
The New Role for an Old Kinase: Protein Kinase CK2 Regulates Metal Ion Transport

Abstract
:1. CK2—A Pleiotropic Kinase
2. Structure and Function of CK2
3. Functional Genomics and Discovery of Novel CK2 Functionality
4. CK2 and Metal Ion Transport
5. Metal Transporters Regulated by CK2 Are Potential Therapeutic Targets
6. Future Perspective
Conflicts of Interest
References
- Burnett, G.; Kennedy, E.P. The enzymatic phosphorylation of proteins. J. Biol. Chem. 1954, 211, 969–980. [Google Scholar] [PubMed]
- Fabbro, D.; Cowan-Jacob, S.W.; Moebitz, H. Ten things you should know about protein kinases: IUPHAR Review 14. Br. J. Pharmacol. 2015, 172, 2675–2700. [Google Scholar] [CrossRef] [PubMed]
- Krebs, E.G.; Fischer, E.H. Phosphorylase activity of skeletal muscle extracts. J. Biol. Chem. 1955, 216, 113–120. [Google Scholar] [PubMed]
- Pinna, L. A historical view of protein kinase CK2. Cell. Mol. Biol. Res. 1993, 40, 383–390. [Google Scholar]
- Meggio, F.; Pinna, L.A. One-thousand-and-one substrates of protein kinase CK2? FASEB J. 2003, 17, 349–368. [Google Scholar] [CrossRef] [PubMed]
- Litchfield, D.W. Protein kinase CK2: Structure, regulation and role in cellular decisions of life and death. Biochem. J. 2003, 369 Pt 1, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Allende, J.E.; Allende, C.C. Protein kinases. 4. Protein kinase CK2: An enzyme with multiple substrates and a puzzling regulation. FASEB J. 1995, 9, 313–323. [Google Scholar] [PubMed]
- Venerando, A.; Ruzzene, M.; Pinna, L.A. Casein kinase: The triple meaning of a misnomer. Biochem. J. 2014, 460, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, M.; Daniotti, J.L.; Rojo, D.; Allende, C.C.; Allende, J.E. Cloning, expression and properties of the a’ subunit of casein kinase 2 from zebrafish (Danio rerio). Eur. J. Biochem. 1996, 241, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Niefind, K.; Putter, M.; Guerra, B.; Issinger, O.G.; Schomburg, D. GTP plus water mimic ATP in the active site of protein kinase CK2. Nat. Struct. Biol. 1999, 6, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Pinna, L.A. The raison d’etre of constitutively active protein kinases: The Lesson of CK2. Acc. Chem. Res. 2003, 36, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Kappes, F.; Damoc, C.; Knippers, R.; Przybylski, M.; Pinna, L.A.; Gruss, C. Phosphorylation by protein kinase CK2 changes the DNA binding properties of the human chromatin protein DEK. Mol. Cell. Biol. 2004, 24, 6011–6020. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Issinger, O.G. Protein kinase CK2 and its role in cellular proliferation, development and pathology. Electrophoresis 1999, 20, 391–408. [Google Scholar] [CrossRef]
- Tun, N.; O’Doherty, P.; Chen, Z.; Wu, X.; Bailey, T.; Kersaitis, C.; Wu, M. Identification of aluminium transport-related genes via genome-wide phenotypic screening of Saccharomyces cerevisiae. Metallomics 2014, 6, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Hiscox, S.; Nicholson, R.I.; Hogstrand, C.; Kille, P. Protein kinase CK2 triggers cytosolic zinc signaling pathways by phosphorylation of zinc channel ZIP7. Sci. Signal. 2012, 5, ra11. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, N.; Olsen, B.; Raaf, J.; Bretner, M.; Issinger, O.