
Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles and Their Pathogenicity against Plutella xylostella (Linnaeus)

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Collection, Isolation and Identification of Metarhizium anisopliae
2.3. Pathogenicity of Metarhizium anisopliae Isolate SM036 against Plutella xylostella
2.4. Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles
2.5. Characterization of Metarhizium anisopliae–Chitosan Nanoparticles
2.6. Biological Activity of Chitosan-Based Nanoparticles of Metarhizium anisopliae against Plutella xylostella under Laboratory Conditions
2.7. Biological Activity of Chitosan-Based Nanoparticles of Metarhizium anisopliae against Plutella xylostella under Field Conditions
2.8. Data Analysis
3. Results
3.1. Isolation and Identification of Metarhizium anisopliae Isolate SM036
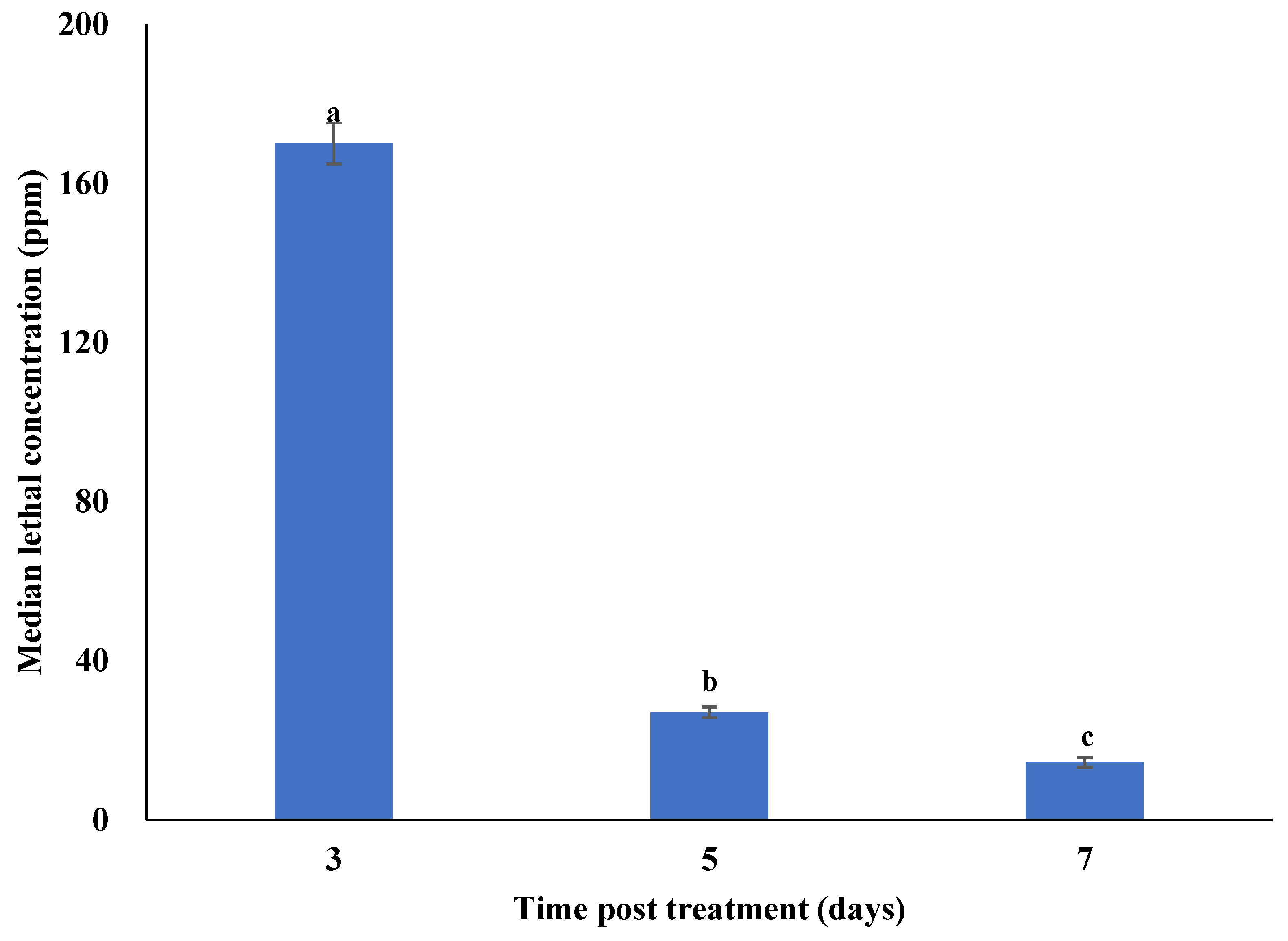
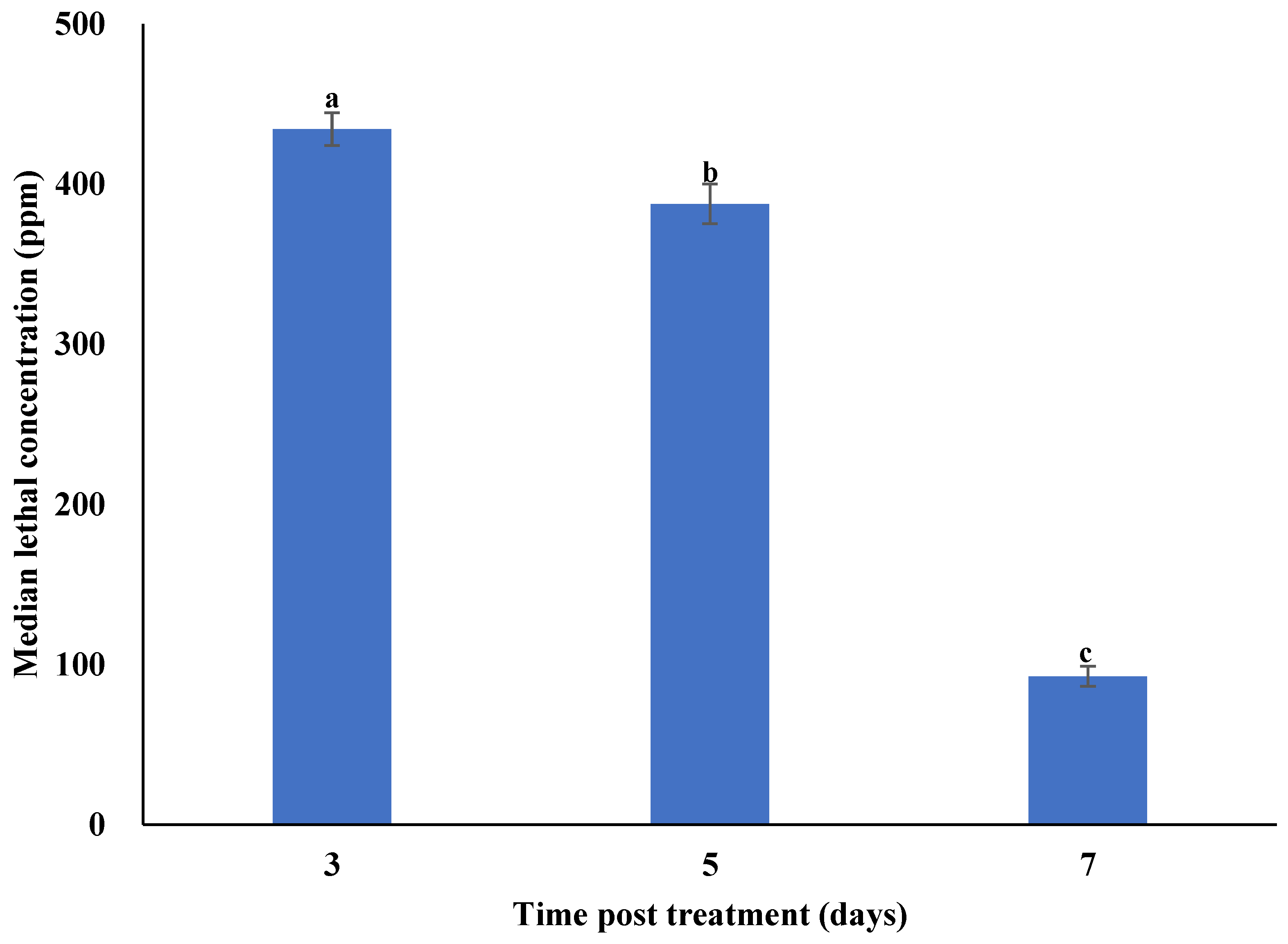
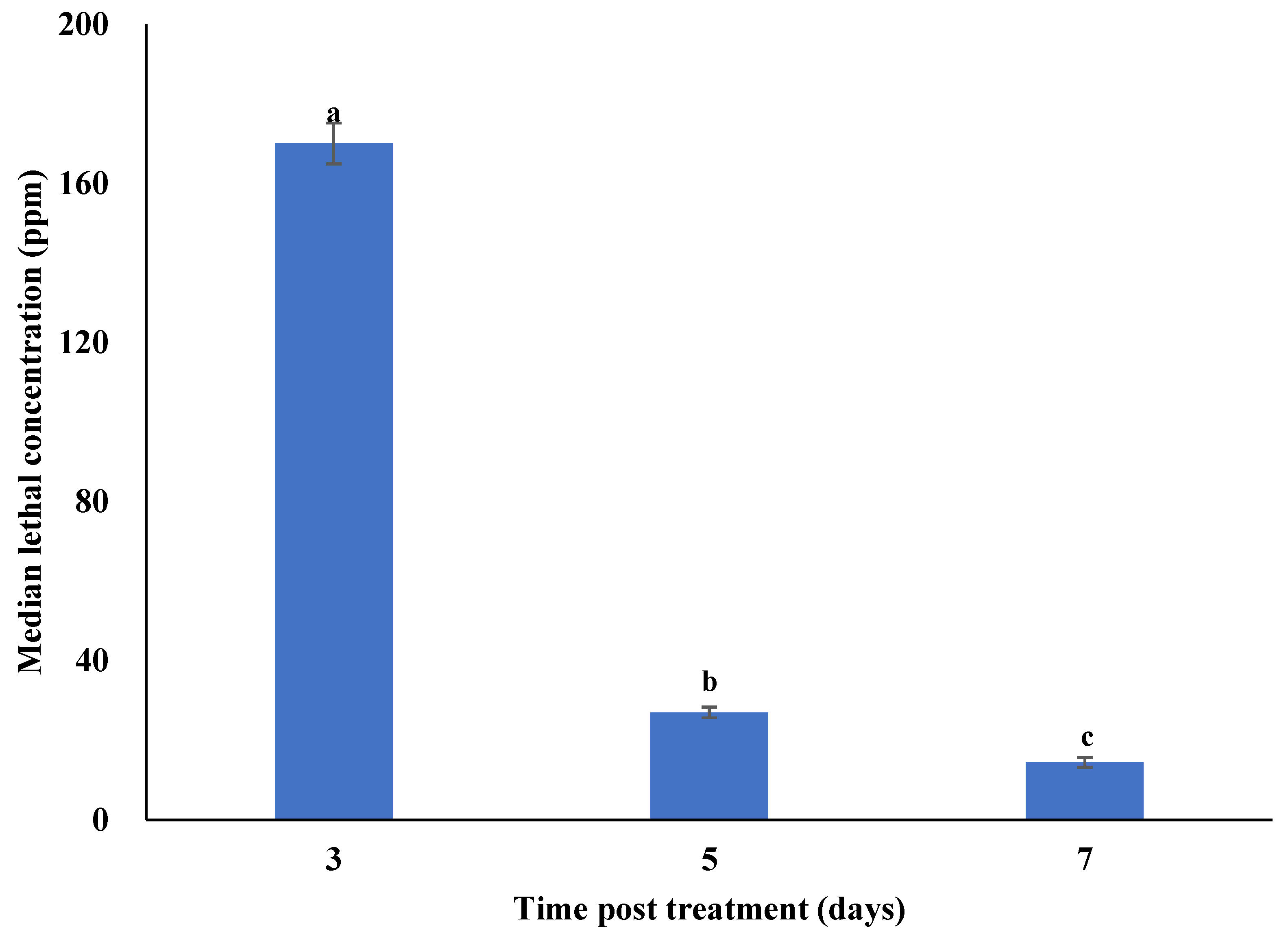
3.2. Concentration–Mortality Response of Plutella xylostella to Metarhizium anisopliae Isolate SM036
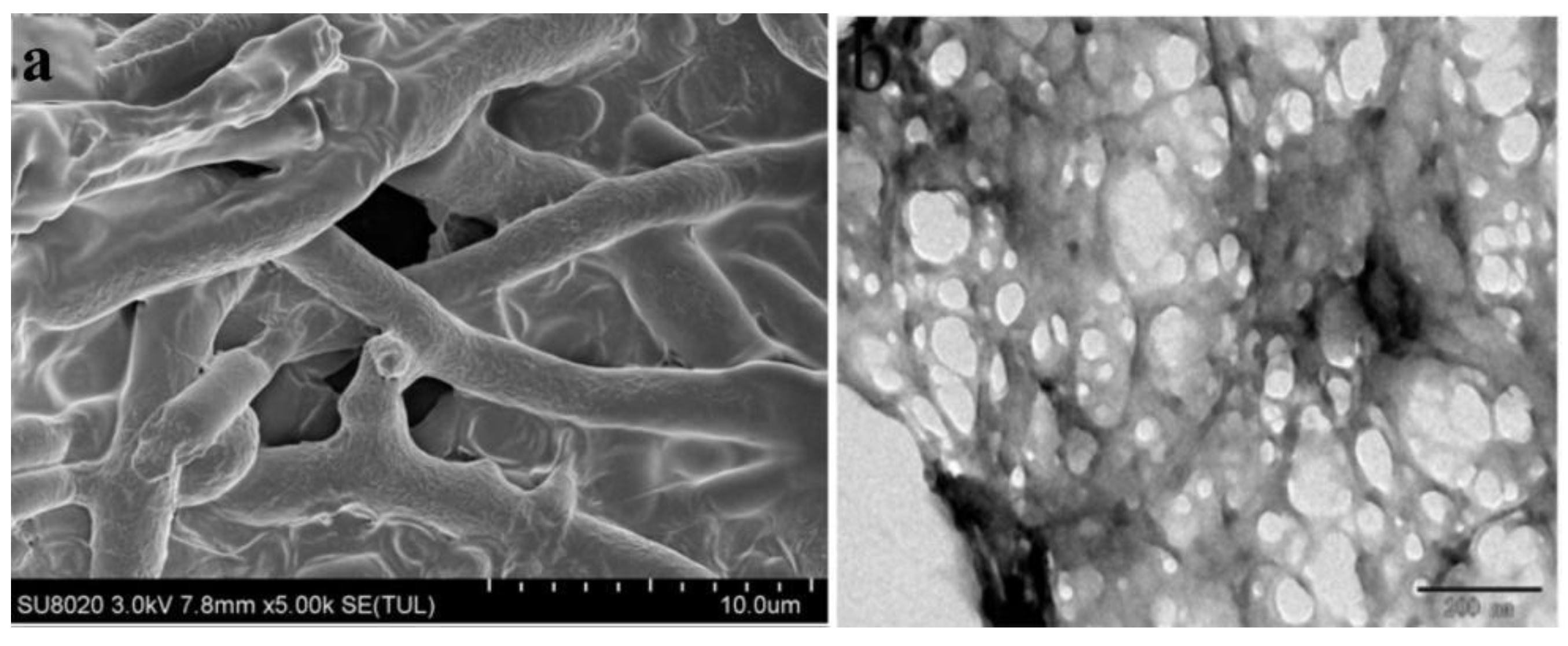
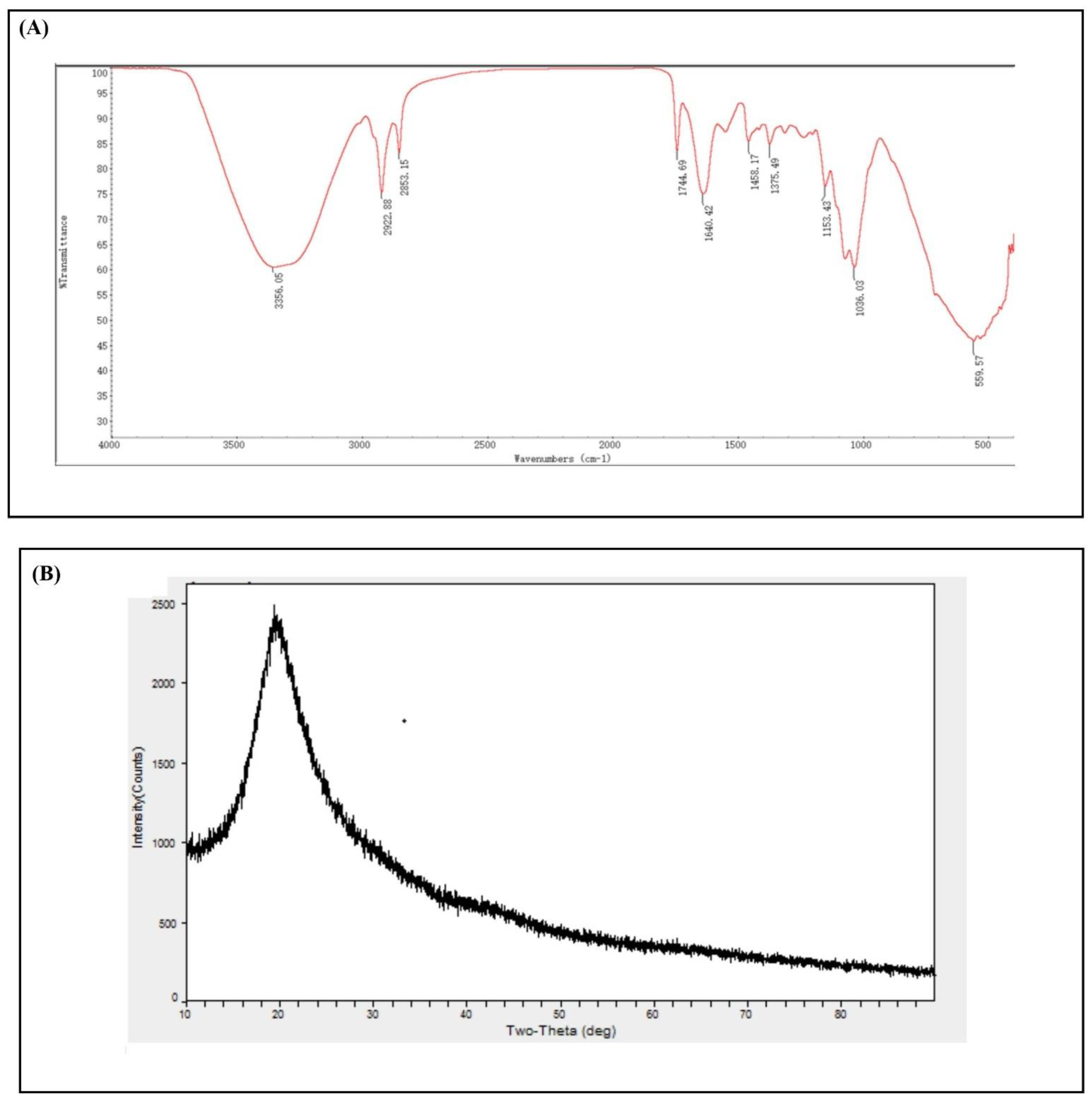
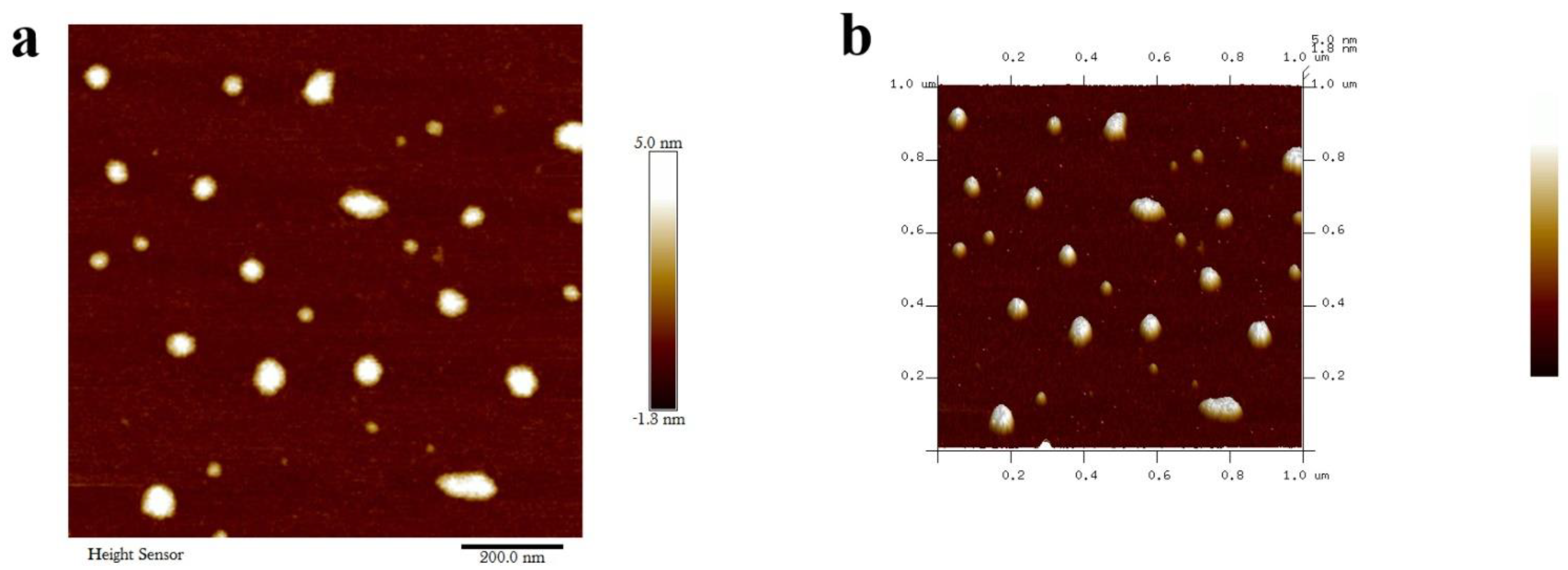
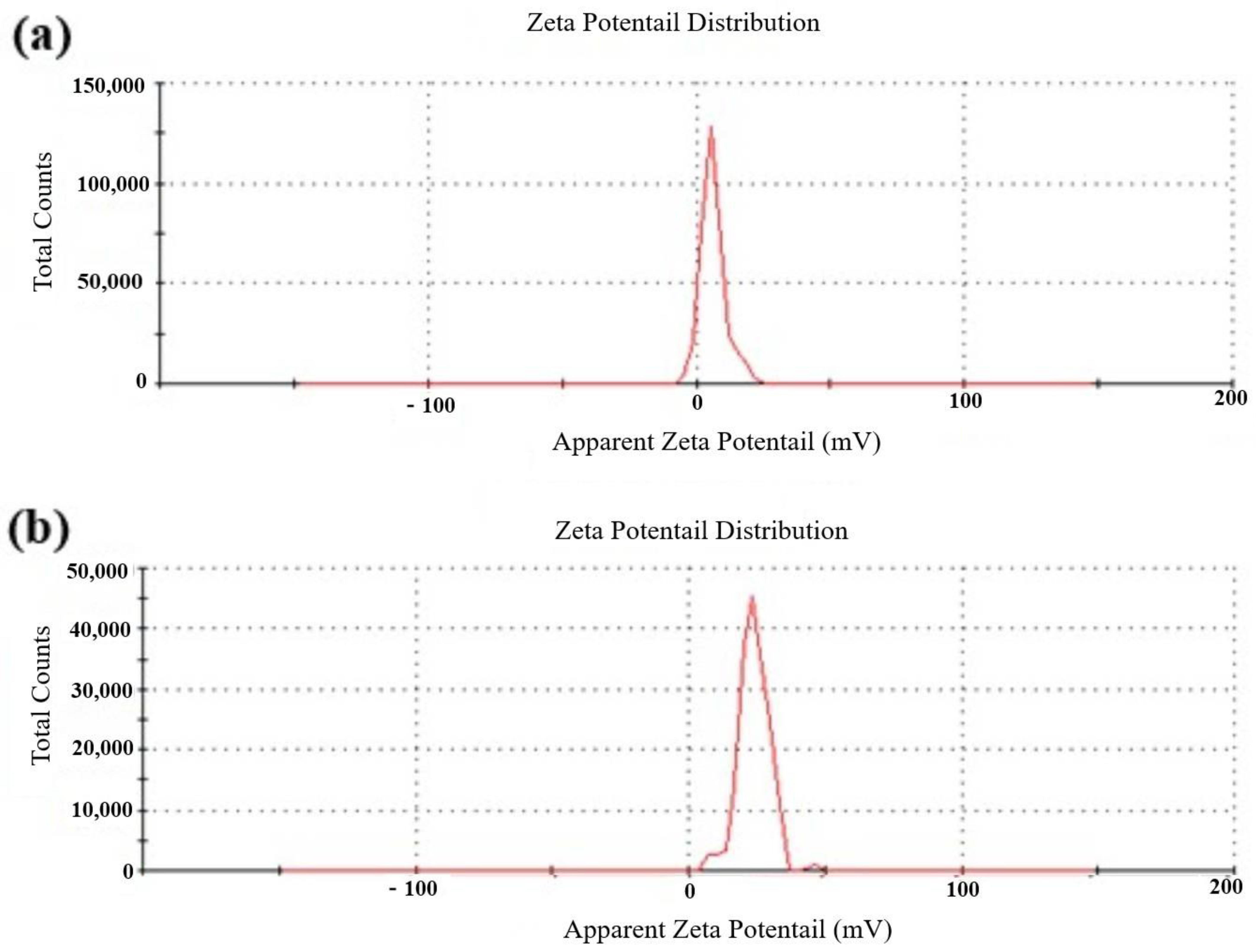
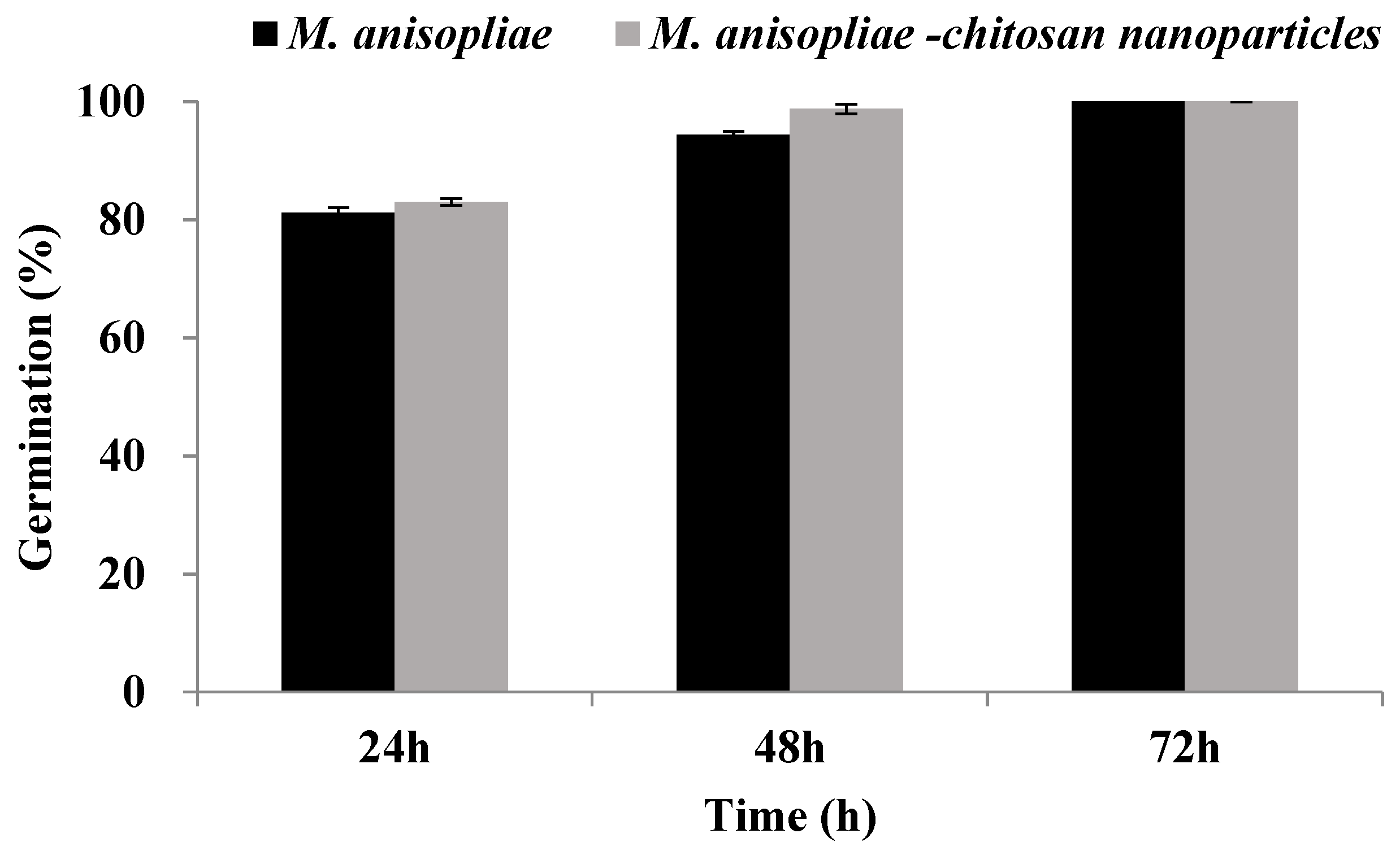
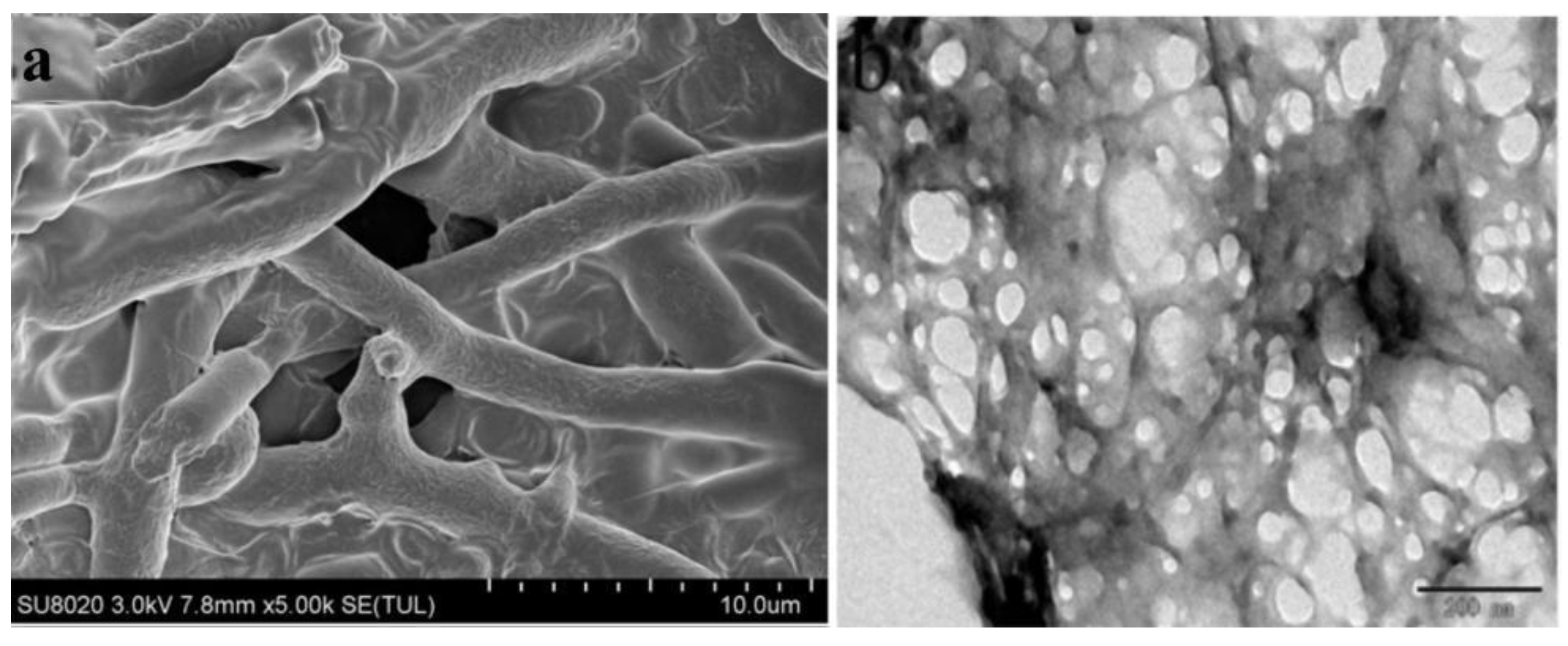
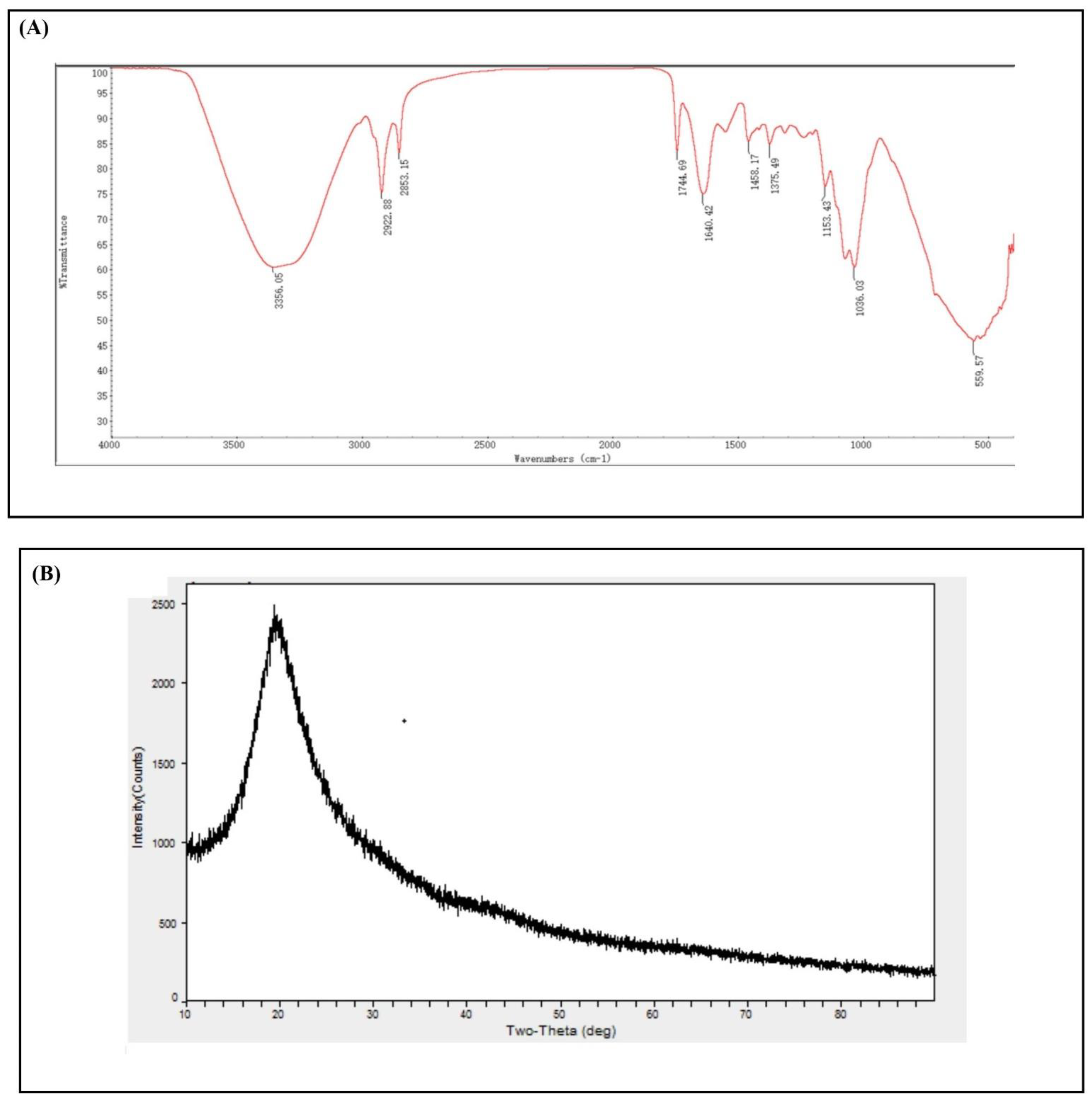
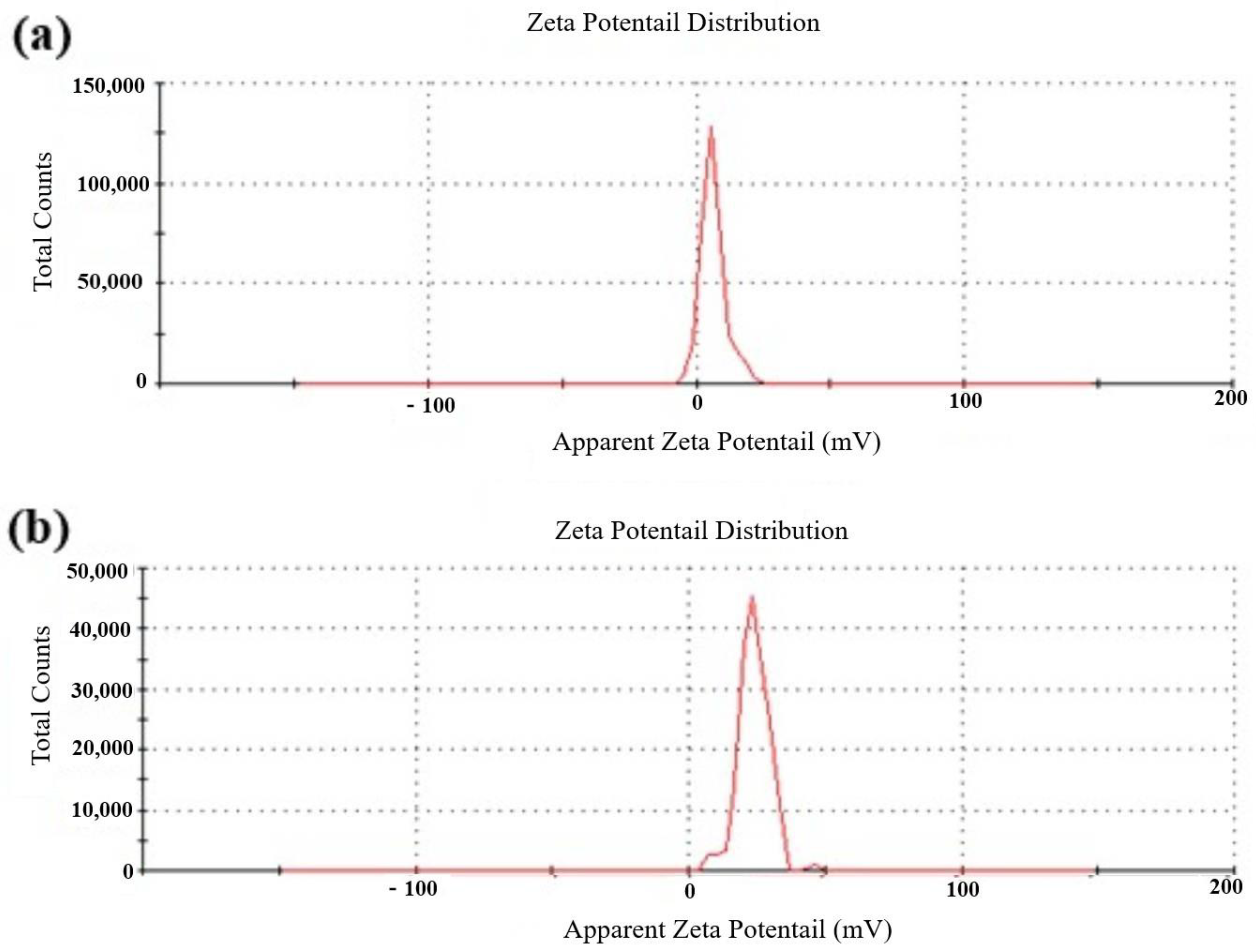
3.3. Characterization of Metarhizium anisopliae–Chitosan Nanoparticles
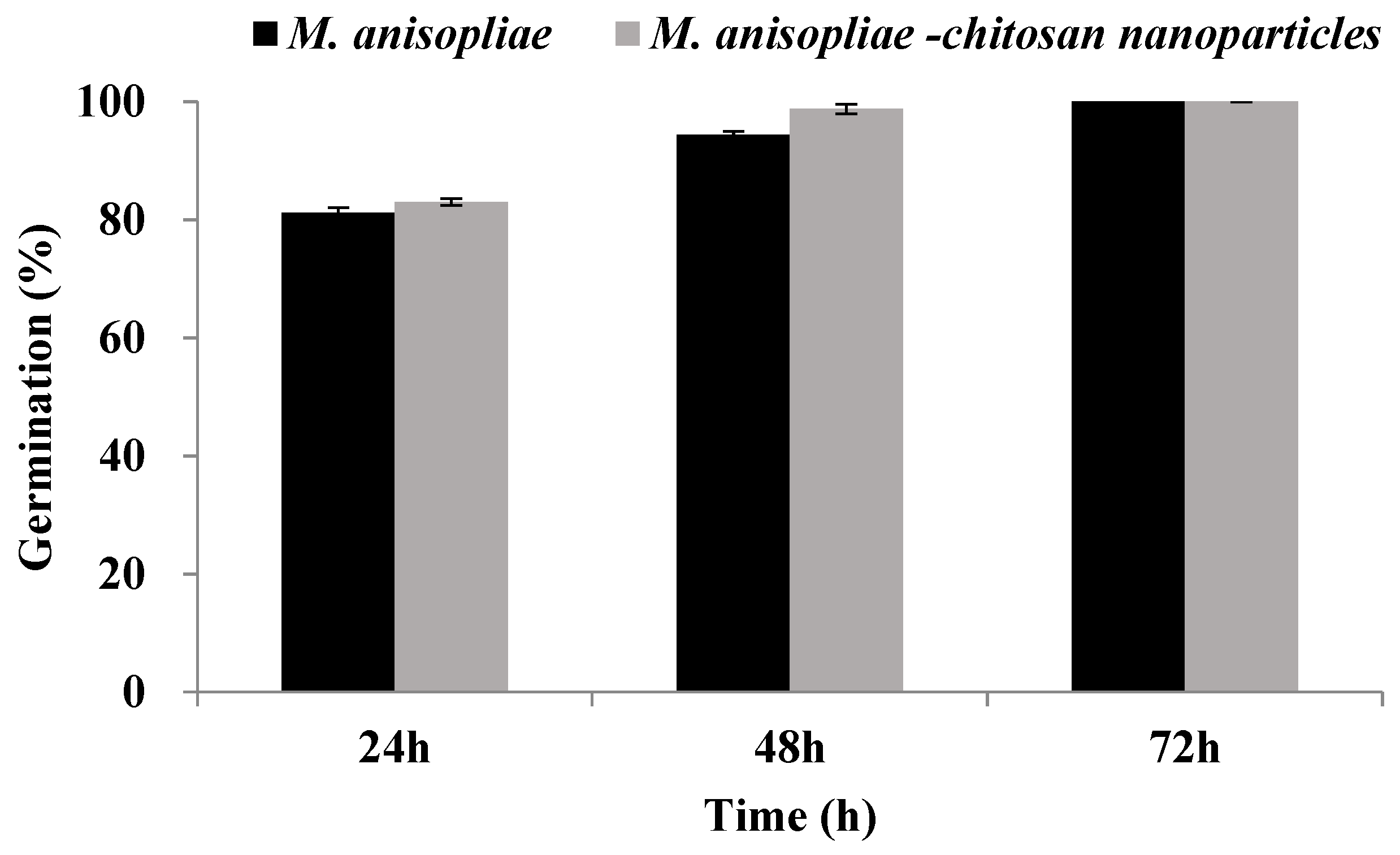
3.4. Biological Activity of Chitosan-Based Nanoparticles of Metarhizium anisopliae against Plutella xylostella under Laboratory Conditions
3.5. Biological Activity of Chitosan-Based Nanoparticles of Metarhizium anisopliae against Plutella xylostella under Field Conditions
4. Discussion
4.1. Isolation and Identification of Metarhizium anisopliae Isolate SM036
4.2. Concentration–Mortality Response of Plutella xylostella to Metarhizium anisopliae Isolate SM036
