
Influence of Breastfeeding on the State of Meta-Inflammation in Obesity—A Narrative Review
 , , ,
, , ,  and
and 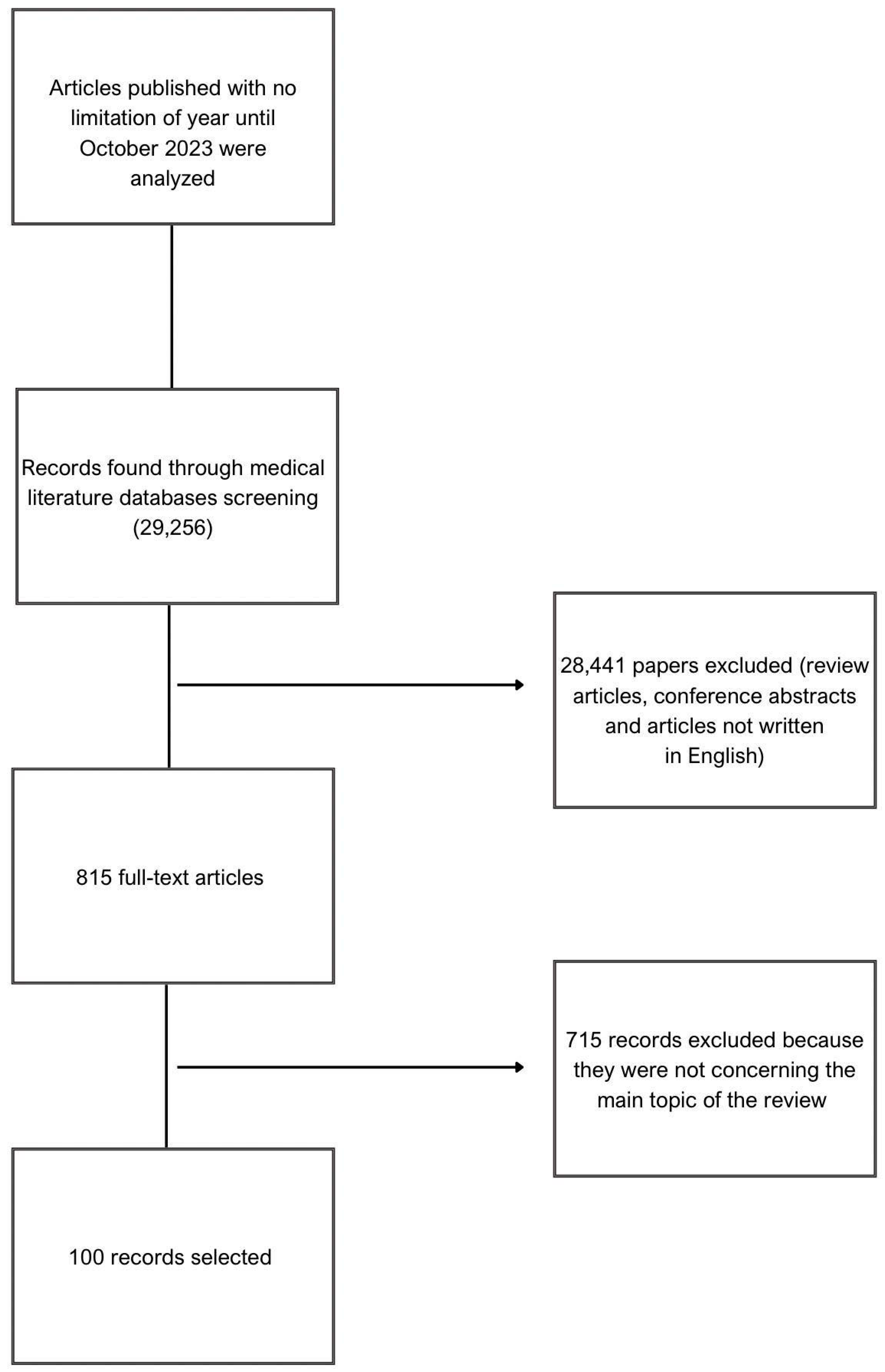{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
3. Meta-Inflammation in Obesity
4. Immunological Changes in the Postpartum Period
5. Breastfeeding and Metabolic Programming
6. Breastfeeding’s Impact on the Meta-Inflammatory Process
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Russo, S.; Kwiatkowski, M.; Govorukhina, N.; Bischoff, R.; Melgert, B.N. Meta-Inflammation and Metabolic Reprogramming of Macrophages in Diabetes and Obesity: The Importance of Metabolites. Front. Immunol. 2021, 12, 746151. [Google Scholar] [CrossRef] [PubMed]
- Pantham, P.; Aye, I.L.M.H.; Powell, T.L. Inflammation in Maternal Obesity and Gestational Diabetes Mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Mariona, F.G. Perspectives in Obesity and Pregnancy. Women’s Health 2016, 12, 523–532. [Google Scholar] [CrossRef]
- Catalano, P.M.; Shankar, K. Obesity and Pregnancy: Mechanisms of Short Term and Long Term Adverse Consequences for Mother and Child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef] [PubMed]