-G.; Niefind, K. Structure of the human protein kinase CK2 catalytic subunit CK2α′ and interaction thermodynamics with the regulatory subunit CK2β. J. Mol. Biol. 2011, 407, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Niefind, K.; Guerra, B.; Ermakowa, I.; Issinger, O.G. Crystal structure of human protein kinase CK2: Insights into basic properties of the CK2 holoenzyme. EMBO J. 2001, 20, 5320–5331. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Niefind, K.; Ermakowa, I.; Issinger, O.-G. Characterization of CK2 holoenzyme variants with regard to crystallization. Mol. Cell. Biochem. 2001, 227, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, A.; Olsen, B.; Issinger, O.-G.; Niefind, K. The protein kinase CK2 (Andante) holoenzyme structure supports proposed models of autoregulation and trans-autophosphorylation. J. Mol. Biol. 2014, 426, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Brown, E.D.; Walsh, C.T. Expression of recombinant human casein kinase II and recombinant heat shock protein 90 in Escherichia coli and characterization of their interactions. Proc. Natl. Acad. Sci. USA 1994, 91, 2767–2771. [Google Scholar] [CrossRef] [PubMed]
- Pechkova, E.; Zanotti, G.; Nicolini, C. Three-dimensional atomic structure of a catalytic subunit mutant of human protein kinase CK2. Acta Crystallogr. 2003, 59 Pt 12, 2133–2139. [Google Scholar] [CrossRef]
- Chantalat, L.; Leroy, D.; Filhol, O.; Nueda, A.; Benitez, M.J.; Chambaz, E.M.; Cochet, C.; Dideberg, O. Crystal structure of the human protein kinase CK2 regulatory subunit reveals its zinc finger-mediated dimerization. EMBO J. 1999, 18, 2930–2940. [Google Scholar] [CrossRef] [PubMed]
- Raaf, J.; Brunstein, E.; Issinger, O.G.; Niefind, K. The interaction of CK2α and CK2β, the subunits of protein kinase CK2, requires CK2β in a preformed conformation and is enthalpically driven. Protein Sci. 2008, 17, 2180–2186. [Google Scholar] [CrossRef] [PubMed]
- Mazzorana, M.; Pinna, L.; Battistutta, R. A structural insight into CK2 inhibition. Mol. Cell. Biochem. 2008, 316, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Pinna, L.A.; Meggio, F. Protein kinase CK2 (“casein kinase-2”) and its implication in cell division and proliferation. Prog. Cell Cycle Res. 1997, 3, 77–97. [Google Scholar] [PubMed]
- Pinna, L.A. Casein kinase 2: An ‘eminence grise’ in cellular regulation? Biochim. Biophys. Acta 1990, 24, 267–284. [Google Scholar] [CrossRef]
- Blanquet, P.R. Casein kinase 2 as a potentially important enzyme in the nervous system. Prog. Neurobiol. 2000, 60, 211–246. [Google Scholar] [CrossRef]
- Perez, D.I.; Gil, C.; Martinez, A. Protein kinases CK1 and CK2 as new targets for neurodegenerative diseases. Med. Res. Rev. 2010, 31, 924–954. [Google Scholar] [CrossRef] [PubMed]
- Faust, M.; Montenarh, M. Subcellular localization of protein kinase CK2. A key to its function? Cell Tissue Res. 2000, 301, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Bibby, A.C.; Litchfield, D.W. The multiple personalities of the regulatory subunit of protein kinase CK2: CK2 dependent and CK2 independent roles reveal a secret identity for CK2beta. Int. J. Biol. Sci. 2005, 1, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Boldyreff, B.; Issinger, O.G. A-Raf kinase is a new interacting partner of protein kinase CK2 beta subunit. FEBS Lett. 1997, 403, 197–199. [Google Scholar] [CrossRef]
