

Hypoxic Preconditioned Neural Stem Cell-Derived Extracellular Vesicles Contain Distinct Protein Cargo from Their Normal Counterparts


Abstract
1. Introduction
2. Results
2.1. NSCs Characterization, Identification, and Differentiation
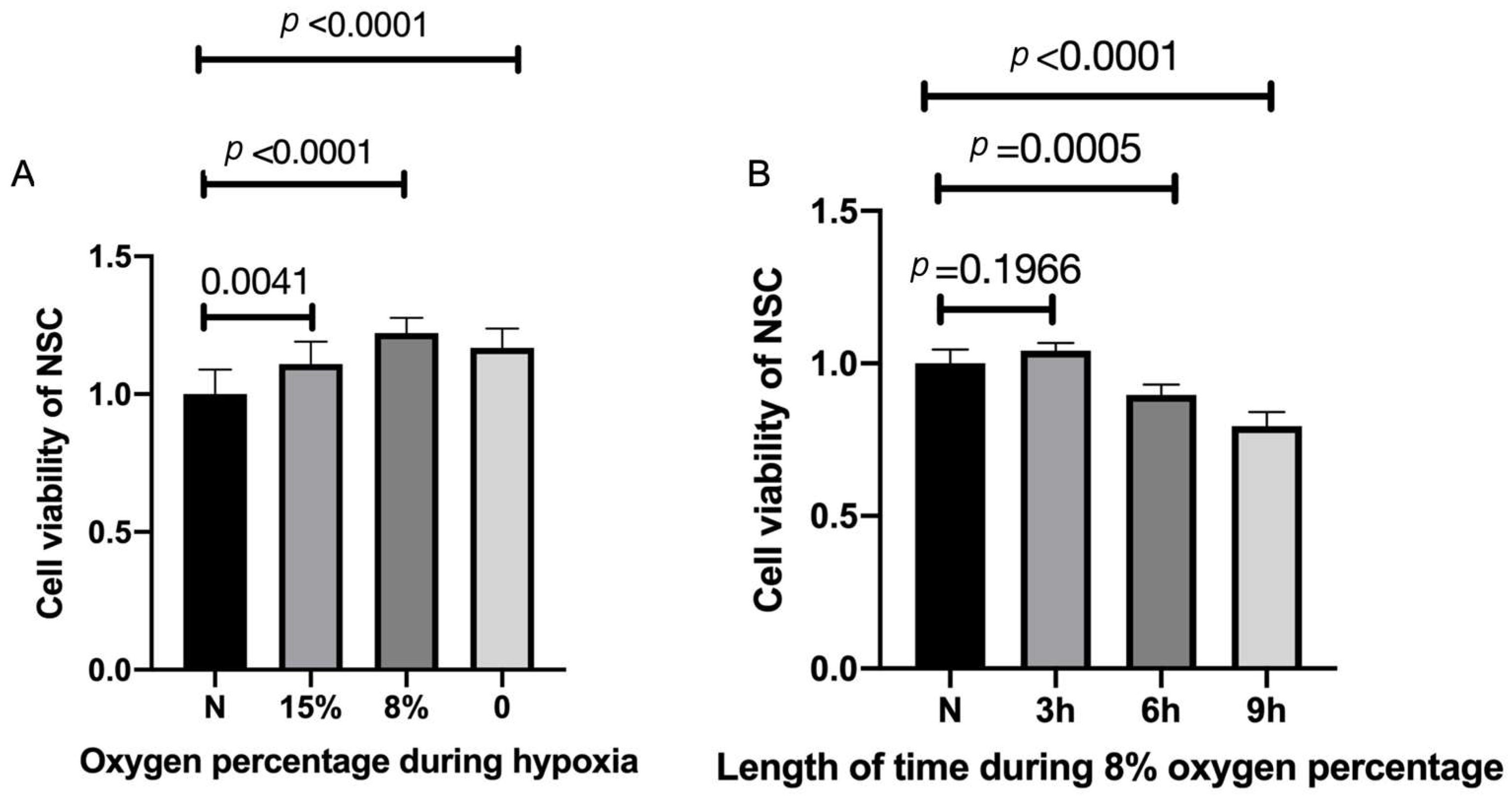
2.2. Hypoxic Preconditioning Affects NSC Viability
2.3. Characterization of Normal vs. Hypoxic Preconditioned NSC-Derived EVs
2.4. Proteomic Profiling of Normal vs. Hypoxic Preconditioned NSC-Derived EVs
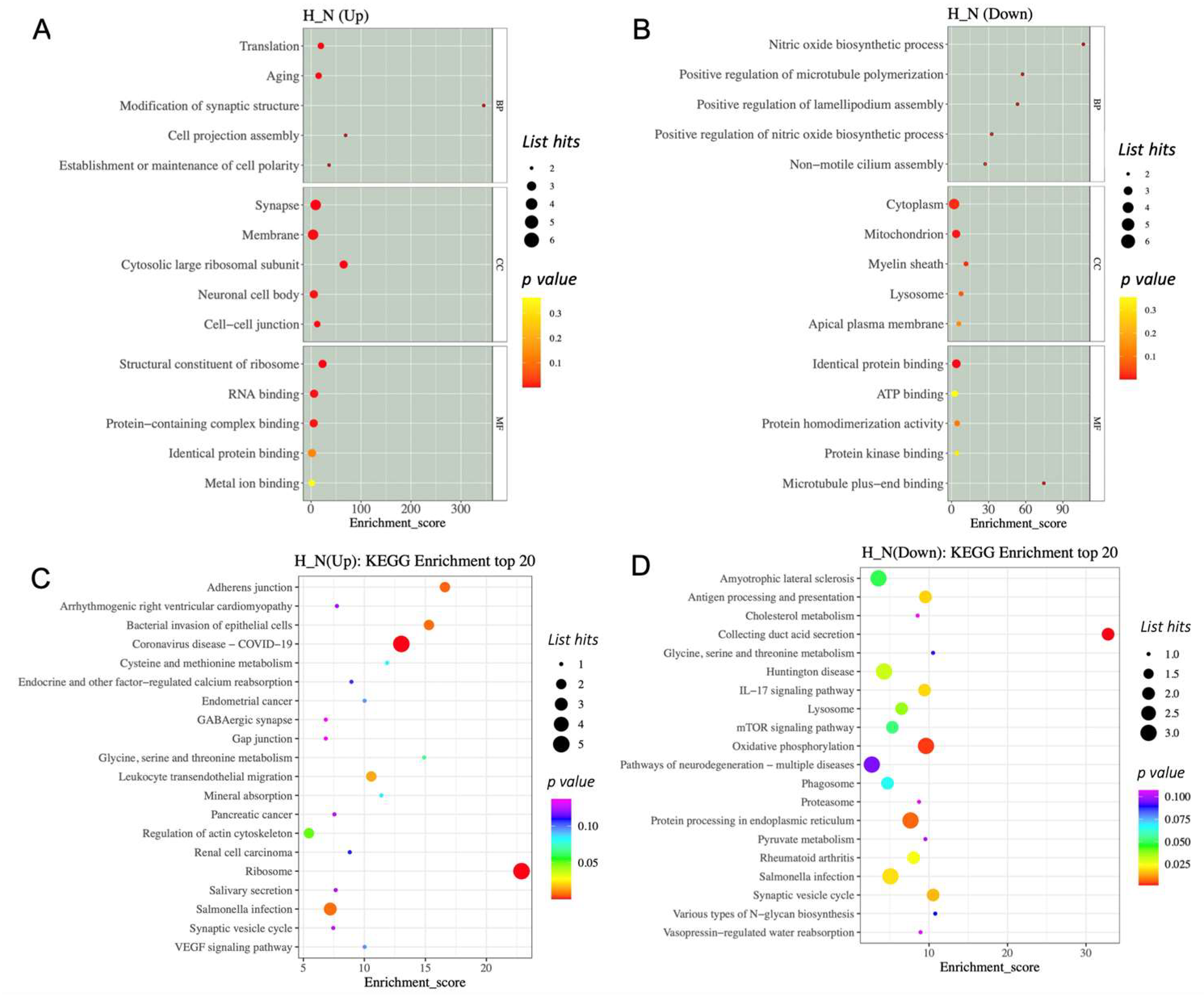
2.5. Differentially Expressed Proteins Enriched in Specific Biological Pathways
2.6. Detection of Elevated mRNA Transcripts of Upregulated Proteins within HP-EVs
3. Discussion
4. Methods and Materials
4.1. Primary NSC Isolation, Culture, and Passaging
4.2. Immunohistochemistry
4.3. Hypoxic Preconditioning
4.4. NSC Proliferation and Cell Viability Assay
4.5. Extracellular Vesicle Isolation
4.6. Characterization of EVs
4.7. Western Blotting
4.8. EV Analysis with ExoView
4.9. Extracellular Vesicle Proteome Profiling
4.10. EV RNA Isolation
4.11. RT-qPCR
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Boese, A.C.; Le, Q.E.; Pham, D.; Hamblin, M.H.; Lee, J.P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem. Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Sart, S.; Ma, T.; Li, Y. Preconditioning stem cells for in vivo delivery. BioRes. Open Access 2014, 3, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Sakata, H.; Niizuma, K.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Katsu, M.; Narasimhan, P.; Maier, C.M.; Nishiyama, Y.; Chan, P.H. Minocycline-preconditioned neural stem cells enhance neuroprotection after ischemic stroke in rats. J. Neurosci. 2012, 32, 3462–3473. [Google Scholar] [CrossRef]
- Sakata, H.; Narasimhan, P.; Niizuma, K.; Maier, C.M.; Wakai, T.; Chan, P.H. Interleukin 6-preconditioned neural stem cells reduce ischaemic injury in stroke mice. Brain 2012, 135 Pt 11, 3298–3310. [Google Scholar] [CrossRef]
- Theus, M.H.; Wei, L.; Cui, L.; Francis, K.; Hu, X.; Keogh, C.; Yu, S.P. In vitro hypoxic preconditioning of embryonic stem cells as a strategy of promoting cell survival and functional benefits after transplantation into the ischemic rat brain. Exp. Neurol. 2008, 210, 656–670. [Google Scholar] [CrossRef]
- D’Ascenzo, F.; Femmino, S.; Ravera, F.; Angelini, F.; Caccioppo, A.; Franchin, L.; Grosso, A.; Comita, S.; Cavallari, C.; Penna, C.; et al. Extracellular vesicles from patients with Acute Coronary Syndrome impact on ischemia-reperfusion injury. Pharmacol. Res. 2021, 170, 105715. [Google Scholar] [CrossRef] [PubMed]
- Francis, K.R.; Wei, L. Human embryonic stem cell neural differentiation and enhanced cell survival promoted by hypoxic preconditioning. Cell Death Dis. 2010, 1, e22. [Google Scholar] [CrossRef]
- Wakai, T.; Narasimhan, P.; Sakata, H.; Wang, E.; Yoshioka, H.; Kinouchi, H.; Chan, P.H. Hypoxic preconditioning enhances neural stem cell transplantation therapy after intracerebral hemorrhage in mice. J. Cereb. Blood Flow Metab. 2016, 36, 2134–2145. [Google Scholar] [CrossRef]
- Mung, K.L.; Tsui, Y.P.; Tai, E.W.; Chan, Y.S.; Shum, D.K.; Shea, G.K. Rapid and efficient generation of neural progenitors from adult bone marrow stromal cells by hypoxic preconditioning. Stem. Cell Res. Ther. 2016, 7, 146. [Google Scholar] [CrossRef]
- Sun, C.; Fu, J.; Qu, Z.; Li, D.; Si, P.; Qiao, Q.; Zhang, W.; Xue, Y.; Zhen, J.; Wang, W. Chronic mild hypoxia promotes hippocampal neurogenesis involving Notch1 signaling in epileptic rats. Brain Res. 2019, 1714, 88–98. [Google Scholar] [CrossRef]
