
Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection

 ,
,  , ,
, , 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of Nanoparticles
2.2. Virus and Anti-Viral Tests
2.3. Cell Lines and Infection
2.4. Cell Viability Assay
2.5. Virus Titration
2.6. Ethical Statement
2.7. Genital HSV-2 Infection and Virus Challenge
2.8. Flow Cytometry Phenotypic Analysis
2.9. qPCR for Cytokines and Chemokines
2.10. Statistical Methods
3. Results
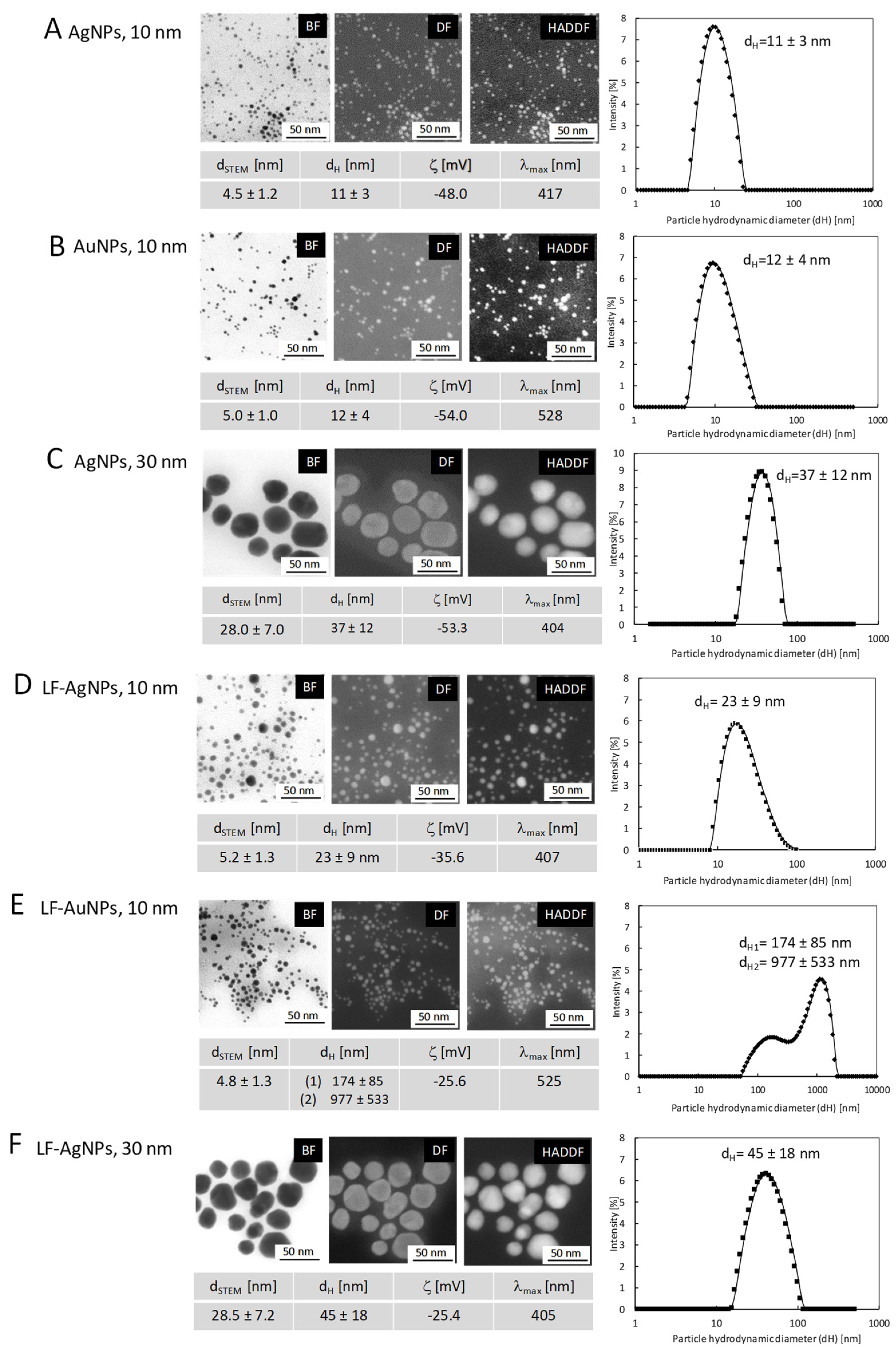
3.1. Characterization of Nanoparticles
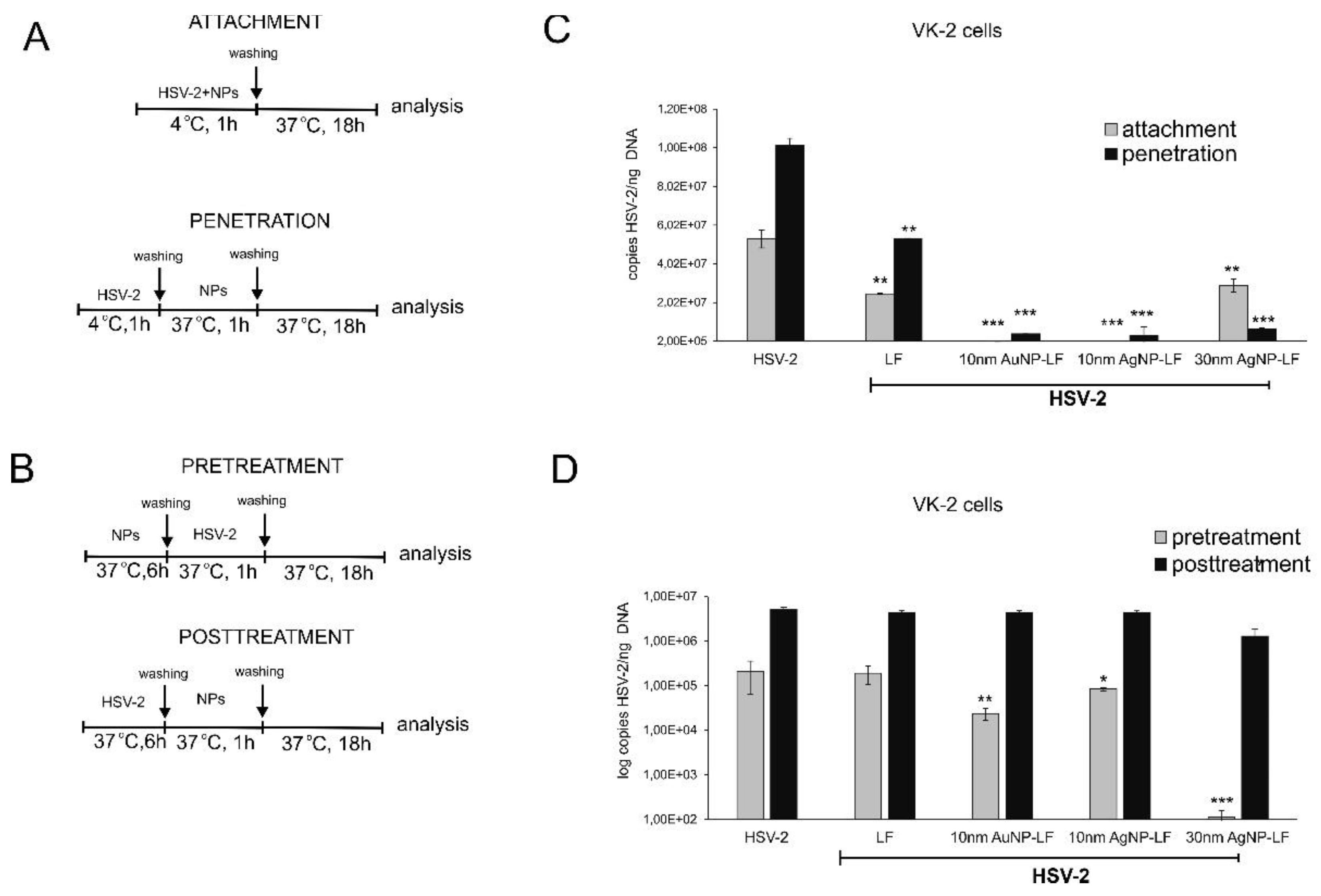
3.2. Antiviral Effects of Lactoferrin Conjugates with Nanoparticles In Vitro
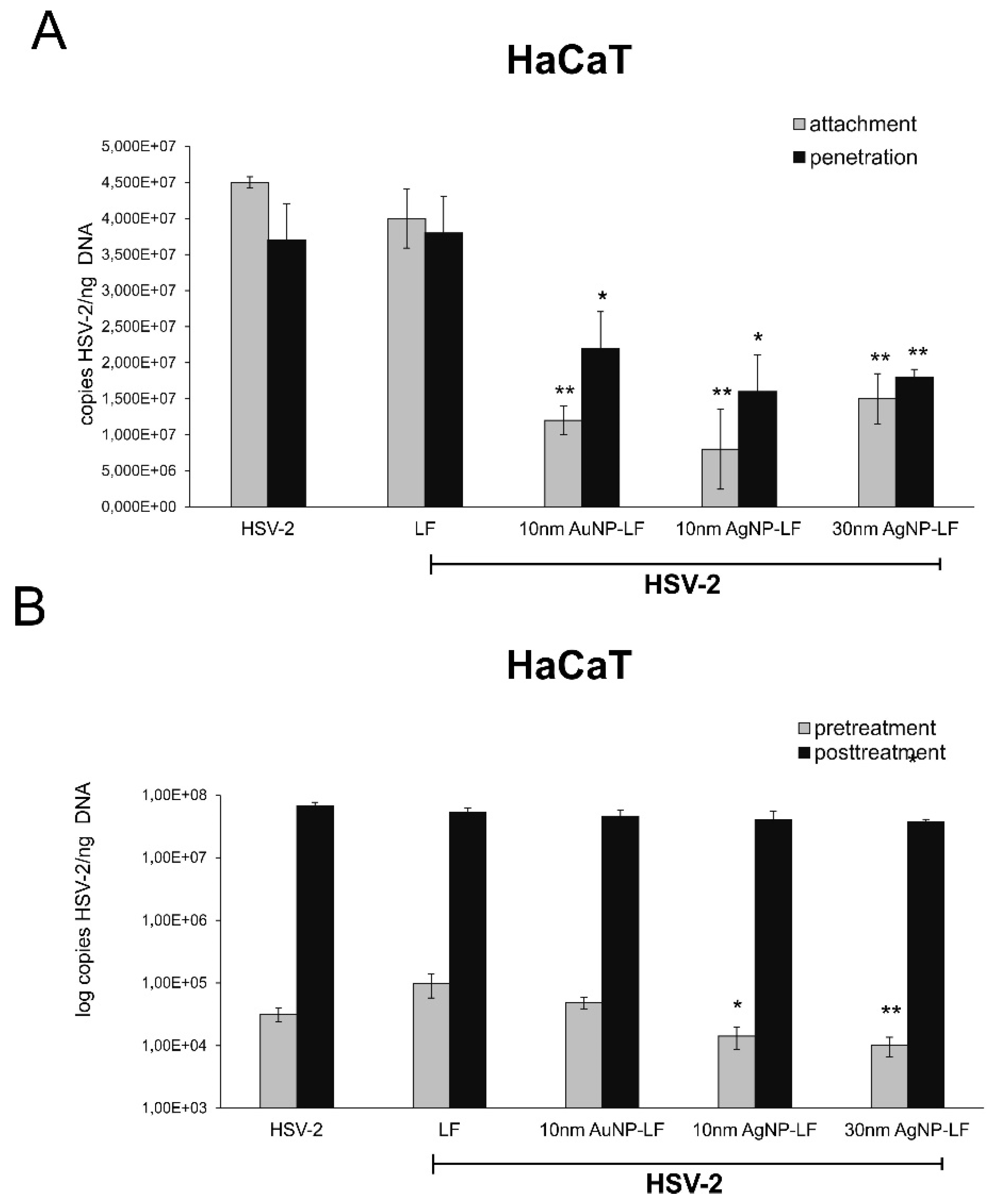
3.3. Lactoferrin-Modified Ag/AuNPs Show Differentiated Antiviral Mechanisms Depending on the Keratinocyte Origin
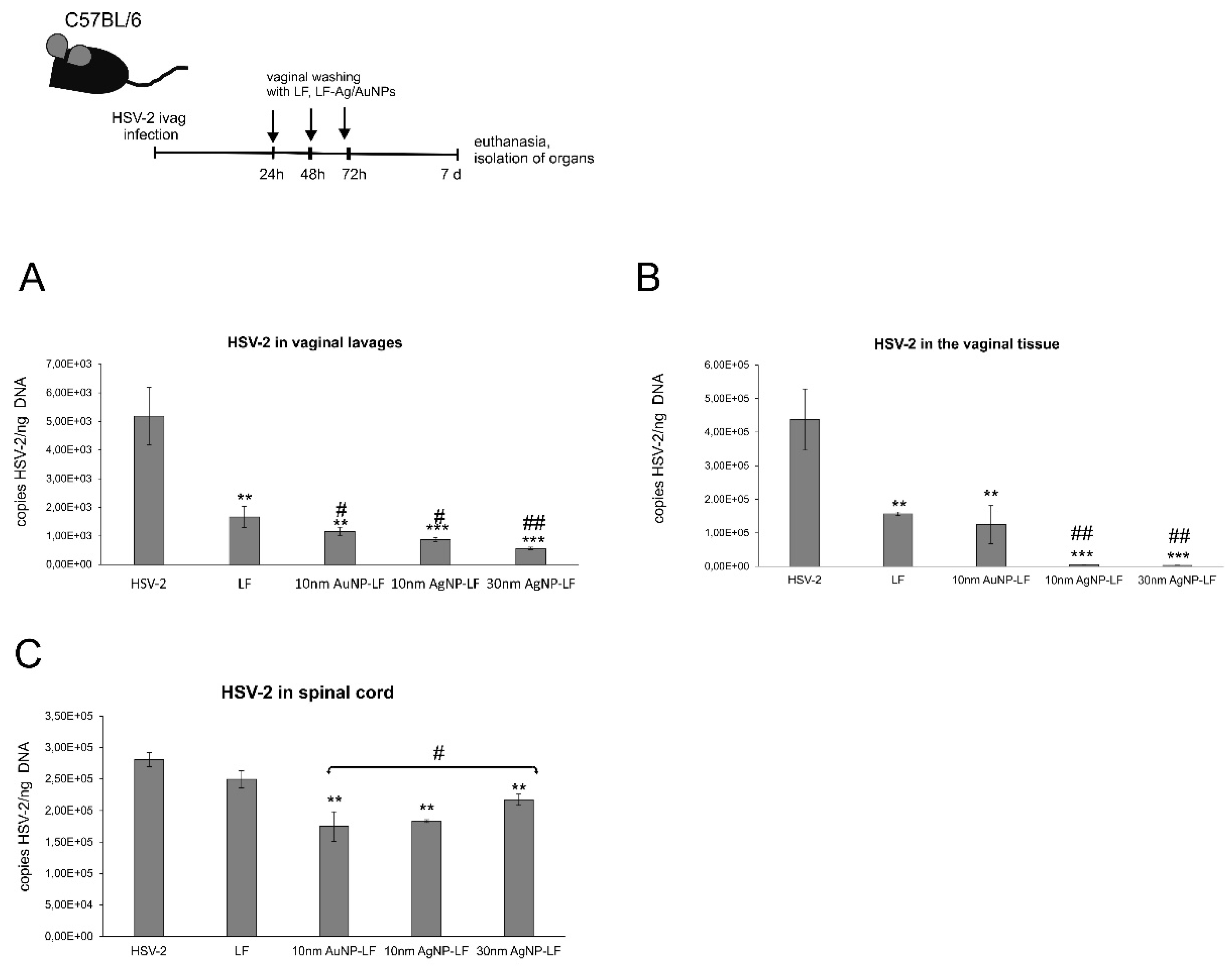
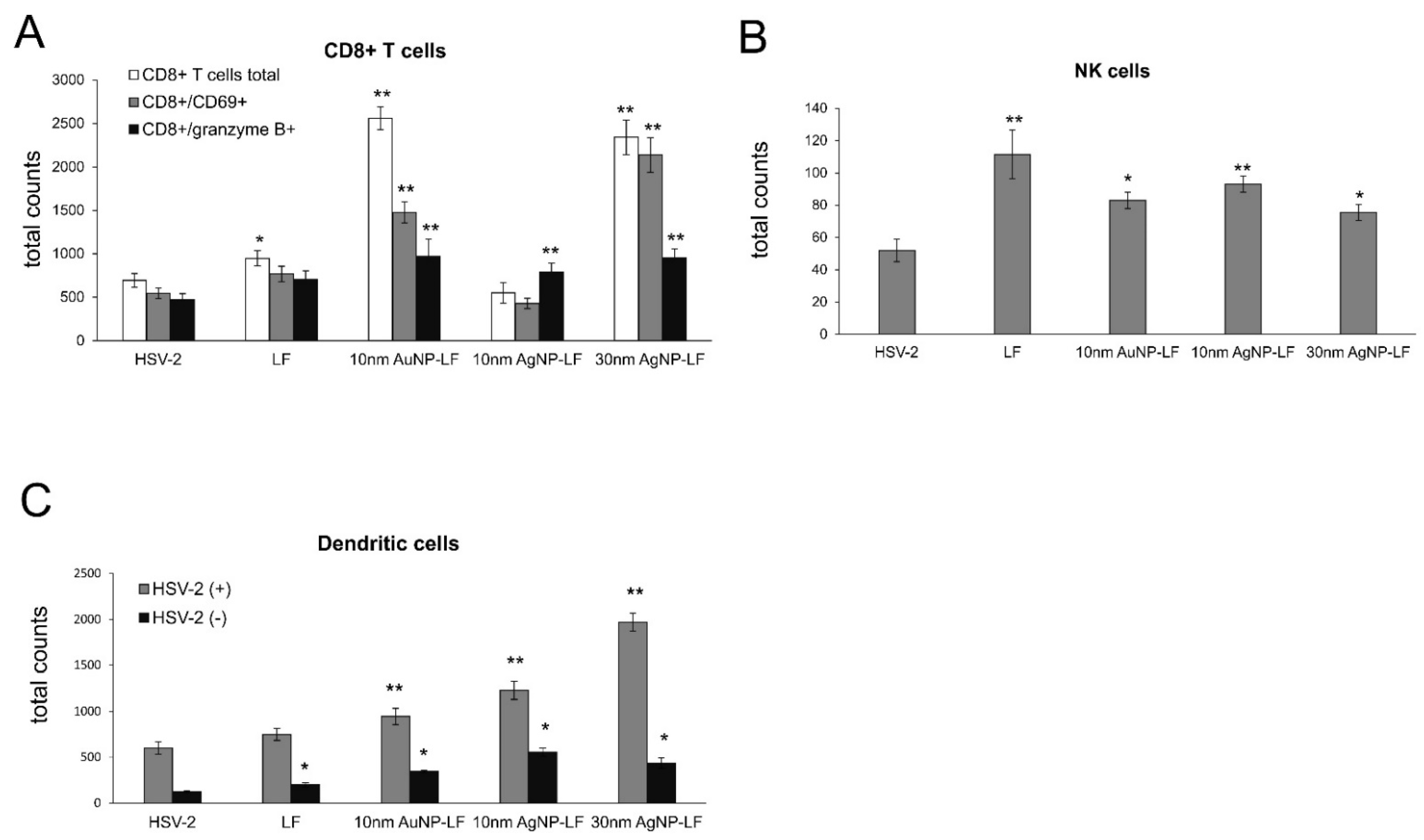
3.4. Lactoferrin-Modified Ag/AuNPs Reduce HSV-2 Infection In Vivo
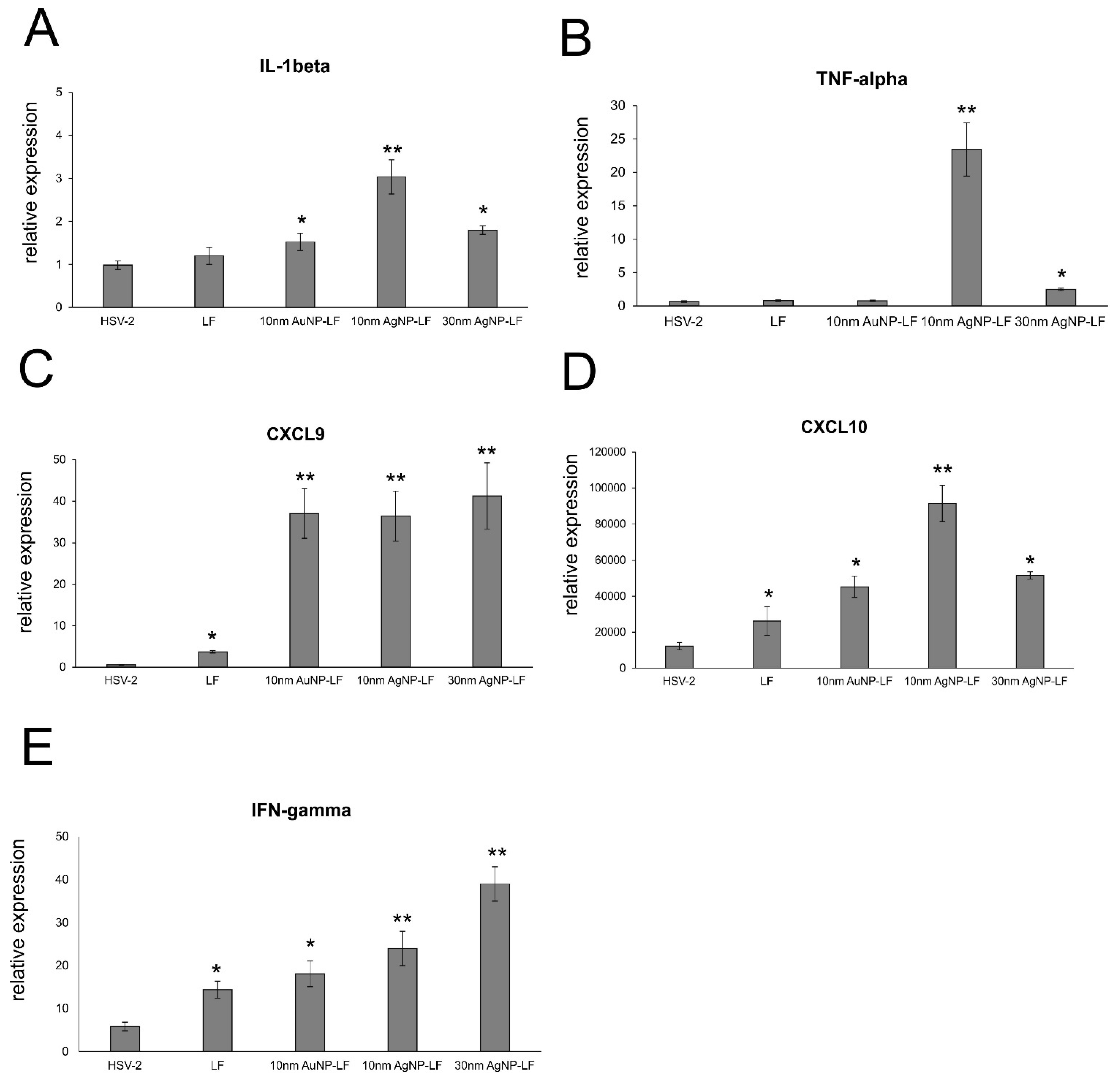
3.5. Lactoferrin-Modified Ag/AuNPs Help to Activate Early Antiviral Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knipe, D.M.; Howley, P. Field’s Virology; Wolters Kluwer Health/Lippincott: Philadelphia, PA, USA; Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- James, C.; Harfouche, M.; Welton, N.J.; Turner, K.M.; Abu-Raddad, L.J.; Gottlieb, S.L.; Looker, K.J. Herpes simplex virus: Global infection prevalence and incidence estimates, 2016. Bull. World Health Organ. 2020, 98, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Wald, A.; Celum, C.L.; Quinn, T.C. The effects of herpes simplex virus-2 on HIV-1 acquisition and transmission: A review of two overlapping epidemics. J. Acquir. Immune Defic. Syndr. 2004, 35, 435–445. [Google Scholar] [CrossRef]