- Denizli, M.; Capitano, M.L.; Kua, K.L. Maternal Obesity and the Impact of Associated Early-Life Inflammation on Long-Term Health of Offspring. Front. Cell. Infect. Microbiol. 2022, 12, 940937. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, T.V.; Goldani, M.Z.; IVAPSA Group. Maternal Pre-Pregnancy Overweight/Obesity and Gestational Diabetes Interaction on Delayed Breastfeeding Initiation. PLoS ONE 2018, 13, e0194879. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Cotelo, M.D.C.; Movilla-Fernández, M.J.; Pita-García, P.; Arias, B.F.; Novío, S. Breastfeeding Knowledge and Relation to Prevalence. Rev. Esc. Enferm. USP 2019, 53, e03433. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Alshahrani, M.A.; Al-Thubaity, D.D.; Sayed, S.H.; Almedhesh, S.A.; Elgzar, W.T. Associated Factors of Exclusive Breastfeeding Intention among Pregnant Women in Najran, Saudi Arabia. Nutrients 2023, 15, 3051. [Google Scholar] [CrossRef]
- Wallenborn, J.T.; Levine, G.A.; Carreira Dos Santos, A.; Grisi, S.; Brentani, A.; Fink, G. Breastfeeding, Physical Growth, and Cognitive Development. Pediatrics 2021, 147, e2020008029. [Google Scholar] [CrossRef]
- Victora, C.G.; Bahl, R.; Barros, A.J.D.; França, G.V.A.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C. Breastfeeding in the 21st Century: Epidemiology, Mechanisms, and Lifelong Effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- Obesity and Overweight. Available online: https://www.who.int/En/News-Room/Fact-Sheets/Detail/Obesity-and-Overweight (accessed on 9 June 2023).