4.3. Characterization of Chitosan-Based Nanoparticles of Metarhizium anisopliae
4.4. Toxicity of Chitosan-Based Nanoparticles of Metarhizium anisopliae against Plutella xylostella
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Shu-Sheng, L.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Farooq, M.; Nasim, W.; Akram, W.; Khan, F.Z.A.; Jaleel, W. Environment polluting conventional chemical control compared to an environmentally friendly IPM approach for control of diamondback moth, Plutella xylostella (L.), in China: A review. Environ. Sci. Pollut. Res. 2017, 24, 14537–14550. [Google Scholar] [CrossRef]
- Altre, J.A.; Vandenberg, J.D.; Cantone, F.A. Pathogenicity of Paecilomyces fumosoroseus isolates to diamondback moth, Plutella xylostella: Correlation with spore size, germination speed, and attachment to cuticle. J. Invertebr. Pathol. 1999, 73, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Huang, Z.; Ren, S.X. Media composition influences on growth, enzyme activity, and virulence of the entomopathogen hyphomycete Isaria fumosoroseus. Entomol. Exp. Appl. 2009, 131, 30–38. [Google Scholar] [CrossRef]
- Leemon, D.M.; Jonsson, N.N. Laboratory studies on Australian isolates of Metarhizium anisopliae as a biopesticide for the cattle tick Boophilus microplus. J. Invertebr. Pathol. 2008, 97, 40–49. [Google Scholar] [CrossRef]
- Huang, Z.; Ali, S.; Ren, S.X.; Wu, J.H. Effect of Isaria fumosoroseus on mortality and fecundity of Bemisia tabaci and Plutella xylostella. Insect Sci. 2010, 17, 140–148. [Google Scholar] [CrossRef]
- Du, C.L.; Wu, J.H.; Cuthbertson, A.G.S.; Bashir, M.H.; Sun, T.F.; Ali, S. Morphological, molecular and virulence characterisation of six Cordyceps spp. isolates infecting the diamondback moth, Plutella xylostella. Biocontrol Sci. Technol. 2021, 31, 373–386. [Google Scholar] [CrossRef]
- Wu, J.H.; Ali, S.; Huang, Z.; Ren, S.X.; Cai, S.J. Media composition influences on growth, enzyme activity, and virulence of the entomopathogen hyphomycete Metarhizium anisopliae. Pak. J. Zool. 2010, 131, 30–38. [Google Scholar]