- Jenssen, H.; Sandvik, K.; Andersen, J.H.; Hancock, R.E.; Gutteberg, T.J. Inhibition of HSV cell-to-cell spread by lactoferrin and lactoferricin. Antiviral Res. 2008, 79, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.H. The physiology of lactoferrin. Biochem. Cell Biol. 2002, 80, 1–6. [Google Scholar] [CrossRef]
- Jenssen, H. Anti herpes simplex virus activity of lactoferrin/lactoferricin—An example of antiviral activity of antimicrobial protein/peptide. Cell. Mol. Life Sci 2005, 62, 3002–3013. [Google Scholar] [CrossRef] [PubMed]
- Van Der Strate, B.W.; Beljaars, L.; Molema, G.; Harmsen, M.C.; Meijer, D.K. Antiviral activities of lactoferrin. Antiviral Res. 2001, 52, 225–239. [Google Scholar] [CrossRef]
- El-Fakharany, E.M.; Sánchez, L.; Al-Mehdar, H.A.; Redwan, E.M. Effectiveness of human, camel, bovine and sheep lactoferrin on the hepatitis C virus cellular infectivity: Comparison study. Virol. J. 2013, 10, 199. [Google Scholar] [CrossRef]
- Groot, F.; Geijtenbeek, T.B.; Sanders, R.W.; Baldwin, C.E.; Sanchez-Hernandez, M.; Floris, R.; van Kooyk, Y.; de Jong, E.C.; Berkhout, B.J. Lactoferrin prevents dendritic cell-mediated human immunodeficiency virus type 1 transmission by blocking the DC-SIGN—gp120 interaction. J. Virol. 2005, 79, 3009–3015. [Google Scholar] [CrossRef]