- Obesity in Pregnancy: ACOG Practice Bulletin, Number 230. Obstet. Gynecol. 2021, 137, e128–e144. [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation, and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory Mechanisms Linking Obesity and Metabolic Disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, T.; Ackerman, S.E.; Shen, L.; Engleman, E. Role of Innate and Adaptive Immunity in Obesity-Associated Metabolic Disease. J. Clin. Investg. 2017, 127, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Carobbio, S.; Vidal-Puig, A. Adipose Tissue Plasticity: How Fat Depots Respond Differently to Pathophysiological Cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef]
- Woo, C.-Y.; Jang, J.E.; Lee, S.E.; Koh, E.H.; Lee, K.-U. Mitochondrial Dysfunction in Adipocytes as a Primary Cause of Adipose Tissue Inflammation. Diabetes Metab. J. 2019, 43, 247. [Google Scholar] [CrossRef]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in Healthy Aging and Disease. Nat. Aging 2021, 1, 634–650. [Google Scholar] [CrossRef]
- Gan, L.; Liu, Z.; Luo, D.; Ren, Q.; Wu, H.; Li, C.; Sun, C. Reduced Endoplasmic Reticulum Stress-Mediated Autophagy Is Required for Leptin Alleviating Inflammation in Adipose Tissue. Front. Immunol. 2017, 8, 1507. [Google Scholar] [CrossRef]
- Nepal, S.; Kim, M.J.; Hong, J.T.; Kim, S.H.; Sohn, D.-H.; Lee, S.H.; Song, K.; Choi, D.Y.; Lee, E.S.; Park, P.-H. Autophagy Induction by Leptin Contributes to Suppression of Apoptosis in Cancer Cells and Xenograft Model: Involvement of P53/FoxO3A Axis. Oncotarget 2015, 6, 7166–7181. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity Is Associated with Macrophage Accumulation in Adipose Tissue. J. Clin. Investg. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity Induces a Phenotypic Switch in Adipose Tissue Macrophage Polarization. J. Clin. Investg. 2007, 117, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Gollinger, K.; Kriehuber, E.; Kiefer, F.W.; Neuhofer, A.; Stulnig, T.M. Newly Identified Adipose Tissue Macrophage Populations in Obesity with Distinct Chemokine and Chemokine Receptor Expression. Int. J. Obes. 2010, 34, 1684–1694. [Google Scholar] [CrossRef] [PubMed]
- Castoldi, A.; Naffah De Souza, C.; Câmara, N.O.S.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2016, 6, 637. [Google Scholar] [CrossRef]
- Boscá, L.; González-Ramos, S.; Prieto, P.; Fernández-Velasco, M.; Mojena, M.; Martín-Sanz, P.; Alemany, S. Metabolic Signatures Linked to Macrophage Polarization: From Glucose Metabolism to Oxidative Phosphorylation. Biochem. Soc. Trans. 2015, 43, 740–744. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose Expression of Tumor Necrosis Factor-α: Direct Role in Obesity-Linked Insulin Resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- O’Rourke, R.W.; Metcalf, M.D.; White, A.E.; Madala, A.; Winters, B.R.; Maizlin, I.I.; Jobe, B.A.; Roberts, C.T.; Slifka, M.K.; Marks, D.L. Depot-Specific Differences in Inflammatory Mediators, and a Role for NK Cells and IFN-γ in Inflammation in Human Adipose Tissue. Int. J. Obes. 2009, 33, 978–990. [Google Scholar] [CrossRef]
- Duffaut, C.; Galitzky, J.; Lafontan, M.; Bouloumié, A. Unexpected Trafficking of Immune Cells within the Adipose Tissue during the Onset of Obesity. Biochem. Biophys. Res. Commun. 2009, 384, 482–485. [Google Scholar] [CrossRef]
