Lambda Interferons: New Cytokines with Old Functions

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Regulation of Type III IFN Synthesis
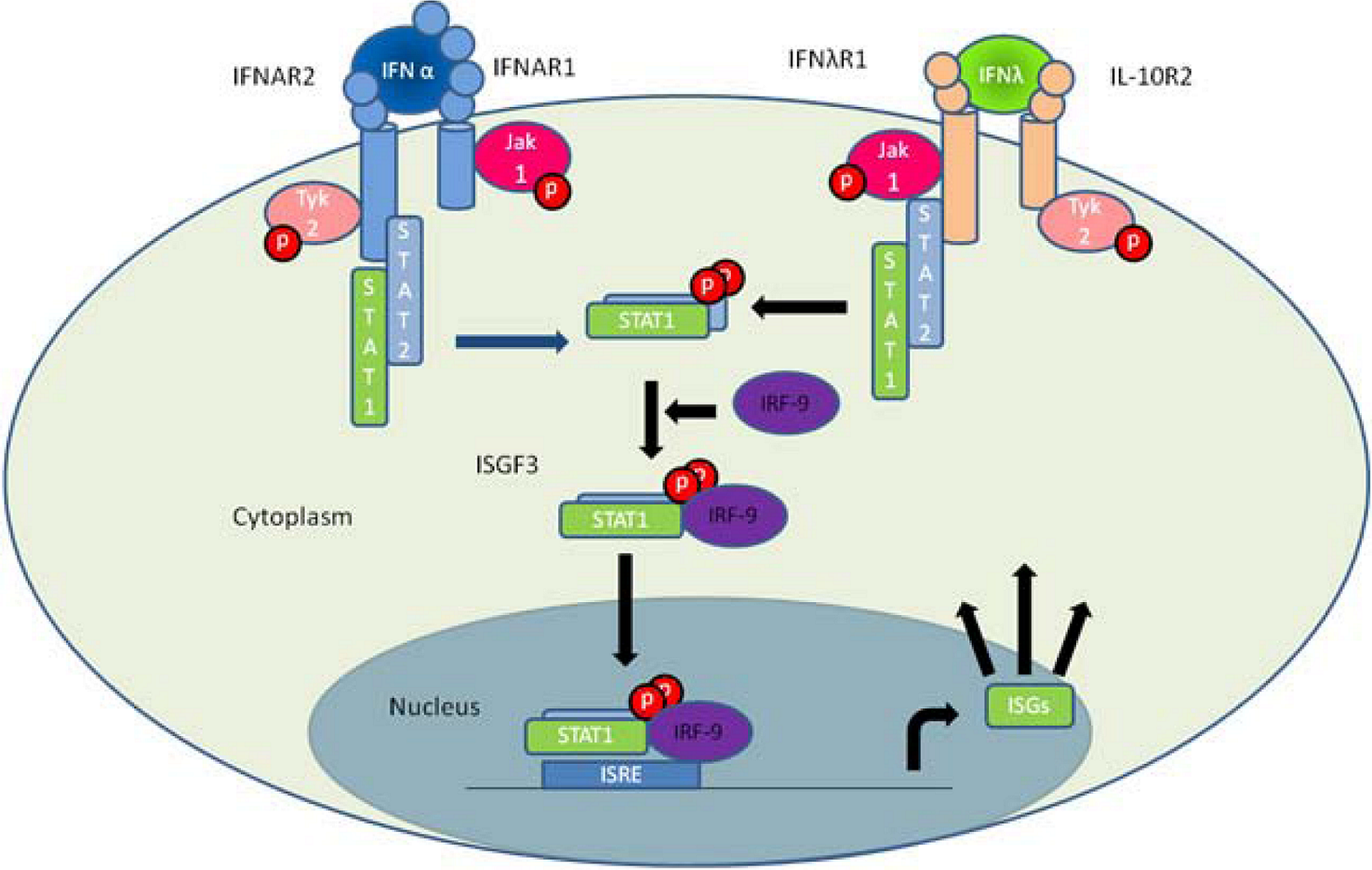
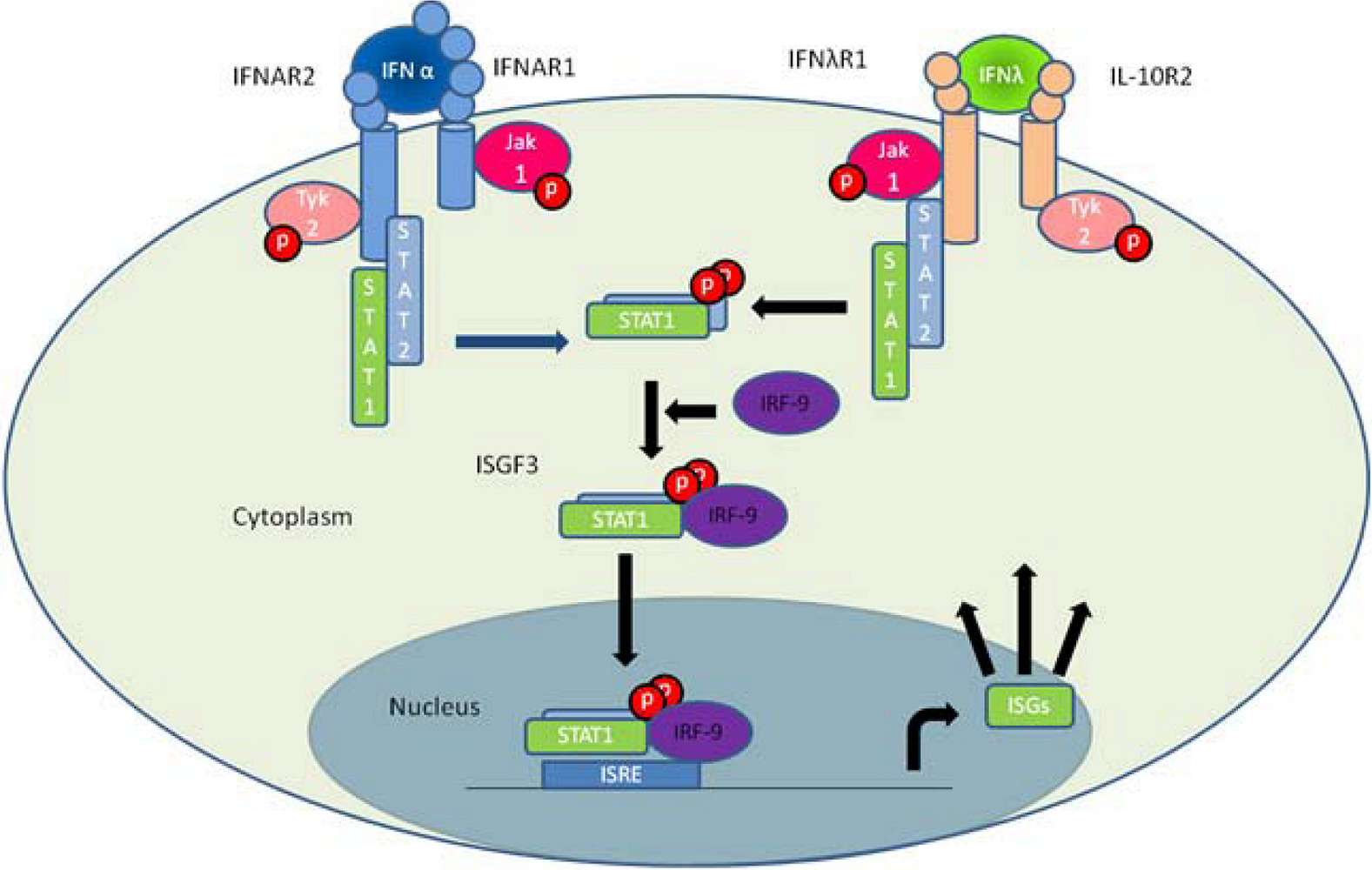
3. IFN-λ Receptor Function and Signaling
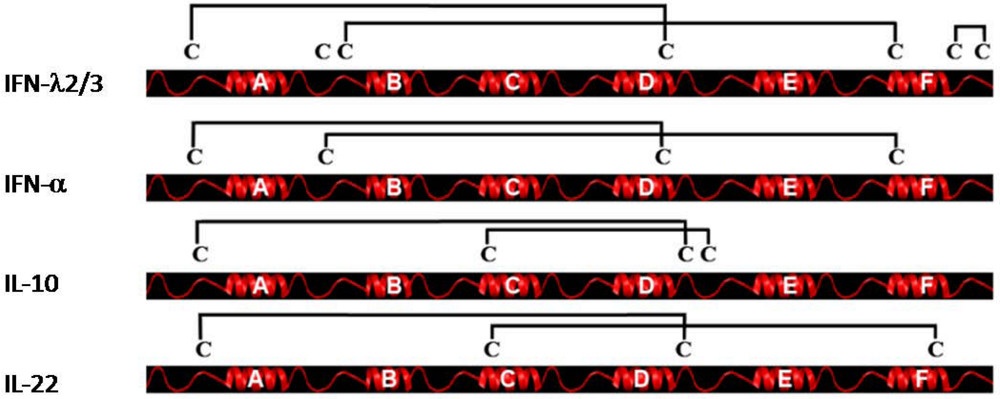
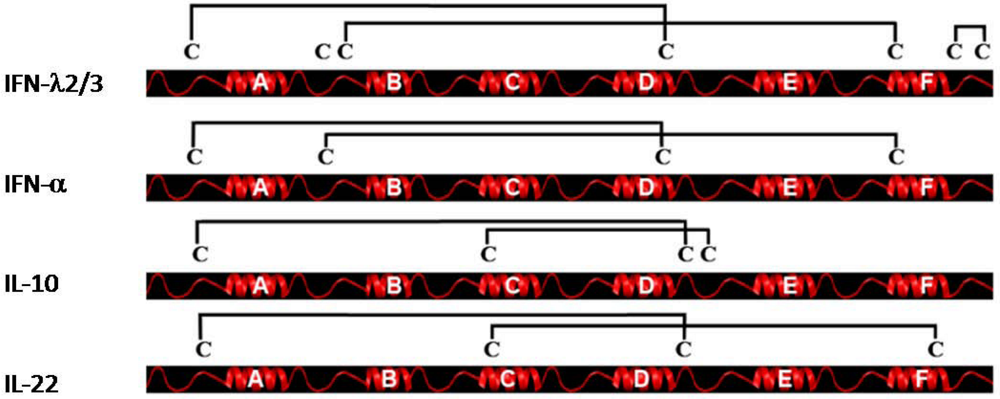
4. Genomic Organization
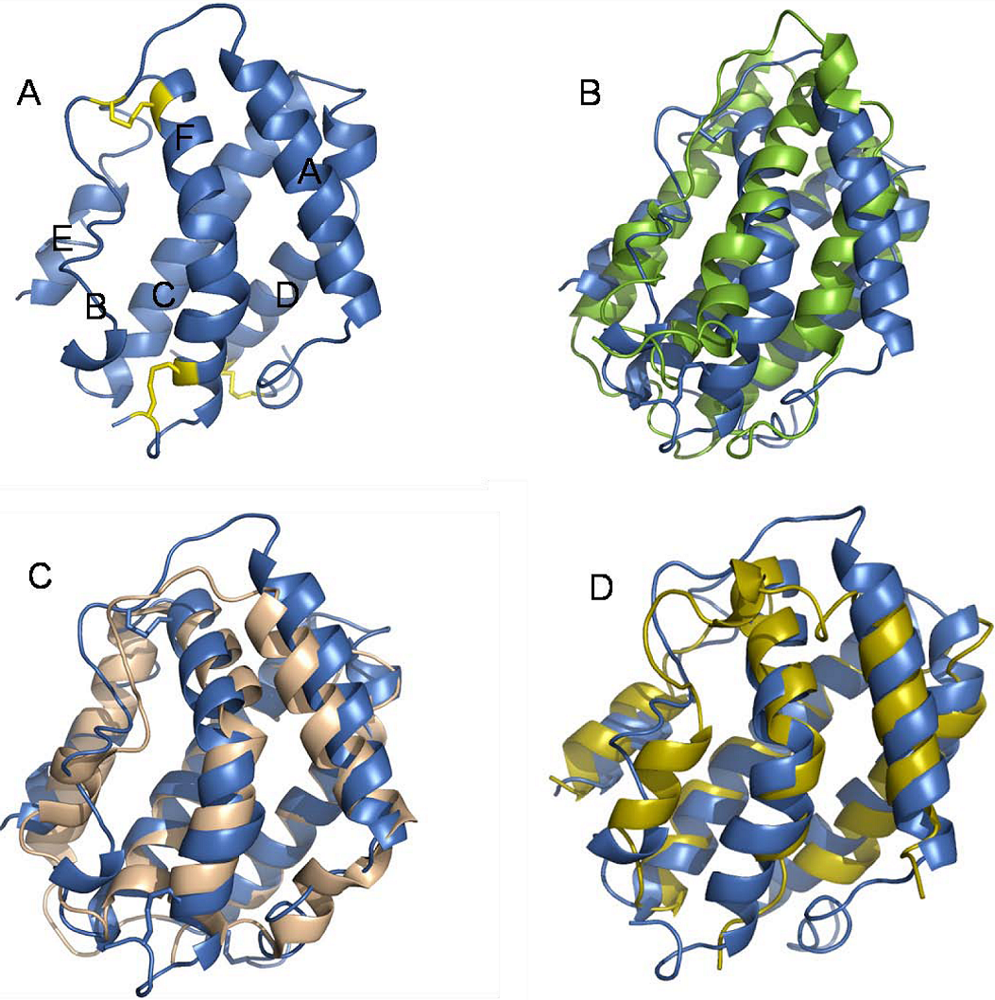
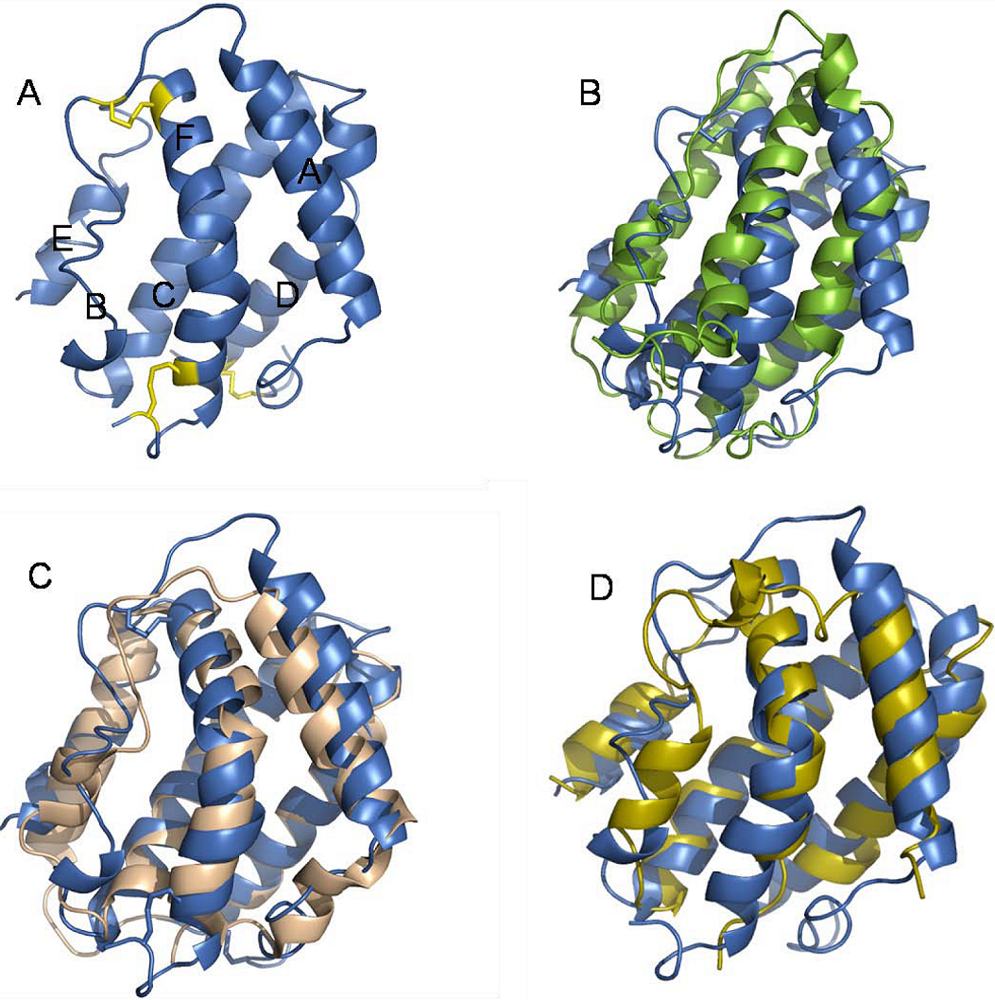
5. Structure of IFN-λ in Relation to Other Class II Cytokines
5. Potency of Different IFN-λ Subtypes
6. Role of IFN-λ in Vivo: Lessons from Animal Models and Studies on Human Diseases
7. IFN-λ in Adaptive Immunity
8. Anticancer Effect of Type III Interferon
9. Summery
Acknowledgements
References and Notes
- Sheppard, P.; Kindsvogel, W.; Xu, W.; Henderson, K.; Schlutsmeyer, S.; Whitmore, T.E.; Kuestner, R.; Garrigues, U.; Birks, C.; Roraback, J.; Ostrander, C.; Dong, D.; Shin, J.; Presnell, S.; Fox, B.; Haldeman, B.; Cooper, E.; Taft, D.; Gilbert, T.; Grant, F.J.; Tackett, M.; Krivan, W.; McKnight, G.; Clegg, C.; Foster, D.; Klucher, K.M. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nat. Immunol. 2003, 4, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.; Shah, N.K.; Langer, J.A.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Hamming, O.J.; Ank, N.; Paludan, S.R.; Nielsen, A.L.; Hartmann, R. Type III interferon (IFN) induces a type I IFN-like response in a restricted subset of cells through signaling pathways involving both the Jak-STAT pathway and the mitogen-activated protein kinases. J. Virol. 2007, 81, 7749–7758. [Google Scholar] [CrossRef] [PubMed]