- Huang, L.; Wan, Y.; Dang, Z.; Yang, P.; Yang, Q.; Wu, S. Hypoxic preconditioning ameliorated neuronal injury after middle cerebral artery occlusion by promoting neurogenesis. Brain Behav. 2020, 10, e01804. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; De Montpellier, S. Hypoxic-preconditioning enhances the regenerative capacity of neural stem/progenitors in subventricular zone of newborn piglet brain. Stem. Cell Res. 2013, 11, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.L.; Guan, Y.J.; Ma, D.D.; Du, H.M. Optimal concentration and time window for proliferation and differentiation of neural stem cells from embryonic cerebral cortex: 5% oxygen preconditioning for 72 hours. Neural Regen. Res. 2015, 10, 1516–1522. [Google Scholar] [CrossRef]
- Fan, W.L.; Liu, P.; Wang, G.; Pu, J.G.; Xue, X.; Zhao, J.H. Transplantation of hypoxic preconditioned neural stem cells benefits functional recovery via enhancing neurotrophic secretion after spinal cord injury in rats. J. Cell. Biochem. 2018, 119, 4339–4351. [Google Scholar] [CrossRef] [PubMed]
- Motomura, A.; Shimizu, M.; Kato, A.; Motomura, K.; Yamamichi, A.; Koyama, H.; Ohka, F.; Nishikawa, T.; Nishimura, Y.; Hara, M.; et al. Remote ischemic preconditioning protects human neural stem cells from oxidative stress. Apoptosis 2017, 22, 1353–1361. [Google Scholar] [CrossRef]
- Gharbi, T.; Zhang, Z.; Yang, G.Y. The Function of Astrocyte Mediated Extracellular Vesicles in Central Nervous System Diseases. Front. Cell Dev. Biol. 2020, 8, 568889. [Google Scholar] [CrossRef]
- Rooj, A.K.; Mineo, M.; Godlewski, J. MicroRNA and extracellular vesicles in glioblastoma: Small but powerful. Brain Tumor Pathol. 2016, 33, 77–88. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, L.; Wang, Y. Enhanced differentiation of neural stem cells to neurons and promotion of neurite outgrowth by oxygen-glucose deprivation. Int. J. Dev. Neurosci. 2015, 43, 50–57. [Google Scholar] [CrossRef]
- Wei, Z.Z.; Zhu, Y.B.; Zhang, J.Y.; McCrary, M.R.; Wang, S.; Zhang, Y.B.; Yu, S.P.; Wei, L. Priming of the Cells: Hypoxic Preconditioning for Stem Cell Therapy. Chin. Med. J. 2017, 130, 2361–2374. [Google Scholar] [PubMed]
- De Filippis, L.; Delia, D. Hypoxia in the regulation of neural stem cells. Cell. Mol. Life Sci. 2011, 68, 2831–2844. [Google Scholar] [CrossRef]
- Abati, E.; Bresolin, N.; Comi, G.P.; Corti, S. Preconditioning and Cellular Engineering to Increase the Survival of Transplanted Neural Stem Cells for Motor Neuron Disease Therapy. Mol. Neurobiol. 2019, 56, 3356–3367. [Google Scholar] [CrossRef] [PubMed]
- Bister, N.; Pistono, C.; Huremagic, B.; Jolkkonen, J.; Giugno, R.; Malm, T. Hypoxia and extracellular vesicles: A review on methods, vesicular cargo and functions. J. Extracell. Vesicles 2020, 10, e12002. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Hsiang, F.; Chang, J.H.; Yao, X.Q.; Zhao, H.; Zou, H.Y.; Wang, L.; Zhang, Q.X. Houshiheisan and its components promote axon regeneration after ischemic brain injury. Neural Regen. Res. 2018, 13, 1195–1203. [Google Scholar]
- Yang, Y.; Zhang, K.; Chen, X.; Wang, J.; Lei, X.; Zhong, J.; Xian, J.; Quan, Y.; Lu, Y.; Huang, Q.; et al. SVCT2 Promotes Neural Stem/Progenitor Cells Migration Through Activating CDC42 After Ischemic Stroke. Front Cell Neurosci. 2019, 13, 429. [Google Scholar] [CrossRef]
- Liu, J.; Chen, M.; Dong, R.; Sun, C.; Li, S.; Zhu, S. Ghrelin Promotes Cortical Neurites Growth in Late Stage After Oxygen-Glucose Deprivation/Reperfusion Injury. J. Mol. Neurosci. 2019, 68, 29–37. [Google Scholar] [CrossRef]
- Iguchi, Y.; Katsuno, M.; Niwa, J.I.; Yamada, S.I.; Sone, J.; Waza, M.; Adachi, H.; Tanaka, F.; Nagata, K.I.; Arimura, N.; et al. TDP-43 depletion induces neuronal cell damage through dysregulation of Rho family GTPases. J. Biol. Chem. 2009, 284, 22059–22066. [Google Scholar] [CrossRef]
- Huang, L.Y.; Ma, J.Y.; Song, J.X.; Xu, J.J.; Hong, R.; Fan, H.D.; Cai, H.; Wang, W.; Wang, Y.L.; Hu, Z.L.; et al. Ischemic accumulation of succinate induces Cdc42 succinylation and inhibits neural stem cell proliferation after cerebral ischemia/reperfusion. Neural Regen. Res. 2023, 18, 1040–1045. [Google Scholar]