- Jagannathan, M.; McDonnell, M.; Liang, Y.; Hasturk, H.; Hetzel, J.; Rubin, D.; Kantarci, A.; Van Dyke, T.E.; Ganley-Leal, L.M.; Nikolajczyk, B.S. Toll-like Receptors Regulate B Cell Cytokine Production in Patients with Diabetes. Diabetologia 2010, 53, 1461–1471. [Google Scholar] [CrossRef]
- Lempesis, I.G.; Van Meijel, R.L.J.; Manolopoulos, K.N.; Goossens, G.H. Oxygenation of Adipose Tissue: A Human Perspective. Acta Physiol. 2020, 228, e13298. [Google Scholar] [CrossRef]
- Murano, I.; Barbatelli, G.; Parisani, V.; Latini, C.; Muzzonigro, G.; Castellucci, M.; Cinti, S. Dead Adipocytes, Detected as Crown-like Structures, Are Prevalent in Visceral Fat Depots of Genetically Obese Mice. J. Lipid Res. 2008, 49, 1562–1568. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gonzalez, D.M.; Kopparapu, A.K. Postpartum Care of the New Mother. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Bergink, V.; Burgerhout, K.M.; Weigelt, K.; Pop, V.J.; De Wit, H.; Drexhage, R.C.; Kushner, S.A.; Drexhage, H.A. Immune System Dysregulation in First-Onset Postpartum Psychosis. Biol. Psychiatry 2013, 73, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, T.; Yang, Y.; Deng, W.; Lu, H.; Wang, S.; Liu, R.; Chang, M.; Wu, S.; Gao, Y.; et al. Postpartum Hepatitis and Host Immunity in Pregnant Women with Chronic HBV Infection. Front. Immunol. 2023, 13, 1112234. [Google Scholar] [CrossRef]
- Groer, M.W.; El-Badri, N.; Djeu, J.; Williams, S.N.; Kane, B.; Szekeres, K. Suppression of Natural Killer Cell Cytotoxicity in Postpartum Women: Time Course and Potential Mechanisms. Biol. Res. Nurs. 2014, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, Y.; Amino, N.; Iwatani, Y.; Kaneda, T.; Mitsuda, N.; Morimoto, Y.; Tanizawa, O.; Miyai, K. Changes in Natural Killer Cell Activity in Normal Pregnant and Postpartum Women: Increases in the First Trimester and Postpartum Period and Decrease in Late Pregnancy. J. Reprod. Immunol. 1991, 20, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Porter, K. Longitudinal Changes in Serum Proinflammatory Markers across Pregnancy and Postpartum: Effects of Maternal Body Mass Index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef]
- Giaglis, S.; Sur Chowdhury, C.; Van Breda, S.V.; Stoikou, M.; Tiaden, A.N.; Daoudlarian, D.; Schaefer, G.; Buser, A.; Walker, U.A.; Lapaire, O.; et al. Circulatory Neutrophils Exhibit Enhanced Neutrophil Extracellular Trap Formation in Early Puerperium: NETs at the Nexus of Thrombosis and Immunity. Int. J. Mol. Sci. 2021, 22, 13646. [Google Scholar] [CrossRef]
- Giaglis, S.; Stoikou, M.; Sur Chowdhury, C.; Schaefer, G.; Grimolizzi, F.; Rossi, S.W.; Hoesli, I.M.; Lapaire, O.; Hasler, P.; Hahn, S. Multimodal Regulation of NET Formation in Pregnancy: Progesterone Antagonizes the Pro-NETotic Effect of Estrogen and G-CSF. Front. Immunol. 2016, 7, 565. [Google Scholar] [CrossRef]
- Gillespie, S.L.; Porter, K.; Christian, L.M. Adaptation of the Inflammatory Immune Response across Pregnancy and Postpartum in Black and White Women. J. Reprod. Immunol. 2016, 114, 27–31. [Google Scholar] [CrossRef]
- Thippeswamy, H.; Davies, W. A New Molecular Risk Pathway for Postpartum Mood Disorders: Clues from Steroid Sulfatase–Deficient Individuals. Arch. Women’s Ment. Health 2021, 24, 391–401. [Google Scholar] [CrossRef]
- Osborne, L.M.; Gilden, J.; Kamperman, A.M.; Hoogendijk, W.J.G.; Spicer, J.; Drexhage, H.A.; Bergink, V. T-Cell Defects and Postpartum Depression. Brain. Behav. Immun. 2020, 87, 397–403. [Google Scholar] [CrossRef] [PubMed]
