Abstract
The liver enzyme matriptase-2 is a multi-domain, transmembrane serine protease with an extracellular, C-terminal catalytic domain. Synthetic low-molecular weight inhibitors of matriptase-2 have potential as therapeutics to treat iron overload syndromes, in particular in patients with β-thalassemia. A sub-library of 64 compounds was screened for matriptase-2 inhibition and several active compounds were identified. (S)-Ethyl 2-(benzyl(3-((4-carbamidoylphenoxy)methyl)-2,3-dihydrobenzo[b][1,4]dioxin-6-yl)amino)-2-oxoacetate ((S)-12) showed an IC50 value of less than 10 µM. Structure-activity relationships were discussed and proposals to design new matriptase-2 inhibitors were made.
1. Introduction
Thalassemias are among the most common inherited diseases worldwide. They are classified as anaemia and typified by abnormal formation of hemoglobin [1]. One type of disease, β-thalassemia, is characterized by a decreased synthesis of β-globin chains or by the complete lack of it, resulting in a severe anaemia and/or red blood cell abnormalities. The imbalance between the amount of α- and β-globin chain leads to extra medullary expansion and splenomegaly [1]. Patients affected by β-thalassemia major, the most severe form, require chronic red blood cell transfusions. As a result, they develop secondary iron overload. The milder form, β-thalassemia intermedia, does not necessitate blood transfusions, but also leads to iron overload due to chronic suppression of the hepcidin synthesis caused by ineffective erythropoiesis, thereby leading to increased iron absorption in the duodenum [1,2,3,4]. Untreated iron overload causes liver cirrhosis, cardiomyopathy, diabetes, arthritis, hypogonadism, and skin pigmentation and is the main reason for death in these individuals. In other diseases that are correlated with primary iron overload, like HFE-associated hemochromatosis, iron accumulation is hindered by phlebotomy, but this is not possible in the case of β-thalassemia. Therefore, patients must be treated with iron chelation therapy, in most cases through the subcutaneous application of desferoxamine.
