

Neuroprotective Indole Diterpenoids from the Fungus Tolypocladium album DWS131

 , , and
, , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Structural Identification
2.2. Neuroprotective Effects of Compounds 1–9 Against Glu-Induced Excitotoxicity in R28 Cells
2.3. Compounds 8 and 9 Inhibited Ferroptosis Signaling in Glu-Induced Excitotoxicity Model of R28 Cells
2.4. Compounds 8 and 9 Protected Against N-Methyl-D-Aspartic Acid (NMDA)-Induced RGC Injury and Visual Dysfunction in Mice
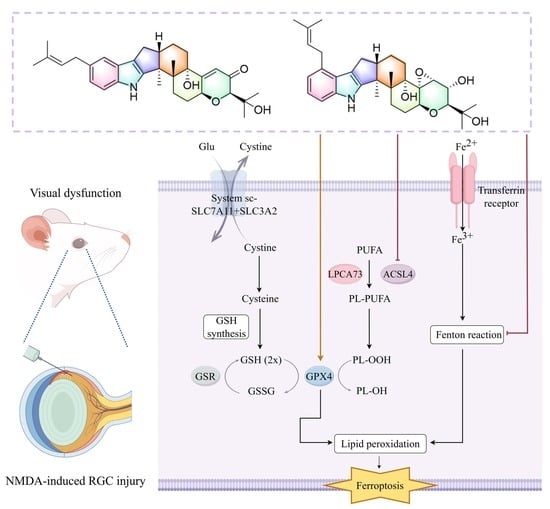
2.5. Modulation of the SLC7A11-GPX4/ACSL4 Ferroptosis Pathway by Compounds 8 and 9 Protected RGCs In Vitro and In Vivo
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Fungal Material
4.3. Fermentation and Extraction
4.4. Isolation
4.5. Quantum Chemical Calculation Methods
4.6. Cell Culture
4.7. Cell Viability Assay and Hoechst 33342/PI Staining
4.8. ROS Detection
4.9. Detection of GSH and MDA
4.10. Detection of Intracellular Fe2+ and Lipid Peroxidation
4.11. Establishment of the NMDA Mouse Model
4.12. Retinal Flat-Mount
4.13. Immunofluorescence of Retinal Cryosections
4.14. f-VEP and f-ERG
4.15. Automated Visual Acuity Testing (OptoDrum)
4.16. H&E Staining
4.17. Western Blotting
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yee, D.A.; Niwa, K.; Perlatti, B.; Chen, M.; Li, Y.; Tang, Y. Genome mining for unknown–unknown natural products. Nat. Chem. Biol. 2023, 19, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ji, M.; Dong, J.; Shi, D.; Wang, Y.; Liu, L.; Feng, S.; Liu, L. New bioactive secondary metabolites from fungi: 2023. Mycology 2024, 15, 283–321. [Google Scholar] [CrossRef]
- Yu, F.-M.; Thilini Chethana, K.W.; Wei, D.-P.; Liu, J.-W.; Zhao, Q.; Tang, S.-M.; Li, L.; Hyde, K.D. Comprehensive review of Tolypocladium and description of a novel lineage from southwest China. Pathogens 2021, 10, 1389. [Google Scholar] [CrossRef]
- Lundgren, B.; Bååth, E.; Söderström, B.E. Antagonistic effects of Tolypocladium species. Trans. Br. Mycol. Soc. 1978, 70, 305–307. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Yuan, L.J.; Chen, Z.J.; Fu, L.; Lu, J.H.; Meng, Q.F.; He, H.; Yu, X.X.; Lin, F.; Teng, L.R. Purification and characterization of β-glucosidase from a newly isolated strain Tolypocladium cylindrosporum Syzx4. Chem. Res. Chin. Univ. 2011, 27, 557–561. [Google Scholar]
- Krasnoff, S.B.; Gupta, S.; Leger, R.J.S.; Renwick, J.A.A.; Roberts, D.W. Antifungal and insecticidal properties of the efrapeptins: Metabolites of the fungus Tolypocladium niveum. J. Invertebr. Pathol. 1991, 58, 180–188. [Google Scholar] [CrossRef]
- Gazis, R.; Skaltsas, D.; Chaverri, P. Novel endophytic lineages of Tolypocladium provide new insights into the ecology and evolution of Cordyceps-like fungi. Mycologia 2014, 106, 1090–1105. [Google Scholar] [CrossRef]