- Marcello, T.; Grakoui, A.; Barba-Spaeth, G.; Machlin, E.S.; Kotenko, S.V.; MacDonald, M.R.; Rice, C.M. Interferons alpha and lambda inhibit hepatitis C virus replication with distinct signal transduction and gene regulation kinetics. Gastroenterology 2006, 131, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.E.; Schreckhise, H.; Khuu-Duong, K.; Henderson, K.; Rosler, R.; Storey, H.; Yao, L.; Liu, H.; Barahmand-pour, F.; Sivakumar, P.; Chan, C.; Birks, C.; Foster, D.; Clegg, C.H.; Wietzke-Braun, P.; Mihm, S.; Klucher, K.M. Interleukin-29 uses a type 1 interferon-like program to promote antiviral responses in human hepatocytes. Hepatology 2006, 44, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Meager, A.; Visvalingam, K.; Dilger, P.; Bryan, D.; Wadhwa, M. Biological activity of interleukins-28 and -29: comparison with type I interferons. Cytokine 2005, 31, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Sommereyns, C.; Paul, S.; Staeheli, P.; Michiels, T. IFN-lambda (IFN-lambda) is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo. PLoS Pathog. 2008, 4, e1000017. [Google Scholar] [CrossRef] [PubMed]
- Onoguchi, K.; Yoneyama, M.; Takemura, A.; Akira, S.; Taniguchi, T.; Namiki, H.; Fujita, T. Viral infections activate types I and III interferon genes through a common mechanism. J. Biol. Chem. 2007, 282, 7576–7581. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, P.I.; Pietila, T.E.; Veckman, V.; Kotenko, S.V.; Julkunen, I. IFN regulatory factor family members differentially regulate the expression of type III IFN (IFN-lambda) genes. J. Immunol. 2007, 179, 3434–3442. [Google Scholar] [PubMed]
- Cella, M.; Fuchs, A.; Vermi, W.; Facchetti, F.; Otero, K.; Lennerz, J.K.; Doherty, J.M.; Mills, J.C.; Colonna, M. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 2009, 457, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, R.P.; Sheikh, F.; Kotenko, S.V.; Dickensheets, H. The expanded family of class II cytokines that share the IL-10 receptor-2 (IL-10R2) chain. J. Leukoc. Biol. 2004, 76, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Harlos, K.; Martin, D.M.; O'Brien, D.P.; Jones, E.Y.; Stuart, D.I.; Polikarpov, I.; Miller, A.; Tuddenham, E.G.; Boys, C.W. Crystal structure of the extracellular region of human tissue factor. Nature 1994, 370, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.R.; Windsor, W.T.; Nagabhushan, T.L.; Lundell, D.J.; Lunn, C.A.; Zauodny, P.J.; Narula, S.K. Crystal structure of a complex between interferon-gamma and its soluble high-affinity receptor. Nature 1995, 376, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Gad, H.H.; Dellgren, C.; Hamming, O.J.; Vends, S.; Paludan, S.R.; Hartmann, R. Interferon-lambda is functionally an interferon but structurally related to the interleukin-10 family. J. Biol. Chem. 2009, 284, 20869–20875. [Google Scholar] [CrossRef] [PubMed]
- Dumoutier, L.; Lejeune, D.; Hor, S.; Fickenscher, H.; Renauld, J.C. Cloning of a new type II cytokine receptor activating signal transducer and activator of transcription (STAT)1, STAT2 and STAT3. Biochem. J. 2003, 370, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Izotova, L.S.; Pollack, B.P.; Muthukumaran, G.; Paukku, K.; Silvennoinen, O.; Ihle, J.N.; Pestka, S. Other kinases can substitute for Jak2 in signal transduction by interferon-gamma. J. Biol. Chem. 1996, 271, 17174–17182. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Witthuhn, B.A.; Matsuda, T.; Kohlhuber, F.; Kerr, I.M.; Ihle, J.N. Activation of Jak2 catalytic activity requires phosphorylation of Y1007 in the kinase activation loop. Mol. Cell Biol. 1997, 17, 2497–2501. [Google Scholar] [PubMed]