Hepcidin, a small hepatic peptide hormone, has a crucial role in iron homeostasis in the human body [2,3]. Hepcidin negatively regulates intestinal iron absorption, iron recycling from macrophages and iron release from hepatic stores and macrophages into the plasma [3,5]. The expression of hepcidin is regulated by the BMP-SMAD pathway. Bone morphogenetic proteins (BMPs) are part of the transforming growth factor-β superfamily of ligands [3,4]. Two factors play a crucial role in the iron homeostasis, BMP6 and hemojuvelin. Hemojuvelin is a glycophosphatidylinositol-membrane-anchored co-receptor, detected mostly in hepatic cells [3,4]. It uses the BMP type I receptors ALK2 and ALK3 to transfer signals as a response to BMP6 [6]. The ligand BMP6 and the BMP-receptor complex activate the SMAD1,5,8/SMAD4 (sons of mother against decapentaplegic) complex. SMAD4 is translocated to the nucleus, where it is recognized by the hepcidin promoter [3,6], which is responsible for the hepcidin expression.
Matriptase-2, encoded by the gene TMPRSS6, known as a member of the type II transmembrane serine protease family, is located mainly at the cell surface of hepatocytes. The structure of this enzyme contains a cytoplasmic N-terminal domain, a transmembrane domain, a SEA (sea-urchin sperm protein, enteropeptidase and agrin) domain, two CUB (complement factor C1s/C1r, urchin embryonic growth factor, bone morphogenetic protein) domains, three LDLRA (low density lipoprotein receptor class A) domains and a C-terminal serine protease domain [7,8,9,10,11].
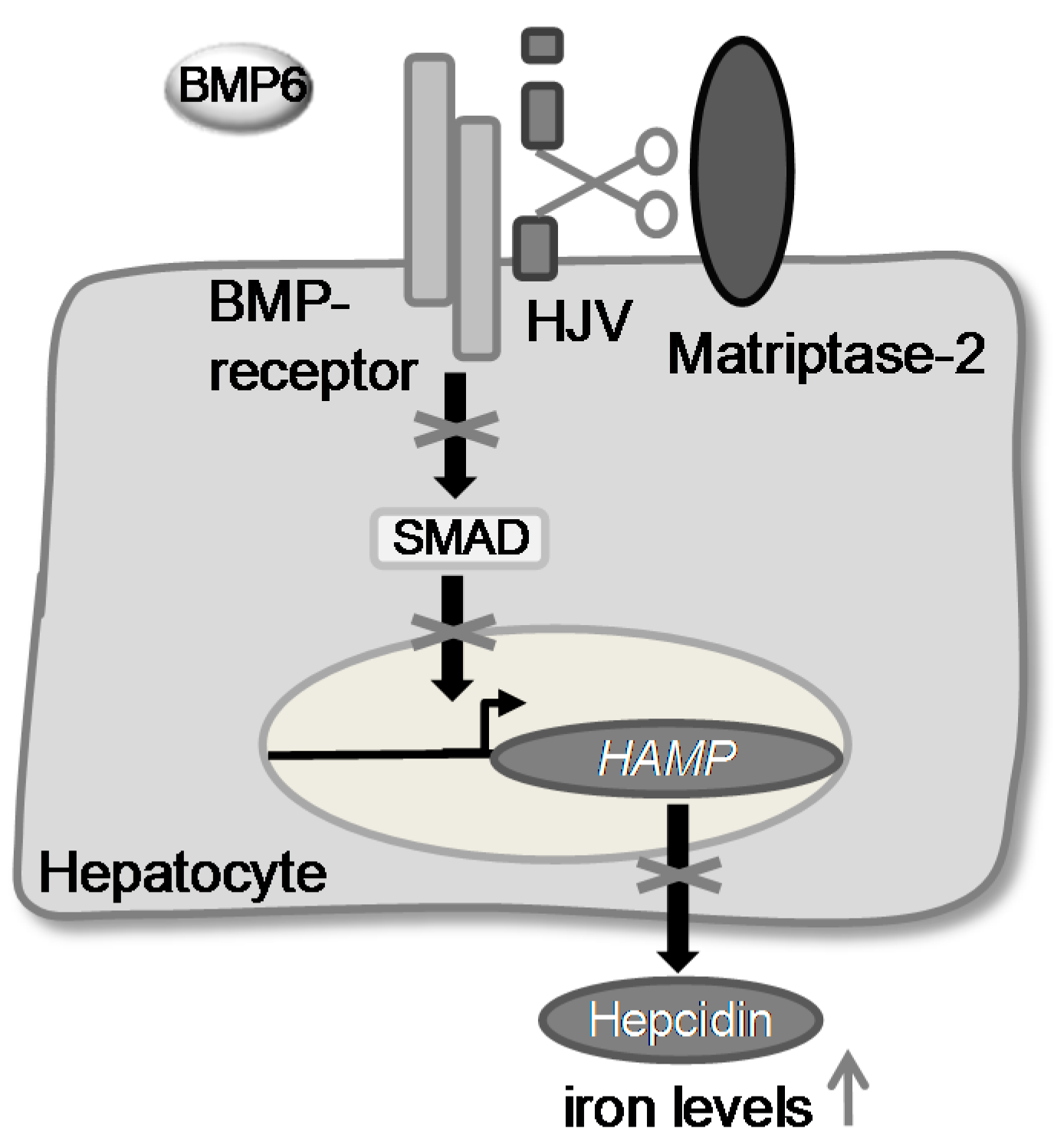
Recently, it was shown that matriptase-2 represents a key enzyme in iron homeostasis [12,13,14]. Mutations in the TMPRSS6 gene were found to cause iron-refractory iron deficieny anaemia (IRIDA) [17]. It was demonstrated that matriptase-2 acts as a suppressor of the expression of the hepatic hormone hepcidin. It probably inactivates the bone morphogenetic protein co-receptor hemojuvelin (m-HJV) by cleaving it into an inactive form [10,15]. As a consequence, the phosphorylation of SMADs (sons of mothers against decapentaplegic homologue) is suppressed and therefore the expression of HAMP, the gene encoding hepcidin, decreases. This leads to a higher level of iron in the blood plasma (Figure 1).
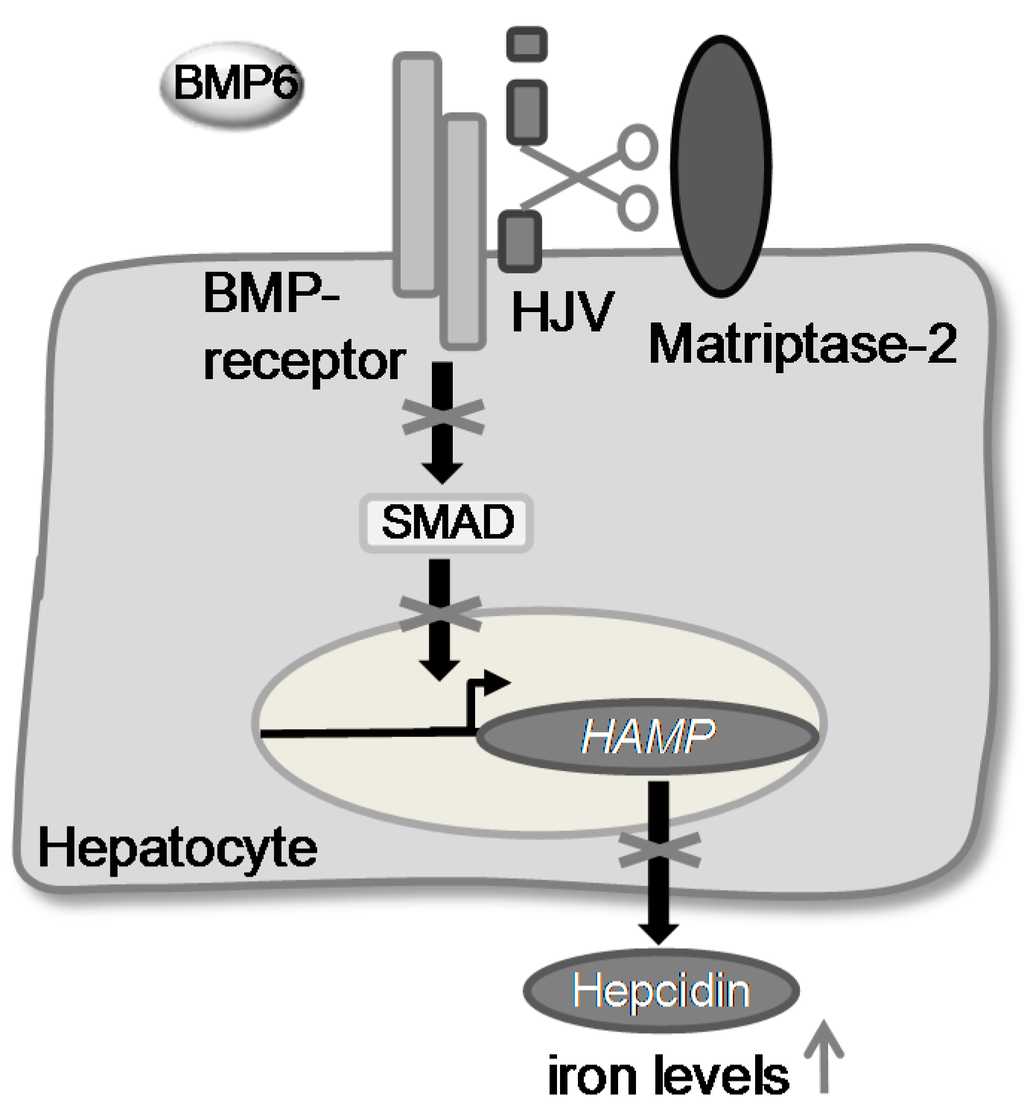
Figure 1.