- Senadeera, S.P.D.; Wang, D.; Kim, C.-K.; Smith, E.A.; Durrant, D.E.; Alexander, P.A.; Wendt, K.L.; Stephen, A.G.; Morrison, D.K.; Cichewicz, R.H.; et al. Tolypocladamides A–G: Cytotoxic peptaibols from Tolypocladium inflatum. J. Nat. Prod. 2022, 85, 1603–1616. [Google Scholar] [CrossRef]
- Dreyfuss, M.; Härri, E.; Hofmann, H.; Kobel, H.; Pache, W.; Tscherter, H. Cyclosporin A and C. Appl. Microbiol. Biotechnol. 1976, 3, 125–133. [Google Scholar]
- Wu, X.Q.; Li, J.; Zhou, X.; Wang, J.; Tan, Y.F.; Mo, J.S.; Liu, S.; Xu, K.P.; Tan, G.S.; Zhang, W.; et al. Liver-cell protective pyridones from the fungi Tolypocladium album dws120. Phytochemistry 2023, 212, 113730. [Google Scholar] [CrossRef]
- Ruan, Q.F.; He, Y.; Jiang, S.Q.; Cui, H.; Jin, J.; Zhao, Z.X. Cytotoxic indole diterpenoids and rare pyridoxatin atropisomers from the endophytic fungus Tolypocladium sp. SHJJ1. Fitoterapia 2024, 175, 105983. [Google Scholar] [CrossRef]
- Byrne, K.M.; Smith, S.K.; Ondeyka, J.G. Biosynthesis of nodulisporic Acid A: precursor studies. J. Am. Chem. Soc. 2002, 124, 7055–7060. [Google Scholar] [CrossRef]
- Saikia, S.; Nicholson, M.J.; Young, C.; Parker, E.J.; Scott, B. The genetic basis for indole-diterpene chemical diversity in filamentous fungi. Mycol. Res. 2008, 112, 184–199. [Google Scholar] [CrossRef]
- Gao, S.-S.; Li, X.-M.; Williams, K.; Proksch, P.; Ji, N.-Y.; Wang, B.-G. Rhizovarins A–F, indole-diterpenes from the mangrove-derived endophytic fungus Mucor irregularis QEN-189. J. Nat. Prod. 2016, 79, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, R.; Lin, M.; Chen, C.; Qi, X.; Zhou, X.; Liu, Y.; Tan, Y.; Luo, X. Antiosteoclastogenic indole alkaloids from the mangrove endophytic fungus Penicillium brefeldianum GXIMD 02511. J. Nat. Prod. 2025, 88, 1671–1683. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, X.-Y.; Zhu, J.-Y.; Cui, Q.-Y.; Lin, Y.-W.; Lu, Q.-R.; Wu, S.-Q.; Fu, Y.-F.; Zhou, Q.-L.; Yuan, T.; et al. HSQC-guided discovery and structural optimization of antiadipogenic indole diterpenoids from endophytic fungus Penicillium janthinellum H-6. Eur. J. Med. Chem. 2025, 297, 117956. [Google Scholar] [CrossRef]
- Wang, Q.-S.; Gao, W.-B.; Gu, H.-Y.; Liu, Z.-Q.; Cao, F. Dipenipenoids C and D, new dimeric indole-diterpenoids from the marine-derived fungus Penicillium sp. CF-06. Tetrahedron 2025, 185, 134850. [Google Scholar] [CrossRef]
- Bo, H.; Li, L.-Y.; Zhang, L.-J.; Liu, X.-N.; Pang, S. Indole-diterpenoids with anti-MRSA activity from the endophytic fungus Aspergillus fumigatus. Chem. Nat. Compd. 2025, 61, 827–830. [Google Scholar] [CrossRef]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and structure elucidation of Janthitrems A and D from Penicillium janthinellum and determination of the tremorgenic and anti-insect activity of Janthitrems A and B. J. Agric. Food. Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Guthridge, K.; Vassiliadis, S.; Hemsworth, J.; Hettiarachchige, I.; Spangenberg, G.; Rochfort, S. Tremorgenic mycotoxins: Structure diversity and biological activity. Toxins 2019, 11, 302. [Google Scholar] [CrossRef]