- Endo, M.; Cerione, R.A. The brain-specific splice variant of the CDC42 GTPase works together with the kinase ACK to downregulate the EGF receptor in promoting neurogenesis. J. Biol. Chem. 2022, 298, 102564. [Google Scholar] [CrossRef]
- Rahman, S.U.; Kim, W.J.; Chung, S.H.; Woo, K.M. Nanofibrous topography-driven altered responsiveness to Wnt5a mediates the three-dimensional polarization of odontoblasts. Mater. Today Bio 2022, 17, 100479. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Matsui, T.K.; Kondo, J.; Kuwako, K.I. N-WASP-Arp2/3 signaling controls multiple steps of dendrite maturation in Purkinje cells in vivo. Development 2022, 149, dev201214. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Chen, F.Z.; Chen, W.; Zhu, H.M.; Chen, Y.; Li, Z.L.; Yan, F.; Liu, Z.Y.; Dong, W.R.; Zhang, L.; et al. Cdc42 Promotes Axonogenesis of Primary Hippocampal Neurons by Inhibiting Glycogen Synthase Kinase-3beta. J. Integr. Neurosci. 2022, 21, 133. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, P.; Puschel, A.W. An isoform-specific function of Cdc42 in regulating mammalian Exo70 during axon formation. Life Sci. Alliance 2023, 6, e202201722. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ye, J.; Zhang, X.; Meng, K. Longitudinal Variations of CDC42 in Patients with Acute Ischemic Stroke during 3-Year Period: Correlation with CD4(+) T Cells, Disease Severity, and Prognosis. Front. Neurol. 2022, 13, 848933. [Google Scholar] [CrossRef]
- Rahimi, M.J.; Urban, N.; Wegler, M.; Sticht, H.; Schaefer, M.; Popp, B.; Gaunitz, F.; Morleo, M.; Nigro, V.; Maitz, S.; et al. De novo variants in ATP2B1 lead to neurodevelopmental delay. Am. J. Hum. Genet. 2022, 109, 944–952. [Google Scholar] [CrossRef]
- Linseman, D.A.; Laessig, T.; Meintzer, M.K.; McClure, M.; Barth, H.; Aktories, K.; Heidenreich, K.A. An essential role for Rac/Cdc42 GTPases in cerebellar granule neuron survival. J. Biol. Chem. 2001, 276, 39123–39131. [Google Scholar] [CrossRef]
- Govek, E.E.; Newey, S.E.; Van Aelst, L. The role of the Rho GTPases in neuronal development. Genes Dev. 2005, 19, 1–49. [Google Scholar] [CrossRef]
- Kalpachidou, T.; Spiecker, L.; Kress, M.; Quarta, S. Rho GTPases in the Physiology and Pathophysiology of Peripheral Sensory Neurons. Cells 2019, 8, 591. [Google Scholar] [CrossRef]
- Stankiewicz, T.R.; Linseman, D.A. Rho family GTPases: Key players in neuronal development, neuronal survival, and neurodegeneration. Front. Cell Neurosci. 2014, 8, 314. [Google Scholar] [CrossRef]
- Rojo, D.R.; Prough, D.S.; Falduto, M.T.; Boone, D.R.; Micci, M.A.; Kahrig, K.M.; Crookshanks, J.M.; Jimenez, A.; Uchida, T.; Cowart, J.C.; et al. Influence of stochastic gene expression on the cell survival rheostat after traumatic brain injury. PLoS ONE 2011, 6, e23111. [Google Scholar] [CrossRef] [PubMed]
- Krey, J.F.; Dolmetsch, R.E. Molecular mechanisms of autism: A possible role for Ca2+ signaling. Curr. Opin. Neurobiol. 2007, 17, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Verpelli, C.; Dvoretskova, E.; Vicidomini, C.; Rossi, F.; Chiappalone, M.; Schoen, M.; Di Stefano, B.; Mantegazza, R.; Broccoli, V.; Bockers, T.M.; et al. Importance of Shank3 protein in regulating metabotropic glutamate receptor 5 (mGluR5) expression and signaling at synapses. J. Biol. Chem. 2011, 286, 34839–34850. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim-Verbaas, C.A.; Bressler, J.; Debette, S.; Schuur, M.; Smith, A.V.; Bis, J.C.; Davies, G.; Trompet, S.; Smith, J.A.; Wolf, C.; et al. GWAS for executive function and processing speed suggests involvement of the CADM2 gene. Mol. Psychiatry 2016, 21, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhao, J.; Chen, C.; Xu, J.; Bell, R.L.; Hall, F.S.; Koob, G.F.; Volkow, N.D.; Qing, H.; Lin, Z. Epistatic evidence for gender-dependant slow neurotransmission signalling in substance use disorders: PPP1R12B versus PPP1R1B. EBioMedicine 2020, 61, 103066. [Google Scholar] [CrossRef] [PubMed]