- McTiernan, C.F.; Morel, P.; Cooper, L.T.; Rajagopalan, N.; Thohan, V.; Zucker, M.; Boehmer, J.; Bozkurt, B.; Mather, P.; Thornton, J.; et al. Circulating T-Cell Subsets, Monocytes, and Natural Killer Cells in Peripartum Cardiomyopathy: Results from the Multicenter IPAC Study. J. Card. Fail. 2018, 24, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, C.; Li, Y.; Guan, H.; Fan, C.; Teng, Y.; Ouyang, Y.; Shan, Z.; Teng, W. Circulating Lymphocyte Subsets and Regulatory T Cells in Patients with Postpartum Thyroiditis during the First Postpartum Year. Clin. Exp. Med. 2009, 9, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Herrock, O.; Deer, E.; LaMarca, B. Setting a Stage: Inflammation during Preeclampsia and Postpartum. Front. Physiol. 2023, 14, 1130116. [Google Scholar] [CrossRef]
- Koletzko, B.; Brands, B.; Chourdakis, M.; Cramer, S.; Grote, V.; Hellmuth, C.; Kirchberg, F.; Prell, C.; Rzehak, P.; Uhl, O.; et al. The Power of Programming and the EarlyNutrition Project: Opportunities for Health Promotion by Nutrition during the First Thousand Days of Life and Beyond. Ann. Nutr. Metab. 2014, 64, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Giussani, D.A.; Forhead, A.J. Intrauterine Programming of Physiological Systems: Causes and Consequences. Physiology 2006, 21, 29–37. [Google Scholar] [CrossRef]
- Mellanby, E. Nutrition and Child-Bearing. Lancet 1933, 222, 1131–1137. [Google Scholar] [CrossRef]
- Barker, D.J. The Fetal and Infant Origins of Adult Disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. Fetal Origins of Coronary Heart Disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef]
- Barker, D. Infant Mortality, Childhood Nutrition, and Ischaemic Heart Disease in England and Wales. Lancet 1986, 327, 1077–1081. [Google Scholar] [CrossRef]
- Barker, D.J.P.; Osmond, C.; Winter, P.D.; Margetts, B.; Simmonds, S.J. Weight in Infancy and Death from Ischaemic Heart Disease. Lancet 1989, 334, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P.; Godfrey, K.M.; Gluckman, P.D.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal Nutrition and Cardiovascular Disease in Adult Life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Lucas, A. Programming by Early Nutrition in Man. Ciba Found. Symp. 1991, 156, 38–50, discussion 50–55. [Google Scholar] [PubMed]
- Hales, C.N.; Barker, D.J.P. The Thrifty Phenotype Hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Hofman, P.L. Insulin Resistance in Short Children with Intrauterine Growth Retardation. J. Clin. Endocrinol. Metab. 1997, 82, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Connelly, A.; Bland, R.M.; Reilly, J.J. Health Impact of Catch-up Growth in Low-birth Weight Infants: Systematic Review, Evidence Appraisal, and Meta-analysis. Matern. Child. Nutr. 2017, 13, mcn.12297. [Google Scholar] [CrossRef]
- Ong, K.K.L. Association between Postnatal Catch-up Growth and Obesity in Childhood: Prospective Cohort Study. BMJ 2000, 320, 967–971. [Google Scholar] [CrossRef]
- Galjaard, S.; Devlieger, R.; Van Assche, F.A. Fetal Growth and Developmental Programming. J. Perinat. Med. 2013, 41, 101–105. [Google Scholar] [CrossRef]
- Low, F.M.; Gluckman, P.D.; Hanson, M.A. Developmental Plasticity and Epigenetic Mechanisms Underpinning Metabolic and Cardiovascular Diseases. Epigenomics 2011, 3, 279–294. [Google Scholar] [CrossRef]