- Wang, X.-X.; Chen, Z.-L.; Zhang, J.-S.; Liu, H.-S.; Ma, R.-P.; Liu, X.-P.; Li, M.-Y.; Ge, D.; Bao, J.; Zhang, H. Indole diterpenes from mangrove sediment-derived fungus Penicillium sp. UJNMF0740 protect PC12 cells against 6-OHDA-induced neurotoxicity via regulating the PI3K/Akt pathway. Mar. Drugs 2023, 21, 593. [Google Scholar] [CrossRef]
- Chen, Z.W.; Wang, Y.; Chao, R.; Wu, T.Z.; Zhong, T.H.; Yang, X.W.; Wang, B. Chemical Constituents with Ferroptosis Inhibitory Activity of the Deep-Sea-Derived Penicillium paxilli. Chem. Biodivers. 2025, 22, e202500540. [Google Scholar] [CrossRef]
- Mo, J.; Tan, Y.; Ai, W.; Li, Y.; Yuan, Y.; Jiang, Y.; Xu, K.; Tan, G.; Wang, W.; Li, J.; et al. Macrocyclic trichothecenes from Myrothecium verrucaria PA 57 and their cytotoxic activity. Chin. J. Nat. Med. 2024, 22, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Chen, X.; Wang, C.; Liang, A.; Lu, W.; Xia, X.; Li, J.; Liu, S.; Wang, W.-X. A novel fungal alkaloid protects retinal ganglion cells against glutamate-induced excitotoxicity in vitro and in vivo via activation of GABA pathway. Bioorgan. Chem. 2025, 166, 109170. [Google Scholar] [CrossRef]
- Nogawa, T.; Kawatani, M.; Okano, A.; Futamura, Y.; Aono, H.; Shimizu, T.; Kato, N.; Kikuchi, H.; Osada, H. Structure and biological activity of Metarhizin C, a stereoisomer of BR-050 from Tolypocladium album RK17-F0007. J. Antibiot. 2019, 72, 996–1000. [Google Scholar] [CrossRef]
- Fukuda, T.; Sudoh, Y.; Tsuchiya, Y.; Okuda, T.; Matsuura, N.; Motojima, A.; Oikawa, T.; Igarashi, Y. Tolypoalbin, a new tetramic acid from Tolypocladium album TAMA 479. J. Antibiot. 2015, 68, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.-K.; Wang, W.-X. GIAO 13C NMR calculation with sorted training sets improves accuracy and reliability for structural assignation. J. Org. Chem. 2020, 85, 11350–11358. [Google Scholar] [CrossRef]
- Ai, W.-J.; Li, J.; Cao, D.; Liu, S.; Yuan, Y.-Y.; Li, Y.; Tan, G.-S.; Xu, K.-P.; Yu, X.; Kang, F.; et al. A very deep graph convolutional network for 13C NMR chemical shift calculations with density functional theory level performance for structure assignment. J. Nat. Prod. 2024, 87, 743–752. [Google Scholar] [CrossRef]
- Sharpe, R.J.; Johnson, J.S. Asymmetric total synthesis of the indole diterpene alkaloid paspaline. J. Org. Chem. 2015, 80, 9740–9766. [Google Scholar] [CrossRef]