- Andersen, J.H.; Jenssen, H.; Sandvik, K.; Gutteberg, T.J. Anti-HSV activity of lactoferrin and lactoferricin is dependent on the presence of heparan sulphate at the cell surface. J. Med. Virol. 2004, 74, 262–271. [Google Scholar] [CrossRef]
- Välimaa, H.; Tenovuo, J.; Waris, M.; Hukkanen, V. Human lactoferrin but not lysozyme neutralizes HSV-1 and inhibits HSV-1 replication and cell-to-cell spread. Virol. J. 2009, 6, 1–7. [Google Scholar] [CrossRef]
- Blessing, A.A. Metal-based nanoparticles for the treatment of infectious diseases. Molecules 2017, 22, 1370. [Google Scholar]
- Singh, L.; Kruger, H.G.; Maguire, G.E.; Govender, T.; Parboosing, R. The role of nanotechnology in the treatment of viral infections. Ther. Adv. Infect. Dis. 2017, 4, 105–131. [Google Scholar] [CrossRef]
- Orlowski, P.; Tomaszewska, E.; Gniadek, M.; Baska, P.; Nowakowska, J.; Sokolowska, J.; Nowak, Z.; Donten, M.; Celichowski, G.; Grobelny, J.; et al. A Tannic acid modified silver nanoparticles show antiviral activity in herpes simplex virus type 2 infection. PLoS ONE 2014, 9, e104113. [Google Scholar] [CrossRef]
- Suleman Ismail Abdalla, S.; Katas, H.; Chan, J.Y.; Ganasan, P.; Azmi, F.; Fauzi, M.B. Gelatin hydrogels loaded with lactoferrin-functionalized bio-nanosilver as a potential antibacterial and anti-biofilm dressing for infected wounds: Synthesis, characterization, and deciphering of cytotoxicity. Mol. Pharm. 2021, 18, 1956–1969. [Google Scholar] [CrossRef]
