loss, including the first instance of the loss of CamCYC2 in a bilaterally symmetrical group [27]. The CamCYC1 gene was included in duplication events in the radially symmetrical Campanuloideae species, whereas CamCYC2 was duplicated but CamCYC3 was lost at an early stage of divergence, in the bilaterally symmetrical and inverted Lobelioideae species [27]. In addition, the bilaterally symmetrical and non-inverted Cyphioideae species lost CamCYC2, but replicated CamCYC3 [27]. The late expression of CamCYC2 along the dorsoventral axis of the inverted flower was confirmed, and was not regulated by external factors, such as gravity [27].
8. Outlook
Researchers have conducted systematic and detailed studies on the CYC-like genes of many angiosperm families, such as Fabaceae, Asteraceae, Scrophulariaceae, Gesneriaceae, and Orchidaceae. However there are still many issues regarding the function and evolution of CYC-like genes that require exploration in greater detail.
8.1. Conduct Systematic Functional and Evolutionary Research, Especially Regarding CYC1 and CYC3 Clade Members
The CYC-like genes have extensive and important roles affecting plant development [26,29,30]. The current relevant research has mainly focused on the CYC2 clade, which is primarily associated with the regulation of floral symmetry, with less research conducted on the CYC1 and CYC3 clades [53,54,55,56,57,112]. Therefore, the functions and evolution of CYC1 and CYC3 genes should be investigated, to expand our understanding of the contributions of CYC-like genes to the growth and development of angiosperms.
8.2. Functionally Characterize the CYC-like Genes in More Plant Groups
Through developmental biology, genetics, and evolutionary genetics, scientists revealed that the ancestors of CYC in core eudicots were expressed in the dorsal flower organs, thus affecting floral symmetry [19,26,112,157]. The CYC genes are expressed in the ventral floral organs of several monocot groups (Zingiberaceae, Alstroemeriaceae, and Commelinaceae) [138,141,158]. In Alstroemeriaceae, Hoshino et al. observed that the CYC-like genes AaTCP1, AmTCP1, ApTCP1, and ApTCP2, which belong to the ECE clade, are involved in the development of floral asymmetry and the identity of ventral floral organs in Alstroemeria aurea, Alstroemeria magenta, and Alstroemeria pelegrina with bilaterally symmetrical flowers [158]. In addition, AaTCP1 transcripts were specifically accumulated in flower buds and located at the paraxial perianth base of A. aurea. These results reflect the complexity of the CYC expression pattern in angiosperms. Additional research on these genes and their regulatory effects on floral symmetry will require the inclusion of more plant groups.
8.3. Investigate the Regulatory Elements Upstream of CYC-like Genes
Increasing numbers of studies have isolated and analyzed the phylogenetic relationships, expression patterns, and functions of CYC-like homologs in different angiosperm groups, but there has been minimal research on the upstream regulatory elements. Yang and co-workers determined that the bilateral symmetry of the flowers in Gesneriaceae may have involved the evolution of an automatic regulatory loop for the CYC-like gene [119,120]. In the double-flowered (dbl) sunflower mutant, HaCYC2c inserted into the promoter region is usually expressed specifically in wild-type ray florets, but not throughout the capitulum, possibly resulting in the inability to observe radially symmetrical flowers [79]. CmWUS can bind to the cis-acting element TAAT in the CmCYC3a promoter in yeast, and activate the expression of resistance genes, while also regulating floral symmetry and flower organ development together with ECE TFs in chrysanthemum [89]. The chrysanthemum TF CmCYC2c can bind to the cis-acting element of CmCYC2f to activate its expression, but it can also form heterodimers with CmCYC2c-2, CmCYC2d, and CmCYC2e, which may participate in the regulation of floral organ symmetry [88]. The spatiotemporal expression patterns and functions of CYC-like genes in different flower organs in different taxonomic groups are diverse, which may be related to changes in the upstream regulatory elements, ultimately resulting in a variety of angiosperm flower types. Therefore, the regulatory elements upstream of CYC-like genes must be studied, which will help to clarify the evolution and functional differentiation of these genes in angiosperms.
8.4. Study the Phylogenetic Relationships and Expression of CYC-like Genes with New Techniques and Methods
In addition to traditional methods for verifying gene functions, methylation analyses and other technical methods should be used to study the apparent modifications to CYC-like genes. Zhang et al. used qRT-PCR and bisulfite sequencing PCR techniques to determine the expression patterns and DNA methylation patterns of CYC2-like genes in two types of chrysanthemum florets, thereby providing new epigenetic-related insights into the formation of the capitulum in Asteraceae [98]. Sun et al. confirmed that CYC2c is the main factor influencing the Gaillardia ray–floret phenotype by applying RNA resequencing technology as well as qRT-PCR and gene-silencing methods [73]. In future studies, additional new technologies and methods including comparative genomics can be used to elucidate the role of CYC-like genes in angiosperm floral development, which will lead to new ideas for future research on the evolution and development of angiosperms.
9. Method
We randomly selected CYC-like genes from the published literature to construct a phylogenetic tree (Figure 1) using MEGA 11 software. The nucleotide sequences of the homologous CYC genes were downloaded from NCBI GenBank (https://www.ncbi.nlm.nih.gov/) (accessed on 9 October 2022) and aligned with ClustalW. The phylogenetic tree was constructed with MEGA 11 software using the maximum likelihood method [159]. The accession numbers of the sequence data used to construct the phylogenetic tree are present in the Supplemental Table S1. To assess support for the topology of the tree, a bootstrap analysis with 1000 replications was performed. In the phylogenetic tree, the CYC1, CYC2, and CYC3 clades are labeled with reference to the literature, whereas other CYC-like genes do not have an explicit classification at present [21,37,74,75,76].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cimb45030131/s1, Supplementary Table S1. The accession numbers of the genes used to construct the phylogenetic tree.