- Frei, J.A.; Andermatt, I.; Gesemann, M.; Stoeckli, E.T. The SynCAM synaptic cell adhesion molecules are involved in sensory axon pathfinding by regulating axon-axon contacts. J. Cell Sci. 2014, 127 Pt 24, 5288–5302. [Google Scholar] [CrossRef] [PubMed]
- Pellissier, F.; Gerber, A.; Bauer, C.; Ballivet, M.; Ossipow, V. The adhesion molecule Necl-3/SynCAM-2 localizes to myelinated axons, binds to oligodendrocytes and promotes cell adhesion. BMC Neurosci. 2007, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.Q.; Lu, Y.P.; Sun, H.L.; Gao, L.N.; Song, P.P.; Feng, Z.J.; You, C.G. Sex Difference of Ribosome in Stroke-Induced Peripheral Immunosuppression by Integrated Bioinformatics Analysis. BioMed Res. Int. 2020, 2020, 3650935. [Google Scholar] [CrossRef]
- Wei, X.Y.; Zhang, T.Q.; Suo, R.; Qu, Y.Y.; Chen, Y.; Zhu, Y.L. Long non-coding RNA RPL34-AS1 ameliorates oxygen-glucose deprivation-induced neuronal injury via modulating miR-223-3p/IGF1R axis. Hum. Cell 2022, 35, 1785–1796. [Google Scholar] [CrossRef]
- Ma, Q.; Zhang, L. C-type natriuretic peptide functions as an innate neuroprotectant in neonatal hypoxic-ischemic brain injury in mouse via natriuretic peptide receptor 2. Exp. Neurol. 2018, 304, 58–66. [Google Scholar] [CrossRef]
- Sun, F.; Zhou, K.; Tian, K.Y.; Zhang, X.Y.; Liu, W.; Wang, J.; Zhong, C.P.; Qiu, J.H.; Zha, D.J. Atrial Natriuretic Peptide Promotes Neurite Outgrowth and Survival of Cochlear Spiral Ganglion Neurons in vitro through NPR-A/cGMP/PKG Signaling. Front. Cell Dev. Biol. 2021, 9, 681421. [Google Scholar] [CrossRef] [PubMed]
- Haan, N.; Westacott, L.J.; Carter, J.; Owen, M.J.; Gray, W.P.; Hall, J.; Wilkinson, L.S. Haploinsufficiency of the schizophrenia and autism risk gene Cyfip1 causes abnormal postnatal hippocampal neurogenesis through microglial and Arp2/3 mediated actin dependent mechanisms. Transl. Psychiatry 2021, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Habela, C.W.; Yoon, K.J.; Kim, N.S.; Taga, A.; Bell, K.; Bergles, D.E.; Maragakis, N.J.; Ming, G.L.; Song, H. Persistent Cyfip1 Expression Is Required to Maintain the Adult Subventricular Zone Neurogenic Niche. J. Neurosci. 2020, 40, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.; Harony-Nicolas, H.; Buxbaum, J.D.; Bozdagi-Gunal, O.; Benson, D.L. Cyfip1 Regulates Presynaptic Activity during Development. J. Neurosci. 2016, 36, 1564–1576. [Google Scholar] [CrossRef]
- Yoon, K.J.; Nguyen, H.N.; Ursini, G.; Zhang, F.; Kim, N.S.; Wen, Z.; Makri, G.; Nauen, D.; Shin, J.H.; Park, Y.; et al. Modeling a genetic risk for schizophrenia in iPSCs and mice reveals neural stem cell deficits associated with adherens junctions and polarity. Cell Stem Cell 2014, 15, 79–91. [Google Scholar] [CrossRef]
- Biembengut, I.V.; Silva, I.L.Z.; Souza, T.; Shigunov, P. Cytoplasmic FMR1 interacting protein (CYFIP) family members and their function in neural development and disorders. Mol. Biol. Rep. 2021, 48, 6131–6143. [Google Scholar] [CrossRef]
- Kim, G.H.; Zhang, Y.; Kang, H.R.; Lee, S.H.; Shin, J.; Lee, C.H.; Kang, H.; Ma, R.; Jin, C.; Kim, Y.; et al. Altered presynaptic function and number of mitochondria in the medial prefrontal cortex of adult Cyfip2 heterozygous mice. Mol. Brain 2020, 13, 123. [Google Scholar] [CrossRef]
- Atasoy, D.; Schoch, S.; Ho, A.; Nadasy, K.A.; Liu, X.; Zhang, W.; Mukherjee, K.; Nosyreva, E.D.; Fernandez-Chacon, R.; Missler, M.; et al. Deletion of CASK in mice is lethal and impairs synaptic function. Proc. Natl. Acad. Sci. USA 2007, 104, 2525–2530. [Google Scholar] [CrossRef]
- Mori, T.; Kasem, E.A.; Suzuki-Kouyama, E.; Cao, X.; Li, X.; Kurihara, T.; Uemura, T.; Yanagawa, T.; Tabuchi, K. Deficiency of calcium/calmodulin-dependent serine protein kinase disrupts the excitatory-inhibitory balance of synapses by down-regulating GluN2B. Mol. Psychiatry 2019, 24, 1079–1092. [Google Scholar] [CrossRef]