- Springer, J.P.; Clardy, J.; Wells, J.M.; Cole, R.J.; Kirksey, J.W. The structure of paxilline, a tremorgenic metabolite of Penicillium paxilli bainier. Tetrahedron Lett. 1975, 16, 2531–2534. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Antiinsectan alkaloids: Shearinines A-C and a new paxilline derivative from the ascostromata of Eupenicillium shearii. Tetrahedron 1995, 51, 3959–3968. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Gallagher, R.T.; Hawkes, A.D.; Munday, S.C.; Towers, N.R. Synthesis and tremorgenicity of paxitriols and lolitriol: Possible biosynthetic precursors of lolitrem B. J. Agric. Food Chem. 1992, 40, 234–238. [Google Scholar] [CrossRef]
- Tan, Y.F.; Chen, X.Y.; Wang, C.; Liang, A.L.; Lu, W.Y.; Feng, L.M.; Huang, Q.; Jiang, Y.P.; Liu, S.; Xia, X.B.; et al. Tolypaxillines A–J: Indole diterpenoids with neuroprotective activities from a soil-derived fungus Tolypocladium album DWS131. Phytochemistry 2026, 247, 114855. [Google Scholar] [CrossRef]
- Itabashi, T.; Hosoe, T.; Wakana, D.; Fukushima, K.; Takizawa, K.; Yaguchi, T.; Okada, K.; de Campos Takaki, G.M.; Kawai, K.-I. A new indoloditerpene derivative, penijanthine A, isolated from Penicillium janthinellum. J. Nat. Med. 2009, 63, 96–99. [Google Scholar] [CrossRef]
- Roll, D.M.; Barbieri, L.R.; Bigelis, R.; McDonald, L.A.; Arias, D.A.; Chang, L.-P.; Singh, M.P.; Luckman, S.W.; Berrodin, T.J.; Yudt, M.R. The lecanindoles, nonsteroidal progestins from the terrestrial fungus Verticillium lecanii 6144. J. Nat. Prod. 2009, 72, 1944–1948. [Google Scholar] [CrossRef]
- Tani, K.; Tsuruta, T.; Yoshino, Y.; Fukushige, A.; Doe, M.; Miyake, H.; Morimoto, Y.; Hashimoto, M.; Ukiya, M.; Marumoto, S.; et al. Skeletal Diversity of Diterpenoids Isolated from the Soft Coral Sclerophytum heterospiculatum in Kikai Island. J. Nat. Prod. 2026, 89, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wang, Y.; Liu, P.; Fu, P.; Zhu, T.; Wang, W.; Zhu, W. Indole-diterpenoids with anti-H1N1 activity from the aciduric fungus Penicillium camemberti OUCMDZ-1492. J. Nat. Prod. 2013, 76, 1328–1336. [Google Scholar] [CrossRef]
- Su, S.-S.; Song, A.-H.; Chen, G.; Wang, H.-F.; Li, Z.-Q.; Pei, Y.-H. Two new indole-diterpenoids from the fungus Penicillium crustosum YN-HT-15. J. Asian Nat. Prod. Res. 2014, 16, 285–289. [Google Scholar] [CrossRef]
- Zhao, J.-C.; Wang, Y.-L.; Zhang, T.-Y.; Chen, Z.-J.; Yang, T.-M.; Wu, Y.-Y.; Sun, C.-P.; Ma, X.-C.; Zhang, Y.-X. Indole diterpenoids from the endophytic fungus Drechmeria sp. as natural antimicrobial agents. Phytochemistry 2018, 148, 21–28. [Google Scholar] [CrossRef]
- Yuan, Y.-Y.; Li, Y.; Lu, W.-Y.; Liang, A.-L.; Li, J.; Wang, W.-X. Xylariaides A and B, novel cytochalasans with a unique 5/6/5/3 ring system from a soil fungus Xylaria sp. Y01. Nat. Prod. Bioprosp. 2025, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.H.; Nishida, H.; Tomoda, H.; Tabata, N.; Shiomi, K.; Yang, D.J.; Takayanagi, H.; Omura, S. Terpendoles, novel ACAT inhibitors produced by Albophoma yamanashiensis. II. structure elucidation of terpendoles A, B, C and D. J. Antibiot. 