- Namvar, L.; Olofsson, S.; Bergstrom, T.; Lindh, M. Detection and typing of herpes simplex virus (HSV) in mucocutaneous samples by TaqMan PCR targeting a gB segment homologous for HSV types 1 and 2. J. Clin. Microbiol. 2005, 43, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Orłowski, P.; Kowalczyk, A.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Węgrzyn, A.; Grzesiak, J.; Celichowski, G.; Grobelny, J.; Eriksson, K.; Krzyzowska, M. Antiviral activity of tannic acid modified silver nanoparticles: Potential to activate immune response in herpes genitalis. Viruses 2018, 10, 524. [Google Scholar] [CrossRef]
- Orlowski, P.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Gniadek, M.; Labedz, O.; Malewski, T.; Nowakowska, J.; Chodaczek, G.; Celichowski, G.; Grobelny, J.; et al. Tannic acid-modified silver and gold nanoparticles as novel stimulators of dendritic cells activation. Front. Immunol. 2018, 9, 1115. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, E.; Orłowski, P.; Winnicka, K.; Tomaszewska, E.; Bąska, P.; Celichowski, G.; Grobelny, J.; Basa, A.; Krzyżowska, M. Multifunctional tannic acid/silver nanoparticle-based mucoadhesive hydrogel for improved local treatment of HSV infection: In vitro and in vivo studies. Int. J. Mol. Sci. 2018, 19, 387. [Google Scholar] [CrossRef]
- Orlowski, P.; Soliwoda, K.; Tomaszewska, E.; Bien, K.; Fruba, A.; Gniadek, M.; Labedz, O.; Nowak, Z.; Celichowski, G.; Grobelny, J.; et al. Toxicity of tannic acid-modified silver nanoparticles in keratinocytes: Potential for immunomodulatory applications. Toxicol. In Vitro 2016, 35, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Yi, J.; Choi, K.; Park, K. Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol. In Vitro 2010, 24, 872–878. [Google Scholar] [CrossRef]