- Wu, X.; Cai, Q.; Chen, Y.; Zhu, S.; Mi, J.; Wang, J.; Zhang, M. Structural Basis for the High-Affinity Interaction between CASK and Mint1. Structure 2020, 28, 664–673.e3. [Google Scholar] [CrossRef]
- Ngo, L.; Haas, M.; Qu, Z.; Li, S.S.; Zenker, J.; Teng, K.S.; Gunnersen, J.M.; Breuss, M.; Habgood, M.; Keays, D.A.; et al. TUBB5 and its disease-associated mutations influence the terminal differentiation and dendritic spine densities of cerebral cortical neurons. Hum. Mol. Genet. 2014, 23, 5147–5158. [Google Scholar] [CrossRef] [PubMed]
- Breuss, M.; Heng, J.I.; Poirier, K.; Tian, G.; Jaglin, X.H.; Qu, Z.; Braun, A.; Gstrein, T.; Ngo, L.; Haas, M.; et al. Mutations in the beta-tubulin gene TUBB5 cause microcephaly with structural brain abnormalities. Cell Rep. 2012, 2, 1554–1562. [Google Scholar] [CrossRef]
- Walkley, S.U.; Suzuki, K. Consequences of NPC1 and NPC2 loss of function in mammalian neurons. Biochim. Biophys. Acta 2004, 1685, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Hetmanczyk-Sawicka, K.; Iwanicka-Nowicka, R.; Fogtman, A.; Ciesla, J.; Wlodarski, P.; Zyzynska-Granica, B.; Filocamo, M.; Dardis, A.; Peruzzo, P.; Bednarska-Makaruk, M.; et al. Changes in global gene expression indicate disordered autophagy, apoptosis and inflammatory processes and downregulation of cytoskeletal signalling and neuronal development in patients with Niemann-Pick C disease. Neurogenetics 2020, 21, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Ratovitski, T.; Chighladze, E.; Waldron, E.; Hirschhorn, R.R.; Ross, C.A. Cysteine proteases bleomycin hydrolase and cathepsin Z mediate N-terminal proteolysis and toxicity of mutant huntingtin. J. Biol. Chem. 2011, 286, 12578–12589. [Google Scholar] [CrossRef]
- Suszynska-Zajczyk, J.; Luczak, M.; Marczak, L.; Jakubowski, H. Hyperhomocysteinemia and bleomycin hydrolase modulate the expression of mouse brain proteins involved in neurodegeneration. J. Alzheimers Dis. 2014, 40, 713–726. [Google Scholar] [CrossRef]
- Luo, Y.; Yue, W.; Quan, X.; Wang, Y.; Zhao, B.; Lu, Z. Asymmetric dimethylarginine exacerbates Abeta-induced toxicity and oxidative stress in human cell and Caenorhabditis elegans models of Alzheimer disease. Free Radic. Biol. Med. 2015, 79, 117–126. [Google Scholar] [CrossRef]
- Uyeda, A.; Quan, L.; Kato, Y.; Muramatsu, N.; Tanabe, S.; Sakai, K.; Ichinohe, N.; Kawahara, Y.; Suzuki, T.; Muramatsu, R. Dimethylarginine dimethylaminohydrolase 1 as a novel regulator of oligodendrocyte differentiation in the central nervous system remyelination. Glia 2021, 69, 2591–2604. [Google Scholar] [CrossRef]
- Lopez-Toledo, G.; Silva-Lucero, M.D.; Herrera-Diaz, J.; Garcia, D.E.; Arias-Montano, J.A.; Cardenas-Aguayo, M.D. Patient-Derived Fibroblasts with Presenilin-1 Mutations, That Model Aspects of Alzheimer’s Disease Pathology, Constitute a Potential Object for Early Diagnosis. Front. Aging Neurosci. 2022, 14, 921573. [Google Scholar] [CrossRef]
- Shim, J.S.; Song, M.Y.; Yim, S.V.; Lee, S.E.; Park, K.S. Global analysis of ginsenoside Rg1 protective effects in beta-amyloid-treated neuronal cells. J. Ginseng Res. 2017, 41, 566–571. [Google Scholar] [CrossRef]
- Cassano, T.; Giamogante, F.; Calcagnini, S.; Romano, A.; Lavecchia, A.M.; Inglese, F.; Paglia, G.; Bukke, V.N.; Romano, A.D.; Friuli, M.; et al. PDIA3 Expression Is Altered in the Limbic Brain Regions of Triple-Transgenic Mouse Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 3005. [Google Scholar] [CrossRef] [PubMed]
- Mendsaikhan, A.; Tooyama, I.; Serrano, G.E.; Beach, T.G.; Walker, D.G. Loss of Lysosomal Proteins Progranulin and Prosaposin Associated with Increased Neurofibrillary Tangle Development in Alzheimer Disease. J. Neuropathol. Exp. Neurol. 2021, 80, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Callizot, N.; Estrella, C.; Burlet, S.; Henriques, A.; Brantis, C.; Barrier, M.; Campanari, M.L.; Verwaerde, P. AZP2006, a new promising treatment for Alzheimer’s and related diseases. Sci. Rep. 2021, 11, 16806. [Google Scholar] [CrossRef] [PubMed]