Author Contributions
Conceptualization, H.L.; methodology, X.Z.; software, Y.C., W.C., C.G. and H.C.; writing—original draft preparation, Y.C. and H.L.; writing—review and editing, Y.C., H.L. and C.H.; funding acquisition, X.C., C.L. and D.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31901354), Beijing Innovation Consortium of Agriculture Research System (BAIC09-2022), and the Innovation Foundation of the Beijing Academy of Agriculture and Forestry Sciences (KJCX20200112).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No confidential/unpublished data have been used in this article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Luo, D.; Carpenter, R.; Vincent, C.; Copsey, L.; Coen, E. Origin of floral asymmetry in Antirrhinum. Nature 1996, 383, 794–799. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Kosugi, S.; Ohashi, Y. PCF1 and PCF2 specifically bind to cis elements in the rice proliferating cell nuclear antigen gene. Plant Cell 1997, 9, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Carpenter, R.; Copsey, L.; Vincent, C.; Clark, J.; Coen, E. Control of organ asymmetry in flowers of Antirrhinum. Cell 1999, 99, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Manassero, N.G.; Viola, I.L.; Welchen, E.; Gonzalez, D.H. TCP transcription factors: Architectures of plant form. Biomol. Concepts 2013, 4, 111–127. [Google Scholar] [CrossRef]
- Nicolas, M.; Torres-Perez, R.; Wahl, V.; Cruz-Oro, E.; Rodriguez-Buey, M.L.; Zamarreno, A.M.; Martin-Jouve, B.; Garcia-Mina, J.M.; Oliveros, J.C.; Prat, S.; et al. Spatial control of potato tuberization by the TCP transcription factor BRANCHED1b. Nat. Plants 2022, 8, 281–294. [Google Scholar] [CrossRef]
- Damerval, C.; Claudot, C.; Le Guilloux, M.; Conde, E.S.N.; Brunaud, V.; Soubigou-Taconnat, L.; Caius, J.; Delannoy, E.; Nadot, S.; Jabbour, F.; et al. Evolutionary analyses and expression patterns of TCP genes in Ranunculales. Front. Plant Sci. 2022, 13, 1055196. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, P.; Fri, N.; Ijast, I. Genome wide analysis and identification of TCP gene family in Wheat (Triticum aestivum L.). Int. J. Appl. Sci. Technol. 2022, 8, 19–35. [Google Scholar]
- Parapunova, V.; Busscher, M.; Busscher-Lange, J.; Lammers, M.; Karlova, R.; Bovy, A.G.; Angenent, G.C.; de Maagd, R.A. Identification, cloning and characterization of the tomato TCP transcription factor family. BMC Plant Biol. 2014, 14, 157. [Google Scholar] [CrossRef]
- Baulies, J.L.; Bresso, E.G.; Goldy, C.; Palatnik, J.F.; Schommer, C. Potent inhibition of TCP transcription factors by miR319 ensures proper root growth in Arabidopsis. Plant Mol. Biol. 2022, 108, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Igartua, E.; Contreras-Moreira, B.; Casas, A.M. TB1: From domestication gene to tool for many trades. J. Exp. Bot. 2020, 71, 4621–4624. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.T.; Van Es, S.W.; Hernandez-Pinzon, I.; Kirschner, G.K.; Van Der Wal, F.; Da, S.S.; Busscher-Lange, J.; Angenent, G.C.; Moscou, M.; Immink, R.; et al. The TCP transcription factor HvTB2 heterodimerizes with VRS5 and controls spike architecture in barley. Plant Reprod. 2022, 35, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.E.; Pasquariello, M.; Boden, S.A. TEOSINTE BRANCHED1 regulates height and stem internode length in bread wheat. J. Exp. Bot. 2020, 71, 4742–4750. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, D.; Gao, Q.; Luo, Y.; Zhang, H.; Ma, B.; Chen, C.; Whibley, A.; Zhang, Y.; Cao, Y.; et al. Genome structure and evolution of Antirrhinum majus L. Nat. Plants 2019, 5, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Steiner, E.; Livne, S.; Kobinson-Katz, T.; Tal, L.; Pri-Tal, O.; Mosquna, A.; Tarkowska, D.; Mueller, B.; Tarkowski, P.; Weiss, D. The Putative O-Linked N-Acetylglucosamine Transferase SPINDLY Inhibits Class I TCP Proteolysis to Promote Sensitivity to Cytokinin. Plant Physiol. 2016, 171, 1485–1494. [Google Scholar] [CrossRef]
- Busch, A.; Deckena, M.; Almeida-Trapp, M.; Kopischke, S.; Kock, C.; Schussler, E.; Tsiantis, M.; Mithofer, A.; Zachgo, S. MpTCP1 controls cell proliferation and redox processes in Marchantia polymorpha. New Phytol. 2019, 224, 1627–1641. [Google Scholar] [CrossRef]
- Spears, B.J.; McInturf, S.A.; Collins, C.; Chlebowski, M.; Cseke, L.J.; Su, J.; Mendoza-Cozatl, D.G.; Gassmann, W. Class I TCP transcription factor AtTCP8 modulates key brassinosteroid-responsive genes. Plant Physiol. 2022, 190, 1457–1473. [Google Scholar] [CrossRef]
- Cubas, P. Floral zygomorphy, the recurring evolution of a successful trait. Bioessays 2004, 26, 1175–1184. [Google Scholar] [CrossRef]
- Martín-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef]
- Bello, M.A.; Cubas, P.; Alvarez, I.; Sanjuanbenito, G.; Fuertes-Aguilar, J. Evolution and expression patterns of CYC/TB1 genes in Anacyclus: Phylogenetic insights for floral symmetry genes in Asteraceae. Front. Plant Sci. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Levin, K.A.; Boden, S.A. A new branch of understanding for barley inflorescence development. J. Exp. Bot. 2020, 71, 6869–6871. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P. Role of TCP genes in the evolution of morphological characters in angiosperms. In Role of TCP Genes in the Evolution of Key Morphological Characters in Angiosperms; Cronk, Q.C.B., Hawkins, J., Bateman, R.M., Eds.; Taylor & Francis: London, UK, 2002; pp. 247–266. [Google Scholar]