- Dong, T.Y.; Zhang, B.W.; Weng, Q.F.; Hu, Q.B. The production relationship of destruxins and blastospores of Metarhizium anisopliae with virulence against Plutella xylostella. J. Integr. Agr. 2016, 15, 1313–1320. [Google Scholar] [CrossRef]
- Zafar, J.; Shoukat, R.F.; Zhang, Y.; Freed, S.; Xu, X.X.; Jin, F. Metarhizium anisopliae challenges immunity and demography of Plutella xylostella. Insects 2020, 11, 694. [Google Scholar] [CrossRef]
- Vandenberg, J.D.; Ramos, M.; Altre, J.A. Dose-response and age- and temperature-related susceptibility of the Diamondback moth (Lepidoptera: Plutellidae) to two isolates of Beauveria bassiana (Hyphomycetes: Moniliaceae). Environ. Entomol. 1998, 27, 1017–1021. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Zhang, C.; Wang, Z.Q.; Wang, X.M.; Wu, J.H.; Cuthbertson, A.G.S.; Shao, Z.F.; Qiu, B.L. Toxicological and biochemical basis of synergism between the entomopathogenic fungus Lecanicillium muscarium and the insecticide matrine against Bemisia tabaci (Gennadius). Sci. Rep. 2017, 7, 46558. [Google Scholar] [CrossRef] [PubMed]
- Gul, H.T.; Saeed, S.; Khan, F.Z.A.; Manzoor, S.A. Potential of nanotechnology in agriculture and crop protection: A review. Appl. Sci. Bus. Econ. 2014, 1, 23–28. [Google Scholar]
- Namasivayam, S.K.R.; Bharani, R.S.A.; Karunamoorthy, K. Insecticidal fungal metabolites fabricated chitosan naocomposite (IM-CNC) preparation for the enhanced larvicidal activity—An effective strategy for green pesticide against economic important insect pests. Int. J. Biol. Macromol. 2018, 120, 921–944. [Google Scholar] [CrossRef] [PubMed]
- Ehdaie, B. Application of nanotechnology in cancer research: Review of progress in the National Cancer Institute’s Alliance for Nanotechnology. Int. J. Biol. Sci. 2007, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Parisi, C.; Vigani, M.; Rodriguez-Cerezo, E. Agricultural nanotechnologies: What are the current possibilities? Nano Today 2015, 10, 124–127. [Google Scholar] [CrossRef]