The postulated role of matriptase-2 in iron homeostasis.
Figure 1.
The postulated role of matriptase-2 in iron homeostasis.
Several lines of evidence indicate that matriptase-2, as a trypsin-like serine protease, has a specificity to cleave the peptide bond after basic amino acids. For example, putative cleavage sites in hemojuvelin as well as autoprocessing cleavage sites feature arginine in the P1 position [10,11,15,16,17]. A preferred P4–P1 substrate sequence (Ile–Arg–Ala–Arg), obtained by a combinatorial approach, confirmed this primary substrate specificity [18], which is facilitated by the negatively charged aspartyl side chain at the bottom of their S1 pocket, able to interact with positively charged moieties, e.g., of arginine or arginine mimetics. Moreover, the S3/S4 region of matriptase-2 has also been found to be occupied by positively charged ligand moieties [18,19,20].
Inhibitors of matriptase-2 have potential as therapeutic compounds to treat iron overload syndromes, which are present in β-thalassemia patients [21,22,23,24]. Therefore, matriptase-2 is a promising pharmaceutical target for the development of synthetic inhibitors [10]. Meanwhile, first reports on low-molecular weight inhibitors of matriptase-2 have appeared, including dipeptide amides with a amidinobenzylamide residue [25], amidinophenylalanine derivatives [26], peptidic ketones [27], and sunflower trypsin inhibitor-1 analogues [28]. Moreover, bis- and trisbenzamidines [19,20] have been reported as matriptase-2 inhibitors, so former type of compounds also being valued as antiprotozoal and antifungal agents [29,30,31]. This study attempted to provide further heterocyclic, non-peptidic matriptase-2 inhibitors. By taking the primary substrate specificity of matriptase-2 into account, a focused screening approach was used and is described herein.
2. Results and Discussion
A sub-library of 64 compounds was evaluated for inhibition of matriptase-2. Structures of relevant compounds are shown in Table 1 and Table 2. Two classes were identified which include active compounds. The first class comprises 4H-3,1-benzothiazin-4-ones. The corresponding data are listed in Table 1. 4H-3,1-Benzothiazin-4-ones have previously been reported to exhibit dual activities as adenosine receptors antagonists and inhibitors of monoamine oxidase B [32,33]. Moreover, certain members act as oxoeicosanoid receptor ligands [34]. The structure of 4H-3,1-benzothiazin-4-ones can be considered to result from a bioisosteric replacement of the ring oxygen by sulfur. The oxygen analogues, 4H-3,1-benzoxazin-4-ones, have attracted attention as inhibitors for serine proteases, for example for human leukocyte elastase or chymase [35,36], but are less stable against unspecific nucleophiles than the 4H-3,1-benzothiazin-4-ones [37].
As a first representative of matriptase-2 inhibiting 4H-3,1-benzothiazin-4-ones, we identified compound 1 (Table 1). This molecule contains a basic side chain at position 6 of the heterocyclic skeleton. It was assumed that this basic group might be able to interact with the S1 pocket of matriptase-2. Thus, all 4H-3,1-benzothiazin-4-ones available in our library were considered and those compounds were selected which bear a basic residue at optional positions of the heterocyclic scaffold. Derivatives closely related to 1 were, however, found to be inactive. For example, a shift of the residue at position 6 to position 7 led to a loss of activity (1 versus 2). Compound 5 with an extended 6-residue and the more embedded basic nitrogen was also inactive, as the positively charged group might be prevented from interacting with the S1 pocket. We have evaluated three 4H-3,1-benzothiazin-4-ones which bear a basic moiety within the 2-substituent. While the presence of a primary amine structure in 7 yielded a second, active compound, derivatives 8 and 9 with tertiary amine substructures were inactive.
The second class of test compounds from which we have identified active representatives mainly consists of heterocycles which exclusively contain a benzamidine moiety. The benzamidine group is known to be efficiently accommodated in the S1 pocket of trypsin-like serine proteases. Benzamidine itself was also tested in the course of this study as an inhibitor of matriptase-2, but exhibited only weak activity with an IC50 value of more than 400 µM. The structures of the benzamidine-containing heterocycles and their IC50 values for matriptase-2 inhibition are outlined in Table 2. The first five entries include 2,3-dihydro-1,4-benzodioxines ((S)-10 to (S)-12). These members have previously been reported to exhibit a dual activity, against thrombin and the fibrinogen receptor αIIbβ3, with inhibition of the latter target producing an anti-platelet activity. Besides thrombin, activity against related serine proteases, e.g., trypsin and factor Xa, has also been identified. The enantiomers (S)-10 and (R)-10 as well as (S)-11 and (R)-11, represent 6- and 7-substituted isomers [38].
Compounds 13–21 are racemic 3,4-dihydro-2H-1,4-benzoxazine derivatives with an oxymethylene spacer connecting the heterocyclic core with a para-benzamidine moiety. In 21, the direction of spacer is inverted. Compounds 13–21 bear different residues, either at the 6 or 7 position, with fluorinated aryl groups as a typical substructure present in 13–18 [39,40,41]. These compounds have also been evaluated towards thrombin, trypsin, factor Xa and at the fibrinogen receptor. Except for 14, the fluorinated derivatives showed a strong thrombin inhibition and, moreover, 13, 15 and 16 inhibited thrombin better than trypsin and factor Xa. [39,41]. Compound 18 was also characterized with respect radical scavenging activity, lipid peroxidation of linoleic acid and lipoxygenase inhibition [40]. The last five entries in Table 2 comprise benzamidine derivatives with more dissimilar structures [41,42,43]. Compound 21 has an anilide substructure, 22 and 23 bear other residues than methyl at position 4, and (R)-24 and 25 lack the 3,4-dihydro-2H-1,4-benzoxazine scaffold.