- Howarth, D.G.; Donoghue, M.J. Duplications in CYC-like genes from Dipsacales correlate with floral form. Int. J. Plant Sci. 2005, 166, 357–370. [Google Scholar] [CrossRef][Green Version]
- Citerne, H.L.; Le Guilloux, M.; Sannier, J.; Nadot, S.; Damerval, C. Combining phylogenetic and syntenic analyses for understanding the evolution of TCP ECE genes in eudicots. PLoS ONE 2013, 8, e74803. [Google Scholar] [CrossRef] [PubMed]
- Howarth, D.G.; Donoghue, M.J. Phylogenetic analysis of the “ECE” (CYC/TB1) clade reveals duplications predating the core eudicots. Proc. Natl. Acad. Sci. USA 2006, 103, 9101–9106. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Knox, E.B.; Morden, C.W.; Cellinese, N.; Mossolem, F.; Zubair, A.S.; Howarth, D.G. Duplication and expression patterns of CYCLOIDEA-like genes in Campanulaceae. Evodevo 2022, 13, 5. [Google Scholar] [CrossRef]
- Ramage, E.; Soza, V.L.; Yi, J.; Deal, H.; Chudgar, V.; Hall, B.D.; Di Stilio, V.S. Gene Duplication and Differential Expression of Flower Symmetry Genes in Rhododendron (Ericaceae). Plants 2021, 10, 1994. [Google Scholar] [CrossRef]
- Carlson, S.E.; Howarth, D.G.; Donoghue, M.J. Diversification of CYCLOIDEA-like genes in Dipsacaceae (Dipsacales): Implications for the evolution of capitulum inflorescences. BMC Evol. Biol. 2011, 11, 325. [Google Scholar] [CrossRef]
- Busch, A.; Zachgo, S. Flower symmetry evolution: Towards understanding the abominable mystery of angiosperm radiation. Bioessays 2009, 31, 1181–1190. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Karaaslan, E.S.; Wang, N.; Faiss, N.; Liang, Y.; Montgomery, S.A.; Laubinger, S.; Berendzen, K.W.; Berger, F.; Breuninger, H.; Liu, C. Marchantia TCP transcription factor activity correlates with three-dimensional chromatin structure. Nat. Plants 2020, 6, 1250–1261. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, L.V.; Gastaldi, V.; Ariel, F.D.; Viola, I.L.; Gonzalez, D.H. Class I TCP proteins TCP14 and TCP15 are required for elongation and gene expression responses to auxin. Plant Mol. Biol. 2021, 105, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Tabarelli, M.; Malnoy, M.; Janik, K. Chasing Consistency: An Update of the TCP Gene Family of Malus x Domestica. Genes 2022, 13, 1696. [Google Scholar] [CrossRef] [PubMed]
- Sinou, C.; Cardinal-McTeague, W.; Bruneau, A. Testing generic limits in Cercidoideae (Leguminosae): Insights from plastid and duplicated nuclear gene sequences: Phylogeny and generic limits in Cercidoideae. Taxon 2020, 69, 67–86. [Google Scholar] [CrossRef]
- Fambrini, M.; Pugliesi, C. CYCLOIDEA-2 Clade Genes: Key Players in the Control of Floral Symmetry, Inflorescence Architecture, and Reproductive Organ Development. Plant Mol. Biol. Rep. 2017, 35, 20–36. [Google Scholar] [CrossRef]
- Fambrini, M.; Pugliesi, C. Presence/absence of a CACTA transposon in the CYC2c gene of two genotypes of Helianthus × multiflorus cv. “Meteor” characterized by a radiate inflorescence with different shape of disk flower corollas. Biologia 2019, 74, 1675–1686. [Google Scholar] [CrossRef]
- Fambrini, M.; Bernardi, R.; Pugliesi, C. Ray flower initiation in the Helianthus radula inflorescence is influenced by a functional allele of the HrCYC2c gene. GENESIS 2020, 58, e23401. [Google Scholar] [CrossRef]
- Bukhari, G.; Zhang, J.; Stevens, P.F.; Zhang, W. Evolution of the process underlying floral zygomorphy development in pentapetalous angiosperms. Am. J. Bot. 2017, 104, 1846–1856. [Google Scholar] [CrossRef]
- Sengupta, A.; Hileman, L.C. Novel Traits, Flower Symmetry, and Transcriptional Autoregulation: New Hypotheses From Bioinformatic and Experimental Data. Front. Plant Sci. 2018, 9, 1561. [Google Scholar] [CrossRef]
- Kalisz, S.; Ree, R.H.; Sargent, R.D. Linking floral symmetry genes to breeding system evolution. Trends Plant Sci. 2006, 11, 568–573. [Google Scholar] [CrossRef]
- Endress, P.K. Evolution of floral symmetry. Curr. Opin. Plant Biol. 2010, 4, 86–91. [Google Scholar] [CrossRef]
- Coen, E.; Nugent, J.; Luo, D.; Bradley, D.; Cubas, P.; Chadwick, M.; Copsey, L.; Carpenter, R. Evolution of floral symmetry. Philos. Trans. R. Soc. B 1995, 350, 35–38. [Google Scholar] [CrossRef][Green Version]
- Zhang, J.; Stevens, P.F.; Zhang, W. Evolution and development of inflorescences and floral symmetry in Solanaceae. Am. J. Bot. 2022, 109, 746–767. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Moubayidin, L. Floral symmetry: The geometry of plant reproduction. Emerg. Top. Life Sci. 2022, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Rosin, F.M.; Kramer, E.M. Old dogs, new tricks: Regulatory evolution in conserved genetic modules leads to novel morphologies in plants. Dev. Biol. 2009, 332, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; Fernandez-Mazuecos, M.; Vargas, P. Evolution in the Model Genus Antirrhinum Based on Phylogenomics of Topotypic Material. Front. Plant Sci. 2021, 12, 631178. [Google Scholar] [CrossRef]