- Guerra, B.; Boldyreff, B.; Sarno, S.; Cesaro, L.; Issinger, O.G.; Pinna, L.A. CK2: A protein kinase in need of control. Pharmacol. Ther. 1999, 82, 303–313. [Google Scholar] [CrossRef]
- Cochet, C.; Chambaz, E.M. Oligomeric structure and catalytic activity of G type casein kinase. Isolation of the two subunits and renaturation experiments. J. Biol. Chem. 1983, 258, 1403–1406. [Google Scholar] [PubMed]
- Kubiński, K.; Domańska, K.; Sajnaga, E.; Mazur, E.; Zieliński, R.; Szyszka, R. Yeast holoenzyme of protein kinase CK2 requires both β and β′ regulatory subunits for its activity. Mol. Cell. Biochem. 2007, 295, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Mortimer, R.K.; Culotti, J.; Culotti, M. Genetic control of the cell division cycle in yeast: V. genetic analysis of cdc mutants. Genetics 1973, 74, 267–286. [Google Scholar] [CrossRef]
- Lee, M.G.; Nurse, P. Complementation used to clone a human homologue of the fission yeast cell cycle control gene cdc2. Nature 1987, 327, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Loewith, R. A brief history of TOR. Biochem. Soc. Trans. 2011, 39, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.; Hall, M. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Padmanabha, R.; Chen-Wu, J.; Hanna, D.; Glover, C. Isolation, sequencing, and disruption of the yeast CKA2 gene: Casein kinase II is essential for viability in Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 4089–4099. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.J.; Veljanoski, F.; O’Doherty, P.J.; Zaman, M.S.; Petersingham, G.; Bailey, T.D.; Munch, G.; Kersaitis, C.; Wu, M.J. Revelation of molecular basis for chromium toxicity by phenotypes of Saccharomyces cerevisiae gene deletion mutants. Metallomics 2016, 8, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Rethinaswamy, A.; Birnbaum, M.J.; Glover, C.V.C. Temperature-sensitive mutations of the CKA1 gene reveal a role for casein kinase II in maintenance of cell polarity in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 5869–5877. [Google Scholar] [CrossRef] [PubMed]
- Blond, O.; Jensen, H.; Buchou, T.; Cochet, C.; Issinger, O.-G.; Boldyreff, B. Knocking out the regulatory b subunit of protein kinase CK2 in mice: Gene dosage effects in ES cells and embryos. Mol. Cell. Biochem. 2005, 274, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Toselli, P.A.; Russell, L.D.; Seldin, D.C. Globozoospermia in mice lacking the casein kinase II a’ catalytic subunit. Nat. Genet. 1999, 23, 118–121. [Google Scholar] [PubMed]
- Lou, D.Y.; Dominguez, I.; Toselli, P.; Landesman-Bollag, E.; O’Brien, C.; Seldin, D.C. The alpha catalytic subunit of protein kinase CK2 is required for mouse embryonic development. Mol. Cell. Biol. 2008, 28, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Trembley, J.H.; Wang, G.; Unger, G.; Slaton, J.; Ahmed, K. Protein kinase CK2 in health and disease: CK2: A key player in cancer biology. Cell. Mol. Life Sci. 2009, 66, 1858–1867. [Google Scholar] [CrossRef] [PubMed]