- Hallberg, K. Towards a responsible research in nanoscience and nanotechnology. Pasi Nano-Bio 2010, 1, 1–47. [Google Scholar]
- Wang, X.S.; Xu, J.; Wang, X.M.; Qiu, B.L.; Cuthbertson, A.G.S.; Du, C.L.; Wu, J.H.; Ali, S. Isaria fumosorosea-based-zero-valent iron nanoparticles affect the growth and survival of sweet potato whitefly, Bemisia tabaci (Gennadius). Pest Manag. Sci. 2019, 75, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, X.H.; Cuthbertson, A.G.S.; Du, C.L.; Ali, S. Toxicity and biological effects of Beauveria brongniartii Fe0 nanoparticles against Spodoptera litura (Fabricius). Insects 2020, 11, 895. [Google Scholar] [CrossRef]
- Wang, X.; Xu, J.; Sun, T.; Ali, S. Synthesis of Cordyceps fumosorosea-biochar nanoparticles and their effects on growth and survival of Bemisia tabaci (Gennadius). Front. Microbiol. 2021, 12, 630220. [Google Scholar] [CrossRef]
- Chen, L.C.; Kung, S.K.; Chen, H.H.; Lin, S.B. Evaluation of zeta potential difference as an indicator for antibacterial strength of low molecular weight chitosan. Carbohydr. Polym. 2010, 82, 913–919. [Google Scholar] [CrossRef]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Anti-fungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Moran, B.; Lopez-Goti, C.; Alonso-Prados, J.L.; Sandis-Espana, P. Consideration of nano-QSAR/QSPR models for nonpesticide risk assessment within the Eupopean legeslative framework. Sci. Total Environ. 2018, 634, 1530–1539. [Google Scholar] [CrossRef]
- Forest, V.; Hochepied, J.F.; Leclerc, L.; Trouve, A.; Abdelkebir, K.; Sarry, G.; Augusto, V.; Pourchez, J. Towards an alternative to nano-QSAR for nanoparticle toxicity ranking in case of small datasets. Perspectives 2019, 21, 95. [Google Scholar] [CrossRef] [Green Version]
- Villaverde, J.J.; Sevilla-Moran, B.; Lopez-Goti, C.; Alonso-Prados, J.L.; Sandis-Espana, P. Computational methodologies for the risk assessment of the pesticides in the European Union. J. Agric. Food Chem. 2017, 65, 2017–2018. [Google Scholar] [CrossRef] [PubMed]