- Nicolas, M.; Cubas, P. The Role of TCP Transcription Factors in Shaping Flower Structure, Leaf Morphology, and Plant Architecture. In Plant Transcription Factors; Gonzalez, D.H., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 249–267. [Google Scholar]
- Preston, J.C.; Hileman, L.C.; Cubas, P. Reduce, reuse, and recycle: Developmental evolution of trait diversification. Am. J. Bot. 2011, 98, 397–403. [Google Scholar] [CrossRef]
- Preston, J.C.; Hileman, L.C. Developmental genetics of floral symmetry evolution. Trends Plant Sci. 2009, 14, 147–154. [Google Scholar] [CrossRef]
- Hsin, K.T.; Lu, J.Y.; Moller, M.; Wang, C.N. Gene duplication and relaxation from selective constraints of GCYC genes correlated with various floral symmetry patterns in Asiatic Gesneriaceae tribe Trichosporeae. PLoS ONE 2019, 14, e210054. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Coen, E.; Zapater, J.M. Ancient asymmetries in the evolution of flowers. Curr. Biol. 2001, 11, 1050–1052. [Google Scholar] [CrossRef]
- Preston, J.C.; Martinez, C.C.; Hileman, L.C. Gradual disintegration of the floral symmetry gene network is implicated in the evolution of a wind-pollination syndrome. Proc. Natl. Acad. Sci. USA 2011, 108, 2343–2348. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, X.G.; Li, C.Q.; Liu, J.; Qiu, Z.J.; Dong, Y.; Wang, Y.Z. Distinct Regulatory Changes Underlying Differential Expression of TEOSINTE BRANCHED1-CYCLOIDEA-PROLIFERATING CELL FACTOR Genes Associated with Petal Variations in Zygomorphic Flowers of Petrocosmea spp. of the Family Gesneriaceae. Plant Physiol. 2015, 169, 2138–2151. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.A.; Thompson, V.; Lim, A.; Ricigliano, V.; Howarth, D.G. Elaboration of bilateral symmetry across Knautia macedonica capitula related to changes in ventral petal expression of CYCLOIDEA-like genes. Evodevo 2016, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Kramer, E.M.; Davis, C.C. Differential Expression of CYC2 Genes and the Elaboration of Floral Morphologies in Hiptage, an Old World Genus of Malpighiaceae. Int. J. Plant Sci. 2016, 177, 551–558. [Google Scholar] [CrossRef]
- Hsu, H.C.; Wang, C.N.; Liang, C.H.; Wang, C.C.; Kuo, Y.F. Association between Petal Form Variation and CYC2-like Genotype in a Hybrid Line of Sinningia speciosa. Front. Plant Sci. 2017, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Citerne, H.L.; Pennington, R.T.; Cronk, Q.C. An apparent reversal in floral symmetry in the legume Cadia is a homeotic transformation. Proc. Natl. Acad. Sci. USA 2006, 103, 12017–12020. [Google Scholar] [CrossRef]
- Feng, X.; Zhao, Z.; Tian, Z.; Xu, S.; Luo, Y.; Cai, Z.; Wang, Y.; Yang, J.; Wang, Z.; Weng, L.; et al. Control of petal shape and floral zygomorphy in Lotus japonicus. Proc. Natl. Acad. Sci. USA 2006, 103, 4970–4975. [Google Scholar] [CrossRef]
- Citerne, H.L.; Luo, D.; Pennington, R.T.; Coen, E.; Cronk, Q.C. A phylogenomic investigation of CYCLOIDEA-like TCP genes in the Leguminosae. Plant Physiol. 2003, 131, 1042–1053. [Google Scholar] [CrossRef]
- Ree, R.H.; Citerne, H.L.; Lavin, M.; Cronk, Q.C. Heterogeneous selection on LEGCYC paralogs in relation to flower morphology and the phylogeny of Lupinus (Leguminosae). Mol. Biol. Evol. 2004, 21, 321–331. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Luo, D. LjCYC genes constitute floral dorsoventral asymmetry in Lotus japonicus. J. Integr. Plant Biol. 2010, 52, 959–970. [Google Scholar] [CrossRef]
- Woollacott, C.; Cronk, Q. The hooded mutant of Lathyrus odoratus (Fabaceae) is associated with a cycloidea gene mutation. Botany 2017, 96, 47–55. [Google Scholar] [CrossRef]
- Woollacott, C.; Wang, L.; Beyer, S.; Walus, K.; Cronk, Q. CYCLOIDEA gene activity, local growth and curvature in the dorsal petal of Lathyrus odoratus (Fabaceae). Bot. Lett. 2018, 166, 64–69. [Google Scholar] [CrossRef]
- Ojeda, D.I.; Jaen-Molina, R.; Santos-Guerra, A.; Caujape-Castells, J.; Cronk, Q. Temporal, but not spatial, changes in expression patterns of petal identity genes are associated with loss of papillate conical cells and the shift to bird pollination in Macaronesian Lotus (Leguminosae). Plant Biol. 2017, 19, 420–427. [Google Scholar] [CrossRef]
- Feng, Z.; Liu, N.; Zhang, G.; Xu, S.; Gong, Y. Identification of Branch Related CYL Gene in Soybean and Analysis of Hormone Expression Pattern in Vegetable Soybean Species. Mol. Plant Breed. 2019, 17, 4865–4872. [Google Scholar] [CrossRef]
- Zhao, Z.; Hu, J.; Chen, S.; Luo, Z.; Luo, D.; Wen, J.; Tu, T.; Zhang, D. Evolution of CYCLOIDEA-like genes in Fabales: Insights into duplication patterns and the control of floral symmetry. Mol. Phylogenet. Evol. 2019, 132, 81–89. [Google Scholar] [CrossRef]
- Li, X.; Sun, M.; Jia, Y.; Qiu, D.; Peng, Q.; Zhuang, L. Genetic control of the lateral petal shape and identity of asymmetric flowers in mungbean (Vigna radiata L.). Front. Plant Sci. 2022, 13, 996239. [Google Scholar] [CrossRef]
- Zoulias, N.; Duttke, S.; Garces, H.; Spencer, V.; Kim, M. The Role of Auxin in the Pattern Formation of the Asteraceae Flower Head (Capitulum). Plant Physiol. 2019, 179, 391–401. [Google Scholar] [CrossRef]