- Filhol, O.; Nueda, A.; Martel, V.; Gerber-Scokaert, D.; Benitez, M.J.; Souchie, C.; Saoudi, Y.; Cochet, C. Live-cell fluorescence imaging reveals the dynamics of protein kinase CK2 individual subunits. Mol. Cell. Biol. 2003, 23, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Kalathur, M.; Toso, A.; Chen, J.; Revandkar, A.; Danzer-Baltzer, C.; Guccini, I.; Alajati, A.; Sarti, M.; Pinton, S.; Brambilla, L.; et al. A chemogenomic screening identifies CK2 as a target for pro-senescence therapy in PTEN-deficient tumours. Nat. Commun. 2015, 6, 7227. [Google Scholar] [CrossRef] [PubMed]
- Hathaway, G.M.; Lubben, T.H.; Traugh, J.A. Inhibition of casein kinase II by heparin. J. Biol. Chem. 1980, 255, 8038–8041. [Google Scholar] [PubMed]
- Gatica, M.; Hinrichs, M.V.; Jedlicki, A.; Allende, C.C.; Allende, J.E. Effect of metal ions on the activity of cascein kinase II from Xenopus laevis. FEBS Lett. 1993, 315, 173–177. [Google Scholar] [CrossRef]
- Hathaway, G.M.; Traugh, J.A. Interaction of polyamines and magnesium with casein kinase II. Arch. Biochem. Biophys. 1984, 233, 133–138. [Google Scholar] [CrossRef]
- Jiménez, J.; Benítez, M.; Lechuga, C.; Collado, M.; González-Nicólas, J.; Moreno, F. Casein kinase 2 inactivation by Mg2+, Mn2+ and Co2+ ions. Mol. Cell. Biochem. 1995, 152, 1–6. [Google Scholar] [PubMed]
- Botstein, D.; Fink, G.R. Yeast: An experimental organism for 21st century biology. Genetics 2011, 189, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Goffeau, A.; Barrell, B.; Bussey, H.; Davis, R.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.; Jacq, C.; Johnston, M. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Arita, A.; Zhou, X.; Ellen, T.P.; Liu, X.; Bai, J.; Rooney, J.P.; Kurtz, A.; Klein, C.B.; Dai, W.; Begley, T.J.; et al. A genome-wide deletion mutant screen identifies pathways affected by nickel sulfate in Saccharomyces cerevisiae. BMC Genom. 2009, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, M.; Pagano, L.; Pasquali, F.; Zappettini, A.; Tosato, V.; Bruschi, C.V.; Marmiroli, N. A genome-wide nanotoxicology screen of Saccharomyces cerevisiae mutants reveals the basis for cadmium sulphide quantum dot tolerance and sensitivity. Nanotoxicology 2015, 10, 84–93. [Google Scholar] [PubMed]
- Johnson, A.J.; Veljanoski, F.; O’Doherty, P.J.; Zaman, M.S.; Petersingham, G.; Bailey, T.D.; Munch, G.; Kersaitis, C.; Wu, M.J. Molecular insight into arsenic toxicity via the genome-wide deletion mutant screening of Saccharomyces cerevisiae. Metallomics 2016, 8, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, M.; Perrone, G.; Kristiansson, E.; Traini, M.; Ye, T.; Dawes, I.; Nerman, O.; Tamas, M. Genetic basis of arsenite and cadmium tolerance in Saccharomyces cerevisiae. BMC Genom. 2009, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Cao, C.; Jiang, L. Genome-scale genetic screen of lead ion-sensitive gene deletion mutations in Saccharomyces cerevisiae. Gene 2015, 563, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Tun, N.M.; Lennon, B.R.; O’Doherty, P.J.; Johnson, A.J.; Petersingham, G.; Bailey, T.D.; Kersaitis, C.; Wu, M.J. Effects of metal ions and hydrogen peroxide on the phenotype of yeast hom6Δ mutant. Lett. Appl. Microbiol. 2015, 60, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.H.; Laurent, J.M.; Yellman, C.M.; Meyer, A.G.; Wilke, C.O.; Marcotte, E.M. Systematic humanization of yeast genes reveals conserved functions and genetic modularity. Science 2015, 348, 921–925. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.P.; Remm, M.; Sonnhammer, E.L.L. Inparanoid: A comprehensive database of eukaryotic orthologs. Nucleic Acids Res. 2005, 33, D476–D480. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.A.; Bedalov, A. Yeast as a model system for anticancer drug discovery. Nat. Rev. Cancer 2004, 4, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Mager, W.H.; Winderickx, J. Yeast as a model for medical and medicinal research. Trends Pharmacol. Sci. 2005, 26, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.S.; Johnson, A.J.; Bobek, G.; Kueh, S.; Kersaitis, C.; Bailey, T.D.; Buskila, Y.; Wu, M.J. Protein kinase CK2 regulates metal toxicity in neuronal cells. Metallomics 2016, 8, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Kelleher, S.L. Cellular mechanisms of zinc dysregulation: A perspective on zinc homeostasis as an etiological factor in the development and progression of breast cancer. Nutrients 2012, 4, 875–903. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Ho, E. Zinc and prostatic cancer. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 640–645. [Google Scholar]
- Lightman, A.; Brandes, J.M.; Binur, N.; Drugan, A.; Zinder, O. Use of the serum copper/zinc ratio in the differential diagnosis of ovarian malignancy. Clin. Chem. 1986, 32, 101–103. [Google Scholar] [PubMed]
- Li, M.; Zhang, Y.; Liu, Z.; Bharadwaj, U.; Wang, H.; Wang, X.; Zhang, S.; Liuzzi, J.P.; Chang, S.-M.; Cousins, R.J. Aberrant expression of zinc transporter ZIP4 (SLC39A4) significantly contributes to human pancreatic cancer pathogenesis and progression. Proc. Natl. Acad. Sci. USA 2007, 104, 18636–18641. [Google Scholar] [CrossRef] [PubMed]
- Ebara, M.; Fukuda, H.; Hatano, R.; Saisho, H.; Nagato, Y.; Suzuki, K.; Nakajima, K.; Yukawa, M.; Kondo, F.; Nakayama, A. Relationship between copper, zinc and metallothionein in hepatocellular carcinoma and its surrounding liver parenchyma. J. Hepatol. 2000, 33, 415–422. [Google Scholar] [CrossRef]
- Eide, D.J. The molecular biology of metal ion transport in Saccharomyces cerevisiae. Annu. Rev. Nutr. 1998, 18, 441–469. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N. Metal ion transporters and homeostasis. EMBO J. 1999, 18, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Zatta, P.; Drago, D.; Bolognin, S.; Sensi, S.L. Alzheimer’s disease, metal ions and metal homeostatic therapy. Trends Pharmacol. Sci. 2009, 30, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.H.; Dunlap, P.E.; McBride, S.J.; Al-Refai, H.; Bushel, P.R.; Freedman, J.H. Global transcriptome and deletome profiles of yeast exposed to transition metals. PLoS Genet. 2008, 4, e1000053. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Danku, J.M.; Baxter, I.; Kim, S.; Vatamaniuk, O.; Vitek, O.; Ouzzani, M.; Salt, D. High-resolution genome-wide scan of genes, gene-networks and cellular systems impacting the yeast ionome. BMC Genom. 2012, 13, 623. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Shi, X. Gene expression profile in response to chromium-induced cell stress in A549 cells. Mol. Cell. Biochem. 