- Imoulan, A.; Alaoui, A.; Meziane, A.E. Natural occurrence of soil-borne entomopathogenic fungi in the Moroccan endemic forest of Argania spinosa and their pathogenicity to Ceratitis capitata. World J. Microbiol. Biotechnol. 2011, 27, 2619–2628. [Google Scholar] [CrossRef]
- Du, C.L.; Yang, B.; Wu, J.H.; Ali, S. Identification and virulence characterization of two Akanthomyces attenuatus isolates against Megalurothrips usitatus (Thysanoptera: Thripidae). Insects 2019, 10, 168. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Minnis, D.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Banu, A.N.; Balasubramanian, C. Optimization and synthesis of silver nanoparticles using Isaria fumosorosea against human vector mosquitoes. Parasitol. Res. 2014, 113, 3843–3851. [Google Scholar] [CrossRef] [PubMed]
- Amerasan, D.; Nataraj, T.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Nicoletti, M. Myco-synthesis of silver nanoparticles using Metarhizium anisopliae against the rural malaria vector Anopheles culicifacies Giles (Diptera: Culicidae). J. Pest Sci. 2016, 89, 249–256. [Google Scholar] [CrossRef]
- Finney, J.; Smith, D.; Skeeters, D.; Auvenshine, C. MMPI alcoholism scales: Factor structure and content analysis. J. Stud. Alcohol. 1971, 32, 1055–1060. [Google Scholar] [CrossRef]
- SAS Institute. SAS User’s Guide; Statistics SAS Institute: Cary, NC, USA, 2000. [Google Scholar]
- Imoulan, A.; Wu, H.J.; Lu, W.L.; Li, Y.; Li, B.B.; Yang, R.H.; Wang, W.J.; Wang, X.L.; Kirk, P.M.; Yao, Y.J. Beauveria medogensis sp. nov.; a new fungus of the entomopathogenic genus from China. J. Invertebr. Pathol. 2016, 139, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, M.; Vargas, M.; Gerding, M.; Ceballos, R.; Oyarzua, P. Molecular, morphological and pathogenic characterization of six strains of Metarhizium spp. (Deuteromycotina: Hyphomycetes) for the control of Aegorhinus superciliosus (Coleoptera: Curculionidae). Chil. J. Agric. Res. 2016, 76, 77–83. [Google Scholar] [CrossRef]