- Warner, M.J.; Ozanne, S.E. Mechanisms Involved in the Developmental Programming of Adulthood Disease. Biochem. J. 2010, 427, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.G.; Forsen, T.; Tuomiletho, J.; Winter, P.D.; Osmond, C.; Baker, D.J. Catch–up Growth in Childhood and Death from Coronary Heart Disease: Longitudinal Study. BMJ 1999, 318, 427–431. [Google Scholar] [CrossRef]
- Roseboom, T.; De Rooij, S.; Painter, R. The Dutch Famine and Its Long-Term Consequences for Adult Health. Early Hum. Dev. 2006, 82, 485–491. [Google Scholar] [CrossRef]
- Roseboom, T.J.; Painter, R.C.; Van Abeelen, A.F.M.; Veenendaal, M.V.E.; De Rooij, S.R. Hungry in the Womb: What Are the Consequences? Lessons from the Dutch Famine. Maturitas 2011, 70, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Hult, M.; Tornhammar, P.; Ueda, P.; Chima, C.; Edstedt Bonamy, A.-K.; Ozumba, B.; Norman, M. Hypertension, Diabetes and Overweight: Looming Legacies of the Biafran Famine. PLoS ONE 2010, 5, e13582. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Forrester, T.; Barker, D.J.P.; Jackson, A.A.; Landman, J.P.; Hall, J.S.E.; Cox, V.; Osmond, C. Maternal Nutritional Status in Pregnancy and Blood Pressure in Childhood. BJOG Int. J. Obstet. Gynaecol. 1994, 101, 398–403. [Google Scholar] [CrossRef]
- Silveira, J.A.C.; Colugnati, F.A.B.; Poblacion, A.P.; Taddei, J.A.A.C. The Role of Exclusive Breastfeeding and Sugar-Sweetened Beverage Consumption on Preschool Children’s Weight Gain. Pediatr. Obes. 2015, 10, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Meek, J.Y.; Noble, L. Section on Breastfeeding Policy Statement: Breastfeeding and the Use of Human Milk. Pediatrics 2022, 150, e2022057988. [Google Scholar] [CrossRef]
- Quigley, M.A.; Carson, C.; Sacker, A.; Kelly, Y. Exclusive Breastfeeding Duration, and Infant Infection. Eur. J. Clin. Nutr. 2016, 70, 1420–1427. [Google Scholar] [CrossRef]
- Bowatte, G.; Tham, R.; Allen, K.; Tan, D.; Lau, M.; Dai, X.; Lodge, C. Breastfeeding and Childhood Acute Otitis Media: A Systematic Review and Meta-analysis. Acta Paediatr. 2015, 104, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Lodge, C.; Tan, D.; Lau, M.; Dai, X.; Tham, R.; Lowe, A.; Bowatte, G.; Allen, K.; Dharmage, S. Breastfeeding and Asthma and Allergies: A Systematic Review and Meta-analysis. Acta Paediatr. 2015, 104, 38–53. [Google Scholar] [CrossRef]
- Dogaru, C.M.; Nyffenegger, D.; Pescatore, A.M.; Spycher, B.D.; Kuehni, C.E. Breastfeeding and Childhood Asthma: Systematic Review and Meta-Analysis. Am. J. Epidemiol. 2014, 179, 1153–1167. [Google Scholar] [CrossRef]
- Xu, L.; Lochhead, P.; Ko, Y.; Claggett, B.; Leong, R.W.; Ananthakrishnan, A.N. Systematic Review with Meta-analysis: Breastfeeding and the Risk of Crohn’s Disease and Ulcerative Colitis. Aliment. Pharmacol. Ther. 2017, 46, 780–789. [Google Scholar] [CrossRef]
- Horta, B.L.; Loret De Mola, C.; Victora, C.G. Long-term Consequences of Breastfeeding on Cholesterol, Obesity, Systolic Blood Pressure and Type 2 Diabetes: A Systematic Review and Meta-analysis. Acta Paediatr. 2015, 104, 30–37. [Google Scholar] [CrossRef]