2001, 222, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Liu, G.; He, Z.; Ma, W.Y.; Bode, A.M.; Dong, Z. Arsenite inhibits p53 phosphorylation, DNA binding activity, and p53 target gene p21 expression in mouse epidermal JB6 cells. Mol. Carcinog. 2006, 45, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Bachhuber, T.; Almaça, J.; Aldehni, F.; Mehta, A.; Amaral, M.D.; Schreiber, R.; Kunzelmann, K. Regulation of the epithelial Na+ channel by the protein kinase CK2. J. Biol. Chem. 2008, 283, 13225–13232. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, D.; Villella, V.; Esposito, S.; Tosco, A.; Sepe, A.; de Gregorio, F.; Salvadori, L.; Grassia, R.; Leone, C.; de Rosa, G.; et al. Restoration of CFTR function in patients with cystic fibrosis carrying the F508del-CFTR mutation. Autophagy 2014, 10, 2053–2074. [Google Scholar] [CrossRef] [PubMed]
- Venerando, A.; Pagano, M.A.; Tosoni, K.; Meggio, F.; Cassidy, D.; Stobbart, M.; Pinna, L.A.; Mehta, A. Understanding protein kinase CK2 mis-regulation upon F508del CFTR expression. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 384, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Treharne, K.J.; Xu, Z.; Chen, J.-H.; Best, O.G.; Cassidy, D.M.; Gruenert, D.C.; Hegyi, P.; Gray, M.A.; Sheppard, D.N.; Kunzelmann, K.; et al. Inhibition of protein kinase CK2 closes the CFTR Cl− channel, but has no effect on the cystic fibrosis mutant ΔF508-CFTR. Cell. Physiol. Biochem. 2009, 24, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.K.; Saydam, N.; Steiner, F.; Schaffner, W.; Freedman, J.H. Activation of gene expression by metal-responsive signal transduction pathways. Environ. Health Perspect. 2002, 110 (Suppl. 5), S813–S817. [Google Scholar] [CrossRef]
- Xue, Y.; Ren, J.; Gao, X.; Jin, C.; Wen, L.; Yao, X. GPS 2.0, a tool to predict kinase-specific phosphorylation sites in hierarchy. Mol. Cell. Proteom. 2008, 7, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, B.; Kuo, Y.-M.; Zemansky, J.; Gitschier, J. A novel member of a zinc transporter family is defective in acrodermatitis enteropathica. Am. J. Hum. Genet. 2002, 71, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Muraina, I.A.; Brethour, D.; Schmitt-Ulms, G.; Nimmanon, T.; Ziliotto, S.; Kille, P.; Hogstrand, C. Zinc transporter ZIP10 forms a heteromer with ZIP6 which regulates embryonic development and cell migration. Biochem. J. 2016, 473, 2531–2544. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Ulms, G.; Ehsani, S.; Watts, J.C.; Westaway, D.; Wille, H. Evolutionary descent of prion genes from the ZIP family of metal ion transporters. PLoS ONE 2009, 4, e7208. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Tepaamorndech, S. The SLC30 family of zinc transporters—A review of current understanding of their biological and pathophysiological roles. Mol. Asp. Med. 2013, 34, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Fu, D. Structure of the zinc transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef] [PubMed]
- Monteith, G.R.; Davis, F.M.; Roberts-Thomson, S.J. Calcium channels and pumps in cancer: Changes and consequences. J. Biol. Chem. 2012, 287, 31666–31673. [Google Scholar] [CrossRef] [PubMed]
- Mahe, I.; Chassany, O.; Grenard, A.S.; Caulin, C.; Bergmann, J.F. Defining the role of calcium channel antagonists in heart failure due to systolic dysfunction. Am. J. Cardiovasc. Drugs 2003, 3, 33–41. [Google Scholar] [PubMed]