1995, 48, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-L.; Hai, P.; Zhang, S.-B.; Xiao, J.-F.; Gao, Y.; Ma, B.-J.; Fu, H.-Y.; Chen, Y.-M.; Yang, X.-L. Prenylated indole diterpene alkaloids from a mine-soil-derived Tolypocladium sp. J. Nat. Prod. 2019, 82, 221–231. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.-M.; Li, X.; Wang, B.-G. New indole-diterpenoids from the algal-associated fungus Aspergillus nidulans. Phytochem. Lett. 2015, 12, 182–185. [Google Scholar] [CrossRef]
- Nozawa, K.; Nakajima, S.; Kawai, K.-I.; Udagawa, S.-I. Isolation and structures of indoloditerpenes, possible biosynthetic intermediates to the tremorgenic mycotoxin, paxilline, from Emericella striata. J. Chem. Soc. Perkin Trans. 1 1988, 2607–2610. [Google Scholar] [CrossRef]
- Xu, M.; Gessner, G.; Groth, I.; Lange, C.; Christner, A.; Bruhn, T.; Deng, Z.; Li, X.; Heinemann, S.H.; Grabley, S.; et al. Shearinines D–K, new indole triterpenoids from an endophytic Penicillium sp. (strain HKI0459) with blocking activity on large-conductance calcium-activated potassium channels. Tetrahedron 2007, 63, 435–444. [Google Scholar] [CrossRef]
- Zhu, L.; Han, X.; Cui, X.; Li, S.; Qiao, L.; Chen, R. Two New Alkaloids from Pleurotus ostreatus (Jacq.: Pers.) Roll. Rec. Nat. Prod. 2020, 14, 319–325. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A regulated cell death nexus linking metabolism, redox biology, and disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, C.; Zhang, C.; Zhang, W.; Zhu, W.; He, Y.; Xia, Z.; Song, W. p38 MAPK inhibitor SB202190 suppresses ferroptosis in the glutamate-induced retinal excitotoxicity glaucoma model. Neural Regen. Res. 2024, 19, 2299–2309. [Google Scholar] [CrossRef]
- Nehme, J.; Raad, P.; Jalkh, E.; Karkouh, R.; Tamer, Z.; Jalkh, A.; Quoc, E.B. Pattern ERG, pattern VEP, and GCL thickness in diabetic patients with no diabetic retinopathy. J. Français D’ophtalmol. 2025, 48, 104513. [Google Scholar] [CrossRef]
- Dubois, M.-A.; Pelletier, C.-A.; Mérette, C.; Jomphe, V.; Turgeon, R.; Bélanger, R.E.; Grondin, S.; Hébert, M. Evaluation of electroretinography (ERG) parameters as a biomarker for ADHD. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2023, 127, 110807. [Google Scholar] [CrossRef]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Stockwell, B.R. Ferroptosis turns 10: Emerging mechanisms, physiological functions, and therapeutic applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef]
- Niu, J.W.; Qi, J.Z.; Wang, P.C.; Liu, C.W.; Gao, J.M. The chemical structures and biological activities of indole diterpenoids. Nat. Prod. Bioprospect. 2023, 13, 3. [Google Scholar] [CrossRef]