- Johnston, H.J.; Hutchinson, G.; Christensen, F.M.; Peters, S.; Hankin, S.; Stone, V. A review of the in vivo and in vitro toxicity of silver and gold particulates: Particle attributes and biological mechanisms responsible for the observed toxicity. Crit. Rev. Toxicol. 2010, 40, 328–346. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.N.; Baker, H.M.; Kidd, R.D. Lactoferrin and transferrin: Functional variations on a common structural framework. Biochem. Cell Biol. 2002, 80, 27–34. [Google Scholar] [CrossRef]
- Nayak, P.S.; Borah, S.M.; Gogoi, H.; Asthana, S.; Bhatnagar, R.; Jha, A.N.; Jha, S. Lactoferrin adsorption onto silver nanoparticle interface: Implications of corona on protein conformation, nanoparticle cytotoxicity and the formulation adjuvanticity. Chem. Eng. J. 2019, 361, 470–484. [Google Scholar] [CrossRef]
- Marr, A.K.; Jenssen, H.; Moniri, M.R.; Hancock, R.E.; Pante, N. Bovine lactoferrin and lactoferricin interfere with intracellular trafficking of Herpes simplex virus-1. Biochimie 2009, 91, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Ziere, G.J.; van Dijk, M.C.; Bijsterbosch, M.K.; van Berkel, T.J. Lactoferrin uptake by the rat liver. Characterization of the recognition site and effect of selective modification of arginine residues. J. Biol. Chem 1992, 267, 11229–11235. [Google Scholar] [CrossRef]
- Weng, T.Y.; Chen, L.C.; Shyu, H.W.; Chen, S.H.; Wang, J.R.; Yu, C.K.; Lei, H.Y.; Yeh, T.M. Lactoferrin inhibits enterovirus 71 infection by binding to VP1 protein and host cells. Antiviral. Res. 2005, 67, 31–37. [Google Scholar] [CrossRef]