- Hernandez, L.F.; Palmer, J.H. Regeneration of the sunflower capitulum after cylindrical wounding of the receptacle. Am. J. Bot. 1988, 75, 1253–1261. [Google Scholar] [CrossRef]
- Broholm, S.K.; Tahtiharju, S.; Laitinen, R.A.; Albert, V.A.; Teeri, T.H.; Elomaa, P. A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence. Proc. Natl. Acad. Sci. USA 2008, 105, 9117–9122. [Google Scholar] [CrossRef]
- Garces, H.M.; Spencer, V.M.; Kim, M. Control of Floret Symmetry by RAY3, SvDIV1B, and SvRAD in the Capitulum of Senecio vulgaris. Plant Physiol. 2016, 171, 2055–2068. [Google Scholar] [CrossRef]
- Sun, P.; Bao, Y.; Zhu, Y.; Huang, N.; Wang, X.; Wu, Z. Possible role of the CYC2c gene in the cornflower-like ray floret phenotype of Gaillardia cultivars. J. Plant Res. 2022, 135, 465–472. [Google Scholar] [CrossRef]
- Tahtiharju, S.; Rijpkema, A.S.; Vetterli, A.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Evolution and diversification of the CYC/TB1 gene family in Asteraceae—A comparative study in Gerbera (Mutisieae) and sunflower (Heliantheae). Mol. Biol. Evol. 2012, 29, 1155–1166. [Google Scholar] [CrossRef]
- Juntheikki-Palovaara, I.; Tahtiharju, S.; Lan, T.; Broholm, S.K.; Rijpkema, A.S.; Ruonala, R.; Kale, L.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Functional diversification of duplicated CYC2 clade genes in regulation of inflorescence development in Gerbera hybrida (Asteraceae). Plant J. 2014, 79, 783–796. [Google Scholar] [CrossRef]
- Spencer, V.; Kim, M. Re”CYC”ling molecular regulators in the evolution and development of flower symmetry. Semin. Cell Dev. Biol. 2018, 79, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.A.; Leebens-Mack, J.H.; Burke, J.M. Positive selection and expression divergence following gene duplication in the sunflower CYCLOIDEA gene family. Mol. Biol. Evol. 2008, 25, 1260–1273. [Google Scholar] [CrossRef] [PubMed]
- Fambrini, M.; Salvini, M.; Pugliesi, C. A transposon-mediate inactivation of a CYCLOIDEA-like gene originates polysymmetric and androgynous ray flowers in Helianthus annuus. Genetica 2011, 139, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.A.; Tang, S.; Draeger, D.; Nambeesan, S.; Shaffer, H.; Barb, J.G.; Knapp, S.J.; Burke, J.M. Genetic analysis of floral symmetry in Van Gogh’s sunflowers reveals independent recruitment of CYCLOIDEA genes in the Asteraceae. PLoS Genet. 2012, 8, e1002628. [Google Scholar] [CrossRef] [PubMed]
- Fambrini, M.; Salvini, M.; Basile, A.; Pugliesi, C. Transposon-dependent induction of Vincent van Gogh’s sunflowers: Exceptions revealed. Genesis 2014, 52, 315–327. [Google Scholar] [CrossRef]
- Mizzotti, C.; Fambrini, M.; Caporali, E.; Masiero, S.; Pugliesi, C. A CYCLOIDEA -like gene mutation in sunflower determines an unusual floret type able to produce filled achenes at the periphery of the pseudanthium. Botany 2015, 93, 171–181. [Google Scholar] [CrossRef]
- He, Z.; Zeng, W.; Chen, W.; Wu, Y.; Wen, G.; Chen, X.; Wang, Q.; Zhou, J.; Li, Y.; Yang, Z.; et al. HaCYC2c regulating the heteromorphous development and functional differentiation of florets by recognizing HaNDUA2 in sunflower. Plant Cell Rep. 2022, 41, 1025–1041. [Google Scholar] [CrossRef]
- Fambrini, M.; Bellanca, M.; Costa, M.M.; Usai, G.; Cavallini, A.; Pugliesi, C. Ligulate inflorescence of Helianthus x multiflorus, cv. Soleil d’Or, correlates with a mis-regulation of a CYCLOIDEA gene characterised by insertion of a transposable element. Plant Biol. 2018, 20, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Broholm, S. The Role of MADS and TCP Transcription Factors in Gerbera Hybrida Flower Development. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2009. [Google Scholar]
- Zhao, Y.; Broholm, S.K.; Wang, F.; Rijpkema, A.S.; Lan, T.; Albert, V.A.; Teeri, T.H.; Elomaa, P. TCP and MADS-Box Transcription Factor Networks Regulate Heteromorphic Flower Type Identity in Gerbera hybrida. Plant Physiol. 2020, 184, 1455–1468. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Cui, M.L.; Cubas, P.; Gillies, A.; Lee, K.; Chapman, M.A.; Abbott, R.J.; Coen, E. Regulatory genes control a key morphological and ecological trait transferred between species. Science 2008, 322, 1116–1119. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Po, C.; Hao, Y.; Feng, L.; Zhou, J.; Luan, S.; Liu, L.; Li, F.; Yuan, S.; Cui, M. Cloning and functional analysis of CYCLOIDEA(CYC)-like SvRAY1 gene from Senecio vulgaris. J. Zhejiang AF Univ. 2021, 38, 1153–1160. [Google Scholar] [CrossRef]
- Huang, D.; Sun, M.; Yuan, C.; Cheng, T.; Wang, J.; Zhang, Q. Isolation and functional analysis of CYC2d orthologous genes from several plants of the tribe Anthemideae. J. Beijing For. Univ. 2017, 39, 63–71. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, M.; Yuan, C.; Han, Y.; Zheng, T.; Cheng, T.; Wang, J.; Zhang, Q. Interactions between WUSCHEL and CYC2-like Transcription Factors in Regulating the Development of Reproductive Organs in Chrysanthemum morifolium. Int. J. Mol. Sci. 2019, 20, 1276. [Google Scholar] [CrossRef]