- Bischoff, J.; Rehner, S.; Humber, R. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, É.K.K.; Keyser, C.A.; Chong, J.P.; Rangel, D.E.N.; Miller, M.P.; Roberts, D.E. Characterization of Metarhizium species and varieties based on molecular analysis, heat tolerance and cold activity. J. Appl. Microbiol. 2010, 108, 115–128. [Google Scholar] [CrossRef]
- Loc, N.T.; Chi, V.T.B. Biocontrol potential of Metarhizium anisopliae and Beauveria bassiana against diamondback moth, Plutella xylostella. Omonrice 2007, 15, 86–93. [Google Scholar]
- Mehinto, J.T.; Atachi, P.; Kpindou, O.K.D.; Dannon, E.A.; Tamò, M. Mortality of Maruca vitrata (Lepidoptera: Crambidae) larval stages induced by different doses of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana. Int. J. Adv. Res. 2014, 2, 273–285. [Google Scholar]
- Freed, S.; Feng-Liang, J.; Naeem, M.; Ren, S.X.; Hussian, M. Toxicity of proteins secreted by entomopathogenic fungi against Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Agric. Biol. 2012, 14, 291–295. [Google Scholar]
- Xia, J.; Huang, Z.; Hu, Q. Histopathological study of Plutella xylostella infected by three entomopathogenic fungal species. Adv. Entomol. 2013, 1, 15–19. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J. Pest Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Pauli, G.; Moura, M.G.; Eilenberg, J.; Delalibera, J.I. Within-host competition between two entomopathogenic fungi and a granulovirus in Diatraea saccharalis (Lepidoptera: Crambidae). Insects 2018, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Gajbhiye, M.; Kesharwani, J.; Ingle, A.; Gade, A.; Rai, M. Fungus mediated synthesis of silver nanoparticles and its activity against pathogenic fungi in combination of fluconazole. Nanomedicine 2009, 5, 282–286. [Google Scholar]
- Han, J.; Zhou, Z.; Yin, R.; Yang, D.; Nie, J. Alginate-chitosan/hydroxyapatite polyelectrolyte complex porous scaffolds: Preparation and characterization. Int. J. Biol. Macromol. 2010, 46, 199–205. [Google Scholar] [CrossRef]