- Shimizu, K.; Matsuzawa, H.; Okada, K.; Tazume, S.; Dosako, S.; Kawasaki, Y.; Hashimoto, K.; Koga, Y. Lactoferrin-mediated protection of the host from murine cytomegalovirus infection by a T-cell-dependent augmentation of natural killer cell activity. Arch. Virol. 1996, 141, 1875–1889. [Google Scholar] [CrossRef]
- Fujihara, T.; Hayashi, K. Lactoferrin inhibits herpes simplex virus type-1 (HSV-1) infection to mouse cornea. Arch. Virol. 1995, 140, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Shestakov, A.; Jenssen, H.; Nordström, I.; Eriksson, K. Lactoferricin but not lactoferrin inhibit herpes simplex virus type 2 infection in mice. Antiviral Res. 2012, 93, 340–345. [Google Scholar] [CrossRef]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Haseggawa, H.; Kajino, K.; Ninomya, T.; et al. Gold nanoparticles as a vaccine platform: Influence of size and shape on immunological responses in vitro and in vivo. ACS Nano. 2013, 7, 3926–3938. [Google Scholar] [CrossRef]
- Sanchez-Guzman, D.; le Guen, P.; Villeret, B.; Sola, N.; Borgne, R.L.; Guyard, A.; Kemmel, A.; Crestani, B.; Sallenave, J.-M.; Verdugo, I.G. Silver nanoparticle-adjuvanted vaccine protects against lethal influenza infection through inducing BALT and IgA-mediated mucosal immunity. Biomaterials 2019, 217, 119308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Deng, X.; Guan, X.; Geng, L.; Fu, M.; Zhang, B.; Chen, R.; Hu, H.; Hu, K.; Zhang, D.; et al. Herpes simplex virus type 2 infection-induced expression of CXCR3 ligands promotes CD4+ T cell migration and is regulated by the viral immediate-early protein ICP4. Front. Immunol. 2018, 9, 2932. [Google Scholar] [CrossRef]