- Pu, Y.; Huang, H.; Wen, X.; Lu, C.; Zhang, B.; Gu, X.; Qi, S.; Fan, G.; Wang, W.; Dai, S. Comprehensive transcriptomic analysis provides new insights into the mechanism of ray floret morphogenesis in chrysanthemum. BMC Genom. 2020, 21, 728. [Google Scholar] [CrossRef]
- Yuan, C.; Huang, D.; Yang, Y.; Sun, M.; Cheng, T.; Wang, J.; Pan, H.; Zhang, Q. CmCYC2-like transcription factors may interact with each other or bind to the promoter to regulate floral symmetry development in Chrysanthemum morifolium. Plant Mol. Biol. 2020, 103, 159–171. [Google Scholar] [CrossRef]
- Liu, H.; Sun, M.; Pan, H.; Cheng, T.; Wang, J.; Zhang, Q. Two Cyc2CL transcripts (Cyc2CL-1 and Cyc2CL-2) may play key roles in the petal and stamen development of ray florets in chrysanthemum. BMC Plant Biol. 2021, 21, 105. [Google Scholar] [CrossRef]
- Chen, J.; Shen, C.Z.; Guo, Y.P.; Rao, G.Y. Patterning the Asteraceae Capitulum: Duplications and Differential Expression of the Flower Symmetry CYC2-Like Genes. Front. Plant Sci. 2018, 9, 551. [Google Scholar] [CrossRef]
- Wen, X.; Qi, S.; Huang, H.; Wu, X.; Zhang, B.; Fan, G.; Yang, L.; Hong, Y.; Dai, S. The expression and interactions of ABCE-class and CYC2-like genes in the capitulum development of Chrysanthemum lavandulifolium and C. × morifolium. Plant Growth Regul. 2019, 88, 205–214. [Google Scholar] [CrossRef]
- Kironji, G.S. Patterning of the Chrysanthemum Inflorescence Roles of B-Class and CYC2 Subclade Genes. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2019. [Google Scholar]
- Yang, Y.; Yang, Y.J.; Yuan, C.Q.; Zhang, Q.X. Cloning and Expression Analysis of CmTCP7 in Chrysanthemum morifolium. Acta Bot. Boreali-Occident. Sin. 2019, 39, 595–602. [Google Scholar] [CrossRef]
- Shen, C.Z.; Chen, J.; Zhang, C.J.; Rao, G.Y.; Guo, Y.P. Dysfunction of CYC2g is responsible for the evolutionary shift from radiate to disciform flowerheads in the Chrysanthemum group (Asteraceae: Anthemideae). Plant J. 2021, 106, 1024–1038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lu, C.; Qi, S.; Dai, S. Difference Analysis of ClCYC2-Like Genes Expression and DNA Methylation between the Two Types of Florets in Chrysanthemum lavandulifolium. J. Plant Growth Regul. 2022, 41, 1316–1330. [Google Scholar] [CrossRef]
- Elomaa, P.; Zhao, Y.; Zhang, T. Flower heads in Asteraceae-recruitment of conserved developmental regulators to control the flower-like inflorescence architecture. Hortic. Res. 2018, 5, 36. [Google Scholar] [CrossRef]
- Zhong, J.; Kellogg, E.A. Duplication and expression of CYC2-like genes in the origin and maintenance of corolla zygomorphy in Lamiales. New Phytol. 2014, 205, 852–868. [Google Scholar] [CrossRef]
- Ha, Y.H.; Kim, C.; Choi, K.; Kim, J.H. Molecular Phylogeny and Dating of Forsythieae (Oleaceae) Provide Insight into the Miocene History of Eurasian Temperate Shrubs. Front. Plant Sci. 2018, 9, 99. [Google Scholar] [CrossRef]
- Smyth, D.R. Evolution and genetic control of the floral ground plan. New Phytol. 2018, 220, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Ajani, Y.; Jamzad, Z.; Claßen-Bockhoff, R. Floral biology in the endemic Iranian Salvia majdae—Implications for taxonomy, character evolution and conservation. Flora 2021, 287, 151986. [Google Scholar] [CrossRef]
- Wessinger, C.; Hileman, L. Parallelism in Flower Evolution and Development. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 387–408. [Google Scholar] [CrossRef]
- Zhong, J.; Kellogg, E.A. Stepwise evolution of corolla symmetry in CYCLOIDEA2-like and RADIALIS-like gene expression patterns in Lamiales. Am. J. Bot. 2015, 102, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Poulin, V.; Amesefe, D.; Gonzalez, E.; Alexandre, H.; Joly, S. Testing candidate genes linked to corolla shape variation of a pollinator shift in Rhytidophyllum (Gesneriaceae). PLoS ONE 2022, 17, e267540. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Preston, J.C.; Hileman, L.C.; Kellogg, E.A. Repeated and diverse losses of corolla bilateral symmetry in the Lamiaceae. Ann. Bot. 2017, 119, 1211–1223. [Google Scholar] [CrossRef] [PubMed]
- Hileman, L.C.; Cubas, P. An expanded evolutionary role for flower symmetry genes. J. Biol. 2009, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Neel, M.C. Conservation implications of the reproductive ecology of Agalinis acuta (Scrophulariaceae). Am. J. Bot. 2002, 89, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Vincent, C.; Coen, E. An epigenetic mutation responsible for natural variation in floral symmetry. Nature 1999, 401, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Hileman, L.C.; Kramer, E.M.; Baum, D.A. Differential regulation of symmetry genes and the evolution of floral morphologies. Proc. Natl. Acad. Sci. USA 2003, 100, 12814–12819. [Google Scholar] [CrossRef]
- Su, S.; Xiao, W.; Guo, W.; Yao, X.; Xiao, J.; Ye, Z.; Wang, N.; Jiao, K.; Lei, M.; Peng, Q.; et al. The CYCLOIDEA-RADIALIS module regulates petal shape and pigmentation, leading to bilateral corolla symmetry in Torenia fournieri (Linderniaceae). New Phytol. 2017, 215, 1582–1593. [Google Scholar] [CrossRef]
- Clark, J.I.; Coen, E.S. The cycloidea gene can respond to a common dorsoventral prepattern in Antirrhinum. Plant J. 2002, 30, 639–648. [Google Scholar] [CrossRef]