- Ram, K.T.; Bobby, P.; Hailpern, S.M.; Lo, J.C.; Schocken, M.; Skurnick, J.; Santoro, N. Duration of Lactation Is Associated with Lower Prevalence of the Metabolic Syndrome in Midlife—SWAN, the Study of Women’s Health across the Nation. Am. J. Obstet. Gynecol. 2008, 198, 268.e1–268.e6. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Sinha, B.; Sankar, M.J.; Taneja, S.; Bhandari, N.; Rollins, N.; Bahl, R.; Martines, J. Breastfeeding and Maternal Health Outcomes: A Systematic Review and Meta-analysis. Acta Paediatr. 2015, 104, 96–113. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, H.W.; Zhang, Y.; Hood, W.R.; Kavazis, A.N. Lactation Has Persistent Effects on a Mother’s Metabolism and Mitochondrial Function. Sci. Rep. 2017, 7, 17118. [Google Scholar] [CrossRef]
- Anhê, G.F.; Bordin, S. The Adaptation of Maternal Energy Metabolism to Lactation and Its Underlying Mechanisms. Mol. Cell. Endocrinol. 2022, 553, 111697. [Google Scholar] [CrossRef]
- Hamosh, M.; Clary, T.R.; Chernick, S.S.; Scow, R.O. Lipoprotein Lipase Activity of Adipose and Mammary Tissue and Plasma Triglyceride in Pregnant and Lactating Rats. Biochim. Biophys. Acta BBA-Lipids Lipid Metab. 1970, 210, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Steingrimsdottir, L.; Brasel, J.A.; Greenwood, M.R.C. Diet, Pregnancy, and Lactation: Effects on Adipose Tissue, Lipoprotein Lipase, and Fat Cell Size. Metabolism 1980, 29, 837–841. [Google Scholar] [CrossRef]
- Moore, B.J.; Olsen, J.L.; Marks, F.; Brasel, J.A. The Effects of High Fat Feeding during One Cycle of Reproduction Consisting of Pregnancy, Lactation and Recovery on Body Composition and Fat Pad Cellularity in the Rat. J. Nutr. 1984, 114, 1566–1573. [Google Scholar] [CrossRef]
- Zhong, S.; Almario, R.; Dubrinsky, M.; Rose, K.; Lin, P.K.H.; Grunberger, G.; Jen, K.-L.C. Repeated Pregnancy without Lactation: Effects on Maternal Glycemic Control, Pregnancy Outcome, Carcass Composition, and Fat Distribution in Rats. Metabolism 1990, 39, 1127–1132. [Google Scholar] [CrossRef]
- He, X.; Zhu, M.; Hu, C.; Tao, X.; Li, Y.; Wang, Q.; Liu, Y. Breast-Feeding, and Postpartum Weight Retention: A Systematic Review and Meta-Analysis. Public Health Nutr. 2015, 18, 3308–3316. [Google Scholar] [CrossRef]
- Lambrinou, C.-P.; Karaglani, E.; Manios, Y. Breastfeeding and Postpartum Weight Loss. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 413–417. [Google Scholar] [CrossRef]
- Butte, N.F.; King, J.C. Energy Requirements during Pregnancy and Lactation. Public Health Nutr. 2005, 8, 1010–1027. [Google Scholar] [CrossRef]
- Dewey, K.G.; Cohen, R.J.; Brown, K.H.; Rivera, L.L. Effects of Exclusive Breastfeeding for Four versus Six Months on Maternal Nutritional Status, and Infant Motor Development: Results of Two Randomized Trials in Honduras. J. Nutr. 2001, 131, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Weinstock, R.; Thusu, K.; Abdel-Rahman, E.; Aljada, A.; Wadden, T. Tumor Necrosis Factor-α in Sera of Obese Patients: Fall with Weight Loss. J. Clin. Endocrinol. Metab. 1998, 83, 2907–2910. [Google Scholar] [CrossRef]
- Turner, L.; Santosa, S. Putting ATM to BED: How Adipose Tissue Macrophages Are Affected by Bariatric Surgery, Exercise, and Dietary Fatty Acids. Adv. Nutr. 2021, 12, 1893–1910. [Google Scholar] [CrossRef]
- Butte, N.F.; Hopkinson, J.M. Body Composition Changes during Lactation Are Highly Variable among Women. J. Nutr. 1998, 128, 381S–385S. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Gamez, X.; Morales-Castillo, P.; Fernandez-Mejia, C. Maternal Adaptations of the Pancreas and Glucose Homeostasis in Lactation and after Lactation. Mol. Cell. Endocrinol. 2023, 559, 111778. [Google Scholar] [CrossRef] [PubMed]