- Mendsaikhan, A.; Tooyama, I.; Bellier, J.P.; Serrano, G.E.; Sue, L.I.; Lue, L.F.; Beach, T.G.; Walker, D.G. Characterization of lysosomal proteins Progranulin and Prosaposin and their interactions in Alzheimer’s disease and aged brains: Increased levels correlate with neuropathology. Acta Neuropathol. Commun. 2019, 7, 215. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, L.; Bracko, O.; Choi, J.W.; Jia, Y.; Nana, A.L.; Brady, O.A.; Hernandez, J.C.C.; Nishimura, N.; Seeley, W.W.; et al. Impaired prosaposin lysosomal trafficking in frontotemporal lobar degeneration due to progranulin mutations. Nat. Commun. 2017, 8, 15277. [Google Scholar] [CrossRef]
- Wang, L.; Chen, J.; Hu, Y.; Liao, A.; Zheng, W.; Wang, X.; Lan, J.; Shen, J.; Wang, S.; Yang, F.; et al. Progranulin improves neural development via the PI3K/Akt/GSK-3beta pathway in the cerebellum of a VPA-induced rat model of ASD. Transl. Psychiatry 2022, 12, 114. [Google Scholar] [CrossRef]
- Sikora, J.; Harzer, K.; Elleder, M. Neurolysosomal pathology in human prosaposin deficiency suggests essential neurotrophic function of prosaposin. Acta Neuropathol. 2007, 113, 163–175. [Google Scholar] [CrossRef]
- Troncoso, M.; Bannoud, N.; Carvelli, L.; Asensio, J.; Seltzer, A.; Sosa, M.A. Hypoxia-ischemia alters distribution of lysosomal proteins in rat cortex and hippocampus. Biol. Open 2018, 7, bio036723. [Google Scholar] [CrossRef]
- Martinez, B.; Peplow, P.V. MicroRNAs in Parkinson’s disease and emerging therapeutic targets. Neural Regen. Res. 2017, 12, 1945–1959. [Google Scholar]
- Oji, Y.; Hatano, T.; Ueno, S.I.; Funayama, M.; Ishikawa, K.I.; Okuzumi, A.; Noda, S.; Sato, S.; Satake, W.; Toda, T.; et al. Variants in saposin D domain of prosaposin gene linked to Parkinson’s disease. Brain 2020, 143, 1190–1205. [Google Scholar] [CrossRef]
- Araki, E.; Tsuboi, Y.; Daechsel, J.; Milnerwood, A.; Vilarino-Guell, C.; Fujii, N.; Mishima, T.; Oka, T.; Hara, H.; Fukae, J.; et al. A novel DCTN1 mutation with late-onset parkinsonism and frontotemporal atrophy. Mov. Disord. 2014, 29, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; You, P.; Yu, H.; Su, B. REEP1 Preserves Motor Function in SOD1(G93A) Mice by Improving Mitochondrial Function via Interaction with NDUFA4. Neurosci. Bull. 2022, 1–18. [Google Scholar] [CrossRef]
- Finsterer, J.; Burgunder, J.M. Recent progress in the genetics of motor neuron disease. Eur. J. Med. Genet. 2014, 57, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Henty-Ridilla, J.L. Multiple roles for the cytoskeleton in ALS. Exp. Neurol. 2022, 355, 114143. [Google Scholar] [CrossRef]
- Castellanos-Montiel, M.J.; Chaineau, M.; Durcan, T.M. The Neglected Genes of ALS: Cytoskeletal Dynamics Impact Synaptic Degeneration in ALS. Front. Cell Neurosci. 2020, 14, 594975. [Google Scholar] [CrossRef]
- Wada, Y.; Nagai, A.; Sheikh, A.M.; Onoda, K.; Terashima, M.; Shiota, Y.; Araki, A.; Yamaguchi, S. Co-localization of cystatin C and prosaposin in cultured neurons and in anterior horn neurons with amyotrophic lateral sclerosis. J. Neurol. Sci. 2018, 384, 67–74. [Google Scholar] [CrossRef]
- Gustavsson, E.K.; Trinh, J.; Guella, I.; Szu-Tu, C.; Khinda, J.; Lin, C.H.; Wu, R.M.; Stoessl, J.; Appel-Cresswell, S.; McKeown, M.; et al. DCTN1 p.K56R in progressive supranuclear palsy. Park. Relat. Disord. 2016, 28, 56–61. [Google Scholar] [CrossRef]
- Deshimaru, M.; Kinoshita-Kawada, M.; Kubota, K.; Watanabe, T.; Tanaka, Y.; Hirano, S.; Ishidate, F.; Hiramoto, M.; Ishikawa, M.; Uehara, Y.; et al. DCTN1 Binds to TDP-43 and Regulates TDP-43 Aggregation. Int. J. Mol. Sci. 2021, 22, 3985. [Google Scholar] [CrossRef]
- Mishima, T.; Fujioka, S.; Tomiyama, H.; Yabe, I.; Kurisaki, R.; Fujii, N.; Neshige, R.; Ross, O.A.; Farrer, M.J.; Dickson, D.W.; et al. Establishing diagnostic criteria for Perry syndrome. J. Neurol. Neurosurg. Psychiatry 2018, 89, 482–487. [Google Scholar] [CrossRef]