- Inzitari, M.; Di Bari, M.; Marchionni, N. Calcium channel blockers and coronary heart disease. Aging Clin. Exp. Res. 2005, 17, S6–S15. [Google Scholar]
- Chai, F.; Truong-Tran, A.Q.; Ho, L.H.; Zalewski, P.D. Regulation of caspase activation and apoptosis by cellular zinc fluxes and zinc deprivation: A review. Immunol. Cell Biol. 1999, 77, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-L.; Hung, T.-C.; Hsieh, B.-S.; Chen, Y.-H.; Chen, T.-F.; Cheng, H.-L. Zinc at pharmacologic concentrations affects cytokine expression and induces apoptosis of human peripheral blood mononuclear cells. Nutrition 2006, 22, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Costello, L.C.; Franklin, R.B. The clinical relevance of the metabolism of prostate cancer; zinc and tumor suppression: Connecting the dots. Mol. Cancer 2006, 5, 17. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, Y.; Zhu, X.; Zhu, J.; Liao, S.; Tang, Q.; Liu, K.; Guan, X.; Zhang, J.; Feng, Z. Identification of differential expression of genes in hepatocellular carcinoma by suppression subtractive hybridization combined cDNA microarray. Oncol. Rep. 2007, 18, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.N.; Lee, Y.S.; Kim, M.Y.; Kim, J.D.; Park, L.O. Antiproliferative and apoptotic effects of zinc–citrate compound (CIZAR®) on human epithelial ovarian cancer cell line, OVCAR-3. Gynecol. Oncol. 2006, 103, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.-Y.; Liu, Y.-Y.; Zou, J.; Franklin, R.B.; Costello, L.C.; Feng, P. Inhibitory effect of zinc on human prostatic carcinoma cell growth. Prostate 1999, 40, 200–207. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Y.; Nguyen, Q.; Novikoff, P.M.; Czaja, M.J. Induction of hepatoma cell apoptosis by c-myc requires zinc and occurs in the absence of DNA fragmentation. Am. J. Physiol. Gastrointest. Liver Physiol. 1996, 270, G60–G70. [Google Scholar]
- Margalioth, E.J.; Schenker, J.G.; Chevion, M. Copper and zinc levels in normal and malignant tissues. Cancer 1983, 52, 868–872. [Google Scholar] [CrossRef]
- Rizk, S.L.; Sky-Peck, H.H. Comparison between concentrations of trace elements in normal and neoplastic human breast tissue. Cancer Res. 1984, 44, 5390–5394. [Google Scholar] [PubMed]
- Kagara, N.; Tanaka, N.; Noguchi, S.; Hirano, T. Zinc and its transporter ZIP10 are involved in invasive behavior of breast cancer cells. Cancer Sci. 2007, 98, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Manning, D.; Robertson, J.; Ellis, I.; Elston, C.; McClelland, R.A.; Gee, J.M.W.; Jones, R.; Green, C.; Cannon, P.; Blamey, R. Oestrogen-regulated genes in breast cancer: Association of pLIV1 with lymph node involvement. Eur. J. Cancer 1994, 30, 675–678. [Google Scholar] [CrossRef]
- Manning, D.L.; McClelland, R.A.; Knowlden, J.M.; Bryant, S.; Gee, J.M.; Green, C.D.; Robertson, J.F.; Blamey, R.W.; Sutherland, R.L.; Ormandy, C.J. Differential expression of oestrogen regulated genes in breast cancer. Acta Oncol. 