- Bragg, W.H.; Bragg, W.L. The reflection of X-rays by crystals. Proc. R. Soc. A 1913, 88, 428–438. [Google Scholar] [CrossRef]
- Miller. Indices of crystal faces—Miller indices. In Dictionary of Gems and Gemology; Manutchehr-Danai, M., Ed.; Springer: Berlin, Germany, 2009. [Google Scholar] [CrossRef]
- Du, W.L.; Niu, S.S.; Xu, Y.L.; Xu, Z.R.; Fan, C.L. Antibacterial activity of chitosan tripolyphosphate nanoparticles loaded with various metal ions. Carbohydr. Polym. 2009, 75, 385–389. [Google Scholar] [CrossRef]
- Zain, M.N.; Stapley, A.G.F.; Shama, G. Green synthesis of silver and copper nanoparticles using ascorbic acid and chitosan for antibacterial applications. Carbohydr. Polym. 2014, 112, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Meng, H.; Xia, T.; George, S.; Nel, A.E. A predictive toxicological paradigm for the safety assessment of nanomaterials. ACS Nano 2009, 3, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.S.; Silva, T.M.; Cardoso, J.C.; Albuquerque-Júnior, R.L.C.D.; Zielinska, A.; Souto, E.B.; Severino, P.; Mendonça, M.D.C. Biosynthesis of silver nanoparticles mediated by entomopathogenic fungi: Antimicrobial resistance, nanopesticides, and toxicity. Antibiotics 2021, 10, 852. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.S.; dos Passos, E.M.; de Jesus Seabra, M.G.; Souto, E.B.; Severino, P.; da Costa Mendonça, M. Entomopathogenic fungi biomass production and extracellular biosynthesis of silver nanoparticles for bioinsecticide action. Appl. Sci. 2021, 11, 2465. [Google Scholar] [CrossRef]
- Brandt, C.R.; Adang, M.J.; Spence, K.D. The peritrophic membrane: Ultrastructural analysis and function as a mechanical barrier to microbial infection in Orgyia pseudotsugata. J. Invertebr. Pathol. 1978, 32, 12–24. [Google Scholar] [CrossRef]