Table 1.
Matriptase-2 inhibition by 4H-3,1-benzothiazin-4-ones.
| Compd | Structure | IC50 (µM) ± SEM a |
|---|---|---|
| 1 [34] | 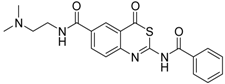 | 148 ± 28 |
| 2 [34] | 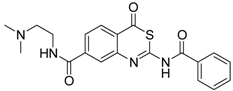 | >160 |
| 3 [32] | 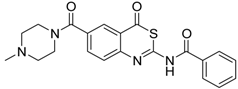 | >160 |
| 4 [32] | 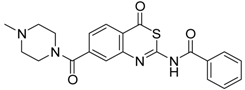 | >160 |
| 5 [32] | 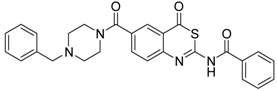 | >160 |
| 6 [32] | 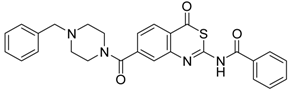 | >160 |
| 7 [32] | 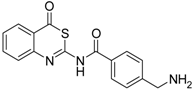 | 119 ± 9 |
| 8 [32] | 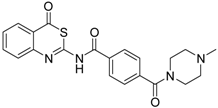 | >160 |
| 9 [32] | 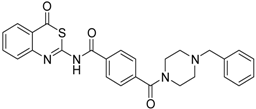 | >160 |
a Values with SEM refer to duplicate measurements with five different inhibitor concentrations. Limits refer to duplicate measurements with a single inhibitor concentration of 40 µM.
Table 2.
Matriptase-2 inhibition by 2,3-dihydro-1,4-benzodioxines and 3,4-dihydro-2H-1,4-benzoxazines.
| Compd. | Structure | IC50 (µM) ± SEM a |
|---|---|---|
| (S)-10 [38] | 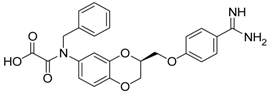 | 16.7 ± 1.7 |
| (R)-10 [38] | 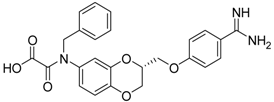 | 27.3 ± 2.0 |
| (S)-11 [38] | 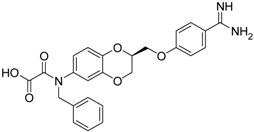 | 29.8 ± 1.6 |
| (R)-11 [38] | 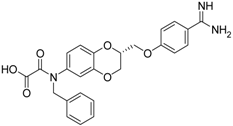 | 33.6 ± 1.1 |
| (S)-12 [38] | 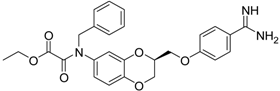 | 8.47 ± 0.76 |
| 13 [39] | 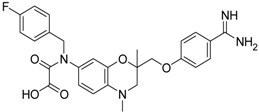 | >40 |
| 14 [39] | 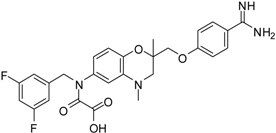 | 35.8 ± 1.1 |
| 15 [39] | 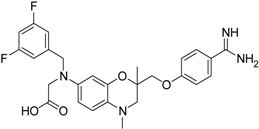 | 25.8 ± 4.0 |
| 16 [39] | 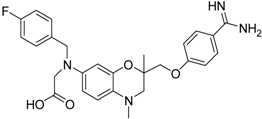 | 13.6 ± 2.31 |
| 17 [39] | 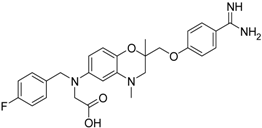 | 11.2 ± 1.61 |
| 18 [39,40] | 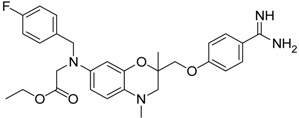 | 31.6 ± 2.69 |
| 19 [41] | 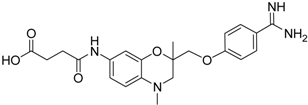 | 38.8 ± 3.14 |
| 20 [41] | 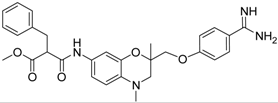 | 31.4 ± 3.6 |
| 21 [41] | 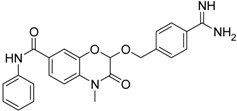 | 42.6 ± 4.7 |
| 22 [41] | 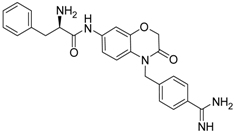 | 30.9 ± 3.4 |
| 23 [42] | 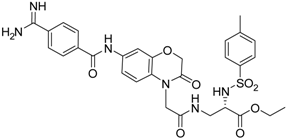 | 20.5 ± 2.0 |
| (R)-24 [43] | 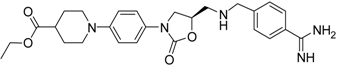 | >40 |
| 25 b | 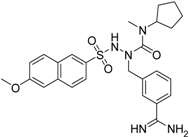 | >40 |
a Values with SEM refer to duplicate measurements with five different inhibitor concentrations. Limits refer to duplicate measurements with a single inhibitor concentration of 40 µM; b Compound 25 was prepared using a protocol described in reference [44].
In the course of this study, we identified several benzamidine-substituted heterocycles as inhibitors of matriptase-2. For one of these active compounds, 19, the influence of the substrate concentration on the inhibition was assessed. The Lineweaver-Burk plot is shown in Figure 2. Unexpectedly, compound 19 did not behave as a competitive inhibitor, but showed a mixed type of inhibition.