- Dai, L.T.; Yang, L.; Kong, F.D.; Ma, Q.Y.; Xie, Q.Y.; Dai, H.F.; Yu, Z.F.; Zhao, Y.X. Cytotoxic Indole-Diterpenoids from the Marine-Derived Fungus Penicillium sp. KFD28. Mar. Drugs 2021, 19, 613. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Y.L.; Zhou, X.; Ni, W.; Dai, Z.; Yang, D.; Hao, J.J.; Luo, L.; Liu, Y.P.; Luo, X.D.; et al. Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair. Toxins 2017, 9, 150. [Google Scholar] [CrossRef]
- Lin, W.W.; Li, H.P.; Wu, Z.W.; Su, J.Y.; Zhang, Z.H.; Yang, L.; Deng, X.M.; Xu, Q.Y. Paspalines C-D and Paxillines B-D: New Indole Diterpenoids from Penicillium brefeldianum WZW-F-69. Mar. Drugs 2022, 20, 684. [Google Scholar] [CrossRef]
- Dai, L.-T.; Yang, L.; Guo, J.-C.; Ma, Q.-Y.; Xie, Q.-Y.; Jiang, L.; Yu, Z.-F.; Dai, H.-F.; Zhao, Y.-X. Anti-diabetic and anti-inflammatory indole diterpenes from the marine-derived fungus Penicillium sp. ZYX-Z-143. Bioorg. Chem. 2024, 145, 107205. [Google Scholar] [CrossRef]
- Pracht, P.; Bohle, F.; Grimme, S. Automated exploration of the low-energy chemical space with fast quantum chemical methods. Phys. Chem. Chem. Phys. 2020, 22, 7169–7192. [Google Scholar] [CrossRef] [PubMed]
- Mathew, B.; Ravindran, S.; Liu, X.; Torres, L.; Chennakesavalu, M.; Huang, C.C.; Feng, L.; Zelka, R.; Lopez, J.; Sharma, M.; et al. Mesenchymal stem cell-derived extracellular vesicles and retinal ischemia-reperfusion. Biomaterials 2019, 197, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Manabe, S.; Lipton, S.A. Divergent NMDA signals leading to proapoptotic and antiapoptotic pathways in the rat retina. Investig. Ophthalmol. Vis. Sci. 2003, 44, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.-W.; Liang, Y.-X.; Liu, J.-F.; Sun, Z.-Q.; So, K.-F.; Chiu, K. Retinal safety and toxicity study of artesunate in vitro and in vivo. Adv. Ophthalmol. Pract. Res. 2023, 3, 47–54. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Liang, A.-L.; Wang, C.; Chen, X.-Y.; Tan, Y.-F.; Lu, W.-Y.; Xu, P.-J.; Long, H.-P.; Liu, S.; Li, J.; Wang, W.-X.; et al. Neuroprotective Indole Diterpenoids from the Fungus Tolypocladium album DWS131. Pharmaceuticals 2026, 19, 807. https://doi.org/10.3390/ph19060807
Liang A-L, Wang C, Chen X-Y, Tan Y-F, Lu W-Y, Xu P-J, Long H-P, Liu S, Li J, Wang W-X, et al. Neuroprotective Indole Diterpenoids from the Fungus Tolypocladium album DWS131. Pharmaceuticals. 2026; 19(6):807. https://doi.org/10.3390/ph19060807
Chicago/Turabian StyleLiang, Ai-Lin, Chao Wang, Xing-Yi Chen, Yu-Feng Tan, Wen-Yu Lu, Peng-Ju Xu, Hong-Ping Long, Shao Liu, Jing Li, Wen-Xuan Wang, and et al. 2026. "Neuroprotective Indole Diterpenoids from the Fungus Tolypocladium album DWS131" Pharmaceuticals 19, no. 6: 807. https://doi.org/10.3390/ph19060807
APA StyleLiang, A.-L., Wang, C., Chen, X.-Y., Tan, Y.-F., Lu, W.-Y., Xu, P.-J., Long, H.-P., Liu, S., Li, J., Wang, W.-X., & Xia, X. (2026). Neuroprotective Indole Diterpenoids from the Fungus Tolypocladium album DWS131. Pharmaceuticals, 19(6), 807. https://doi.org/10.3390/ph19060807