- Dumoutier, L.; Tounsi, A.; Michiels, T.; Sommereyns, C.; Kotenko, S.V.; Renauld, J.C. Role of the interleukin (IL)-28 receptor tyrosine residues for antiviral and antiproliferative activity of IL-29/interferon-lambda 1: similarities with type I interferon signaling. J. Biol. Chem. 2004, 279, 32269–32274. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Vinkemeier, U.; Zhao, Y.; Jeruzalmi, D.; Darnell, J.E., Jr.; Kuriyan, J. Crystal structure of a tyrosine phosphorylated STAT-1 dimer bound to DNA. Cell 1998, 93, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Mordstein, M.; Kochs, G.; Dumoutier, L.; Renauld, J.C.; Paludan, S.R.; Klucher, K.; Staeheli, P. Interferon-lambda contributes to innate immunity of mice against influenza A virus but not against hepatotropic viruses. PLoS Pathog 2008, 4, e1000151. [Google Scholar] [CrossRef] [PubMed]
- Mennechet, F.J.; Uze, G. Interferon-lambda-treated dendritic cells specifically induce proliferation of FOXP3-expressing suppressor T cells. Blood 2006, 107, 4417–4423. [Google Scholar] [CrossRef] [PubMed]
- Witte, K.; Gruetz, G.; Volk, H.D.; Looman, A.C.; Asadullah, K.; Sterry, W.; Sabat, R.; Wolk, K. Despite IFN-lambda receptor expression, blood immune cells, but not keratinocytes or melanocytes, have an impaired response to type III interferons: implications for therapeutic applications of these cytokines. Genes Immun. 2009, 10, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Lasfar, A.; Lewis-Antes, A.; Smirnov, S.V.; Anantha, S.; Abushahba, W.; Tian, B.; Reuhl, K.; Dickensheets, H.; Sheikh, F.; Donnelly, R.P.; Raveche, E.; Kotenko, S.V. Characterization of the mouse IFN-lambda ligand-receptor system: IFN-lambdas exhibit antitumor activity against B16 melanoma. Cancer Res. 2006, 66, 4468–4477. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef]
- Walter, M.R. Structural analysis of IL-10 and Type I interferon family members and their complexes with receptor. Adv. Protein Chem. 2004, 68, 171–223. [Google Scholar] [PubMed]