- Shahabuddin, M.; Toyoshima, T.; Aikawa, M.; Kaslow, D.C. Transmission-blocking activity of a chitinase inhibitor and activation of malarial parasite chitinase by mosquito protease. Proc. Natl. Acad. Sci. USA 1993, 90, 4266–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Wu, J.H.; Huang, Z.; Ren, S.X. Production and regulation of extracellular chitinase from the entomopathogenic fungus Isaria fumosorosea. Biocontrol Sci. Technol. 2010, 20, 723–738. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Treatment Description | Concentration |
|---|---|---|
| T1 | M. anisopliae–chitosan nanoparticles | 31.25 ppm |
| T2 | M. anisopliae–chitosan nanoparticles | 62.5 ppm |
| T3 | M. anisopliae–chitosan nanoparticles | 125 ppm |
| T4 | M. anisopliae–chitosan nanoparticles | 250 ppm |
| T5 | M. anisopliae–chitosan nanoparticles | 500 ppm |
| T6 | M. anisopliae conidial suspension | 106 conidia/ml |
| T7 | Chitosan nanoparticles | 200 ppm |
| T8 | Control (ddH2O) | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Du, C.; Zhang, J.; Yang, B.; Cuthbertson, A.G.S.; Ali, S. Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles and Their Pathogenicity against Plutella xylostella (Linnaeus). Microorganisms 2022, 10, 1. https://doi.org/10.3390/microorganisms10010001
Wu J, Du C, Zhang J, Yang B, Cuthbertson AGS, Ali S. Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles and Their Pathogenicity against Plutella xylostella (Linnaeus). Microorganisms. 2022; 10(1):1. https://doi.org/10.3390/microorganisms10010001
Chicago/Turabian StyleWu, Jianhui, Cailian Du, Jieming Zhang, Bo Yang, Andrew G. S. Cuthbertson, and Shaukat Ali. 2022. "Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles and Their Pathogenicity against Plutella xylostella (Linnaeus)" Microorganisms 10, no. 1: 1. https://doi.org/10.3390/microorganisms10010001
APA StyleWu, J., Du, C., Zhang, J., Yang, B., Cuthbertson, A. G. S., & Ali, S. (2022). Synthesis of Metarhizium anisopliae–Chitosan Nanoparticles and Their Pathogenicity against Plutella xylostella (Linnaeus). Microorganisms, 10(1), 1. https://doi.org/10.3390/microorganisms10010001