- Preston, J.C.; Kost, M.A.; Hileman, L.C. Conservation and diversification of the symmetry developmental program among close relatives of snapdragon with divergent floral morphologies. New Phytol. 2009, 182, 751–762. [Google Scholar] [CrossRef]
- Gubitz, T.; Caldwell, A.; Hudson, A. Rapid molecular evolution of CYCLOIDEA-like genes in Antirrhinum and its relatives. Mol. Biol. Evol. 2003, 20, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Corley, S.B.; Carpenter, R.; Copsey, L.; Coen, E. Floral asymmetry involves an interplay between TCP and MYB transcription factors in Antirrhinum. Proc. Natl. Acad. Sci. USA 2005, 102, 5068–5073. [Google Scholar] [CrossRef] [PubMed]
- Theissen, G. Evolutionary developmental genetics of floral symmetry: The revealing power of Linnaeus’ monstrous flower. Bioessays 2000, 22, 209–213. [Google Scholar] [CrossRef]
- Kondo, M.; Tanikawa, N.; Nishijima, T. Mutation of CYCLOIDEA expands variation of dorsal-ventral flower asymmetry expressed as a pigmentation pattern in Torenia fournieri cultivars. Hortic. J. 2020, 89, 481–487. [Google Scholar] [CrossRef]
- Yang, X.; Cui, H.; Yuan, Z.; Wang, Y. Significance of consensus CYC-binding sites found in the promoters of both ChCYC and ChRAD genes in Chirita heterotricha (Gesneriaceae). J. Syst. Evol. 2010, 48, 249–256. [Google Scholar] [CrossRef]
- Yang, X.; Pang, H.B.; Liu, B.L.; Qiu, Z.J.; Gao, Q.; Wei, L.; Dong, Y.; Wang, Y.Z. Evolution of double positive autoregulatory feedback loops in CYCLOIDEA2 clade genes is associated with the origin of floral zygomorphy. Plant Cell 2012, 24, 1834–1847. [Google Scholar] [CrossRef]
- Hsu, H.J.; He, C.W.; Kuo, W.H.; Hsin, K.T.; Lu, J.Y.; Pan, Z.J.; Wang, C.N. Genetic analysis of floral symmetry transition in African violet suggests the involvement of trans-acting factor for CYCLOIDEA expression shifts. Front. Plant Sci. 2018, 9, 1008. [Google Scholar] [CrossRef]
- Liu, J.; Jie, W.; Yang, X.; Wang, Y. Regulatory pathways of CYC-like genes in patterning floral zygomorphy exemplified in Chirita pumila. J. Syst. Evol. 2020, 59, 567–580. [Google Scholar] [CrossRef]
- Yang, M.; Xu, J.; Pang, J. Flower Shape Changes of African Violets Caused by LjCYC1 Gene in Lotus japonicus. Acta Horticulturae Sinica 2020, 47, 708–716. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.J.; Wu, J.; Wang, Y.; Liu, Q.; Liu, F.P.; Yang, X.; Wang, Y.Z. An Optimized Transformation System and Functional Test of CYC-Like TCP Gene CpCYC in Chirita pumila (Gesneriaceae). Int. J. Mol. Sci. 2021, 22, 4544. [Google Scholar] [CrossRef]
- Ferraro, B.J. Examining the roles of CYCLOIDEA, RADIALIS and DIVARICATA in driving the evolution of flower shape Californian Diplacus pictus (Curran ex Greene) Nesom (Phrymaceae). Master’s Thesis, California State University, Long Beach, CA, USA, 2014. [Google Scholar]
- Lucibelli, F.; Valoroso, M.C.; Aceto, S. Radial or bilateral? The molecular basis of floral symmetry. Genes 2020, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Lucibelli, F.; Valoroso, M.; Theißen, G.; Nolden, S.; Mondragon Palomino, M.; Aceto, S. Extending the toolkit for beauty: Differential co-expression of DROOPING LEAF-like and class B MADS-Box genes during Phalaenopsis flower development. Int. J. Mol. Sci. 2021, 22, 7025. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, Y.; Pabon-Mora, N.; Alzate, J. Unraveling the genetic basis of floral symmetry variation in monocots with an emphasis on Asparagales. In Proceedings of the World Orchid Conference 2017, Guayaquil, Ecuador, 8–12 November 2017. [Google Scholar]
- Klepikova, A.V.; Kasianov, A.S.; Ezhova, M.A.; Penin, A.A.; Logacheva, M.D. Transcriptome atlas of Phalaenopsis equestris. PeerJ 2021, 9, e12600. [Google Scholar] [CrossRef]
- Lin, Y.F.; Chen, Y.Y.; Hsiao, Y.Y.; Shen, C.Y.; Hsu, J.L.; Yeh, C.M.; Mitsuda, N.; Ohme-Takagi, M.; Liu, Z.J.; Tsai, W.C. Genome-wide identification and characterization of TCP genes involved in ovule development of Phalaenopsis equestris. J. Exp. Bot. 2016, 67, 5051–5066. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.K.; Zhang, C.; Zhao, X.; Ke, S.; Li, Y.; Zhang, D.; Zheng, Q.; Li, M.H.; Lan, S.; Liu, Z.J. Genome-wide analysis of the TCP gene family and their expression pattern in Cymbidium goeringii. Front. Plant Sci. 2022, 13, 1068969. [Google Scholar] [CrossRef]
- Madrigal, Y.; Alzate, J.F.; Pabon-Mora, N. Evolution and expression patterns of TCP genes in Asparagales. Front. Plant Sci. 2017, 8, 9. [Google Scholar] [CrossRef]
- Zhang, J.; Stevens, P.; Zhang, W. Evolution of floral zygomorphy in androecium and corolla in Solanaceae. J. Syst. Evol. 2017, 55, 581–590. [Google Scholar] [CrossRef]
- Shimada, A.; Kimura, Y. Influence of glyphosate on flower morphogenesis and pigmentation in Petunia hybrida. Z. Naturforsch. C 2006, 61, 578–582. [Google Scholar] [CrossRef][Green Version]
- Zou, S.; Wang, H.; Yu, Y.; Ma, J.; Guo, Y.; Li, M. Studies on molecular cloning, expression pattern of ECE Clade TCP genes in Petunia. Acta Hortic. Sin. 2013, 40, 307–316. [Google Scholar] [CrossRef]
- Zhang, S.; Zhou, Q.; Chen, F.; Wu, L.; Liu, B.; Li, F.; Zhang, J.; Bao, M.; Liu, G. Genome-Wide Identification, Characterization and Expression Analysis of TCP Transcription Factors in Petunia. Int. J. Mol. Sci. 2020, 21, 6594. [Google Scholar] [CrossRef]