- Senda, T.; Shimazu, T.; Matsuda, S.; Kawano, G.; Shimizu, H.; Nakamura, K.T.; Mitsui, Y. Three-dimensional crystal structure of recombinant murine interferon-beta. EMBO J. 1992, 11, 3193–3201. [Google Scholar] [PubMed]
- Radhakrishnan, R.; Walter, L.J.; Hruza, A.; Reichert, P.; Trotta, P.P.; Nagabhushan, T.L.; Walter, M.R. Zinc mediated dimer of human interferon-alpha 2b revealed by X-ray crystallography. Structure 1996, 4, 1453–1463. [Google Scholar] [CrossRef]
- Zdanov, A.; Schalk-Hihi, C.; Gustchina, A.; Tsang, M.; Weatherbee, J.; Wlodawer, A. Crystal structure of interleukin-10 reveals the functional dimer with an unexpected topological similarity to interferon gamma. Structure 1995, 3, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Magracheva, E.; Kozlov, S.; Fong, S.; Tobin, G.; Kotenko, S.; Wlodawer, A.; Zdanov, A. Crystal structure of interleukin-19 defines a new subfamily of helical cytokines. J. Biol. Chem. 2003, 278, 3308–3313. [Google Scholar] [CrossRef] [PubMed]
- Nagem, R.A.; Colau, D.; Dumoutier, L.; Renauld, J.C.; Ogata, C.; Polikarpov, I. Crystal structure of recombinant human interleukin-22. Structure 2002, 10, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.A.; Sheppard, P.O.; O'Hara, P.J. The role of genomic data in the discovery, annotation and evolutionary interpretation of the interferon-lambda family. PLoS One 2009, 4, e4933. [Google Scholar] [CrossRef] [PubMed]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda interferon (IFN-lambda), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. J. Virol. 2006, 80, 4501–4509. [Google Scholar] [CrossRef] [PubMed]
- Dellgren, C.; Gad, H.H.; Hamming, O.J.; Melchjorsen, J.; Hartmann, R. Human interferon-lambda3 is a potent member of the type III interferon family. Genes Immun. 2009, 10, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, N.W.; Buttigieg, K.; Kotenko, S.V.; Smith, G.L. Murine interferon lambdas (type III interferons) exhibit potent antiviral activity in vivo in a poxvirus infection model. J. Gen. Virol. 