- Karnik, S.K.; Chen, H.; McLean, G.W.; Heit, J.J.; Gu, X.; Zhang, A.Y.; Fontaine, M.; Yen, M.H.; Kim, S.K. Menin Controls Growth of Pancreatic SS-Cells in Pregnant Mice and Promotes Gestational Diabetes Mellitus. Science 2007, 318, 806–809. [Google Scholar] [CrossRef]
- McManus, R.M.; Cunningham, I.; Watson, A.; Harker, L.; Finegood, D.T. Beta-Cell Function and Visceral Fat in Lactating Women with a History of Gestational Diabetes. Metabolism 2001, 50, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Ozisik, H.; Suner, A.; Cetinkalp, S. Prolactin Effect on Blood Glucose and Insülin in Breastfeeding Women. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1765–1767. [Google Scholar] [CrossRef]
- Stuebe, A.M. Duration of Lactation and Incidence of Type 2 Diabetes. JAMA 2005, 294, 2601. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Gao, Y.-T.; Yang, G.; Li, H.L.; Elasy, T.; Zheng, W.; Shu, X.-O. Duration of Breast-Feeding, and the Incidence of Type 2 Diabetes Mellitus in the Shanghai Women’s Health Study. Diabetologia 2008, 51, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.H.; Kim, H.; Kim, H.; Park, J.; Choi, W.; Choi, W.; Hong, H.J.; Ro, H.-J.; Jun, S.; Choi, S.H.; et al. Lactation Improves Pancreatic β Cell Mass and Function through Serotonin Production. Sci. Transl. Med. 2020, 12, eaay0455. [Google Scholar] [CrossRef]
- Roman, T.; Kechichian, T.; Gamble, P.; Ramsey, P.S.; Saade, G.; Prewit, E.B. 1026: Effect of Lactation on Maternal Metabolic and Inflammatory Markers: A Murine Model. Am. J. Obstet. Gynecol. 2019, 220, S658–S659. [Google Scholar] [CrossRef]
- Ahn, S.; Corwin, E.J. The Association between Breastfeeding, the Stress Response, Inflammation, and Postpartum Depression during the Postpartum Period: Prospective Cohort Study. Int. J. Nurs. Stud. 2015, 52, 1582–1590. [Google Scholar] [CrossRef]
- Groer, M.W.; Davis, M.W. Cytokines, Infections, Stress, and Dysphoric Moods in Breastfeeders and Formula Feeders. J. Obstet. Gynecol. Neonatal Nurs. 2006, 35, 599–607. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur, D.; Satora, M.; Rekowska, A.K.; Kabała, Z.; Łomża, A.; Kimber-Trojnar, Ż.; Leszczyńska-Gorzelak, B. Influence of Breastfeeding on the State of Meta-Inflammation in Obesity—A Narrative Review. Curr. Issues Mol. Biol. 2023, 45, 9003-9018. https://doi.org/10.3390/cimb45110565
Mazur D, Satora M, Rekowska AK, Kabała Z, Łomża A, Kimber-Trojnar Ż, Leszczyńska-Gorzelak B. Influence of Breastfeeding on the State of Meta-Inflammation in Obesity—A Narrative Review. Current Issues in Molecular Biology. 2023; 45(11):9003-9018. https://doi.org/10.3390/cimb45110565
Chicago/Turabian StyleMazur, Dominika, Małgorzata Satora, Anna K. Rekowska, Zuzanna Kabała, Aleksandra Łomża, Żaneta Kimber-Trojnar, and Bożena Leszczyńska-Gorzelak. 2023. "Influence of Breastfeeding on the State of Meta-Inflammation in Obesity—A Narrative Review" Current Issues in Molecular Biology 45, no. 11: 9003-9018. https://doi.org/10.3390/cimb45110565
APA StyleMazur, D., Satora, M., Rekowska, A. K., Kabała, Z., Łomża, A., Kimber-Trojnar, Ż., & Leszczyńska-Gorzelak, B. (2023). Influence of Breastfeeding on the State of Meta-Inflammation in Obesity—A Narrative Review. Current Issues in Molecular Biology, 45(11), 9003-9018. https://doi.org/10.3390/cimb45110565