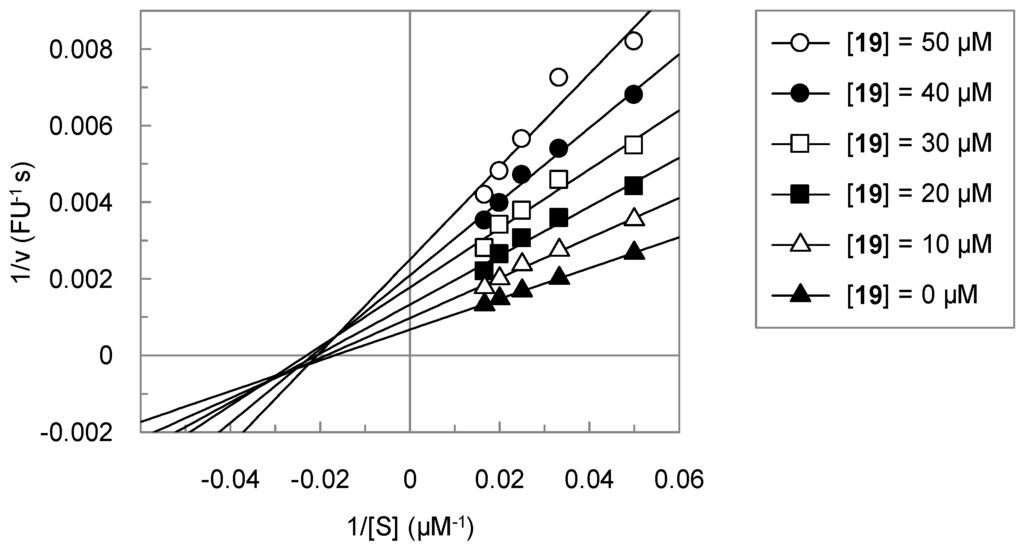
Figure 2.
Double reciprocal plot for the inhibition of matriptase-2 by 19. Substrate concentrations of 20, 30, 40, 50 and 60 µM were used.
Figure 2.
Double reciprocal plot for the inhibition of matriptase-2 by 19. Substrate concentrations of 20, 30, 40, 50 and 60 µM were used.
Among the active compounds, (S)-12 was found to be a potent inhibitor of matriptase-2 with an IC50 value of 8.47 µM. A comparison of the activity of the two analogues (S)-12 and (S)-10 revealed a slightly stronger activity of the oxamic ester (S)-12 than that of the oxamic acid (S)-10. Among the pairs of enantiomers, the (S)-configuration was somewhat preferred for matriptase-2 inhibition ((S)-10 versus (R)-10 and (S)-11 versus (R)-11). It should be noted that (S)-12 was described to be a highly potent thrombin inhibitor [38]. However, thrombin inhibition is not always accompanied by matriptase-2 inhibition. For example, 13 was inactive at matriptase-2, but highly active at thrombin [39]. Among the 3,4-dihydro-2H-1,4-benzoxazines with a methyl group at 4-position (13–21), several members inhibited matriptase-2 with IC50 values of less than 30 µM. The presence of an oxamate moiety (in 13 and 14) appeared to be less favorable. This could be concluded from the results of the inactive compound 13 and of 16 (IC50 = 13.6 µM). The higher flexibility of the glycine substructure (in 15–17) compared to the oxamate substructure (in 13 and 14) might account for this effect. The position of the N-substituted glycine moiety, as either 7- or 6-substituent, did not exert a remarkable influence on matriptase-2 inhibition (16 versus 17).
The common feature of the fluorine-free compounds 19 and 20 is the NHCO group at position 7. Both compounds were moderately active. The 3,4-dihydro-2H-1,4-benzoxazine derivatives 21–23 did not show an improved inhibitory activity, and (R)-24 and 25 were inactive. The finding that the latter two compounds did not affect matriptase-2 activity indicated that the presence of a benzamidine moiety does not necessarily lead to matriptase-2 inhibition. This was in accordance with the lack of inhibitory activity of benzamidine itself. On the one hand, the absence of the benzo-fused heterocyclic core in (R)-24 and 25 was obviously unfavorable. On the other hand, since the majority of 2,3-dihydro-1,4-benzodioxines and 3,4-dihydro-2H-1,4-benzoxazines were active, these scaffolds are suitable for the positioning of various residues and for directing them to the target’s binding pockets.
In summary, representatives of three heterocyclic classes (4H-3,1-benzothiazin-4-ones, 2,3-dihydro-1,4-benzodioxines and 3,4-dihydro-2H-1,4-benzoxazines) were identified as inhibitors of matriptase-2. The three heterocyclic scaffolds are similar as they consist of a benzene ring fused to a six-membered heterocyclic ring. The results enabled us to assess the effect of certain residues on biological activity. Even though these compounds are not expected to be selective, this set of data can be used for the future design of new compounds in which such residues were placed at different positions at the bicyclic core in a combinatorial way. For example, the 4-benzamidino-oxymethylene group might be introduced into the 4H-3,1-benzothiazin-4-one scaffold. The first attempts to decorate the 4H-3,1-benzothiazin-4-one heterocycle with a benzamidine moiety failed, because the scaffold was found to be unstable under the conditions used to convert a nitrile to an amidine group. Moreover, the substituents at positions 7 or 6 present in the active compounds (S)-12 and 17 might be introduced into the 4H-3,1-benzothiazin-4-one scaffold. The 6-substituent of 1 or the 2-substituent of 7 might also be considered for the design of new members of the 2,3-dihydro-1,4-benzodioxine and 3,4-dihydro-2H-1,4-benzoxazine series. Such investigations are planned for the future in our laboratories.