- Cumberbatch, M.; Dearman, R.J.; Kimber, I. Langerhans cells require signals from both tumour necrosis factor alpha and interleukin 1 beta for migration. Adv. Exp. Med. Biol 1997, 92, 125–128. [Google Scholar]
- Iijima, N.; Mattei, L.M.; Iwasaki, A. Recruited inflammatory monocytes stimulate antiviral Th1 immunity in infected tissue. Proc. Natl. Acad. Sci. USA 2011, 108, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Belisle, S.E.; Tisoncik, J.R.; Korth, M.J.; Carter, V.S.; Proll, S.C.; Swayne, D.E.; Pantin-Jackwood, M.; Tumpey, T.M.; Katze, M.G. Genomic profiling of tumor necrosis factor alpha (TNF-alpha) receptor and interleukin-1 receptor knockout mice reveals a link between TNF-alpha signaling and increased severity of 1918 pandemic influenza virus infection. J. Virol. 2010, 84, 12576–12588. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ag/AuNPs | Vero Cells CC50 2 µg/mL ± SEM | Vero Cells MNTC 3 µg/mL ± SEM | HaCaT Cells CC50 µg/mL ± SEM | HaCaT Cells MNTC µg/mL ± SEM | VK-2/E6/E7 Cells CC50 µg/mL ± SEM | VK-2/E6/E7 Cells MNTC µg/mL ± SEM |
|---|---|---|---|---|---|---|
| LF-AuNPS, 10 nm | 39.2 ± 4.7 | 8.02 ± 1 | 37.6 ± 4.1 | 7.78 ± 0.88 | 28.2 ± 4.2 | 6.99 ± 1.1 |
| LF-AgNPs, 10 nm | 15.01 ± 5.2 | 6.56 ± 0.45 | 16.1 ± 2.9 | 7.2 ± 0.6 | 11.9 ± 1.4 | 5.1 ± 0.5 |
| LF-AgNPs, 30 nm | 33.3 ± 6.1 | 9.1 ± 0.78 | 33.2 ± 7.01 | 8.1 ± 0.94 | 27.2 ± 4.3 | 8.99 ± 0.55 |
| AuNPS, 10 nm | 36.8 ± 7.1 | 9.8 ± 2.5 | 35.8 ± 7.9 | 7.21 ± 0.67 | 26 ± 9.1 | 7.22 ± 1.2 |