- Deshimaru, M.; Mishima, T.; Watanabe, T.; Kubota, K.; Hosoi, M.; Kinoshita-Kawada, M.; Yuasa-Kawada, J.; Ikeda, M.; Mori, M.; Murata, Y.; et al. Behavioral profile in a Dctn1(G71A) knock-in mouse model of Perry disease. Neurosci. Lett. 2021, 764, 136234. [Google Scholar] [CrossRef]
- Honda, H.; Sasagasako, N.; Shen, C.; Shijo, M.; Hamasaki, H.; Suzuki, S.O.; Tsuboi, Y.; Fujii, N.; Iwaki, T. DCTN1 F52L mutation case of Perry syndrome with progressive supranuclear palsy-like tauopathy. Park. Relat. Disord. 2018, 51, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, A.; Sekiya, M.; Beckmann, N.D.; Quan, X.; Schrode, N.; Fernando, M.B.; Yu, A.; Zhu, L.; Cao, J.; et al. Transformative Network Modeling of Multi-omics Data Reveals Detailed Circuits, Key Regulators, and Potential Therapeutics for Alzheimer’s Disease. Neuron 2021, 109, 257–272.e14. [Google Scholar] [CrossRef] [PubMed]
- Dulski, J.; Konno, T.; Wszolek, Z. DCTN1-Related Neurodegeneration. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Wang, N.; Ma, Q.; Peng, P.; Yu, Y.; Xu, S.; Wang, G.; Ying, Z.; Wang, H. Autophagy and Ubiquitin-Proteasome System Coordinate to Regulate the Protein Quality Control of Neurodegenerative Disease-Associated DCTN1. Neurotox. Res. 2020, 37, 48–57. [Google Scholar] [CrossRef]
- Yoneshige, A.; Suzuki, K.; Suzuki, K.; Matsuda, J. A mutation in the saposin C domain of the sphingolipid activator protein (Prosaposin) gene causes neurodegenerative disease in mice. J. Neurosci. Res. 2010, 88, 2118–2134. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, M.; Dou, Y.; Du, K.; Liu, X.; Zhao, Y. DDAH1/ADMA Regulates Adiponectin Resistance in Cerebral Ischemia via the ROS/FOXO1/APR1 Pathway. Oxid. Med. Cell. Longev. 2022, 2022, 2350857. [Google Scholar] [CrossRef]
- Fassio, A.; Esposito, A.; Kato, M.; Saitsu, H.; Mei, D.; Marini, C.; Conti, V.; Nakashima, M.; Okamoto, N.; Olmez Turker, A.; et al. De novo mutations of the ATP6V1A gene cause developmental encephalopathy with epilepsy. Brain 2018, 141, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Li, Y.; Li, R.; Lei, T.Y.; Wang, D.; Yang, X.; Han, J.; Pan, M.; Zhen, L.; Ou, Y.M.; et al. NDUFA4 enhances neuron growth by triggering growth factors and inhibiting neuron apoptosis through Bcl-2 and cytochrome C mediated signaling pathway. Am. J. Transl. Res. 2018, 10, 164–174. [Google Scholar]
- Yu, J.; Mu, J.; Guo, Q.; Yang, L.; Zhang, J.; Liu, Z.; Yu, B.; Zhang, T.; Xie, J. Transcriptomic profile analysis of mouse neural tube development by RNA-Seq. IUBMB Life 2017, 69, 706–719. [Google Scholar] [CrossRef]
- Guan, Z.; Liang, Y.; Wang, X.; Zhu, Z.; Yang, A.; Li, S.; Yu, J.; Niu, B.; Wang, J. Unraveling the Mechanisms of Clinical Drugs-Induced Neural Tube Defects Based on Network Pharmacology and Molecular Docking Analysis. Neurochem. Res. 2022, 47, 3709–3722. [Google Scholar] [CrossRef]
- Song, Y.; Li, Z.; He, T.; Qu, M.; Jiang, L.; Li, W.; Shi, X.; Pan, J.; Zhang, L.; Wang, Y.; et al. M2 microglia-derived exosomes protect the mouse brain from ischemia-reperfusion injury via exosomal miR-124. Theranostics 2019, 9, 2910–2923. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Up/Down-Regulated | Description | p-Value | FC |
|---|---|---|---|---|
| Ints4 | Up | Integrator complex subunit 4 | 0.01973179 | 4.61333333 |
| Rpl18 | Up | 60S ribosomal protein L18 | 0.03111818 | 2.18979266 |
| Moxd1 | Up | DBH-like monooxygenase protein 1 | 0.03943133 | 1.96772997 |
| Rpl7 | Up | 60S ribosomal protein L7 | 0.0180495 | 1.9379361 |
| Cadm2 | Up | Cell adhesion molecule 2 | 0.04979333 | 1.76720858 |
| Rps8 | Up | 40S ribosomal protein S8 | 0.0437712 | 1.69393939 |
| Tubb5 | Up | Tubulin beta-5 chain | 0.04878117 | 1.65571049 |
| Rpl34 | Up | 60S ribosomal protein L34 | 0.04677985 | 1.64879179 |
| Bhmt | Up | Betaine--homocysteine S-methyltransferase 1 | 0.04297693 | 1.63417847 |