2005, 86, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Ank, N.; Iversen, M.B.; Bartholdy, C.; Staeheli, P.; Hartmann, R.; Jensen, U.B.; Dagnaes-Hansen, F.; Thomsen, A.R.; Chen, Z.; Haugen, H.; Klucher, K.; Paludan, S.R. An important role for type III interferon (IFN-lambda/IL-28) in TLR-induced antiviral activity. J. Immunol. 2008, 180, 2474–2485. [Google Scholar] [PubMed]
- Contoli, M.; Message, S.D.; Laza-Stanca, V.; Edwards, M.R.; Wark, P.A.; Bartlett, N.W.; Kebadze, T.; Mallia, P.; Stanciu, L.A.; Parker, H.L.; Slater, L.; Lewis-Antes, A.; Kon, O.M.; Holgate, S.T.; Davies, D.E.; Kotenko, S.V.; Papi, A.; Johnston, S.L. Role of deficient type III interferon-lambda production in asthma exacerbations. Nat. Med. 2006, 12, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, C.A.; Chang, M.; Tang, W.; Campbell, J.; Sullivan, D.G.; Gretch, D.R.; Corey, L.; Coombs, R.W.; Fausto, N. Hepatitis C virus replication in transfected and serum-infected cultured human fetal hepatocytes. Am. J. Pathol. 2007, 170, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Fellay, J.; Thompson, A.J.; Simon, J.S.; Shianna, K.V.; Urban, T.J.; Heinzen, E.L.; Qiu, P.; Bertelsen, A.H.; Muir, A.J.; Sulkowski, M.; McHutchison, J.G.; Goldstein, D.B. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 2009, 461, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.L.; Thio, C.L.; Martin, M.P.; Qi, Y.; Ge, D.; O'Huigin, C.; Kidd, J.; Kidd, K.; Khakoo, S.I.; Alexander, G.; Goedert, J.J.; Kirk, G.D.; Donfield, S.M.; Rosen, H.R.; Tobler, L.H.; Busch, M.P.; McHutchison, J.G.; Goldstein, D.B.; Carrington, M. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature 2009, 461, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Smirnov, S.V.; Lewis-Antes, A.; Balan, M.; Li, W.; Tang, S.; Silke, G.V.; Putz, M.M.; Smith, G.L.; Kotenko, S.V. Inhibition of type I and type III interferons by a secreted glycoprotein from Yaba-like disease virus. Proc. Natl. Acad. Sci. USA 2007, 104, 9822–9827. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Morrow, M.P.; Pankhong, P.; Laddy, D.J.; Schoenly, K.A.; Yan, J.; Cisper, N.; Weiner, D.B. Comparative ability of IL-12 and IL-28B to regulate Treg populations and enhance adaptive cellular immunity. Blood 2009, 113, 5868–5877. [Google Scholar] [CrossRef] [PubMed]
- Jordan, W.J.; Eskdale, J.; Srinivas, S.; Pekarek, V.; Kelner, D.; Rodia, M.; Gallagher, G. Human interferon lambda-1 (IFN-lambda1/IL-29) modulates the Th1/Th2 response. Genes Immun. 2007, 8, 254–261. [Google Scholar] [CrossRef]
- Dai, J.; Megjugorac, N.J.; Gallagher, G.E.; Yu, R.Y.; Gallagher, G. IFN-lambda1 (IL-29) inhibits GATA3 expression and suppresses Th2 responses in human naive and memory T cells. Blood 2009, 113, 5829–5838. [Google Scholar] [CrossRef] [PubMed]
- Golding, B.; Zaitseva, M.; Golding, H. The potential for recruiting immune responses toward type 1 or type 2 T cell help. Am. J. Trop. Med. Hyg. 1994, 50, 33–40. [Google Scholar] [PubMed]
- Khaitov, M.R.; Laza-Stanca, V.; Edwards, M.R.; Walton, R.P.; Rohde, G.; Contoli, M.; Papi, A.; Stanciu, L.A.; Kotenko, S.V.; Johnston, S.L. Respiratory virus induction of alpha-, beta- and lambda-interferons in bronchial epithelial cells and peripheral blood mononuclear cells. Allergy 2009, 64, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Prather, A.A.; Rabinovitz, M.; Pollock, B.G.; Lotrich, F.E. Cytokine-induced depression during IFN-alpha treatment: the role of IL-6 and sleep quality. Brain Behav. Immun. 