| AgNPs, 10 nm | 11.22 ± 3.4 | 10 ± 3 | 13.8 ± 1 | 8.1 ± 0.77 | 12.3 ± 0.9 | 7.77 ± 1.4 |
| AgNPs, 30 nm | 31.1 ± 2.1 | 9.39 ± 2.9 | 30.8 ± 2.2 | 8.88 ± 0.81 | 24.2 ± 1.4 | 8.5 ± 1.9 |
| LF solution 4 | 34.7 ± 7.2 | 11.45 ± 3.01 | 39.1 ± 5.4 | 12.34 ± 2.1 | 38.5 ± 2.1 | 16.2 ± 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyzowska, M.; Chodkowski, M.; Janicka, M.; Dmowska, D.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Bednarczyk, K.; Celichowski, G.; Grobelny, J. Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection. Microorganisms 2022, 10, 110. https://doi.org/10.3390/microorganisms10010110
Krzyzowska M, Chodkowski M, Janicka M, Dmowska D, Tomaszewska E, Ranoszek-Soliwoda K, Bednarczyk K, Celichowski G, Grobelny J. Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection. Microorganisms. 2022; 10(1):110. https://doi.org/10.3390/microorganisms10010110
Chicago/Turabian StyleKrzyzowska, Malgorzata, Marcin Chodkowski, Martyna Janicka, Dominika Dmowska, Emilia Tomaszewska, Katarzyna Ranoszek-Soliwoda, Katarzyna Bednarczyk, Grzegorz Celichowski, and Jaroslaw Grobelny. 2022. "Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection" Microorganisms 10, no. 1: 110. https://doi.org/10.3390/microorganisms10010110
APA StyleKrzyzowska, M., Chodkowski, M., Janicka, M., Dmowska, D., Tomaszewska, E., Ranoszek-Soliwoda, K., Bednarczyk, K., Celichowski, G., & Grobelny, J. (2022). Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection. Microorganisms, 10(1), 110. https://doi.org/10.3390/microorganisms10010110