3. Experimental Section
3.1. Assays for Human Matriptase-2 Inhibition
The conditioned medium of HEK-MT2 cells was used as a source of matriptase-2 activity and assay conditions were as follows [11,19,25]. Assay buffer was 50 mM Tris–HCl, 150 mM NaCl, pH 8.0. The conditioned medium was collected and concentrated, and aliquots of the supernatant were stored at −20 °C. After thawing, it was diluted with assay buffer (1:10 or 1:20 depending on the enzyme activity) and kept at 0 °C not longer than 8 h. The assays were performed at a FLUOstar OPTIMA PlateReader (BMG Labtech, Ortenberg, Germany). A 10 mM stock solution of the fluorogenic substrate Boc-Gln-Ala-Arg-AMC (Bachem, Bubendorf, Switzerland) in DMSO was diluted with assay buffer. The final concentration of the substrate was 40 µM and of DMSO was 6%. The substrate concentration of 40 µM refers to 1.24 × Km [19]. Into each well containing 163.8 µL buffer, 11.2 µL of an inhibitor solution in DMSO and 10 µL of a substrate solution (800 µM) were added and thoroughly mixed. At 37 °C the reaction was initiated by adding 15 µL of diluted conditioned medium and followed over 400 s. All measurements were performed in duplicate with a single inhibitor concentration of 40 µM. Active inhibitors were investigated in duplicate with five different concentrations. Benzamidine hydrochloride was purchased from Acros Organics (Geel, Belgium).
3.2. Analysis of the Kinetic Data
Progress curves were analyzed by linear regression. IC50 values were determined by nonlinear regression using the equation vs = v0/(1 + [I]/IC50), where vs is the steady-state rate, v0 is the rate in the absence of the inhibitor, and [I] is the inhibitor concentration. Standard errors of the mean (SEM) values refer to this nonlinear regression.
3.3. Purity of Tested Compounds
After performing the kinetic measurements, the purity of the compounds was exemplarily checked by LC/MS. The purity was determined by HPLC-UV obtained on an LC-MS instrument (Applied Biosystems API 2000 LC/MS/MS (Darmstadt, Germany), HPLC Agilent 1100 (Waldbronn, Germany). UV absorption was detected from 220 to 400 nm using a diode array detector. In some cases, the DMSO stock solutions which were used for the inhibition assays were directly subjected to LC/MS. Elution was performed with a gradient of water/MeOH either containing 2 mM ammonium acetate from 90:10 up to 0:100 for 10 min at a flow rate of 300 μL/min. The compounds (S)-10, 13, 22, 23 and (R)-24 showed a purity of more than 90%. Compound 25 showed a purity of 81%. In case of compounds, from which stock solutions were prepared immediately before the kinetic measurements were performed, purity was checked as follows. Solutions in DMSO (1–6) or acetonitrile (7, 8) were prepared and subjected to LC/MS. Elution was performed with a gradient of water/MeOH either containing 2 mM ammonium acetate from 90:10 up to 0:100 for 10 min at a flow rate of 300 μL/min (compounds 1–6) or with a gradient of water/MeOH either containing 2 mM ammonium acetate from 60:40 up to 0:100 for 10 min at a flow rate of 300 μL/min (compounds 7 and 8) The compounds 1, 2, 3, 4, 5, 6 and 8, showed a purity of more than 90%. Compound 7 showed a purity of 86%.
Acknowledgments
A.M.B. and M.S. are supported by the German Research Foundation (DFG) Grant STI 660/1-1. M.S. is supported by the Maria von Linden Program of the Gender Equality Center of the University of Bonn. The authors thank Miloš Ilić, Petra Dunkel, Uroš Trstenjak and Anamarija Zega for donating compounds.
Author Contributions
M.G. and J.I. designed the study. P.G.R. and A.-M.B. performed kinetic experiments. P.G.P., A.-M.B. and M.G. analyzed the data. A.-M.B., M.S., J.C., D.K. and J.I. provided materials. M.S., J.C., D.K. and J.I. edited the manuscript. M.G., P.G.R. and A.-M.B. wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ginzburg, Y.; Rivella, S. β-Thalassemia: A model for elucidating the dynamic regulation of ineffective erythropoiesis and iron metabolism. Blood 2011, 118, 4321–4330. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, G.; Tzilianos, M.; Christakis, J.I.; Bogdanos, D.; Tsimirika, K.; MacFarlane, J.; Goldberg, Y.P.; Sakellaropoulos, N.; Ganz, T.; Nemeth, E. Hepcidin in iron overload disorders. Blood 2005, 105, 4103–4105. [Google Scholar] [CrossRef] [PubMed]
- Meynard, D.; Babitt, J.L.; Lin, H.Y. The liver: Conductor of systemic iron balance. Blood 2014, 123, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.S. Control of systemic iron homeostasis by the hemojuvelin-hepcidin axis. Adv. Nutr. 2010, 1, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Camaschella, C.; Silvestri, L. Molecular mechanisms regulating hepcidin revealed by hepcidin disorders. Sci. World J. 2011, 11, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.J.; Fleming, M.D. Modulation of hepcidin as therapy for primary and secondary iron overload disorders. Preclinical models and approaches. Hematol. Oncol. Clin. North Am. 2014, 28, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Nai, A.; Pagani, A.; Mandelli, G.; Lidonnici, M.R.; Silvestri, L.; Ferrari, G.; Camaschella, C. Deletion of TMPRSS6 attenuates the phenotype in a mouse model of β-thalassemia. Blood 2012, 119, 5021–5029. [Google Scholar] [CrossRef] [PubMed]