1995, 34, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Morgan, H.E.; Smart, K.; Zahari, N.M.; Pumford, S.; Ellis, I.O.; Robertson, J.F.; Nicholson, R.I. The emerging role of the LIV-1 subfamily of zinc transporters in breast cancer. Mol. Med. 2007, 13, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Donadelli, M.; Dalla Pozza, E.; Costanzo, C.; Scupoli, M.; Scarpa, A.; Palmieri, M. Zinc depletion efficiently inhibits pancreatic cancer cell growth by increasing the ratio of antiproliferative/proliferative genes. J. Cell. Biochem. 2008, 104, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.B.; Costello, L.C. The important role of the apoptotic effects of zinc in the development of cancers. J. Cell. Biochem. 2009, 106, 750–757. [Google Scholar] [CrossRef] [PubMed]


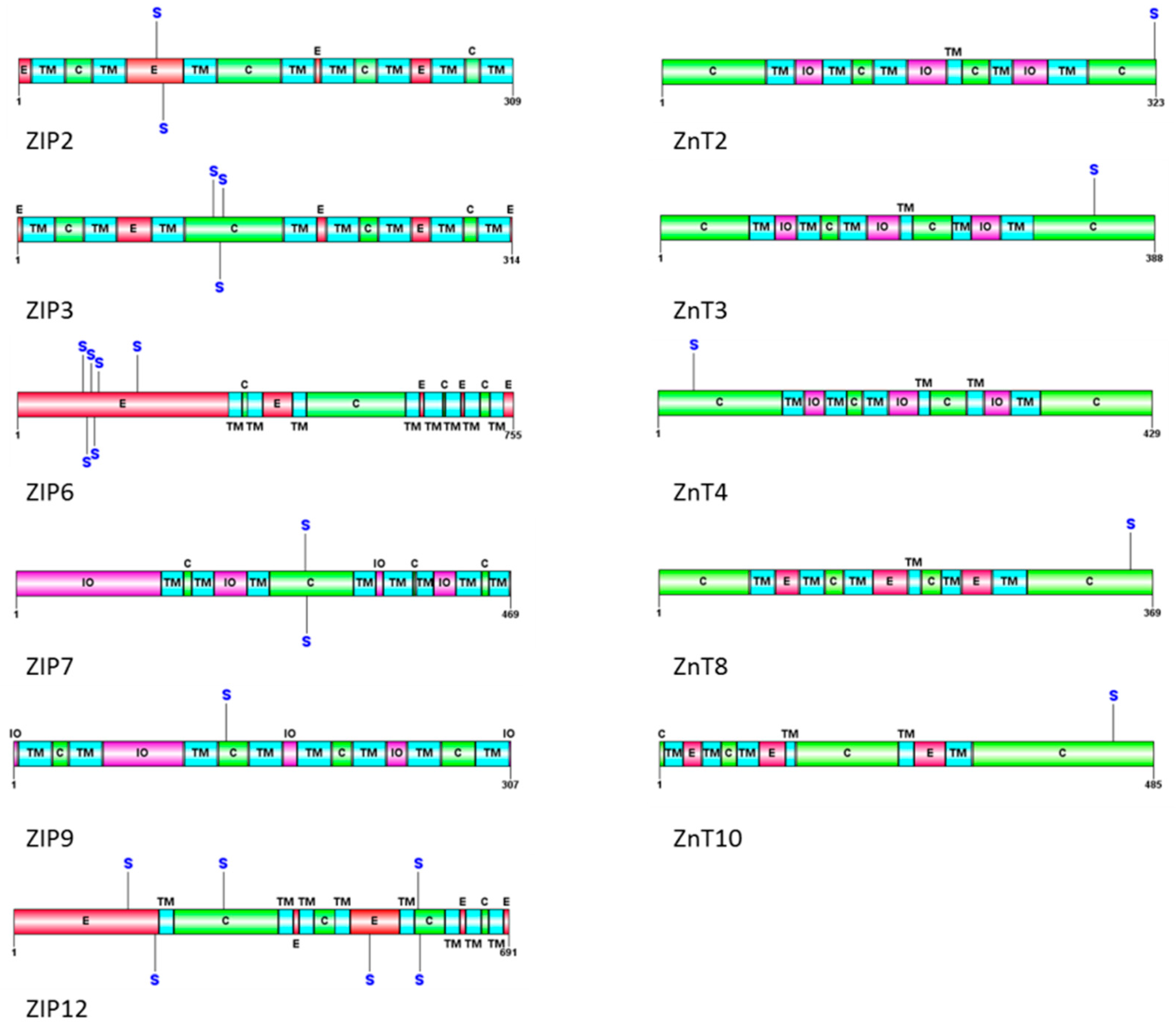{kind=link}
| Protein | Site | Exemplar Sequence | Score | Location of Phosphorylation Site |
|---|---|---|---|---|
| ZIP1 | None | - | - | - |
| ZIP2 | S87 | MVQNRSASERNSSGD | 10.179 | Extracellular |
| ZIP2 | S91 | RSASERNSSGDADSA | 15.628 | Extracellular |
| ZIP3 | S125 | LETFNAGSDVGSDSE | 10.216 | Cytoplasmic |
| ZIP3 | S129 | NAGSDVGSDSEYESP | 16.602 | Cytoplasmic |
| ZIP3 | S131 | GSDVGSDSEYESPFM | 12.602 | Cytoplasmic |
| ZIP4 | None | - | - | - |
| ZIP5 | None | - | - | - |
| ZIP6 | S100 | HHDHDHHSDHEHHSD | 11.864 | Extracellular |
| ZIP6 | S106 | HSDHEHHSDHERHSD | 12.672 | Extracellular |
| ZIP6 | S112 | HSDHERHSDHEHHSE | 12.662 | Extracellular |
| ZIP6 | S118 | HSDHEHHSEHEHHSD | 12.807 | Extracellular |
| ZIP6 | S124 | HSEHEHHSDHDHHSH | 12.309 | Extracellular |
| ZIP6 | S183 | RNVKDSVSASEVTST | 10.324 | Extracellular |
| ZIP7 | S275 | RSTKEKQSSEEEEKE | 16.548 | Cytoplasmic |
| ZIP7 | S276 | RSTKEKQSSEEEEKE | 16.548 | Cytoplasmic |
| ZIP8 | None | - | - | - |
| ZIP9 | S132 | IGNSHVHSTDDPEAA | 11.066 | Cytoplasmic |
| ZIP10 | None | - | - | - |
| ZIP11 | None | - | - | - |
| ZIP12 | S160 | DEDSSFLSQNETEDI | 10.412 | Extracellular |
| ZIP12 | S197 | KKSGIVSSEGANEST | 10.888 | Extracellular |
| ZIP12 | S293 | QDYSNFSSSMEKESE | 11.826 | Cytoplasmic |
| ZIP12 | S497 | LALNSELSDQAGRGK | 9.983 | Extracellular |
| ZIP12 | S565 | AIGAAFSSSSESGVT | 10.276 | Cytoplasmic |
| ZIP12 | S567 | GAAFSSSSESGVTTT | 10.409 | Cytoplasmic |
| ZIP13 | None | - | - | - |
| ZIP14 | None | - | - | - |
| ZNT1 | None | - | - | - |
| ZNT2 | S322 | CQACQGPSD | 10.147 | Cytoplasmic |
| ZNT3 | S341 | SAHLAIDSTADPEAV | 9.921 | Cytoplasmic |
| ZNT4 | S32 | DTSAFDFSDEAGDEG | 13.229 | Cytoplasmic |
| ZNT5 | None | - | - | - |
| ZNT6 | None | - | - | - |
| ZNT7 | None | - | - | - |
| ZNT8 | S353 | SLTIQMESPVDQDPD | 10.017 | Cytoplasmic |
| ZNT9 | None | - | - | - |
| ZNT10 | S446 | TYGSDGLSRRDAREV | 11.562 | Cytoplasmic |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.J.; Wu, M.J. The New Role for an Old Kinase: Protein Kinase CK2 Regulates Metal Ion Transport. Pharmaceuticals 2016, 9, 80. https://doi.org/10.3390/ph9040080
Johnson AJ, Wu MJ. The New Role for an Old Kinase: Protein Kinase CK2 Regulates Metal Ion Transport. Pharmaceuticals. 2016; 9(4):80. https://doi.org/10.3390/ph9040080
Chicago/Turabian StyleJohnson, Adam J., and Ming J. Wu. 2016. "The New Role for an Old Kinase: Protein Kinase CK2 Regulates Metal Ion Transport" Pharmaceuticals 9, no. 4: 80. https://doi.org/10.3390/ph9040080
APA StyleJohnson, A. J., & Wu, M. J. (2016). The New Role for an Old Kinase: Protein Kinase CK2 Regulates Metal Ion Transport. Pharmaceuticals, 9(4), 80. https://doi.org/10.3390/ph9040080