- Sengupta, A.; Hileman, L.C. A CYC-RAD-DIV-DRIF interaction likely pre-dates the origin of floral monosymmetry in Lamiales. Evodevo 2022, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Preston, J.C.; Hileman, L.C. Parallel evolution of TCP and B-class genes in Commelinaceae flower bilateral symmetry. Evodevo 2012, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Classen-Bockhoff, R.; Ruonala, R.; Bull-Herenu, K.; Marchant, N.; Albert, V.A. The unique pseudanthium of Actinodium (Myrtaceae)—Morphological reinvestigation and possible regulation by CYCLOIDEA-like genes. Evodevo 2013, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Pabon-Mora, N.; Theuss, V.S.; Busch, A.; Zachgo, S. Analysis of the CYC/TB1 class of TCP transcription factors in basal angiosperms and magnoliids. Plant J. 2015, 81, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, M.E.; Specht, C.D. Changes in expression pattern of the teosinte branched1-like genes in the Zingiberales provide a mechanism for evolutionary shifts in symmetry across the order. Am. J. Bot. 2011, 98, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Tian, X.; Lin, C.; Specht, C.D.; Liao, J. Expression and function studies of CYC/TB1-like genes in the asymmetric flower Canna (Cannaceae, Zingiberales). Front. Plant Sci. 2020, 11, 580576. [Google Scholar] [CrossRef] [PubMed]
- Iljinska, A.P. The range of the morphological features of Brassicaceae s. l.: Inflorescence, flower. Ukr. Bot. J. 2015, 72, 122–134. [Google Scholar] [CrossRef]
- Nikolov, L.A. Brassicaceae flowers: Diversity amid uniformity. J. Exp. Bot. 2019, 70, 2623–2635. [Google Scholar] [CrossRef]
- Reichling, J.; Saller, R. Iberis amara L. (Bittere Schleifenblume)—Profil einer Heilpflanze. Complement. Med. Res. 2002, 9, 21–33. [Google Scholar] [CrossRef]
- Busch, A.; Zachgo, S. Control of corolla monosymmetry in the Brassicaceae Iberis amara. Proc. Natl. Acad. Sci. USA 2007, 104, 16714–16719. [Google Scholar] [CrossRef]
- Busch, A.; Horn, S.; Zachgo, S. Differential transcriptome analysis reveals insight into monosymmetric corolla development of the crucifer Iberis amara. BMC Plant Biol. 2014, 14, 285. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.; Horn, S.; Muhlhausen, A.; Mummenhoff, K.; Zachgo, S. Corolla monosymmetry: Evolution of a morphological novelty in the Brassicaceae family. Mol. Biol. Evol. 2012, 29, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, F.; Cossard, G.; Le Guilloux, M.; Sannier, J.; Nadot, S.; Damerval, C. Specific duplication and dorsoventrally asymmetric expression patterns of Cycloidea-like genes in zygomorphic species of Ranunculaceae. PLoS ONE 2014, 9, e95727. [Google Scholar] [CrossRef] [PubMed]
- Zalko, J.; Frachon, S.; Morel, A.; Deroin, T.; Espinosa Moreno, F.; Xiang, K.; Wang, W.; Zhang, W.; Lang, S.; Dixon, L.; et al. Floral organogenesis and morphogenesis of Staphisagria (Ranunculaceae): Implications for the evolution of synorganized floral structures in Delphinieae. Int. J. Plant Sci. 2020, 182, 59–70. [Google Scholar] [CrossRef]
- Kolsch, A.; Gleissberg, S. Diversification of CYCLOIDEA-like TCP genes in the basal eudicot families Fumariaceae and Papaveraceae s.str. Plant Biol. 2006, 8, 680–687. [Google Scholar] [CrossRef]
- Damerval, C.; Le Guilloux, M.; Jager, M.; Charon, C. Diversity and evolution of CYCLOIDEA-like TCP genes in relation to flower development in Papaveraceae. Plant Physiol. 2007, 143, 759–772. [Google Scholar] [CrossRef]
- Zhao, Y.; Pfannebecker, K.; Dommes, A.B.; Hidalgo, O.; Becker, A.; Elomaa, P. Evolutionary diversification of CYC/TB1-like TCP homologs and their recruitment for the control of branching and floral morphology in Papaveraceae (basal eudicots). New Phytol. 2018, 220, 317–331. [Google Scholar] [CrossRef]
- Pabon-Mora, N.; Madrigal, Y.; Alzate, J.F.; Ambrose, B.A.; Ferrandiz, C.; Wanke, S.; Neinhuis, C.; Gonzalez, F. Evolution of Class II TCP genes in perianth bearing Piperales and their contribution to the bilateral calyx in Aristolochia. New Phytol. 2020, 228, 752–769. [Google Scholar] [CrossRef]
- Zhang, W.; Steinmann, V.W.; Nikolov, L.; Kramer, E.M.; Davis, C.C. Divergent genetic mechanisms underlie reversals to radial floral symmetry from diverse zygomorphic flowered ancestors. Front. Plant Sci. 2013, 4, 302. [Google Scholar] [CrossRef]
- Berger, B.A.; Ricigliano, V.A.; Savriama, Y.; Lim, A.; Thompson, V.; Howarth, D.G. Geometric morphometrics reveals shifts in flower shape symmetry and size following gene knockdown of CYCLOIDEA and ANTHOCYANIDIN SYNTHASE. BMC Plant Biol. 2017, 17, 205. [Google Scholar] [CrossRef]
- Hileman, L.C. Trends in flower symmetry evolution revealed through phylogenetic and developmental genetic advances. Philos. Trans. R. Soc. B 2014, 369, 20130348. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, Y.; Igarashi, T.; Ohshima, M.; Shinoda, K.; Murata, N.; Kanno, A.; Nakano, M. Characterization of CYCLOIDEA-like genes in controlling floral zygomorphy in the monocotyledon Alstroemeria. Sci. Hortic. 2014, 169, 6–13. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).