- Velasco, G.; Cal, S.; Quesada, V.; Sanchez, L.M.; Lopez-Otin, C. Matriptase-2, a membrane-bound mosaic serine proteinase predominantly expressed in human liver and showing degrading activity against extracellular matrix proteins. J. Biol. Chem. 2002, 277, 37637–37646. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, A.J.; Reid, J.C.; Velasco, G.; Quigley, J.P.; Hooper, J.D. The type II transmembrane serine protease matriptase-2—Identification, structural features, enzymology, expression pattern and potential roles. Front. Biosci. 2008, 13, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Stirnberg, M.; Gütschow, M. Matriptase-2, a regulatory protease of iron homeostasis: Possible substrates, cleavage sites and inhibitors. Curr. Pharm. Des. 2013, 19, 1052–1061. [Google Scholar] [PubMed]
- Stirnberg, M.; Maurer, E.; Horstmeyer, A.; Kolp, S.; Frank, S.; Bald, T.; Arenz, K.; Janzer, A.; Prager, K.; Walter, J.; et al. Proteolytic processing of the serine protease matriptase-2: Identification of the cleavage sites required for its autocatalytic release from the cell surface. Biochem. J. 2010, 430, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; She, E.; Gelbart, T.; Truksa, J.; Lee, P.; Xia, Y.; Khovananth, K.; Mudd, S.; Mann, N.; Moresco, E.M.; et al. The serine protease TMPRSS6 is required to sense iron deficiency. Science 2008, 320, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Finberg, K.E.; Heeney, M.M.; Campagna, D.R.; Aydinok, Y.; Pearson, H.A.; Hartman, K.R.; Mayo, M.M.; Samuel, S.M.; Strouse, J.J.; Markianos, K.; et al. Mutations in TMPRSS6 cause iron-refractory iron deficiency anemia (IRIDA). Nat. Genet. 2008, 40, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Folgueras, A.R.; de Lara, F.M.; Pendas, A.M.; Garabaya, C.; Rodriguez, F.; Astudillo, A.; Bernal, T.; Cabanillas, R.; Lopez-Otin, C.; Velasco, G. Membrane-bound serine protease matriptase-2 (TMPRSS6) is an essential regulator of iron homeostasis. Blood 2008, 112, 2539–2545. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, L.; Pagani, A.; Nai, A.; de Domenico, I.; Kaplan, J.; Camaschella, C. The serine protease matriptase-2 (TMPRSS6) inhibits hepcidin activation by cleaving membrane hemojuvelin. Cell Metable 2008, 8, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Maxson, J.E.; Chen, J.; Enns, C.A.; Zhang, A.S. Matriptase-2- and proprotein convertase-cleaved forms of hemojuvelin have different roles in the down-regulation of hepcidin expression. J. Biol. Chem. 2010, 285, 39021–39028. [Google Scholar] [CrossRef] [PubMed]
- Rausa, M.; Ghitti, M.; Pagani, A.; Nai, A.; Campanella, A.; Musco, G.; Camaschella, C.; Silvestri, L. Identification of TMPRSS6 cleavage sites of hemojuvelin. J. Cell. Mol. Med. 2015, 19, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Wysocka, M.; Gruba, N.; Miecznikowska, A.; Popow-Stellmaszyk, J.; Gütschow, M.; Stirnberg, M.; Furtmann, N.; Bajorath, J.; Lesner, A.; Rolka, K. Substrate specificity of human matriptase-2. Biochimie 2014, 97, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Dosa, S.; Stirnberg, M.; Lülsdorff, V.; Häuβler, D.; Maurer, E.; Gütschow, M. Active site mapping of trypsin, thrombin and matriptase-2 by sulfamoyl benzamidines. Bioorg. Med. Chem. 2012, 20, 6489–6505. [Google Scholar] [CrossRef] [PubMed]
- Furtmann, N.; Häußler, D.; Scheidt, T.; Stirnberg, M.; Steinmetzer, T.; Bajorath, J.; Gütschow, M. Limiting the number of potential binding modes by introducing symmetry into ligands: Structure-based design of inhibitors for trypsin-like serine proteases. Chem. Eur. J. 2016, 22, 610–625. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Casu, C.; Gardenghi, S.; Booten, S.; Aghajan, M.; Peralta, R.; Watt, A.; Freier, S.; Monia, B.P.; Rivella, S. Reducing TMPRSS6 ameliorates hemochromatosis and β-thalassemia in mice. J. Clin. Investig. 2013, 123, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.J.; Racie, T.; Westerman, M.; Fitzgerald, K.; Butler, J.S.; Fleming, M.D. Combination therapy with a Tmprss6 RNAi-therapeutic and the oral iron chelator deferiprone additively diminishes secondary iron overload in a mouse model of β-thalassemia intermedia. Am. J. Hematol. 2015, 90, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.J.; Toudjarska, I.; Sendamarai, A.K.; Racie, T.; Milstein, S.; Bettencourt, B.R.; Hettinger, J.; Bumcrot, D.; Fleming, M.D. An RNAi therapeutic targeting Tmprss6 decreases iron overload in Hfe(−/−) mice and ameliorates anemia and iron overload in murine β-thalassemia intermedia. Blood 2013, 121, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Casu, C.; Aghajan, M.; Oikonomidou, P.R.; Guo, S.; Monia, B.P.; Rivella, S. Combination of Tmprss6-ASO and the iron chelator deferiprone improves erythropoiesis and reduces iron overload in a mouse model of β-thalassemia intermedia. Haematologica 2016, 101, e8–e11. [Google Scholar] [CrossRef] [PubMed]
- Sisay, M.T.; Steinmetzer, T.; Stirnberg, M.; Maurer, E.; Hammami, M.; Bajorath, J.; Gütschow, M. Identification of the first low-molecular-weight inhibitors of matriptase-2. J. Med. Chem. 2010, 53, 5523–5535. [Google Scholar] [CrossRef] [PubMed]