2009, 23, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.M.; Klucher, K.M.; Freeman, J.A.; Hausman, D.F.; Fontana, D.; Williams, D.E. Interferon lambda as a potential new therapeutic for hepatitis C. Ann. N. Y. Acad. Sci. 2009, 1182, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Talpaz, M.; Kantarjian, H.M.; McCredie, K.B.; Keating, M.J.; Trujillo, J.; Gutterman, J. Clinical investigation of human alpha interferon in chronic myelogenous leukemia. Blood 1987, 69, 1280–1288. [Google Scholar] [PubMed]
- Maher, S.G.; Sheikh, F.; Scarzello, A.J.; Romero-Weaver, A.L.; Baker, D.P.; Donnelly, R.P.; Gamero, A.M. IFNalpha and IFNlambda differ in their antiproliferative effects and duration of JAK/STAT signaling activity. Cancer Biol. Ther. 2008, 7, 1109–1115. [Google Scholar] [PubMed]
- Li, W.; Lewis-Antes, A.; Huang, J.; Balan, M.; Kotenko, S.V. Regulation of apoptosis by type III interferons. Cell Prolif. 2008, 41, 960–979. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Ohtsuki, M.; Hata, M.; Kobayashi, E.; Murakami, T. Antitumor activity of IFN-lambda in murine tumor models. J. Immunol. 2006, 176, 7686–7694. [Google Scholar] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hamming, O.J.; Gad, H.H.; Paludan, S.; Hartmann, R. Lambda Interferons: New Cytokines with Old Functions. Pharmaceuticals 2010, 3, 795-809. https://doi.org/10.3390/ph3040795
Hamming OJ, Gad HH, Paludan S, Hartmann R. Lambda Interferons: New Cytokines with Old Functions. Pharmaceuticals. 2010; 3(4):795-809. https://doi.org/10.3390/ph3040795
Chicago/Turabian StyleHamming, Ole J., Hans Henrik Gad, Søren Paludan, and Rune Hartmann. 2010. "Lambda Interferons: New Cytokines with Old Functions" Pharmaceuticals 3, no. 4: 795-809. https://doi.org/10.3390/ph3040795
APA StyleHamming, O. J., Gad, H. H., Paludan, S., & Hartmann, R. (2010). Lambda Interferons: New Cytokines with Old Functions. Pharmaceuticals, 3(4), 795-809. https://doi.org/10.3390/ph3040795