| Cyfip1 | Up | Cytoplasmic FMR1-interacting protein 1 | 0.04924497 | 1.60110533 |
| Rpl7a | Up | 60S ribosomal protein L7a | 0.03507391 | 1.5979214 |
| Up | Uncharacterized protein C2orf72 | 0.0137668 | 1.45398773 | |
| Ctnna1 | Up | Catenin alpha-1 | 0.0172758 | 1.41868198 |
| Cdc42 | Up | Cell division control protein 42 | 0.02439533 | 1.41036457 |
| Adam9 | Up | Disintegrin and metalloproteinase domain-containing protein 9 | 0.02993997 | 1.3840882 |
| Atp2b1 | Up | Plasma membrane calcium-transporting ATPase 1 | 0.03146976 | 1.37318303 |
| Slc6a1 | Up | Sodium- and chloride-dependent GABA transporter 1 | 0.00137502 | 1.34094235 |
| Cnp | Up | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase | 0.01201357 | 1.28534704 |
| Nkain3 | Up | Sodium/potassium-transporting ATPase subunit beta-1-interacting protein 3 | 0.03002822 | 1.2425007 |
| Cask | Up | Peripheral plasma membrane protein CASK | 0.02686701 | 1.23588597 |
| Actr3 | Down | Actin-related protein 3 | 0.03585085 | 0.82959744 |
| Npc2 | Down | NPC intracellular cholesterol transporter 2 | 0.03261727 | 0.80202703 |
| Blmh | Down | Bleomycin hydrolase | 0.04554183 | 0.79856115 |
| Psph | Down | Phosphoserine phosphatase | 0.01691792 | 0.79636282 |
| Atp6v1a | Down | V-type proton ATPase catalytic subunit A | 0.01888445 | 0.78930888 |
| Mapre1 | Down | Microtubule-associated protein RP/EB family member 1 | 0.04938624 | 0.77795066 |
| Elavl1 | Down | ELAV-like protein 1 | 0.0289941 | 0.7691287 |
| Glo1 | Down | Lactoylglutathione lyase | 0.04542562 | 0.76422585 |
| Atp6v1c1 | Down | V-type proton ATPase subunit C 1 | 0.03806311 | 0.73310225 |
| Plbd2 | Down | Putative phospholipase B-like 2 | 0.00952826 | 0.73010381 |
| Ddah1 | Down | N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 | 0.01486318 | 0.72043011 |
| Hsp90aa1 | Down | Heat shock protein HSP 90-alpha | 0.0227131 | 0.70524297 |
| Psmb2 | Down | Proteasome subunit beta type-2 | 0.04881743 | 0.6939856 |
| Ndufa4 | Down | Cytochrome c oxidase subunit NDUFA4 | 0.03014898 | 0.65645061 |
| Dctn1 | Down | Dynactin subunit 1 | 0.02291716 | 0.65439504 |
| Oat | Down | Ornithine aminotransferase | 0.02557392 | 0.6400164 |
| Rpn2 | Down | Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 2 | 0.0427866 | 0.63445035 |
| Ak3 | Down | GTP:AMP phosphotransferase AK3 | 0.04882486 | 0.63211589 |
| Psap | Down | Prosaposin | 0.02450609 | 0.62423873 |
| Hnrnpab | Down | Heterogeneous nuclear ribonucleoprotein A/B | 0.03999623 | 0.61628612 |
| Pdia3 | Down | Protein disulfide-isomerase A3 | 0.0477473 | 0.57666535 |
| Actg2 | Down | Actin, gamma-enteric smooth muscle | 0.01623375 | 0.50677966 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gharbi, T.; Liu, C.; Khan, H.; Zhang, Z.; Yang, G.-Y.; Tang, Y. Hypoxic Preconditioned Neural Stem Cell-Derived Extracellular Vesicles Contain Distinct Protein Cargo from Their Normal Counterparts. Curr. Issues Mol. Biol. 2023, 45, 1982-1997. https://doi.org/10.3390/cimb45030127
Gharbi T, Liu C, Khan H, Zhang Z, Yang G-Y, Tang Y. Hypoxic Preconditioned Neural Stem Cell-Derived Extracellular Vesicles Contain Distinct Protein Cargo from Their Normal Counterparts. Current Issues in Molecular Biology. 2023; 45(3):1982-1997. https://doi.org/10.3390/cimb45030127
Chicago/Turabian StyleGharbi, Tahereh, Chang Liu, Haroon Khan, Zhijun Zhang, Guo-Yuan Yang, and Yaohui Tang. 2023. "Hypoxic Preconditioned Neural Stem Cell-Derived Extracellular Vesicles Contain Distinct Protein Cargo from Their Normal Counterparts" Current Issues in Molecular Biology 45, no. 3: 1982-1997. https://doi.org/10.3390/cimb45030127
APA StyleGharbi, T., Liu, C., Khan, H., Zhang, Z., Yang, G.-Y., & Tang, Y. (2023). Hypoxic Preconditioned Neural Stem Cell-Derived Extracellular Vesicles Contain Distinct Protein Cargo from Their Normal Counterparts. Current Issues in Molecular Biology, 45(3), 1982-1997. https://doi.org/10.3390/cimb45030127