- Hammami, M.; Rühmann, E.; Maurer, E.; Heine, A.; Gütschow, M.; Klebe, G.; Steinmetzer, T. New 3-amidinophenylalanine-derived inhibitors of matriptase. Med. Chem. Commun. 2012, 3, 807–813. [Google Scholar] [CrossRef]
- Duchêne, D.; Colombo, E.; Désilets, A.; Boudreault, P.L.; Leduc, R.; Marsault, E.; Najmanovich, R. Analysis of subpocket selectivity and identification of potent selective inhibitors for matriptase and matriptase-2. J. Med. Chem. 2014, 57, 10198–10204. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, A.; Dębowski, D.; Karna, N.; Łęgowska, A.; Stirnberg, M.; Gütschow, M.; Rolka, K. Inhibitors of matriptase-2 based on the trypsin inhibitor SFTI-1. ChemBioChem 2015, 16, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Bacchi, C.J.; Kode, N.R.; Zhang, Q.; Wang, G.; Yartlet, N.; Rattendi, D.; Londono, I.; Mazumder, L.; Vanden Eynde, J.J.; et al. Trypanocidal activity of piperazine-linked bisbenzamidines and bisbenzamidoxime, an orally active prodrug. Int. J. Antimicrob. Agents 2007, 30, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Vanden Eynde, J.J.; Mayence, A.; Collins, M.S.; Cushion, M.T.; Rattendi, D.; Londono, I.; Mazumder, L.; Bacchi, C.J.; Yarlett, N. Synthesis and SAR of alkanediamide-linked bisbenzamidines with anti-trypanosomal and anti-pneumocystis activity. Bioorg. Med. Chem. Lett. 2009, 19, 5884–5886. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Mayence, A.; Vanden Eynde, J.J. Some non-conventional biomolecular targets for diamidines. A short survey. Bioorg. Med. Chem. 2014, 22, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Gütschow, M.; Schlenk, M.; Gäb, J.; Paskaleva, M.; Alnouri, M.W.; Scolari, S.; Iqbal, J.; Müller, C.E. Benzothiazinones: A novel class of adenosine receptor antagonists structurally unrelated to xanthine and adenine derivatives. J. Med. Chem. 2012, 55, 3331–3341. [Google Scholar] [CrossRef] [PubMed]
- Stössel, A.; Schlenk, M.; Hinz, S.; Küppers, P.; Heer, J.; Gütschow, M.; Müller, C.E. Dual targeting of adenosine A(2A) receptors and monoamine oxidase B by 4H-3,1-benzothiazin-4-ones. J. Med. Chem. 2013, 56, 4580–4596. [Google Scholar] [CrossRef] [PubMed]
- Blättermann, S.; Peters, L.; Ottersbach, P.A.; Bock, A.; Konya, V.; Weaver, C.D.; Gonzalez, A.; Schröder, R.; Tyagi, R.; Luschnig, P.; et al. A biased ligand for OXE-R uncouples Gα and Gβγ signaling within a heterotrimer. Nat. Chem. Biol. 2012, 8, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Krantz, A.; Spencer, R.W.; Tam, T.F.; Liak, T.J.; Copp, L.J.; Thomas, E.M.; Rafferty, S.P. Design and synthesis of 4H-3,1-benzoxazin-4-ones as potent alternate substrate inhibitors of human leukocyte elastase. J. Med. Chem. 1990, 33, 464–479. [Google Scholar] [CrossRef] [PubMed]
- Neumann, U.; Schechter, N.M.; Gütschow, M. Inhibition of human chymase by 2-amino-3,1-benzoxazin-4-ones. Bioorg. Med. Chem. 2001, 9, 947–954. [Google Scholar] [CrossRef]
- Neumann, U.; Gütschow, M. 3,1-Benzothiazin-4-ones and 3,1-benzoxazin-4-ones: Highly different activities in chymotrypsin inactivation. Bioorg. Chem. 1995, 23, 72–88. [Google Scholar] [CrossRef]
- Ilić, M.; Dunkel, P.; Ilaš, J.; Chabielska, E.; Zakrzeska, A.; Mátyus, P.; Kikelj, D. Towards dual antithrombotic compounds—Balancing thrombin inhibitory and fibrinogen GPIIb/IIIa binding inhibitory activities of 2,3-dihydro-1,4-benzodioxine derivatives through regio- and stereoisomerism. Eur. J. Med. Chem. 2013, 62, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Ilić, M.; Kikelj, D.; Ilaš, J. Fluorinated dual antithrombotic compounds based on 1,4-benzoxazine scaffold. Eur. J. Med. Chem. 2012, 50, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Ilić, M.; Kontogiorgis, C.; Hadjipavlou-Litina, D.; Ilaš, J.; Kikelj, D. Thrombin inhibitors with lipid peroxidation and lipoxygenase inhibitory activities. Bioorg. Med. Chem. Lett. 2011, 21, 4705–4709. [Google Scholar] [CrossRef] [PubMed]
- Ilaš, J.; Jakopin, Z.; Borštnar, T.; Stegnar, M.; Kikelj, D. 3,4-Dihydro-2H-1,4-benzoxazine derivatives combining thrombin inhibitory and glycoprotein IIb/IIIa receptor antagonistic activity as a novel class of antithrombotic compounds with dual function. J. Med. Chem. 2008, 51, 5617–5629. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, M.; Cesar, C.; Štefanič, P.; Kikelj, D.; Janeš, D.; Murn, J.; Nadrah, K.; Tominc, M.; Addicks, E.; Giannis, A.; et al. Design and synthesis of novel platelet fibrinogen receptor antagonists with 2H-1,4-benzoxazine-3(4H)-one scaffold. A systematic study. Eur. J. Med. Chem. 2005, 40, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Trstenjak, U.; Ilaš, J.; Kikelj, D. Low molecular weight dual inhibitors of factor Xa and fibrinogen binding to GPIIb/IIIa with highly overlapped pharmacophores. Eur. J. Med. Chem. 2013, 64, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Zega, A.; Mlinsek, G.; Solmajer, T.; Trampus-Bakija, A.; Stegnar, M.; Urleb, U. Thrombin inhibitors built on an azaphenylalanine scaffold. Bioorg. Med. Chem. Lett. 2004, 14, 1563–1567. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).